Abstract
Recent studies suggest that the developmental curves in adolescence, related to the development of executive functions, could be fitted to a non-linear trajectory of development with progressions and retrogressions. Therefore, the present study proposes to analyze the pattern of development in Perceptual Inhibition (PI), considering all stages of adolescence (early, middle, and late) in intervals of one year. To this aim, we worked with a sample of 275 participants between 10 and 25 years, who performed a joint visual and search task (to measure PI). We have fitted ex-Gaussian functions to the probability distributions of the mean response time across the sample and performed a covariance analysis (ANCOVA). The results showed that the 10- to 13-year-old groups performed similarly in the task and differ from the 14- to 19-year-old participants. We found significant differences between the older group and all the rest of the groups. We discuss the important changes that can be observed in relation to the nonlinear trajectory of development that would show the PI during adolescence.
1. Introduction
Executive Functions (EF) are one of the main self-regulation tools [1,2], and, therefore, are also relevant for adaptive and goal-oriented behavior. Probably this has motivated the large number of studies published in the last 30 years [3,4,5], aimed at understanding its development and identifying its main characteristics. Executive Functions (EF) applies to a set of cognitive processes that intervene in the deliberate control of behaviors, emotions, and thoughts, favoring the achievement of goals that are relevant to each individual [6,7]. These functions play an important role in a wide range of behaviors and domains including, among others, negative emotional regulation [8], social competence [9], academic performance [10,11,12] and healthy behaviors [13]. Current literature often considers inhibition, working memory and cognitive flexibility as the main executive processes, as these are supposed to serve as the basis for the development and functioning of other executive and non-executive processes of greater complexity and level of integration [3,6,14,15,16].
With respect to the development of EF during life course, clear manifestations of executive functioning have been found already at 7 months of age, especially related with inhibition and working memory [17] and in recent neurophysiological studies [18]. From there, the literature shows that executive processes begin to improve gradually reaching the maximum parameters of development in adolescence and early adulthood, but not showing significant changes during adulthood (between 25 to 65 years old) [19,20,21]. This stage is characterized by the maintenance of the functioning parameters achieved in adolescence and youth (20 to 25 years), which are usually represented as a plateau in the developmental trajectory [8]. Finally, during old age, evidence shows the beginning of a progressive decline in the EF efficiency [22,23]. In short, the more traditional view considers that EF control is increased in a linear and progressive manner during the preschool years until the beginning of adult life [24,25,26,27,28]. However, most studies on executive development have focused on childhood, adulthood, or old age [29] but the interest and the amount of work aimed at analyzing the executive development in adolescence is substantially less compared to research done on the rest of the developmental phases. This scarce amount of research done in adolescence is probably explained by the existing belief until the 1960s and 1970s that claimed that the brain areas on which the executive function depends (i.e., the frontal lobes) did not present substantial changes during adolescence [30,31].
Nowadays adolescence is conceived as a sensitive period of development, due to the presence of significant brain, cognitive and emotional changes [32,33,34,35,36,37]. In addition, evidence has begun to emerge suggesting that at this developmental stage executive and brain changes are better suited to a non-linear trajectory of development, a position very different from that of traditional approaches based on linear and progressive development conceptions [38]. The nonlinear pattern of development in a developmental stage can be characterized by a trajectory marked by progressions and regressions that assume the form of an inverted U or by moments of improvement in between periods of stability that are represented as plateaus or valleys [39].
The study by McGivern et al. [38] comprises a relevant empirical contribution regarding the existence of nonlinear changes in executive control during adolescence. The authors compared the performance of participants aged 10 to 17 years (pre-pubescent, pubescent, middle and late teens) with the performance of young adults, aged 18 to 22 years old, in an EF task of pairing. The results showed a high demand for working memory and decision making. The results exhibited a clear decrease in performance at the beginning of puberty (at approximately 11 years), which was reflected in the increase from 10% to 20% in Response Times (RT). Additionally, in the group of pubescent women, the drop in performance occurred at 11–12 years, while in the group of pubescent men it was registered at 10–11 years of age. Both gender groups showed a linear downward trend that recovers the level of performance shown before puberty. However, this effect was only verified in the RT and not in the Accuracy (ACC) measured as percentage of correct responses, which, according to the authors, suggests a temporary decrease in the efficiency (RT) of the processes involved in puberty. Other studies provide similar performance patterns like the ones described by [38] (e.g., [39,40,41,42]).
On the other hand, ref. [41] analyzed the performance in a prospective memory task—with demand for EF control—during childhood, adolescence, and adulthood. The authors report a clear improvement in the performance of the task from 6 to 10 years. However, there were no changes between the ages of 10 and 14. Finally, the group of young adults (25 years’ average age) performed better than the group of children. This study reported a maintenance in puberty of the level of performance achieved in childhood, not showing a progressive and linear development.
In the same line of argumentation, ref. [35] argued that traditional neurobiological and cognitive explanations fail to explain risky choices and actions observed in adolescence, which represent a non-linear change in behavior, and they are not typically found in childhood or adulthood. Considering the traditional perspective, if cognitive control and the immaturity of the prefrontal cortex were the basis of inappropriate or maladaptive behaviors, then children should manifest behaviors remarkably similar or even worse than adolescents, since its prefrontal cortex and cognitive abilities are less developed. Therefore, immature prefrontal function alone cannot explain the behavior of adolescents. According to [34], in adolescents there is an imbalance between the limbic regions—functionally mature—and the prefrontal cortex—still in development—compared to children, for which both systems (limbic and prefrontal) are still being developed, and compared to adults, for whom these systems are already mature.
In summary, previous studies suggest that in adolescence, executive control processes may present a trajectory of non-linear development. However, much of the empirical evidence shows that executive improvement advances progressively and linearly to adulthood (e.g., [42,43]). This lack of consensus is likely to be explained by the type of tasks used to assess executive processes, as not all of them impose equivalent demands on the same executive processes (i.e., working memory, inhibition, cognitive flexibility). In addition, it is known that different executive processes can present different developmental trajectories [44,45], so it is possible that while some processes adjust to a linear path, others do not.
As mentioned above, the frontal lobe also undergoes significant changes during adolescence (e.g., [46]). Research conducted through non-invasive brain imaging techniques such as functional magnetic resonance imaging (fMRI) has allowed to report a continuous and sustained linear increase of the white matter and a progressive and non-linear decrease of the gray matter [47,48]. The adolescent’s brain is different from the child’s brain. As they develop, neurons create a layer of myelin around their axon that acts as an insulator by increasing the rate of transmission of electrical impulses. This layer of myelin is formed by fatty tissue (lipids), which gives it the whitish appearance and explains its name (white matter). At puberty there is more white matter than in childhood, and not only in the frontal lobe [46,47,48], but also in the rest of the brain regions [49]. The evidence has shown a linear and progressive increase in white matter density, associated with age (e.g., [50]). The interesting thing is that this increase not only occurs in childhood and adolescence, but also continues until the age of 60 [51].
The other brain change characteristic of this stage is the decrease of the gray matter, constituted by neuronal bodies and dendrites. However, unlike the white matter changes, which implies a linear increase in most brain regions, in the case of grey matter, a non-linear decrease occurs in different brain areas and at different developmental times (e.g., [33]). With respect to the changes that occur in the frontal lobe, an important body of evidence, provided mostly by longitudinal studies, shown that the gray substance reaches a peak of synaptic density that coincides with the onset of puberty. For some authors, this clear and rapid increase reflects a brain reorganization, described as a wave of synaptic proliferation [52,53,54,55] which is reflected in the increase in the number of synaptic connections. However, after puberty and while adolescence, a rapid but constant decrease in synaptic density is observed thanks to the intervention of the synaptic pruning mechanism [47,56,57]. Synaptic pruning allows the elimination of excessive “noisy” synapses that have not been strengthened through experience [55,56]. In this way, an adjustment and refinement of those synapses more efficient for the organism is achieved.
Depending on the brain and executive changes described above, we wonder if some executive processes such as inhibition [57] could present a non-linear pattern of development in adolescence. In this regard, the study by McGivern et al. [38] through a task with high demand for working memory, shows a pattern of such development type. The authors attributed the decrease in puberty performance to the noise generated by the excessive number of synapses. Consequently, the subsequent improvement was explained by the activation of the synaptic pruning mechanism (e.g., ([58]). In this sense, some executive processes may be particularly affected by these brain changes, while in others the effect may be less.
In addition, as mentioned above, studies on executive development in adolescence are scarce compared to other stages such as childhood or old age. On the other hand, when adolescence is included as part of the life course, it is usually considered as a homogeneous group, so rarely is it distinguished between adolescents of different ages or, at most, adolescents are divided into two large groups (older and younger) (see ([59]). Although the conclusions based on this type of study (e.g., [60]) provide a relevant piece of information regarding the differences found between broad developmental stages—childhood, adulthood, old age—they do not provide specific information about the changes that occur within each stage.
In general, adolescence is divided into early adolescence or puberty (11–13 years), middle adolescence (14–16 years) and late adolescence (17–20 years). Therefore, in this study, we intend to assess the following questions: How is the development of inhibition in adolescents? Are there critical moments of improvement associated with a particular stage or phase of adolescence or is there rather a gradual and progressive improvement without abrupt changes? Is there a time of regression or plateau at the onset of puberty or early adolescence? Does the phenomenon of retrogression occur in both women and men? If so, does it happen at the same ages or do differences occur? We consider that the answers to these questions may represent an interesting contribution to an existing lack of research in the executive processes development area in adolescence.
2. Method
2.1. Design, Participants, and Procedure
A non-experimental, transversal design was followed ([61]). We worked with an intentional non-probabilistic sample of 275 participants aged from 10 to 25.
We think that it is important to establish small age ranges because it would reduce the variability and reflect developmental changes in a more accurate way. In this regard, and with the aim of comparing performance between different age groups, most previous studies usually use intervals of two to three years in childhood and adolescence and at least 10 years in adulthood [19,62,63,64]. The potential problem that we observed while working with these intervals, especially during adolescence or adulthood, is that they are so wide that they do not throw precise information to analyze certain inferences about developmental changes. For this reason, the decision was taken to reduce the intervals in adolescence to one year and, in adulthood, to five years. Thus, in the case of adolescents, 10 groups were formed, from 10 to 19 years of age—10 years of age (n = 25; 76% women, 24% men), group of 11 years (n = 25, 64% women, 36% men), group of 12 years (n = 25, 52% women, 48% men), group of 13 years (n=25, 36% women, 64% men), group of 14 years (n = 25, 58% women, 42% men), group of 15 years (n = 25, 64% women, 36% men), 16-year-old group (n = 25, 52% female, 48% male), 17-year-old group (n = 25, 60% female, 40% male), 18-year-old group (n = 25, 91% female, 9% male) and 19-year-old group (n = 25, 68% female, 32% male); and in the case of young adults, only one group between 20 and 25 years of age was formed (n = 25, 74% female, 26% male; age M = 23.04, DE = 2.05). The sample size was first calculated through the G*Power 3.1 software, incorporating the following data: data analysis chosen = F test, ANCOVA—fixed, main and interaction effects; p < 0.05; power = 0.95; n2= 0.2; number of groups = 11; number of covariates = 1. The results indicated that it is necessary to have at least a total of 176 participants (16 per group). Considering the possible loss of subjects and the method of selection of the sample, it was considered appropriate to have 25 subjects per group.
Participants aged 10 to 17 were students from two educational institutions in the city of Mar del Plata (Argentina), at the primary and secondary levels. Participants aged 18 and above were students from an educational institution that groups together several higher education degrees, from the same city. Unfortunately, the 18-year-old age group is biased towards women, due to the fact that the degrees imparted at this educational institution have predominantly female students. The inclusion criteria of the participants in the sample were at the moment of the testing phase: (i) they were not in psychological and/or psychiatric treatment; (ii) they had a typical development (i.e., without deficits or alterations, without a history of learning or developmental disorders); and (iii) with normal or corrected vision and hearing (conditions necessary to carry out the proposed activities).
The activities were carried out in the educational institutions attended by the participants, in specially designed classrooms where environmental sound and changeable light were controlled. The test administration took place in an individual meeting that had a duration of 10 to 12 min. The visual search task was applied on a HP LAPTOP-RJSENA2U computer with Windows 10 and a 15.6” screen.
2.2. Ethical Considerations
Our research team was authorized by the Ethics Committee of the National University of Mar del Plata (Argentina) for the study of executive functions and application of the tasks at different stages of development. The participation of the subjects was voluntary. First, the informed consent explaining the objectives, the test to be administered in this investigation, the treatment and confidential use of the data was submitted, in accordance with the Declaration of Helsinki [65] and in line with the ethical principles and code of conduct of psychologists, established and reformulated by the [66]. The procedures outlined in the [67] on the protection of personal data regulated by [68] were followed. Specifically, the participants from 16 to 25 years old signed the corresponding informed consent. Adolescents aged 13 to 15 also signed the consent and, at the same time, their parents or legal guardians gave the informed consent for their participation. Finally, children aged 10 to 12 years gave their consent, while their parents or legal guardians signed the consent, authorizing their participation in the study.
2.3. Instruments
Visual search task: Search for conjunctions. To evaluate Inhibitory Control (IC) at the perceptual level, we used one of the tasks that integrate the computerized TAC battery—Cognitive self-regulation tasks [69,70,71]. This is a visual search task that has been designed from the Conjunction Visual Search [72]. In the task, participants should identify the presence or absence of a target stimulus (blue square) that is mixed between a variable set of distracting stimuli (red squares and blue circles). Stimuli consist of double conjunctions, which are defined by the combination of two visual characteristics: shape and color. In addition, all distractors share one of these visual characteristics with the “target”, a condition that guarantees the interference effect and the participation of selective attention and perceptual inhibition. Thus, while selective attention is the mechanism that enables orienting the attention towards relevant objects of the environment, perceptual inhibition counteracts the interference yielded by the distractors, diminishing the activation of the perceptual features which are irrelevant to or incompatible with the objective [3]. Therefore, the visual search task allows to guarantee the measurement of the perceptual inhibition.
The task is composed of a block of 10 practice trials, followed by three experimental blocks of 40 trials each. Each experimental block contains 10 trials per distractor quantity condition: 4, 8, 16 and 32 distractors. The 40 trials are randomly distributed in each block; in 50% of the trials in each block the target is present while in the rest it is absent. The participant must always address an answer, either affirmative or negative, as quickly and precisely as possible, by pressing the corresponding key (the “Z” key if the target is present and the “M” key if absent). Once the answer is given, the next trial appears (see Figure 1).
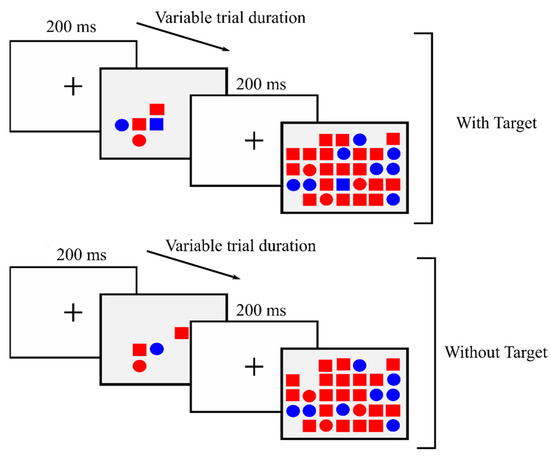
Figure 1.
Example: four trials of 4 and 32 distractors with target and without target. Target: blue square.
In addition, the task contains a block of tests without interference, which is presented first, to bring a basic measurement of the processing speed. In this block there is only one stimulus (a blue square (the target), a blue circle, or a red square). It contains 10 practice trials and 20 experimental trials.
The performance in the task is analyzed through two main measures, the Response Time (RT) and the percentage of hits on the answers given (Accuracy as percentage of hits, ACC). In this way, two indices are obtained for each of the distractor quantity conditions (without distractors, 4, 8, 16 and 32). The TR record is only obtained based on the correct answers. Typical performance corresponds to an increase in mean TRs at the same time as a decrease in response ACCs, as the number of distractors increases. Thus, as the number of distractors increases, there is a significant decrease in performance [72]. Several authors consider that the use of RT or ACC in isolation does not fully reflect performance, so they recommend using measures that combine speed and accuracy [73]. This combined measure was proposed by [74] and consists of dividing the RT by the ratio of correct responses (ACC). Later, it was called Inverse Efficiency (IE) by [75], because the higher the IE score, the less efficient the performance. In this study, IE indexes are used for each of the Interference Conditions (IC) and for the condition without distractors (CWD).
3. Data Analysis
The ex-Gaussian distribution is commonly used to represent the reaction time data [76]. This positive-skewed distribution function comes from the convolution of a Gaussian function and an exponential decay function. The convolution is a mathematical operation on two functions that produces a third function which expresses how the shape of one function is modified by the other. The parameters of the ex-Gaussian function are known to correlate with cognitive processes such as attention or effort control mechanism.
In this work the reaction time (RT) data have been fitted to the following expression of the ex-Gaussian function,
where the symbol “” denotes the convolution operation and,
Equation (1) depends on three parameters, namely, and [the mean and the standard deviation of the Gaussian function, ], and [the decay constant of the exponential component, ],
The first three moments of the ex-Gaussian distribution can be expressed in terms of , and . The mean is expressed as , the variance as , and the skewness as .
We apply a Covariance Analysis (ANCOVA) with an inter-subject factor, an intra-subject factor, and a covariate. The inter-subject factor was age with 11 categories; and the intra-subject factor were the different interference conditions (indexes of IE in 4, 8, 16 and 32 distractors). The covariate used was the measurement obtained in the condition without interference (index of IE in the condition without distractors), in order to control the base performance and the processing speed of the subjects [77]. Prior to this, we evaluated the fulfillment of the assumptions required for the application of the proposed analysis—homogeneity of variance, linearity in terms of the relations between covariate and dependent variables, and homogeneity of regression [78].
4. Results
In Figure 2 the histograms of the variables that represent the RTs through the different conditions in relation to the number of distractors, TR4, TR8, TR16 and TR32, referring to 4, 8, 16 and 32 distractors, respectively ([76] for similar analysis on RTs) are shown. By dividing histograms by the area under the curve, we obtained a probability distribution for each of these RT variables (BVTR4, BVTR8, BVTR16 and BVTR32). Then, we have fitted ex-Gaussian functions to each of the data of the probability distribution functions. The resulting values are shown in Table 1 and represented in Figure 3. The results show that at the level of these parameters, the RTs variables analyzed differ well. The fitting parameters and their uncertainties were obtained by the nonlinear fitting algorithm of Levenberg-Marquardt [79,80], implemented in ORIGIN version 6.1 data analysis software. This is a numeric minimization algorithm that finds the best values of parameters (, and in our case) for the best approach of equation (1) to the RT data. The algorithms follow a series of iterative steps starting from an initial guess for the parameter values until converge is reached.
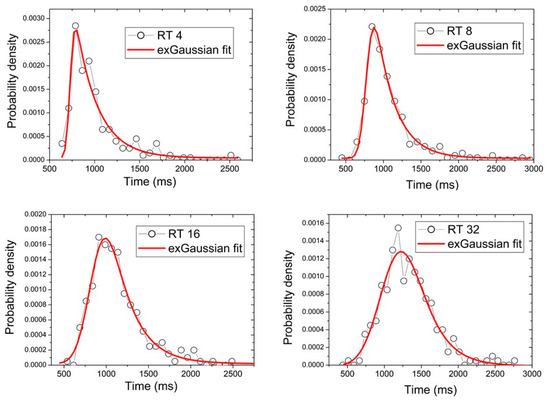
Figure 2.
Histograms RTs variables through distractors amount conditions.

Table 1.
Ex-Gaussian functions (µ, σ, τ) adjusted to each probability distribution RTs data.
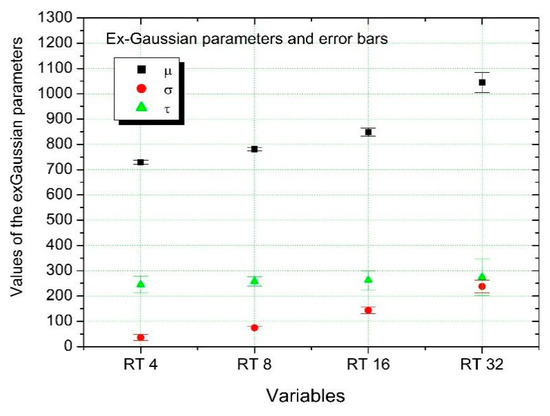
Figure 3.
Ex-Gaussian functions fitted to the probability distribution on RTs values.
In Table 2, we show the descriptive statistics of the main indexes of performance according to the number of distractors shown in age groups.
Table 2.
Descriptive statistics of the main indexes of performance according to the number of distractors shown in age groups.
A statistically significant effect was found in the interference conditions—F (2540) = 8834, p < 0.001, η2p = 0.033—age group—F (10,255) = 11.749, p < 0.001, η2p = 0.315—and an interaction effect between interference conditions and age group—F (21,540) = 14.392, p < 0.001, η2p = 0.361.
Figure 4 shows the performance of participants from different age groups under different interference conditions. The graph shows a trend in which participants show a worse performance the greater the number of distractors (see Figure 3).
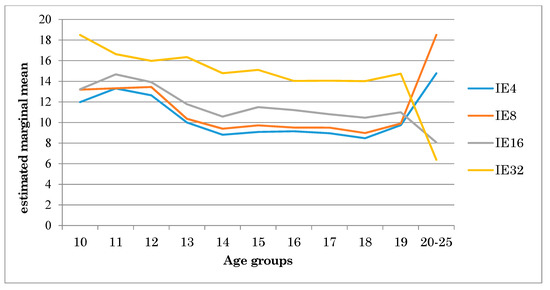
Figure 4.
Performance of different age groups under different interference conditions. Note: IE = Inverse Efficiency Index.
Peer comparisons indicate, in general terms, that the 10-, 11-, 12- and 13-year-old groups are similar in their performance under the different conditions of the visual search task. In turn, these groups differ significantly from participants aged 14 to 19, while the latter groups are similar to each other. Finally, significant differences were observed between the older age group (20–25 years) and all the previous ones (see Table 3).
Table 3.
Peer comparisons between age groups under different interference conditions (IE).
5. Discussion
The general objective of this study was to analyze the development of one of the main executive processes (inhibition) in the adolescent and young adult populations. Current evidence has identified that executive processes do not present the same developmental trajectories during development [3,81]. Within these processes, inhibition plays a fundamental role in the learning and acquisition of complex cognitive skills such as mathematics [82,83], reading comprehension [84], problem-solving ability and intelligence [7,85]. For this reason, we focus specifically on the study of the developmental trajectory of this process, with the purpose of analyzing whether during adolescence and young adulthood, inhibition presents a linear progression or, on the contrary, whether plateaus or even regressions exist, as some authors have suggested [38,41,42,59].
In relation to inhibitory processes, perceptual inhibition showed different characteristics compared to other inhibitory processes such as behavioral inhibition or cognitive inhibition [71,86,87]. Its main function is to reduce or attenuate the interference generated by environmental stimuli. Therefore, to evaluate the functioning of this inhibitory process we used a classic visual search task in which the participant must identify a visual stimulus (target or target) that occurs between a variable number of distracting stimuli.
The results suggest that during adolescence there are important changes in the functioning of this inhibitory process. In general, the data show that between the ages of 10 and 25 there is a substantial improvement in the functioning of perceptual inhibition, and that this is manifested in different interference conditions, that is, under conditions which impose various demands on the inhibitory control.
However, between the ages of 10 and 25, the increase in inhibitory efficiency does not seem to follow a linear path of development, as some studies suggest [42,88,89,90,91]. The performance obtained in the visual search task showed in the present study that, in this age range, the developmental trajectory presents moments of change that correspond to an improvement in inhibitory control, which are represented as peaks, followed by periods of stability visualized as plateaus or valleys (see Figure 2). Thus, between the ages of 10 and 13, there is a plateau that represents the absence of significant differences in performance rates between the different age groups. It is likely that, at this early stage of adolescence, and as life-course theorists suggest, much of the biopsychosocial resources will be used to maintain the achievements made in earlier stages (infancy) [92,93].
Additionally, according to the results obtained, at the age of 14 there seems to be a turning point, that is, a noticeable change in inhibitory efficiency, which is represented as a jump or peak (see Figure 1). This difference seems to constitute a consistent and robust change, since it involves all the conditions of the task, from those that generate a lower interference effect by presenting a smaller number of distractors, even those requiring greater inhibitory control by exposing the participant to higher levels of interference (conditions of 16 and 32 distracting stimuli). From that age, a plateau is again detected that reflects the absence of significant changes between the different groups in most conditions up to 20–25 years, in which a new peak appears that indicates a notable improvement in performance.
It is worth noting that all the differences between adolescents aged 14 to 19 and young adults are detected in the condition of 32 distracting stimuli, that is, in the condition that imposes a greater demand for inhibitory control. However, surprisingly, there are no differences between middle/late teens and young adults (20–25 years old) in the condition of 16 distracting stimuli. On the other hand, contrary to what is observed in the condition of 32 distractors, in the conditions of less interference (4 and 8 distractors), the data show a decrease in performance. That is, from the age of 20, young adults identify a target more efficiently when it occurs between several distractors, but when the target is mixed among a few distractors, the performance is better in adolescents. This difference is probably explained by the implementation of different search strategies. In this regard, some authors [94] argue that, in search conditions with a few distractors, simultaneous or parallel strategies are usually used, while, in conditions with greater demand, in other words, with more distractors, the implementation of a serial search strategy is expected. This type of strategy demands greater inhibitory control [95], and that is why the differences are particularly evident in this condition.
In summary, during adolescence—and as found by [38,41] with respect to other executive functions—perceptual inhibition also appears to have a non-linear trajectory or progression. If we compare the performance obtained in puberty or early adolescence with that obtained in the last phase of this developmental period, the data showed a clear improvement in inhibitory control. However, the improvement is characterized by moments of stability (plateaus or valleys) interspersed by peaks or moments of clear improvement and change. Between the ages of 10 and 14, there is a plateau, followed by a peak at the age of 14, another period of stability and again a change that is detected in early adulthood (20 to 25 years). Clearly, and as proposed by other studies on the development of executive processes [89,90,91], progress continues in early adulthood. The adolescents in our study performed worse than young adults. Which are the cognitive mechanisms that allow us to explain this developmental path? If we consider a classical cognitive theory such as Processing Speed Theory, both periods in inhibition (i.e., plateau periods and periods of rapid increase) may be explained by periods of stability and increased efficiency in processing speed. That is, according to this approach, we proposed that the variations in processing speed would be the core explanation for the concomitant observed variations in inhibitory functioning. Finally, the plateau (absence of linear improvement) obtained from 10 to 14 years in relation to processing speed can be conceived as the result of synaptic proliferation. However, we cannot specify at what point in adulthood the progress occurs, given that at this developmental stage we did not work with intervals of one year as in the case of adolescents, but a single sample was formed with young adults between 20 and 25 years. This prevents the precise identification within this group of the moment when the change is generated. Therefore, this would be an aspect of interest to consider in future studies, since it would allow recording not only the time when the change in the efficiency of the inhibitory control occurs, but also the possible new plateaus or periods of stability that may arise, or new periods of change, whether positive (in the form of new increases or improvements) or negative (as reversals or declines from previous earnings). Likewise, other limitations may be considered. First, the majority presence of women in the sample, especially in the 18-year age group. That bias is due to the fact that the educational institution from which the late adolescents come from has a mostly female student body. Therefore, we also consider it desirable for future studies to use more equitable samples in terms of gender. Likewise, future studies may also include the development of other inhibitory processes, response and cognitive inhibition and thus provide evidence for current models of inhibitory control. Finally, the type of design and the nature of the sample (non-probabilistic) limit the generality of our results, and thus they should be interpreted with caution.
Finally, and in relation to the findings reported in [38], this study detected a slight setback in performance in the task of visual search, at the beginning of adolescence or puberty. However, it should be noted that this trend does not reach the level of significance that would allow inferring the existence of a real regression in inhibitory efficiency and, in turn, was not registered in the condition of greater interference (32 distractors). Taken together, the data show worse performance of 11-year-olds compared to 10-year-olds. For it, we think that in future research it would be interesting to replicate the present study in a larger sample that focuses exclusively on this stage (puberty or early adolescence). The potential relevance of analyzing this aspect in greater detail during this developmental stage lies in the relationships that can be found between brain changes and behavioral modifications. As has been argued, if a regression is found on tasks that measure EF, it is likely that this phenomenon can be explained by the excess of synaptogenesis that occurs prior the activation of the synaptic pruning mechanism that is observed after puberty [47,53,54].
6. Conclusions
The present study proposed to analyze the developmental pattern of Executive Functions by measuring Perceptual Inhibition (PI), considering all stages of adolescence (early, middle, and late) in intervals of one year. The present study proposal tends to fulfill the theoretical and empirical gap that we described, revealing the paucity of studies that have thus far been conducted on EF in adolescents and their lack of clear confluent conclusions. In this sense, it is worth remarking that, to our knowledge, there are not studies that using the same experimental task, measuring the same Executive Function, and analyzing the adolescence in more stages, all of that on the same study and sample. We explored changing curves in adolescence that could be fitted to a non-linear trajectory of development with progressions and retrogressions. Our results clearly reflect an improvement in inhibitory functioning—in conditions of visual interference—during middle adolescence, in line with what was reported by other studies, which describe a minor advance or plateau at the beginning of adolescence [89]. In addition, young adults in this study (20 to 25 years) show better performance than adolescent participants, especially in the condition of greater interference. In this line of argumentation, we mainly discuss the important changes that can be observed in relation to the nonlinear trajectory of development; this would show the PI during adolescence, challenging (a) the idea that executive functioning culminates its development during adolescence and (b) that the youth stage is a period without significant observed changes [28,40,57,96].
Author Contributions
Conceptualization, I.M.I., M.M.R., L.-R.M.F., and E.N.-P.; Writing–original draft preparation, I.M.I., M.M.R., A.G.C., and L.-R.M.F.; Writing—review and editing, E.N.-P.; Software, A.G.C., Y.A., and F.S.; Data curation, F.S.; Methodology, M.M.R., E.V.Z., L.-R.M.F., and E.N.-P.; Visualization, M.M.R., E.V.Z., and Y.A.; Project administration, I.M.I. and M.M.R.; Funding acquisition, I.M.I., L.-R.M.F., and E.N.-P. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki, and approved by the Ethics Committee of National University of Mar del Plata, Argentina (protocol code N°061/2016, Sheet 124, Book 2, and date of approval 14 November 2017).
Informed Consent Statement
Informed consent was obtained from all subjects who participated in the study or from their parents/guardians in the case of minors.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hofmann, W.; Schmeichel, B.J.; Baddeley, A.D. Executive functions and self-regulation. Trends Cogn. Sci. 2012, 16, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Rothbart, M.K.; Sheese, B.E.; Rueda, M.R.; Posner, M.I. Developing mechanisms of self-regulation in early life. Emot. Rev. 2011, 3, 207–213. [Google Scholar] [CrossRef]
- Diamond, A. Executive functions. Annu. Rev. Psychol. 2013, 64, 135–168. [Google Scholar] [CrossRef]
- Schmeichel, B.J.; Tang, D. Individual Differences in Executive Functioning and Their Relationship to Emotional Processes and Responses. Curr. Dir. Psychol. Sci. 2015, 24, 93–98. [Google Scholar] [CrossRef]
- Zelazo, P.D.; Blair, C.B.; Willoughby, M.T. Executive Function: Implications for Education; (NCER 2017–2000); National Center for Education Research, Institute of Education Sciences, U.S. Department of Education: Washington, DC, USA, 2016. Available online: http://ies.ed.gov (accessed on 9 October 2020).
- Diamond, A. Why improving and assessing executive functions early in life is critical. In Executive Functions in Pre-School Age-Children. Integrating Measurement, Neurodevelopment and Translational Research; Griffin, J., McCardle, P., Freund, L., Eds.; American Psychological Association: Washington, DC, USA, 2016; pp. 11–44. [Google Scholar] [CrossRef]
- Friedman, N.P.; Miyake, A. Unity and diversity of executive functions: Individual differences as a window on cognitive structure. Cortex 2017, 86, 186–204. [Google Scholar] [CrossRef] [PubMed]
- Eisenberg, N.; Smith, C.; Spinrad, T. Effortful Control Relations with Emotion Regulation, Adjustment, and Socialization in Childhood. In Handbook of Self-Regulation. Research, Theory, and Applications, 3rd ed.; Vohs, K.D., Baumeister, R.F., Eds.; Guilford Press: New York, NY, USA, 2016; pp. 458–478. [Google Scholar]
- Sharifian, N.; Sol, K.; Zahodne, L.B.; Antonucci, T.C. Social relationships and adaptation in later life. Ref. Mod. Neurosci. Biobehav. Psych. 2020. [Google Scholar] [CrossRef]
- Brock, L.L.; Rimm-Kaufman, S.E.; Nathanson, L.; Grimm, K.J. The contributions of ‘hot’ and ‘cool’ executive function to children’s academic achievement, learning-related behaviors, and engagement in kindergarten. Early Child. Res. Q. 2009, 24, 337–349. [Google Scholar] [CrossRef]
- Jacobson, L.A.; Williford, A.P.; Pianta, R.C. The role of executive function in children’s competent adjustment to middle school. Child Neuropsychol. 2011, 17, 255–280. [Google Scholar] [CrossRef]
- Orbach, L.; Herzog, M.; Fritz, A. Relation of attention deficit hyperactivity disorder (ADHD) to basic number skills and arithmetic fact retrieval in children. Res. Dev. Disabil. 2020, 103, 103697. [Google Scholar] [CrossRef]
- Allan, J.L.; McMinn, D.; Daly, M.A. Bidirectional Relationship between Executive Function and Health Behavior: Evidence, Implications, and Future Directions. Front. Neurosci. 2016, 10, 386. [Google Scholar] [CrossRef]
- Jabłoński, S. Inhibitory control and literacy development among 3- to 5-year-old children. L1-Educ. Stud. Lang. Lit. 2013, 13, 1–25. [Google Scholar]
- Miyake, A.; Friedman, N.P.; Emerson, M.J.; Witzki, A.H.; Howerter, A.; Wager, T.D. The unity and diversity of executive functions and their contributions to complex “frontal lobe” tasks: A latent variable analysis. Cogn. Psychol. 2000, 41, 49–100. [Google Scholar] [CrossRef] [PubMed]
- Nigg, J.T. Annual Research Review: On the relations among self-regulation, self-control, executive functioning, effortful control, cognitive control, impulsivity, risk-taking, and inhibition for developmental psychopathology. J. Child Psychol. Psychiatry 2017, 58, 361–383. [Google Scholar] [CrossRef] [PubMed]
- Diamond, A.; Goldman-Rakic, P. Comparison of human infants and rhesus monkeys on Piaget’s A-not-B task: Evidence for dependence on dorsolateral prefrontal cortex. Exp. Brain Res. 1989, 74, 24–40. [Google Scholar] [CrossRef]
- Klatt, L.; Getzmann, S.; Wascher, E.; Schneider, D. Searching for auditory targets in external space and in working memory: Electrophysiological mechanisms underlying perceptual and retroactive spatial attention. Behav. Brain Res. 2018, 353, 98–107. [Google Scholar] [CrossRef]
- Hommel, B.; Li, K.Z.H.; Li, S.C. Visual search across the life span. Dev. Psychol. 2004, 40, 545–558. [Google Scholar] [CrossRef] [PubMed]
- Lobaugh, N.J.; Cole, S.; Rovet, J.F. Visual search for features and conjunctions in development. Can. J. Exp. 1998, 52, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Trick, L.M.; Enns, J.T. Lifespan changes in attention: The visual search task. Cogn. Dev. 1998, 13, 369–386. [Google Scholar] [CrossRef]
- Jacques, S.; Marcovitch, S. Development of executive function across the life span. In The Handbook of Life-span Development; Overton, W.F., Lerner, R.M., Eds.; Wiley: Hoboken, NJ, USA, 2010; Volume 1, pp. 431–466. [Google Scholar]
- Zelazo, P.D.; Craik, F.I.M.; Booth, L. Executive function across the life span. Acta Psychol. 2004, 115, 167–184. [Google Scholar] [CrossRef]
- Carlson, S. Developmentally Sensitive Measures of Executive Function in Preschool Children. Dev. Neuropsychol. 2005, 28, 595–616. [Google Scholar] [CrossRef]
- León-Carrión, J.; García Orza, J.; Pérez-Santamaría, F.J. Development of the inhibitory component of the executive functions in children and adolescents. Int. J. Neurosci. 2004, 114, 1291–1311. [Google Scholar] [CrossRef] [PubMed]
- Mezzacappa, E. Alerting, orienting, and executive attention: Developmental properties and sociodemographic correlates in an epidemiological sample of young, urban children. Child Dev. 2004, 75, 1373–1386. [Google Scholar] [CrossRef] [PubMed]
- Rueda, M.R.; Fan, J.; McCandliss, B.D.; Halparin, J.D.; Gruber, D.B.; Lercari, L.P.; Posner, M.I. Development of attentional networks in childhood. Neuropsychologia 2004, 42, 1029–1040. [Google Scholar] [CrossRef]
- Rueda, M.R.; Conejero, A. Developing attention and self-regulation in infancy and childhood. In Neural Circuit and Cognitive Development, 2nd ed.; Rubenstein, J., Rakic, P., Chen, B., Kwan, K.Y., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 505–522. [Google Scholar] [CrossRef]
- Poon, K. Hot and Cool Executive Functions in Adolescence: Development and Contributions to Important Developmental Outcomes. Front. Psychol. 2018, 8, 2311. [Google Scholar] [CrossRef]
- Blakemore, S.-J.; Frith, U. The learning brain: Lessons for education: A precis. Dev. Sci. 2005, 8, 459–465. [Google Scholar] [CrossRef] [PubMed]
- Ruotsalainen, I.; Gorbach, T.; Perkola, J.; Renvall, V.; Syväoja, H.J.; Tammelin, T.H.; Parviainen, T. Physical activity, aerobic fitness, and brain white matter: Their role for executive functions in adolescence. Dev. Cogn. Neurosci. 2020, 42, 100765. [Google Scholar] [CrossRef] [PubMed]
- Andrews-Hanna, J.; Mackiewicz Seghete, K.; Claus, E.; Burgess, G.; Ruzic, L.; Banich, M. Cognitive Control in Adolescence: Neural Underpinnings and Relation to Self-Report Behaviors. PLoS ONE 2011, 6, e21598. [Google Scholar] [CrossRef] [PubMed]
- Blakemore, S.J.; Choudhury, S. Development of the adolescent brain: Implications for executive function and social cognition. J. Child Psychol. Psychiatry 2006, 47, 296–312. [Google Scholar] [CrossRef] [PubMed]
- Casey, B. Beyond Simple Models of Self-Control to Circuit-Based Accounts of Adolescent Behavior. Annu. Rev. Psychol. 2015, 66, 295–319. [Google Scholar] [CrossRef] [PubMed]
- Casey, B.; Getza, S.; Galván, A. The adolescent brain. Dev. Rev. 2008, 28, 62–77. [Google Scholar] [CrossRef]
- Giedd, J.N. Structural magnetic resonance imaging of the adolescent brain. Ann. N. Y. Acad. Sci. 2004, 1021, 77–85. [Google Scholar] [CrossRef]
- Simmonds, D.J.; Hallquist, M.N.; Luna, B. Protracted development of executive and mnemonic brain systems underlying working memory in adolescence: A longitudinal fMRI study. Neuroimage 2017, 157, 695–704. [Google Scholar] [CrossRef]
- McGivern, R.F.; Andersen, J.; Byrd, D.A.; Mutter, K.A.; Reilly, J.S. Cognitive efficiency on a match to sample task decreases at the onset of puberty in childrem. Brain Cog. 2002, 50, 73–89. [Google Scholar] [CrossRef]
- Pauls, F.; Macha, T.; Petermann, F. U-shaped development: An old but unsolved problem. Front. Psychol. 2013, 4, 301. [Google Scholar] [CrossRef]
- Andreson, P. Assessment and development of executive function (EF) during childhood. Child Neuropsychol. 2002, 8, 71–82. [Google Scholar] [CrossRef] [PubMed]
- Mackinlay, R.; Charman, T.; Karmiloff-Smith, A. High functioning children with autism spectrum disorder: A novel test of multitasking. Brain Cog. 2006, 61, 14–24. [Google Scholar] [CrossRef]
- Anderson, V.; Anderson, P.; Northam, E.; Jacobs, R.; Catroppa, C. Development of executive functions through late childhood and adolescence in an Australian sample. Dev. Neuropsychol. 2001, 20, 385–406. [Google Scholar] [CrossRef] [PubMed]
- Burnett Heyes, S.; Lau, J.Y.F.; Holmes, E.A. Mental imagery, emotion and psychopathology across child and adolescent development. Dev. Cogn. Neurosci. 2013, 5, 119–133. [Google Scholar] [CrossRef] [PubMed]
- Best, J.R.; Miller, P.H. A developmental perspective on executive function. Child Dev. 2010, 81, 1641–1660. [Google Scholar] [CrossRef] [PubMed]
- Friedman, N.P.; Miyake, A.; Robinson, J.L.; Hewitt, J.K. Developmental trajectories in toddlers’ self-restraint predict individual differences in executive functions 14 years later: A behavioral genetic analysis. Dev. Psychol. 2011, 47, 1410–1430. [Google Scholar] [CrossRef]
- Bodison, S.C.; Colby, J.B.; Sowell, E.R. Structural Brain Development: Birth through Adolescence, 1st ed.; Rubenstein, J., Rakic, P., Chen, B., Kwan, K.Y., Eds.; Academic Press: Cambridge, MA, USA, 2020. [Google Scholar] [CrossRef]
- Giedd, J.N.; Blumenthal, J.; Jeffries, N.O.; Castellanos, F.X.; Liu, H.; Zijdenbos, A.; Paus, T.; Evans, A.C.; Rapoport, J.L. Brain development during childhood and adolescence: A longitudinal MRI study. Nat. Neurosci. 1999, 2, 861–863. [Google Scholar] [CrossRef]
- Giedd, J.N.; Snell, J.W.; Lange, N.; Rajapakse, J.C.; Kaysen, D.; Vaituzis, A.C.; Vauss, Y.C.; Hamburger, S.D.; Kozuch, P.L.; Rapoport, J.L. Quantitative magnetic resonance imaging of human brain development: Ages 4–18. Cereb. Cortex 1996, 6, 551–559. [Google Scholar] [CrossRef] [PubMed]
- Paus, T.; Zijdenbos, A.; Worsley, K.; Collins, D.L.; Blumenthal, J.; Giedd, J.N.; Rapoport, J.L.; Evans, A.C. Structural maturation of neural pathways in children and adolescents: In vivo study. Science 1999, 283, 1908–1911. [Google Scholar] [CrossRef] [PubMed]
- Stramba-Badiale, C.; Mancuso, V.; Cavedoni, S.; Pedroli, E.; Cipresso, P.; Riva, G. Transcranial Magnetic Stimulation Meets Virtual Reality: The Potential of Integrating Brain Stimulation With a Simulative Technology for Food Addiction. Front. Neurosci. 2020, 14, 720. [Google Scholar] [CrossRef] [PubMed]
- Sowell, E.R.; Peterson, B.S.; Thompson, P.M.; Welcome, S.E.; Henkenius, A.L.; Toga, A.W. Mapping cortical change across the life span. Nat. Neurosci. 2003, 6, 309–315. [Google Scholar] [CrossRef]
- Bourgeois, J.P.; Goldman-Rakic, P.S.; Rakic, P. Synaptogenesis in the prefrontal cortex of rhesus monkeys. Cereb. Cortex 1994, 4, 78–96. [Google Scholar] [CrossRef]
- Sowell, E.R.; Thompson, P.M.; Tessner, K.D.; Toga, A.W. Mapping continued brain growth and gray matter density reduction in dorsal frontal cortex: Inverse relationships during postadolescent brain maturation. J. Neurosci. 2001, 21, 8819–8829. [Google Scholar] [CrossRef]
- Gogtay, N.; Giedd, J.N.; Lusk, L.; Hayashi, K.M.; Greenstein, D.; Vaituzis, A.C.; Nugent, T.M.; Herman, D.H.; Clasen, L.S.; Toga, A.W.; et al. Dynamic mapping of human cortical development during childhood through early adulthood. Proc. Natl. Acad. Sci. USA 2004, 101, 8174–8179. [Google Scholar] [CrossRef]
- Huttenlocher, P.R. Synaptic density in human frontal cortex-developmental changes and effects of aging. Brain Res. 1979, 163, 195–205. [Google Scholar] [CrossRef]
- Huttenlocher, P.R.; Dabholkar, A.S. Regional differences in synaptogenesis in human cerebral cortex. J. Comp. Neurol. 1997, 387, 167–178. [Google Scholar] [CrossRef]
- Ólafsdóttir, I.M.; Gestsdóttir, S.; Kristjánsson, Á. Age differences in foraging and executive functions: A cross-sectional study. J. Exp. Child Psychol. 2020, 198, 104910. [Google Scholar] [CrossRef] [PubMed]
- Laviola, G.; Marco, E.M. Passing the knife edge in adolescence: Brain pruning and specification of individual lines of development. Neurosci. Biobehav. R. 2011, 35, 1631–1633. [Google Scholar] [CrossRef] [PubMed]
- Cepeda, N.J.; Kramer, A.F.; Gonzalez de Sather, J. Changes in executive control across the life span: Examination of task-switching performance. Dev. Psychol. 2001, 37, 715–730. [Google Scholar] [CrossRef]
- De Luca, C.; Wood, S.; Anderson, V.; Buchanan, J.; Proffitt, T.; Mahony, K.; Pantelis, C. Normative Data From the Cantab. I: Development of Executive Function Over the Lifespan. J. Clin. Exp. Neuropsychol. 2003, 25, 242–254. [Google Scholar] [CrossRef]
- Hernández Sampieri, R.; Fernández Collado, C.; Baptista Lucio, P. Metodología de la Investigación, 6th ed.; McGraw-Hill: México DF, México, 2014. [Google Scholar]
- Darowski, E.S.; Helder, E.; Zacks, R.T.; Hasher, L.; Hambrick, D.Z. Age-related differences in cognition: The role of distraction control. Neuropsychology 2008, 22, 638–644. [Google Scholar] [CrossRef]
- Huizinga, M.; Dolan, C.V.; Van der Molen, M.W. Age-related change in executive function: Developmental trends and a latent variable analysis. Neuropsychologia 2006, 44, 2017–2036. [Google Scholar] [CrossRef]
- McAuley, T.; White, D.A. A latent variables examination of processing speed, response inhibition, and working memory during typical development. J. Exp. Child Psychol. 2011, 108, 453–468. [Google Scholar] [CrossRef] [PubMed]
- World Medical Association. World Medical Association Declaration of Helsinki: Ethical Principles for Medical Research Involving Human Subjects. JAMA 2013, 310, 2191–2194. [Google Scholar] [CrossRef] [PubMed]
- American Psychological Association (APA). Ethical Principles of Psychologists and Code of Conduct; APA: Whashington DC, USA, 2017; Available online: www.apa.org/ethics/code/ethics-code-2017.pdf (accessed on 9 October 2020).
- Congress of the Argentine Nation. Argentine National Law on the Protection of Personal Data; No. 25.326; Buenos Aires, Argentina, 2000. Available online: https://www.argentina.gob.ar/normativa/nacional/ley-25326-64790 (accessed on 9 October 2020).
- National Executive Government. Personal Data Protection Act; No. 1558/2001; Buenos Aires (Argentina), 2001. Available online: http://servicios.infoleg.gob.ar/infolegInternet/anexos/70000-74999/70368/norma.htm (accessed on 9 October 2020).
- Canet-Juric, L.; Stelzer, F.; Andrés, M.L.; Vernucci, S.; Introzzi, I.; Burin, D. Evidencias de validez de una tarea computarizada de memoria de trabajo verbal y viso-espacial para niños. Interam. J. Psychol. 2018, 52, 112–128. [Google Scholar]
- Introzzi, I.M.; Canet-Juric, L.; Aydmune, Y.; Stelzer, F. Perspectivas teóricas y evidencia empírica sobre la inhibición. Rev. Colomb. Psicol. 2016, 25, 351–368. [Google Scholar] [CrossRef]
- Richard’s, M.; Introzzi, I.; Zamora, E.; Vernucci, S. Analysis of Internal and External Validity Criteria for a Computerized Visual Search Task. A pilot study. Appl. Neuropsychol. Child 2017, 6, 110–119. [Google Scholar] [CrossRef]
- Treisman, A.M.; Gelade, G. A feature-integration theory of attention. Cogn. Psychol. 1980, 12, 97–136. [Google Scholar] [CrossRef]
- Klein, R.M.; Christie, J.J.; Ivanoff, J. Graphical and other methods for representing the speed and accuracy of performance. In Proceedings of the 45th Annual Meeting of the Psychonomic Society, Minneapolis, MN, USA, 18–21 November 2004; Abstract Number 7. Volume 9, p. 1. Available online: https://cdn.ymaws.com/www.psychonomic.org/resource/resmgr/Annual_Meeting/Past_and_Future_Meetings/Abstracts04.pdf (accessed on 9 October 2020).
- Townsend, J.T.; Ashby, F.G. Stochastic Modeling of Elementary Psychological Processes, 1st ed.; Cambridge University Press: Cambridge, UK, 1983. [Google Scholar]
- Christie, J.; Klein, R. Familiarity and attention: Does what we know affect what we notice? Mem. Cogn. 1995, 23, 547–550. [Google Scholar] [CrossRef] [PubMed]
- Castro-Palacio, J.C.; Fernández-de-Córdoba, P.; Isidro, J.M.; Navarro-Pardo, E.; Selvas Aguilar, R. Percentile Study of χ Distribution. Application to Response Time Data. Mathematics 2020, 8, 514. [Google Scholar] [CrossRef]
- Introzzi, I.; Aydmune, Y.; Zamora, E.V.; Vernucci, S.; Ledesma, R. Mecanismos de desarrollo de la atención selectiva en población infantil (The development mechanisms of selective attention in child population). Rev. CES Psicol. 2019, 12, 105–118. [Google Scholar] [CrossRef]
- Tabachnick, B.G.; Fidell, L.S. Using Multivariate Statistics, 6th ed.; Pearson: Boston, MA, USA, 2013. [Google Scholar]
- Levenberg, K. A method for the solution of certain non-linear problems in least squares. Q. Appl. Math. 1944, 2, 164–168. Available online: http://www.jstor.org/stable/43633451 (accessed on 9 October 2020). [CrossRef]
- Marquardt, D. An algorithm for least-squares estimation of nonlinear parameters. J. SIAM 1963, 11, 431–441. [Google Scholar] [CrossRef]
- Bardikoff, N.; Sabbagh, M. The Differentiation of Executive Functioning Across Development: Insights from Developmental Cognitive Neuroscience. In New Perspectives on Human Development, 1st ed.; Budwig, N., Turiel, E., Zelazo, P., Eds.; Cambridge University Press: Cambridge, UK, 2017; pp. 27–46. [Google Scholar] [CrossRef]
- Bull, R.; Lee, K. Executive functioning and mathematics achievement. Child Dev. Perspect. 2014, 8, 36–41. [Google Scholar] [CrossRef]
- Cragg, L.; Keeble, S.; Richardson, S.; Roome, H.E.; Gilmore, C. Direct and indirect influences of executive functions on mathematics achievement. Cognition 2017, 162, 12–26. [Google Scholar] [CrossRef]
- Borella, E.; De Ribaupierre, A. The role of working memory, inhibition, and processing speed in text comprehension in children. Learn. Indiv. Differ. 2014, 34, 86–92. [Google Scholar] [CrossRef]
- Arán Filippetti, V.; Krumm, G.; Raimondi, W. Funciones Ejecutivas y sus correlatos con Inteligencia Cristalizada y Fluida: Un estudio en Niños y Adolescentes. Rev. Neuropsi. Lat. 2015, 7, 24–33. [Google Scholar] [CrossRef]
- Hasher, L.; Lustig, C.; Zacks, R. Inhibitory mechanisms and the control of attention. In Variation in Working Memory; Conway, A., Jarrold, C., Kane, M., Miyake, A., Towse, J., Eds.; Oxford University Press: New York, NY, USA, 2007; pp. 227–249. [Google Scholar] [CrossRef]
- Friedman, N.P.; Miyake, A. The relations among inhibition and interference control functions: A latent-variable analysis. J. Exp. Psychol. 2004, 133, 101–135. [Google Scholar] [CrossRef] [PubMed]
- Bausela Herreras, E. Desarrollo evolutivo de la función ejecutiva. Rev. Galego Port. Psicoloxía Educ. 2005, 10, 85–93. [Google Scholar]
- Flores-Lázaro, J.; Castillo-Preciado, R.; Jiménez-Miramonte, N. Desarrollo de funciones ejecutivas, de la niñez a la juventud. An. Psicol. 2014, 30, 463–473. [Google Scholar] [CrossRef]
- Luciana, M.; Conklin, H.; Hooper, C.; Yarger, R. The Development of Nonverbal Working Memory and Executive Control Processes in Adolescents. Child Dev. 2005, 76, 697–712. [Google Scholar] [CrossRef] [PubMed]
- Romine, C.B.; Reynolds, C.R. A model of the development of frontal lobe functioning: Findings from a meta-analysis. Appl. Neuropsychol. 2005, 12, 190–201. [Google Scholar] [CrossRef]
- Baltes, P.B. On the incomplete architecture of human ontogeny. Am. Psychol. 1997, 52, 366–380. [Google Scholar] [CrossRef]
- Baltes, P.B.; Lindenberger, U.; Staudinger, U.M. Life-span theory in Developmental Psychology. In Handbook of Child Psychology, 6th ed.; Damon, W., Lerner, R.M., Eds.; Wiley: New York, NY, USA, 2006; Volume 1, pp. 569–664. [Google Scholar] [CrossRef]
- Introzzi, I.; Zamora, E.; Aydmune, Y.; Canet-Juric, L.; López, S. El rol de la inhibición en la Teoría de Integración de las Características. Cuad. Neuropsicol. 2017, 11, 135–150. [Google Scholar] [CrossRef]
- Mullane, J.C.; Corkum, P.V.; Klein, R.M.; McLaughlin, E. Interference control in children with and without ADHD: A systematic review of Flanker and Simon task performance. Child Neuropsychol. 2009, 15, 321–342. [Google Scholar] [CrossRef] [PubMed]
- Best, J.R.; Miller, P.H.; Jones, L.L. Executive Functions after Age 5: Changes and Correlates. Dev. Rev. 2009, 29, 180–200. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).