
Atypical Brain Asymmetry in Human Situs Inversus: Gut Feeling or Real Evidence?


Abstract
1. Introduction
2. Left–Right Asymmetry of the Visceral System
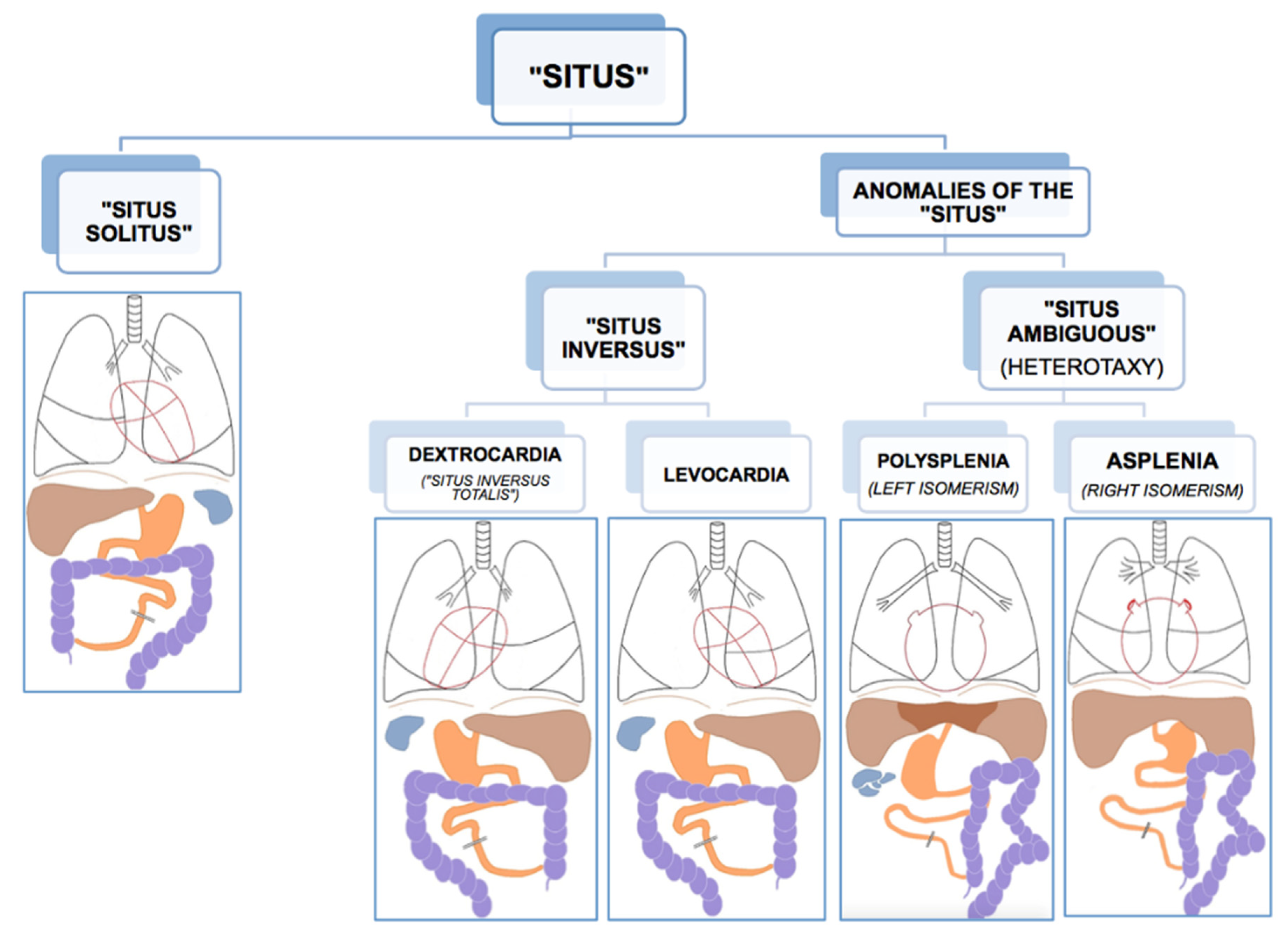
2.1. Phenotypes of Situs Viscerum
2.2. Situs Inversus
2.3. Heterotaxy
2.4. Cause of Visceral Situs Anomalies
3. Left–Right Visceral Development
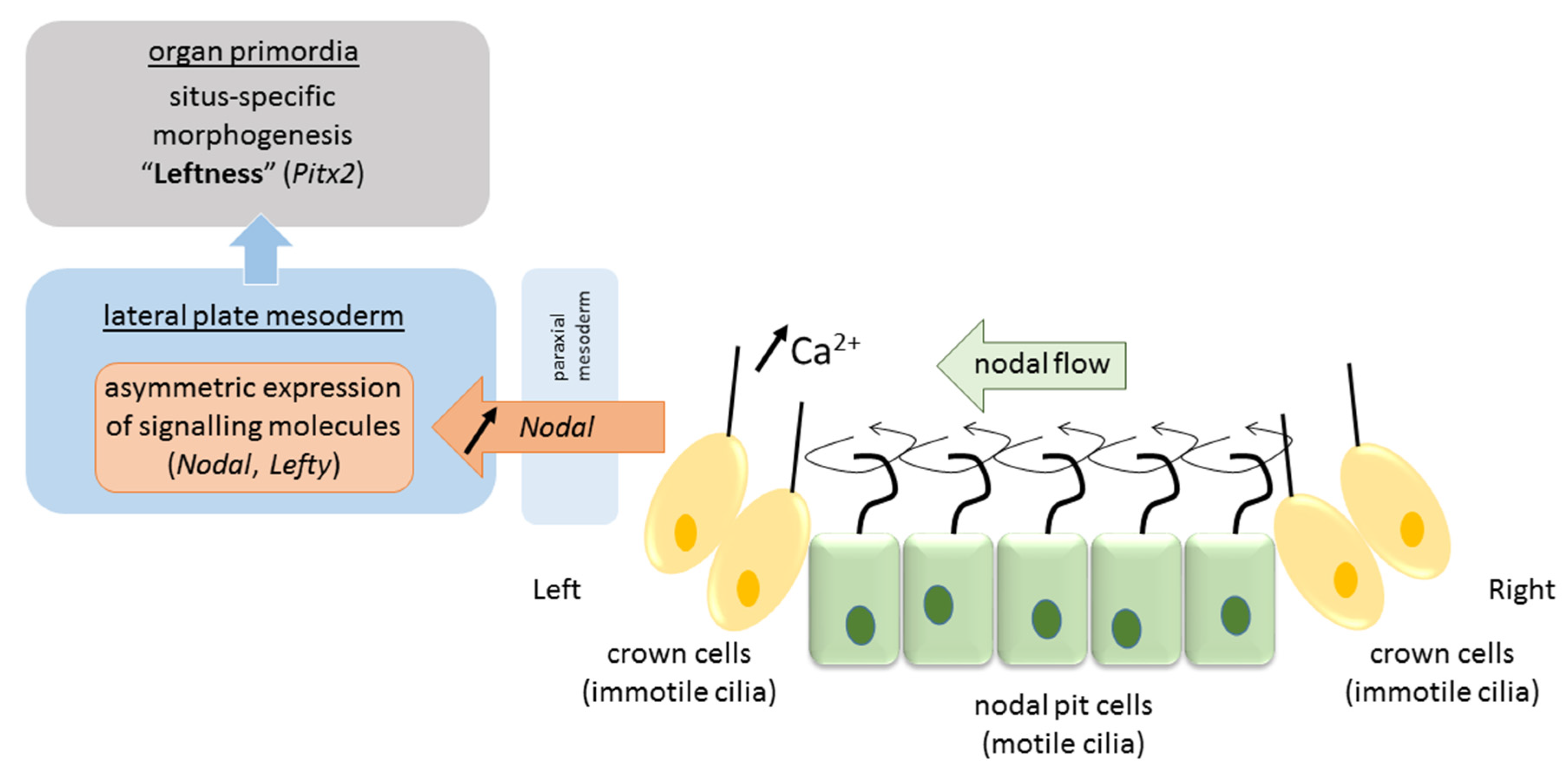
3.1. Motile Cilia at the Primitive Node
3.2. Propagation of the Signal to the Lateral Plate Mesoderm and Organ Primordia
4. Left–Right Asymmetry of the Neurocognitive System
4.1. Phenotypes of Brain Functional Organization
4.2. Reversed Typical Functional Segregation
4.3. Atypical Functional Segregation
5. Left–Right Brain Development
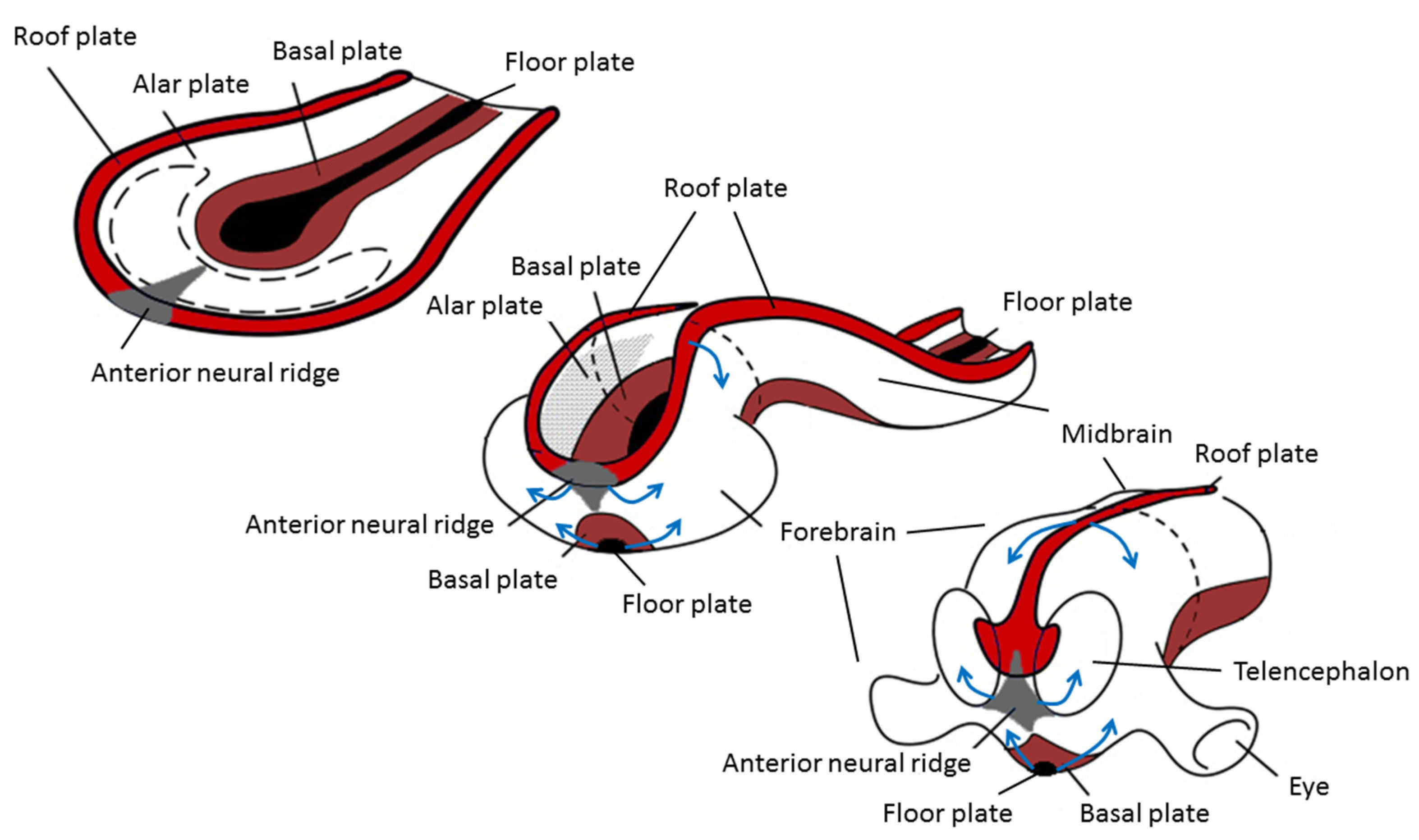
5.1. Neurulation
5.2. Asymmetric Development of the Central Nervous System
6. Are Asymmetries of Visceral and Brain Development Related?
7. Atypical Brain Asymmetry in Human Visceral Situs Anomalies
8. Systematic Review
8.1. Handedness in Situs Inversus
8.2. Brain Functional Asymmetry in Situs Inversus
8.3. Brain Structural Asymmetry in Situs Inversus
9. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Glossary
References
- Hamada, H.; Meno, C.; Watanabe, D.; Saijoh, Y. Establishment of vertebrate left-right asymmetry. Nat. Rev. Genet. 2002, 3, 103–113. [Google Scholar] [CrossRef]
- Vingerhoets, G. Phenotypes in hemispheric functional segregation? Perspectives and challenges. Phys. Life Rev. 2019. [Google Scholar] [CrossRef]
- Long, S.; Ahmad, N.; Rebagliati, M. The zebrafish nodal-related gene southpaw is required for visceral and diencephalic left-right asymmetry. Development 2003, 130, 2303–2316. [Google Scholar] [CrossRef]
- Concha, M.L.; Burdine, R.D.; Russell, C.; Schier, A.F.; Wilson, S.W. A nodal signaling pathway regulates the laterality of neuroanatomical asymmetries in the zebrafish forebrain. Neuron 2000, 28, 399–409. [Google Scholar] [CrossRef]
- Wehrmaker, A. Right-Left Asymmetry and Situs Iversus in Triturus alpestris. Wilhem Roux’ Arch. Entwickl. Org. 1969, 163, 1–32. [Google Scholar] [CrossRef] [PubMed]
- Concha, M.L.; Wilson, S.W. Asymmetry in the epithalamus of vertebrates. J. Anat. 2001, 199, 63–84. [Google Scholar] [CrossRef] [PubMed]
- Barth, K.A.; Miklosi, A.; Watkins, J.; Bianco, I.H.; Wilson, S.W.; Andrew, R.J. fsi zebrafish show concordant reversal of laterality of viscera, neuroanatomy, and a subset of behavioral responses. Curr. Biol. 2005, 15, 844–850. [Google Scholar] [CrossRef] [PubMed]
- Klingenberg, C.P. Developmental instability as a research tool: Using patterns of fluctuating asymmetry to infer the developmental origins of morphological integration. In Developmental Stability: Causes and Consequences; Polak, M., Ed.; Oxford University Press: Oxford, UK, 2003; pp. 427–442. [Google Scholar]
- Palmer, A.R. Symmetry breaking and the evolution of development. Science 2004, 306, 828–833. [Google Scholar] [CrossRef]
- Nakamura, T.; Hamada, H. Left-right patterning: Conserved and divergent mechanisms. Development 2012, 139, 3257–3262. [Google Scholar] [CrossRef]
- Aylsworth, A.S. Clinical aspects of defects in the determination of laterality. Am. J. Med. Genet. 2001, 101, 345–355. [Google Scholar] [CrossRef]
- Lin, A.E.; Krikov, S.; Riehle-Colarusso, T.; Frias, J.L.; Belmont, J.; Anderka, M.; Geva, T.; Getz, K.D.; Botto, L.D.; Prevention, N.B.D. Laterality Defects in the National Birth Defects Prevention Study (1998–2007): Birth Prevalence and Descriptive Epidemiology. Am. J. Med. Genet. Part A 2014, 164, 2581–2591. [Google Scholar] [CrossRef]
- Houyel, L.; Khoshnood, B.; Anderson, R.H.; Lelong, N.; Thieulin, A.C.; Goffinet, F.; Bonnet, D.; Grp, E.S. Population-based evaluation of a suggested anatomic and clinical classification of congenital heart defects based on the International Paediatric and Congenital Cardiac Code. Orphanet J. Rare Dis. 2011, 6. [Google Scholar] [CrossRef]
- Casey, B. Two rights make a wrong: Human left-right malformations. Hum. Mol. Genet. 1998, 7, 1565–1571. [Google Scholar] [CrossRef]
- Kosaki, K.; Casey, B. Genetics of human left-right axis malformations. Semin. Cell Dev. Biol. 1998, 9, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Lin, A.E.; Ticho, B.S.; Houde, K.; Westgate, M.N.; Holmes, L.B. Heterotaxy: Associated conditions and hospital-based prevalence in newborns. Genet. Med. 2000, 2, 157–172. [Google Scholar] [CrossRef] [PubMed]
- Evans, W.N.; Acherman, R.J.; Restrepo, H. Heterotaxy in Southern Nevada: Prenatal Detection and Epidemiology. Pediatric Cardiol. 2015, 36, 930–934. [Google Scholar] [CrossRef]
- Peeters, H.; Devriendt, K. Human laterality disorders. Eur. J. Med. Genet. 2006, 49, 349–362. [Google Scholar] [CrossRef] [PubMed]
- Sutherland, M.J.; Ware, S.M. Disorders of Left-Right Asymmetry: Heterotaxy and Situs Inversus. Am. J. Med. Genet. Part C Semin. Med. Genet. 2009, 151c, 307–317. [Google Scholar] [CrossRef]
- Schmutzer, K.J.; Linde, L.M. Situs Inversus Totalis Associated with Complex Cardiovascular Anomalies. Am. Heart J. 1958, 56, 761–768. [Google Scholar] [CrossRef]
- Chen, W.; Guo, Z.; Qian, L.; Wang, L. Comorbidities in situs inversus totalis: A hospital-based study. Birth Defects Res 2020, 112, 418–426. [Google Scholar] [CrossRef]
- Kennedy, M.P.; Omran, H.; Leigh, M.W.; Dell, S.; Morgan, L.; Molina, P.L.; Robinson, B.V.; Minnix, S.L.; Olbrich, H.; Severin, T.; et al. Congenital heart disease and other heterotaxic defects in a large cohort of patients with primary ciliary dyskinesia. Circulation 2007, 115, 2814–2821. [Google Scholar] [CrossRef]
- Gabriel, G.C.; Lo, C.W. Left-right patterning in congenital heart disease beyond heterotaxy. Am. J. Med. Genet. Part C Semin. Med. Genet. 2020, 184, 90–96. [Google Scholar] [CrossRef]
- Deng, H.; Xia, H.; Deng, S. Genetic basis of human left-right asymmetry disorders. Expert Rev. Mol. Med. 2015, 16, e19. [Google Scholar] [CrossRef]
- Postema, M.C.; Carrion-Castillo, A.; Fisher, S.E.; Vingerhoets, G.; Francks, C. The genetics of situs inversus without primary ciliary dyskinesia. Sci. Rep. 2020, 10. [Google Scholar] [CrossRef]
- Lucas, J.S.; Burgess, A.; Mitchison, H.M.; Moya, E.; Williamson, M.; Hogg, C.; Serv, N.P. Diagnosis and management of primary ciliary dyskinesia. Arch. Dis. Child. 2014, 99, 850–856. [Google Scholar] [CrossRef]
- Afzelius, B.A. Human Syndrome Caused by Immotile Cilia. Science 1976, 193, 317–319. [Google Scholar] [CrossRef]
- Bartoloni, L.; Blouin, J.L.; Pan, Y.Z.; Gehrig, C.; Maiti, A.K.; Scamuffa, N.; Rossier, C.; Jorissen, M.; Armengot, M.; Meeks, M.; et al. Mutations in the DNAH11 (axonemal heavy chain dynein type 11) gene cause one form of situs inversus totalis and most likely primary ciliary dyskinesia. Proc. Natl. Acad. Sci. USA 2002, 99, 10282–10286. [Google Scholar] [CrossRef]
- Leigh, M.W.; Pittman, J.E.; Carson, J.L.; Ferkol, T.W.; Dell, S.D.; Davis, S.D.; Knowles, M.R.; Zariwala, M.A. Clinical and genetic aspects of primary ciliary dyskinesia/Kartagener syndrome. Genet Med. 2009, 11, 473–487. [Google Scholar] [CrossRef]
- Zariwala, M.A.; Omran, H.; Ferkol, T.W. The emerging genetics of primary ciliary dyskinesia. Proc. Am. Thorac. Soc. 2011, 8, 430–433. [Google Scholar] [CrossRef]
- Bisgrove, B.W.; Yost, H.J. The roles of cilia in developmental disorders and disease. Development 2006, 133, 4131–4143. [Google Scholar] [CrossRef]
- Afzelius, B.A.; Stenram, U. Prevalence and genetics of immotile-cilia syndrome and left-handedness. Int. J. Dev. Biol. 2006, 50, 571–573. [Google Scholar] [CrossRef]
- Bush, A.; Cole, P.; Hariri, M.; Mackay, I.; Phillips, G.; O’Callaghan, C.; Wilson, R.; Warner, J.O. Primary ciliary dyskinesia: Diagnosis and standards of care. Eur. Respir. J. 1998, 12, 982–988. [Google Scholar] [CrossRef]
- Kartagener, M. Zur Pathogenese der Bronchiektasien bei Situs viscerum inversus. Beiträge Klin. Tuberk. 1933, 83, 489–501. [Google Scholar] [CrossRef]
- Engesaeth, V.G.; Warner, J.O.; Bush, A. New Associations of Primary Ciliary Dyskinesia Syndrome. Pediatric Pulmonol. 1993, 16, 9–12. [Google Scholar] [CrossRef] [PubMed]
- Noone, P.G.; Bali, D.; Carson, J.L.; Sannuti, A.; Gipson, C.L.; Ostrowski, L.E.; Bromberg, P.A.; Boucher, R.C.; Knowles, M.R. Discordant organ laterality in monozygotic twins with primary ciliary dyskinesia. Am. J. Med. Genet. 1999, 82, 155–160. [Google Scholar] [CrossRef]
- Vandenberg, L.N.; Levin, M. A unified model for left-right asymmetry? Comparison and synthesis of molecular models of embryonic laterality. Dev. Biol. 2013, 379, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, Y.; Mishina, Y. Establishment of left-right asymmetry in vertebrate development: The node in mouse embryos. Cell. Mol. Life Sci. 2013, 70, 4659–4666. [Google Scholar] [CrossRef] [PubMed]
- Hirokawa, N.; Tanaka, Y.; Okada, Y. Left-Right Determination: Involvement of Molecular Motor KIF3, Cilia, and Nodal Flow. Cold Spring Harb. Perspect. Biol. 2009, 1. [Google Scholar] [CrossRef]
- Vandenberg, L.N.; Levin, M. Far From Solved: A Perspective on What We Know About Early Mechanisms of Left-Right Asymmetry. Dev. Dyn. 2010, 239, 3131–3146. [Google Scholar] [CrossRef] [PubMed]
- Basu, B.; Bruedner, M. Cilia: Multifunctional Organelles at the Center of Vertebrate Left-Right Asymmetry. Ciliary Funct. Mamm. Dev. 2008, 85, 151–174. [Google Scholar] [CrossRef]
- Nonaka, S.; Shiratori, H.; Saijoh, Y.; Hamada, H. Determination of left-right patterning of the mouse embryo by artificial nodal flow. Nature 2002, 418, 96–99. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, D.; Saijoh, Y.; Nonaka, S.; Sasaki, G.; Ikawa, Y.; Yokoyama, T.; Hamada, H. The left-right determinant Inversin is a component of node monocilia and other 9+0 cilia. Development 2003, 130, 1725–1734. [Google Scholar] [CrossRef]
- Okada, Y.; Nonaka, S.; Tanaka, Y.; Saijoh, Y.; Hamada, H.; Hirokawa, N. Abnormal nodal flow precedes situs inversus in iv and inv mice. Mol. Cell 1999, 4, 459–468. [Google Scholar] [CrossRef]
- Smith, D.J.; Montenegro-Johnson, T.D.; Lopes, S.S. Symmetry-Breaking Cilia-Driven Flow in Embryogenesis. Annu. Rev. Fluid Mech. 2019, 51, 105–128. [Google Scholar] [CrossRef]
- Rogers, L.J.; Zucca, P.; Vallortigara, G. Advantages of having a lateralized brain. Proc. R. Soc. B Biol. Sci. 2004, 271, S420–S422. [Google Scholar] [CrossRef] [PubMed]
- Vallortigara, G.; Rogers, L.J.; Bisazza, A. Possible evolutionary origins of cognitive brain lateralization. Brain Res. Rev. 1999, 30, 164–175. [Google Scholar] [CrossRef]
- Cai, Q.; Van der Haegen, L.; Brysbaert, M. Complementary hemispheric specialization for language production and visuospatial attention. Proc. Natl. Acad. Sci. USA 2013, 110, E322–E330. [Google Scholar] [CrossRef]
- Vingerhoets, G.; Alderweireldt, A.S.; Vandemaele, P.; Cai, Q.; Van der Haegen, L.; Brysbaert, M.; Achten, E. Praxis and language are linked: Evidence from co-lateralization in individuals, with atypical language dominance. Cortex 2013, 49, 172–183. [Google Scholar] [CrossRef]
- Gerrits, R.; Van der Haegen, L.; Brysbaert, M.; Vingerhoets, G. Laterality for recognizing written words and faces in the fusiform gyrus covaries with language dominance. Cortex J. Devoted Study Nerv. Syst. Behav. 2019, 117, 196–204. [Google Scholar] [CrossRef]
- Gerrits, R.; Verhelst, H.; Vingerhoets, G. Mirrored brain organization: Statistical anomaly or reversal of hemispheric functional segregation bias? Proc. Natl. Acad. Sci. USA 2020, 117, 14057–14065. [Google Scholar] [CrossRef]
- Bryden, M.P.; Hecaen, H.; Deagostini, M. Patterns of Cerebral Organization. Brain Lang. 1983, 20, 249–262. [Google Scholar] [CrossRef]
- McNeely, H.E.; Parlow, S.E. Complementarity of linguistic and prosodic processes in the intact brain. Brain Lang. 2001, 79, 473–481. [Google Scholar] [CrossRef]
- Floel, A.; Buyx, A.; Breitenstein, C.; Lohmann, H.; Knecht, S. Hemispheric lateralization of spatial attention in right- and left-hemispheric language dominance. Behav. Brain Res. 2005, 158, 269–275. [Google Scholar] [CrossRef]
- Whitehouse, A.J.O.; Bishop, D.V.M. Hemispheric division of function is the result of independent probabilistic biases. Neuropsychologia 2009, 47, 1938–1943. [Google Scholar] [CrossRef]
- Badzakova-Trajkov, G.; Haberling, I.S.; Roberts, R.P.; Corballis, M.C. Cerebral Asymmetries: Complementary and independent processes. PLoS ONE 2010, 5, e9682. [Google Scholar] [CrossRef]
- Rosch, R.E.; Bishop, D.V.M.; Badcock, N.A. Lateralised visual attention is unrelated to language lateralisation, and not influenced by task difficulty—A functional transcranial Doppler study. Neuropsychologia 2012, 50, 810–815. [Google Scholar] [CrossRef]
- Groen, M.A.; Whitehouse, A.J.O.; Badcock, N.A.; Bishop, D.V.M. Does cerebral lateralization develop? A study using functional transcranial Doppler ultrasound assessing lateralization for language production and visuospatial memory. Brain Behav. 2012, 2, 256–269. [Google Scholar] [CrossRef]
- Zago, L.; Petit, L.; Mellet, E.; Jobard, G.; Crivello, F.; Joliot, M.; Mazoyer, B.; Tzourio-Mazoyer, N. The association between hemispheric specialization for language production and for spatial attention depends on left-hand preference strength. Neuropsychologia 2016, 93, 394–406. [Google Scholar] [CrossRef]
- Francks, C. Exploring human brain lateralization with molecular genetics and genomics. Ann. N. Y. Acad. Sci. 2015, 1359, 1–13. [Google Scholar] [CrossRef]
- Vingerhoets, G.; Gerrits, R.; Bogaert, S. Atypical brain functional segregation is more frequent in situs inversus totalis. Cortex 2018, 106, 12–25. [Google Scholar] [CrossRef]
- Mazoyer, B.; Zago, L.; Jobard, G.; Crivello, F.; Joliot, M.; Perchey, G.; Mellet, E.; Petit, L.; Tzourio-Mazoyer, N. Gaussian Mixture Modeling of Hemispheric Lateralization for Language in a Large Sample of Healthy Individuals Balanced for Handedness. PLoS ONE 2014, 9, e101165. [Google Scholar] [CrossRef]
- Zago, L.; Herve, P.Y.; Genuer, R.; Laurent, A.; Mazoyer, B.; Tzourio-Mazoyer, N.; Joliot, M. Predicting Hemispheric Dominance for Language Production in Healthy Individuals Using Support Vector Machine. Hum. Brain Mapp. 2017, 38, 5871–5889. [Google Scholar] [CrossRef]
- Bernal, B.; Ardila, A. Bilateral representation of language: A critical review and analysis of some unusual cases. J. Neurolinguist. 2014, 28, 63–80. [Google Scholar] [CrossRef]
- Janecek, J.K.; Swanson, S.J.; Sabsevitz, D.S.; Hammeke, T.A.; Raghavan, M.; Rozman, M.E.; Binder, J.R. Language lateralization by fMRI and Wada testing in 229 patients with epilepsy: Rates and predictors of discordance. Epilepsia 2013, 54, 314–322. [Google Scholar] [CrossRef] [PubMed]
- Moddel, G.; Lineweaver, T.; Schuele, S.U.; Reinholz, J.; Loddenkemper, T. Atypical language lateralization in epilepsy patients. Epilepsia 2009, 50, 1505–1516. [Google Scholar] [CrossRef]
- Sun, T.; Walsh, C.A. Molecular approaches to brain asymmetry and handedness. Nat. Rev. Neurosci. 2006, 7, 655–662. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.; Boutaud, L.; Reilly, M.L.; Benmerah, A. Cilia in hereditary cerebral anomalies. Biol. Cell 2019, 111, 217–231. [Google Scholar] [CrossRef]
- Medina, L. Evolution and Embryological Development of Forebrain. In Encyclopedia of Neuroscience; Binder, M.D., Hirokawa, N., Windhorst, U., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 1172–1192. [Google Scholar] [CrossRef]
- Sun, T.; Patoine, C.; Abu-Khalil, A.; Visvader, J.; Sum, E.; Cherry, T.J.; Orkin, S.H.; Geschwind, D.H.; Walsh, C.A. Early asymmetry of gene transcription in embryonic human left and right cerebral cortex. Science 2005, 308, 1794–1798. [Google Scholar] [CrossRef]
- Fukuchi-Shimogori, T.; Grove, E.A. Neocortex patterning by the secreted signaling molecule FGF8. Science 2001, 294, 1071–1074. [Google Scholar] [CrossRef] [PubMed]
- Puelles, L.; Martínez, S.; Martinez-de-la-Torre, M.; Rubenstein, J.L.R. Gene maps and related histogenic domains in the forebrain and midbrain. In The rat Nervous System; Paxinos, G., Ed.; Elsevier: Amsterdam, The Netherlands, 2004; pp. 3–25. [Google Scholar]
- De Kovel, C.G.F.; Lisgo, S.N.; Fisher, S.E.; Francks, C. Subtle left-right asymmetry of gene expression profiles in embryonic and foetal human brains. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef]
- De Kovel, C.G.F.; Lisgo, S.; Karlebach, G.; Ju, J.; Cheng, G.; Fisher, S.E.; Francks, C. Left-Right Asymmetry of Maturation Rates in Human Embryonic Neural Development. Biol. Psychiatry 2017, 82, 204–212. [Google Scholar] [CrossRef]
- Schmitz, J.; Lor, S.; Klose, R.; Gunturkun, O.; Ocklenburg, S. The Functional Genetics of Handedness and Language Lateralization: Insights from Gene Ontology, Pathway and Disease Association Analyses. Front. Psychol. 2017, 8, 1144. [Google Scholar] [CrossRef] [PubMed]
- Abu-Rustum, R.S.; Ziade, M.F.; Abu-Rustum, S.E. Reference Values for the Right and Left Fetal Choroid Plexus at 11 to 13 Weeks An Early Sign of “Developmental” Laterality? J. Ultrasound Med. 2013, 32, 1623–1629. [Google Scholar] [CrossRef] [PubMed]
- Vasung, L.; Rollins, C.K.; Yun, H.J.; Velasco-Annis, C.; Zhang, J.; Wagstyl, K.; Evans, A.; Warfield, S.K.; Feldman, H.A.; Grant, P.E.; et al. Quantitative In vivo MRI Assessment of Structural Asymmetries and Sexual Dimorphism of Transient Fetal Compartments in the Human Brain. Cereb. Cortex 2020, 30, 1752–1767. [Google Scholar] [CrossRef]
- Kasprian, G.; Langs, G.; Brugger, P.C.; Bittner, M.; Weber, M.; Arantes, M.; Prayer, D. The Prenatal Origin of Hemispheric Asymmetry: An In Utero Neuroimaging Study. Cereb. Cortex 2011, 21, 1076–1083. [Google Scholar] [CrossRef]
- Habas, P.A.; Scott, J.A.; Roosta, A.; Rajagopalan, V.; Kim, K.; Rousseau, F.; Barkovich, A.J.; Glenn, O.A.; Studholme, C. Early Folding Patterns and Asymmetries of the Normal Human Brain Detected from in Utero MRI. Cereb. Cortex 2012, 22, 13–25. [Google Scholar] [CrossRef]
- Yun, H.J.; Vasung, L.; Tarui, T.; Rollins, C.K.; Ortinau, C.M.; Grant, P.E.; Im, K. Temporal Patterns of Emergence and Spatial Distribution of Sulcal Pits During Fetal Life. Cereb. Cortex 2020, 30, 4257–4268. [Google Scholar] [CrossRef]
- Dubois, J.; Benders, M.; Lazeyras, F.; Borradori-Tolsa, C.; Leuchter, R.H.V.; Mangin, J.F.; Huppi, P.S. Structural asymmetries of perisylvian regions in the preterm newborn. Neuroimage 2010, 52, 32–42. [Google Scholar] [CrossRef]
- Karlebach, G.; Francks, C. Lateralization of gene expression in human language cortex. Cortex 2015, 67, 30–36. [Google Scholar] [CrossRef]
- Brandler, W.M.; Morris, A.P.; Evans, D.M.; Scerri, T.S.; Kemp, J.P.; Timpson, N.J.; St Pourcain, B.; Smith, G.D.; Ring, S.M.; Stein, J.; et al. Common Variants in Left/Right Asymmetry Genes and Pathways Are Associated with Relative Hand Skill. PLoS Genet. 2013, 9, e1003751. [Google Scholar] [CrossRef]
- Brandler, W.M.; Paracchini, S. The genetic relationship between handedness and neurodevelopmental disorders. Trends Mol. Med. 2014, 20, 83–90. [Google Scholar] [CrossRef]
- Moher, D.L.A.; Tetzlaff, J.; Altman, D.G.; The PRISMA Group. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef]
- Kienast, P.; Schwartz, E.; Diogo, M.C.; Gruber, G.M.; Brugger, P.C.; Kiss, H.; Ulm, B.; Bartha-Doering, L.; Seidl, R.; Weber, M.; et al. The Prenatal Origins of Human Brain Asymmetry: Lessons Learned from a Cohort of Fetuses with Body Lateralization Defects. Cereb. Cortex 2021. [Google Scholar] [CrossRef] [PubMed]
- McManus, I.C. Reversed bodies, reversed brains, and (some) reversed behaviors: Of zebrafish and men. Dev. Cell 2005, 8, 796–797. [Google Scholar] [CrossRef]
- Cockayne, E.A. The genetics of transposition of the viscera. Q. J. Med. New Ser. 1938, 27, 479–493. [Google Scholar]
- Torgersen, J. Situs inversus, asymmetry and twinning. Am. J. Hum. Genet. 1950, 2, 361–370. [Google Scholar]
- Papadatou-Pastou, M.; Ntolka, E.; Schmitz, J.; Martin, M.; Munafo, M.R.; Ocklenburg, S.; Paracchini, S. Human Handedness: A Meta-Analysis. Psychol. Bull. 2020, 146, 481–524. [Google Scholar] [CrossRef]
- Everett, H.C. Situs Inversus Totalis—A Survey of Laterality and Some Observations on Frequency of Mental Disorder. Am. J. Psychiatry 1963, 119, 884. [Google Scholar] [CrossRef]
- Matsumoto, T.; Kuriya, N.; Akagi, T.; Ohbu, K.; Toyoda, O.; Morita, J.; Ichikawa, K.; Matsuishi, T.; Hayashi, M.; Kato, H. Handedness and laterality of the viscera. Neurology 1997, 49, 1751. [Google Scholar] [CrossRef] [PubMed]
- Mcmanus, I.C.; Martin, N.; Stubbings, G.F.; Chung, E.M.K.; Mitchison, H.M. Handedness and situs inversus in primary ciliary dyskinesia. Proc. R. Soc. B Biol. Sci. 2004, 271, 2579–2582. [Google Scholar] [CrossRef]
- Woods, R.P. Brain Asymmetries in Situs-Inversus—A Case-Report and Review of the Literature. Arch. Neurol. 1986, 43, 1083–1084. [Google Scholar] [CrossRef]
- Tanaka, S.; Kanzaki, R.; Yoshibayashi, M.; Kamiya, T.; Sugishita, M. Dichotic listening in patients with situs inversus: Brain asymmetry and situs asymmetry. Neuropsychologia 1999, 37, 869–874. [Google Scholar] [CrossRef]
- Kennedy, D.N.; O’Craven, K.M.; Ticho, B.S.; Goldstein, A.M.; Makris, N.; Henson, J.W. Structural and functional brain asymmetries in human situs inversus totalis. Neurology 1999, 53, 1260–1265. [Google Scholar] [CrossRef]
- Schuler, A.L.; Kasprian, G.; Schwartz, E.; Seidl, R.; Diogo, M.C.; Mitter, C.; Langs, G.; Prayer, D.; Bartha-Doering, L. Mens inversus in corpore inverso? Language lateralization in a boy with situs inversus totalis. Brain Lang 2017, 174, 9–15. [Google Scholar] [CrossRef]
- Vingerhoets, G.; Li, X.; Hou, L.; Bogaert, S.; Verhelst, H.; Gerrits, R.; Siugzdaite, R.; Roberts, N. Brain structural and functional asymmetry in human situs inversus totalis. Brain Struct. Funct. 2018, 223, 1937–1952. [Google Scholar] [CrossRef]
- Cohen, L.; Geny, C.; Hermine, O.; Gray, F.; Degos, J.D. Crossed Aphasia with Visceral Situs-Inversus. Ann. Neurol. 1993, 33, 215–218. [Google Scholar] [CrossRef]
- Ihara, A.; Hirata, M.; Fujimaki, N.; Goto, T.; Umekawa, Y.; Fujita, N.; Terazono, Y.; Matani, A.; Wei, Q.; Yoshimine, T.; et al. Neuroimaging study on brain asymmetries in situs inversus totalis. J. Neurol. Sci. 2010, 288, 72–78. [Google Scholar] [CrossRef]
- Xiang, L.; Crow, T.; Roberts, N. Cerebral torque is human specific and unrelated to brain size. Brain Struct. Funct. 2019, 224, 1141–1150. [Google Scholar] [CrossRef]
- Balzeau, A.; Gilissen, E.; Grimaud-Herve, D. Shared Pattern of Endocranial Shape Asymmetries among Great Apes, Anatomically Modern Humans, and Fossil Hominins. PLoS ONE 2012, 7. [Google Scholar] [CrossRef]
- Tubbs, R.S.; Wellons, J.C.; Salter, G.; Blount, J.P.; Oakes, W.J. Intracranial anatomic asymmetry in situs inversus totalis. Anat. Embryol. 2003, 206, 199–202. [Google Scholar] [CrossRef]
- Leroy, F.; Cai, Q.; Bogart, S.L.; Dubois, J.; Coulon, O.; Monzalvo, K.; Fischer, C.; Glasel, H.; Van der Haegen, L.; Benezita, A.; et al. New human-specific brain landmark: The depth asymmetry of superior temporal sulcus. Proc. Natl. Acad. Sci. USA 2015, 112, 1208–1213. [Google Scholar] [CrossRef] [PubMed]
- Mannaert, L.; Verhelst, H.; Gerrits, R.; Bogaert, S.; Vingerhoets, G. White matter asymmetries in human situs inversus totalis. Brain Struct. Funct. 2019, 224, 2559–2565. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Author and Year | LH Function | RH Function | N (#Sinistrals) * | Typical | Reversed | Crowded | Method ** |
|---|---|---|---|---|---|---|---|
| Bryden et al., 1983 [52] | Language | Spatial dysfunction | 270 (140) | 72% (RH) | 12% (RH) | 16% (RH) | Lesions |
| 47% (LH) | 12% (LH) | 44% (LH) | |||||
| McNeely and Parlow, 2001 [53] | Language | Prosody | 73 (7) | 78% | 22% | 0% | Dichotic listening |
| Floël et al. 2005 [54] | Language | Spatial attention | 75 (38) | 95% (RH) | 2.5% (RH) | 2.5% (RH) | fTCD |
| 60% (LH) | 8% (LH) | 32% (LH) | |||||
| Whitehouse and Bischop, 2009 [55] | Language | Spatial memory | 75 (30) | 75% | 0% | 25% | fTCD |
| Badzakova-Trajkov et al. 2010 [56] | Language | Spatial attention Face processing | 155 (48) | Majority | 2% | Rest | fMRI |
| Rosch et al., 2012 [57] | Language | Spatial attention | 20 (0) | 65% | 15% | 20% | fTCD |
| Groen et al., 2012 [58] | Language | Spatial memory | 60 (13) | 58% | 5% | 32% | fTCD |
| Zago et al., 2016 [59] | Language | Spatial attention | 293 (151) | 80% | 3.5% | 15% | fMRI |
| Estimated proportion | ±70% | ±10% | ±20% |
| Author and Year | Sample Size | Situs Anomaly | Etiology | Source | Percent Left Handedness | Handedness Determination |
|---|---|---|---|---|---|---|
| Cockayne et al., 1938 [88] | 115 | Situs inversus | Not provided | From literature and the author’s own series; most included were examined on account of illness or a congenital malformation | 6.1% LH and 1.7% ambidexter | Unknown |
| Torgersen et al., 1950 [89] | Convenience sample of 160 from a total sample of 270 | Situs inversus | Not provided | Mass X-ray photographs of the Norwegian population (200) + additional cases through hospitals and municipal health departments (70) | 6.9% in SI compared to 3.5% in (715) siblings and 5.3% in (320) parents | Self-report obtained from the individual with SI |
| Everett et al., 1963, [91] | Convenience sample of 6 from a total sample of 10 | Situs inversus totalis | Not provided | Hospital records search | 0% | Self-report or hospital records |
| Matsumoto et al., 1997 [92] | 16 | Situs inversus | Not provided | Not reported | 6.3% | Writing, drawing, throwing |
| McManus et al., 2004 [93] | 46 PCD-SI and 42 PCD-SS, 334 controls | PCD-related situs inversus | PCD only | Family PCD-support group | 15.2% in PCD-SI and 14.3 in PCD-SS, 8.1 in controls | Writing hand and laterality questionnaires |
| Afzelius and Stenram et al., 2006 [32] | Convenience sample of 112 from a total sample of 239 PCD patients (105 PCD-SI and 134 PCD-SS) | Situs inversus | PCD only | Obtained from contacting many Swedish clinicians | 12.5% LH and 3.6 ambidexter in PCD, 14.3% LH in subgroup with PCD-SI | Self-report |
| Author and Year | Sample Size | Hand Preference * | Situs Anomaly | Etiology | Source | Method ** | Function | Laterality *** |
|---|---|---|---|---|---|---|---|---|
| Woods et al., 1986 [94] | 1 SI | RH | Situs inversus totalis | Sporadic case, no PCD-symptoms | Stroke patient | Lesion | Language | Aphasia following left stroke |
| Cohen et al., 1993 [99] | 1 SI | RH | Heterotaxy (polysplenia) | Sporadic case | Stroke patient | Lesion | Language | Aphasia following right stroke (crossed aphasia) |
| Tanaka et al., 1999 [95] | 9 SI and 24 controls | 9 RH | Situs inversus totalis | 8 sporadic cases, 1 PCD-SIT | Hospital search | DLT | Language | 88.9% of SIT shows REA, 79.1% of controls shows REA |
| Kennedy et al., 1999 [96] | 3 SI and 15 controls | 3 RH (Annett handedness battery) | Situs inversus totalis | 3 sporadic cases, normal general health | Hospital records search | fMRI | Language | 100% of SIT show left-lateralized activation on each of two language tasks (word stem completion and semantics) |
| Ihara et al., 2010 [100] | 3 SI and 11 controls | 1 weak RH (EHI = 38), 1 weak LH (EHI = −20), 1 strong RH (EHI = 100) | Situs inversus totalis | Unknown, 1 SIT case has left temporal epilepsy | Unknown | MEG | Language | 33% of SIT show left-lateralized activation during reading task, 66% show right-lateralized activation |
| Schuler et al., 2017 [97] | 1 SIT and 1 control | RH (EHI = 100) | Situs inversus totalis | Sporadic case, no PCD-symptoms | Fetal screening | fMRI | Language | Typical left lateralization (LI = 0.48) |
| Vingerhoets et al., 2018 [61] | 15 SIT and 15 controls | 9 RH, 6 LH; 20% LH in PCD-SIT, 55% LH in non-PCD SIT, 40% LH overall (EHI) | Situs inversus totalis | 9 sporadic cases, 6 PCD-related | Hospital records search | fMRI | Language | 80% of SIT left lateralized (93% in controls) |
| Praxis | 73% of SIT left lateralized (87% in controls) | |||||||
| Spatial attention | 73% of SIT right lateralized (93% in controls) | |||||||
| Face recognition | 80% of SIT right lateralized (87% in controls) |
| Author and Year | Sample Size | Hand Preference | Situs Anomaly | Etiology | Source | Method * | Cerebral Torque (Petalia) | Planum Temporale | Sylvian Fissure | Other ** |
|---|---|---|---|---|---|---|---|---|---|---|
| Kennedy et al., 1999 [96] | 3 SIT and 15 controls | RH (AHB = 1.24) | Situs inversus totalis | Sporadic cases, normal general health | Hospital records search | MRI | Reversed in all SIT | Volume L > R in 2 SIT and R > L in one SIT | Higher R > L in 2 SIT | |
| Tubbs et al., 2003 [103] | 1 SIT | Unknown | Situs inversus totalis | Unknown, died from “natural causes” | Unknown | Autopsy | Reversed | Higher L > R | Reversed intracranial vasculature | |
| Ihara et al., 2010 [100] | 3 SIT and 11 controls | 2 RH, 1 LH | Situs inversus totalis | Unknown, 1 SIT has temporal lobe epilepsy | Unknown | MRI | Reversed in all SIT | Volume L > R in 3 SIT | IFG volume L > R in 2 SIT, R > L in1 SIT | |
| Leroy et al., 2015 [104] | 6 SIT and 95 controls (48 RH) | 5 RH, 1 LH | Situs inversus totalis | Cases from the Kennedy et al. and Ihara et al. studies | Cases from the Kennedy et al. and Ihara et al. studies | MRI | STS deeper on right (typical) | |||
| Schuler et al., 2017 [97] | 1 SIT and 1 control | RH (EHI = 100) | Situs inversus totalis | Sporadic case, no PCD-symptoms | Fetal screening | MRI | Reversed in SIT | Absent asymmetry in SIT | STS deeper on right (typical) | |
| Vingerhoets et al., 2018 [98] and Mannaert et al., 2019 [105] | 15 SIT and 15 controls | 9 RH, 6 LH | Situs inversus totalis | 9 sporadic cases, 6 PCD-related | Hospital records search | MRI | Reversed in 78% of non-PCD SIT and in 0% of PCD SIT | Not different from controls | Same as controls | IFG volume same as controls; Heschl’s gyrus and anterior insula volume same as controls; Arcuate fasciculus volume same as controls; Uncinate fasciculus lower volume in SIT than controls |
| Kienast et al., 2021 [86] | 5 fetuses with ciliopathies and 21 fetuses with laterality defects; 26 age-matched controls | Not applicable | 4 SIT; 8 dextrocardia; 4 situs ambiguus; 5 situs inversus abdominalis | Prenatal diagnosis reports | Hospital records search | MRI | Perisylvian fetal patterns and asymmetry indices do not differ between cases with laterality defects, ciliopathies, and normal controls |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vingerhoets, G.; Gerrits, R.; Verhelst, H. Atypical Brain Asymmetry in Human Situs Inversus: Gut Feeling or Real Evidence? Symmetry 2021, 13, 695. https://doi.org/10.3390/sym13040695
Vingerhoets G, Gerrits R, Verhelst H. Atypical Brain Asymmetry in Human Situs Inversus: Gut Feeling or Real Evidence? Symmetry. 2021; 13(4):695. https://doi.org/10.3390/sym13040695
Chicago/Turabian StyleVingerhoets, Guy, Robin Gerrits, and Helena Verhelst. 2021. "Atypical Brain Asymmetry in Human Situs Inversus: Gut Feeling or Real Evidence?" Symmetry 13, no. 4: 695. https://doi.org/10.3390/sym13040695
APA StyleVingerhoets, G., Gerrits, R., & Verhelst, H. (2021). Atypical Brain Asymmetry in Human Situs Inversus: Gut Feeling or Real Evidence? Symmetry, 13(4), 695. https://doi.org/10.3390/sym13040695