Abstract
Visual attention can be adjusted to task requirements. We asked participants to switch between judging the symmetry of vertically presented three-letter strings and identifying the central stimulus (i.e., Eriksen task) to investigate anticipatory adjustment of attention. Our experiments provide evidence for anticipatory adjustment of visual attention, depending on the cued task (i.e., focusing and defocusing of attention after the Eriksen task cue and after the symmetry task cue, respectively). Although, symmetry judgments were, overall, considerably slower than the identification of the central letter, the effects of response congruency between tasks were comparable in the two tasks, which suggested strong response priming from concurrent symmetry judgment in Eriksen task trials. Symmetry judgment performance was best for homogeneous letter strings (e.g., HHH), worst for strings that were symmetrical and inhomogeneous (e.g., XHX), and intermediate for asymmetrical strings (e.g., HHX). The difficulty of categorizing symmetrical-inhomogeneous items markedly deviated from the aesthetic ratings of the stimuli, displaying a pronounced preference for symmetrical strings, but only little difference among the symmetrical items, and might be accounted by conflict with response priming based on inhomogeneity detection. Although our study provides little evidence for an effect of aesthetic appreciation in simple symmetry judgments, it demonstrates the strong role of contextual dependencies.
1. Introduction
Aesthetic appreciation constitutes a form of perceptual categorization applicable to a huge variety of stimuli and events, ranging from simple shapes [1,2] to judgments of acts of revenge in work contexts [3]. Its empirical investigation started as early as the 1870s by Fechner [4,5], who founded the discipline Empirical Aesthetics. Fechner already pointed to a symbiosis regarding symmetry and the aesthetic perception and its appearance in creative work such as paintings [4]. The subsequent research shows that symmetry might be a fundamental characteristic of aesthetic appreciation one cannot miss and the in-between positive association: Symmetrical faces are perceived as being more beautiful than asymmetrical faces [6,7,8], amplifying those findings when adding symmetrical decorations [9]. The same holds for human bodies [10]. Artificial, graphical patterns are rated as being more beautiful if they are symmetrical when compared to asymmetrical patterns [11,12,13,14,15].
Besides different types of visual patterns showing a preference for symmetry, there is also a liking for symmetrical acoustic stimuli in comparison to asymmetrical acoustic stimulation [16]. This cross modality, as well as the relation of symmetry and aesthetic value over a variation of stimulus types, allow the inference that symmetry is a relevant and considerable factor when empirically examining aesthetics.
Albeit, this unquestionable link between symmetry and aesthetics, it is not as universal and lawful as one might deduce based on the number of studies focusing on this relationship. Firstly, individual preferences shall not be obliviated when examining the aesthetic value of any kind empirically. Jacobsen [17] was able to show that there are individual preferences in simple patterns, which cannot be integrated into a model adequately. Secondly, art expertise reverses the coherence between symmetry and the aesthetic value: art experts perceive asymmetric patterns as more beautiful opposed to non-experts, who show the classical relation between beauty and symmetry [18]. Hence, symmetry might not be as universal, as assumed previously; still, it allocates a significance, which should not be underestimated due to individuality regarding the subjective perception of beauty. This is supported by the finding that, independent of task requirements, there is a sustained posterior negativity (SPN) that is observable after presenting symmetrical stimuli [13,19,20], indicating that symmetry is processed in any case [21].
Studies show that symmetry in faces and bodies are perceived as more attractive [6,7,8,10]. A reasonable explanation is that symmetry might imply a qualitative good partner for offspring [22]. A further theory comprises the superficial structure of symmetry [23]: the perceptual structure of symmetrical patterns allows for a more fluent processing causing a higher aesthetic appreciation and faster cognitive processing inducing better performance [24].
When having a closer look at the symmetrically induced advantage concerning the performance one can see that task requirements [13], variation in display time [25], orientation of the symmetry axis [25,26,27,28], various types of stimuli (e.g., dot patterns [29], graphical patterns [13], or line patterns [27]), and types of symmetry (reflection/mirror, translation, and rotation [30,31,32,33,34,35]) influence the performance. Contradictions within the different sources of impact cloud an apparent conclusion regarding the underlying processing mechanisms (for an overview on models of the processing of symmetry [36]).
Using single element stimuli, Carmody, Nodine, and Locher [37] observed symmetry to be detectable within a very short display time: a differentiation between symmetrical and asymmetrical patterns is possible within display times as short as 25 milliseconds. Thus, it seems to be a valid conclusion that symmetry is processed preattentively [38].
However, Olivers and van der Helm [39] justifiably pointed out that there might have been confounding factors, such as a given fixation point in advance leading to an already attentional focus onto the region of the (a)symmetrical stimulus’ location prior to its appearance and/or the attention might already be narrowed onto a single element. Thus, Olivers and van der Helm [39] resorted onto the use of visual search tasks, in which one had to determine whether a symmetrical target element was present (or absent) among a varying number of asymmetrical non-target elements and vice versa counteracting the named confounding aspects. Two different results were possible: if symmetry is processed fast and holistic in any case, the performance should not decrease with the increase of non-targets. However, if the perception is not processed preattentively and universally, the number of non-targets should affect the performance on the visual search task. The results clearly support the latter: over several types of stimuli ranging from simple lines over complex line figures to classical dot patterns, there was a linear relationship between the number of non-targets and the reaction time as well as the error rate. Restricting the display time of the stimuli diminished the linear increase. At the same time, it led to a more error-prone performance: the error rate exceeded 50 % when searching for a present, asymmetrical target among three additional symmetrical non-targets, thus, being approximately at chance level. Olivers and van der Helm [39] concluded that symmetry is not processed fast in all cases, and can be subject to a slower serial processing when enlarging the stimulus display to several elements with no prior knowledge of the target’s location. However, they also pointed out that there is an overall advantage of the combination of asymmetrical targets and symmetrical non-target when compared to the reversed combination, which might appear confusing at first. Assuming that single, symmetrical elements are processed faster than asymmetrical elements, non-targets can be more quickly rejected and causing the benefit of the combination of asymmetrical targets and symmetrical non-targets. Thus, concerning the perception of symmetry, there seems to be aspects of a fast processing as well as a serial and slower. One with the allocation of attention being highly influential on symmetry perception.
Cueing the orientation of the axis influences the performance [29,40]. Additionally, varying the axis’ orientation unpredictably is of disadvantage when compared to a single, clearly predictable orientation [41], indicating some sort of attentional processes based on the task requirements. However, a clear understanding of the relation between the anticipation of the stimulus, the respective adjustment of attention, and symmetry perception is not given and it is of further interest, especially if the task requirements do not entail an explicit focus on the symmetrical aspect of the stimulus.
Further, aesthetic appreciation might also have consequences for attentional or cognitive processing strategies. For instance, Goller, Mitrovic, and Leder [42] demonstrated a positive relation of aesthetic judgment regarding pictures of faces or paintings and fixation times for these stimuli [43]. Given the ubiquitous applicability of aesthetic appreciation to all sorts of stimuli, possibly taking place in a highly automatic manner, aesthetic factors might, therefore, affect processing in a variety of tasks that were unrelated to aesthetical issues. Hence, analyzing aesthetic judgments concerning stimuli typically administered in widely used psychological investigations seem to be a valuable endeavor with regard to revealing possible confounds with task-related stimulus categories. In the current study, this was exemplified for an investigation of attentional processing of simple letter string stimuli in two tasks that were associated with opposing demands of perceptual stimulus selection.
Experimental protocols that were frequently used in research on visual attention often involve small sets of stimuli consisting of simple perceptual structures, such as letter strings containing only few constituent elements. Most noteworthy, in the Eriksen flanker task [44,45], participants are presented a string of elementary stimulus objects, such as letters, and they are asked to identify or categorize the element presented at a particular location, disregarding the other elements. The flanker compatibility effect denotes the finding of worse performance in conditions, in which non-target elements (i.e., flankers) are associated with a different response than the target element compared to cases in which all elements of the string are associated with the same response. A frequently used version of the Eriksen task involves no more than two different elements (e.g., the letters H and S. [46]) and letter strings, which consist of the target element presented in the central position, flanked by the same number of either the same letter or the other letter on each side (e.g., HHH, SSHSS). By this arrangement, all of the used strings are symmetrical regarding the sequence of their constituent elements. Homogeneous strings are referred to as compatible and heterogeneous strings as incompatible.
It is widely assumed that stimulus processing in the Eriksen flanker task involves the focusing of visual attention on the location of the central letter. Indeed, Wendt, Kähler, Luna-Rodriguez, and Jacobsen [47] provided evidence that such attentional focusing occurs during task preparation. Experiment 2 of that study employed the stimuli that are displayed in the top left quarter of Table 1 (i.e., HHH, SHS, HSH, SSS). The participants switched between an Eriksen flanker task, assumed to be associated with the focusing of visual attention to the central letter, and a homogeneous/heterogeneous judgment task, assumed to be associated with visual attention that is more evenly distributed across the three letter positions. On a proportion of trials, a visual search task was presented instead of the task indicated by the cue, however, in which a to-be-detected target stimulus equiprobably occurred at any of the three locations of the letters. We refer to the visual search task as the probe task and to its target stimulus as the probe stimulus. Search times in the intermixed trials of the probe task displayed different patterns of dependency of the location of the probe stimulus, consistent with the notion that participants adopt different sets of visual attention during task-specific preparation for the Eriksen task and the homogeneous/heterogeneous judgment task. Specifically, a more pronounced center-to-periphery gradient, that is, faster responses to probes presented in the central as compared to in the peripheral locations, was observed after a cue indicating the likely occurrence of the Eriksen task compared to after a cue indicating the likely occurrence of the homogeneous/heterogeneous judgment task (this was confirmed by a significant contrast of quadratic trends for probe stimuli presented across the possible three positions in trials associated with the cue of the Eriksen task vs. the cue of the homogeneity judgment task).

Table 1.
Categorization scheme of the letter strings used in the experiments of the current study.
One of the major goals of the current study was to corroborate our previous findings of anticipatory adjustment of visual attention to task requirements indicated by an advance task cue by replacing the homogeneous/heterogeneous judgment task by another task requiring the processing of a global stimulus attribute. To this end, we included asymmetrical stimuli (i.e., three-letter strings involving two instances of the same letter at the central and at one of the peripheral positions, as well as another letter that was located at the other peripheral position) and asked the participants to judge the symmetry/asymmetry of these strings. In Experiment 1 of the current study, we administered stimuli involving the letters H and S as constituent elements. In Experiment 2A, we only used (mirror)symmetrical elements (i.e., the letters H and X). In light of the fact that the type of symmetry judgment required in our experiments was equivalent to a judgment of whether the two peripherally letters are identical or not, we replicated Experiment 2A, thus replacing the symmetry judgment instruction by instruction to compare the peripheral letters to investigate whether deployment of attention to the elements of the stimuli was affected by the participants’ instruction-based task-set (Experiment 2B). For all experiments, we expected a higher degree of focusing attention on the location of the central stimulus element after a cue indicating the Eriksen task which should evidence itself in a more pronounced center-to-periphery gradient of search times in the intermixed trials of the probe task.
Although our investigation of attentional deployment in the Eriksen task and the symmetry judgment task heavily relied on the analysis of response performance in the probe task (i.e., comparing search time gradients), performance in the Eriksen task and the symmetry judgment task themselves were also of interest, which allowed for us to assess possible interference effects between the two task-sets. Looking at response priming resulting from covert application of the stimulus-response translation processes of the currently irrelevant task (see below for details), we were particularly interested whether such an impact is asymmetrical between the tasks, arising, for instance, from a larger degree of automatic application of symmetry judgment to a perceived letter string than of applying the (arbitrary) stimulus-response rules of the Eriksen task.
Concerning the investigation of a possible role of aesthetic stimulus appreciation in attention tasks, we first obtained aesthetic beauty ratings for the different kinds of letter strings that we administered in the Eriksen task and in the symmetry judgment task. These ratings yielded a clear pattern of aesthetic preference, allowing for a comparison with response performance in the attention tasks for stimuli falling in different categories of aesthetic value. To foreshadow our results, we obtained an unexpected disadvantage of symmetry judgment for letter strings which were symmetrical but heterogeneous (e.g., HXH) compared to asymmetrical letter strings (e.g., HHX) which strikingly contrasted with the pattern of aesthetic appreciation.
2. Aesthetic Ratings
To relate the findings that were obtained in the Eriksen task and the symmetry task to aesthetical values of the stimuli administered, we obtained ratings of all individual stimuli from an independent sample of participants.
2.1. Method
2.1.1. Participants
One hundred and twenty-four students of the Helmut Schmidt University/University of the Federal Armed Forces Hamburg, majoring in Psychology (1st Trimester) or Educational Science (4th Trimester) volunteered to participate in the study. 87 were men, 36 were women, and one was diverse. Their mean age was 22.51 years with a SD = 2.72 (range = 19-33). One additional participant provided incomplete data, and was thus excluded from further analysis.
2.1.2. Procedure
The study was conducted in a lecture hall. The stimuli were presented in a projection on a central screen. A single presentation sequence was used for all of the participants. After an introduction, the eight stimuli featuring H and S were shown next to each other in one projection in a systematic order (i.e., HHH, HHS, HSH, HSS, SHH, SHS, SSH, and SSS) in order to familiarize the participants with the complete set. Then, the stimuli were shown individually for the ratings. This happened in the sequence given in parenthesis. These were followed by an aesthetic threshold item for each subsample (H and S, H, and X), with which we asked the participants to estimate how well they think that they could judge the aesthetic value of the strings on a scale ranging from 0 to 10 (not well-well). The mean self-assessment of the ability to judge the aesthetic string was 5.67 (SD = 2.45), and 5.57 (SD = 2.53), respectively, for the H and S followed by the H and X string set.
The procedure was repeated for the eight stimuli featuring H and X. The participants were then debriefed. The session lasted about 20 minutes.
2.2. Results and Discussion.
All of the experimental data in this article were analyzed using IBM SPSS 25 [48]. The figures were created using the software RStudio [49] with the packages ggplot2 [50] and Rmisc [51].
The stimuli were categorized according to the structure of the letter string (i.e., symmetrical-homogeneous, symmetrical-heterogeneous, asymmetrical-double letter at top, asymmetrical-double letter at bottom), according to whether the string was taken from the set involving the letter S or from the set that involves the letter X, and according to whether the central letter was an H or an S/X (see Table 1).
Figure 1 displays the mean ratings. An Analysis of Variance (ANOVA) with repeated measures on the factors Stimulus Set (S, X), Central Letter (H, S/X), and String Structure (HHH/SSS, HHS/SSH, HSS/SHH, SHS/HSH), ANOVA yielded a main effect of String Structure (F(2.09, 257.33) = 88.97, p < 0.001, MSE = 13.83, ηp2 = 0.42) (this result is reported with Greenhouse–Geisser correction as the Mauchly test for sphericity indicated a violation of the assumption of independent variance across the four levels of the factor. The Greenhouse–Geisser correction was also applied to all other effects reported in this article for which the Mauchly test was significant). Neither the main effect of S-Set/X-Set nor the main effect of Central Letter H/S or any of the interactions reached statistical significance (all ps > 0.07). Symmetrical strings were overall conceived of as more aesthetic than asymmetrical strings, as can be seen in Figure 1. A separate ANOVA, confined to the symmetrical letter strings (i.e., left half of Table 1), demonstrated that homogeneous letter strings were rated as more aesthetic than heterogeneous letter strings (F(2.09, 257.33) = 88.97, p < 0.001, MSE = 13.83, ηp2 = 0.42).
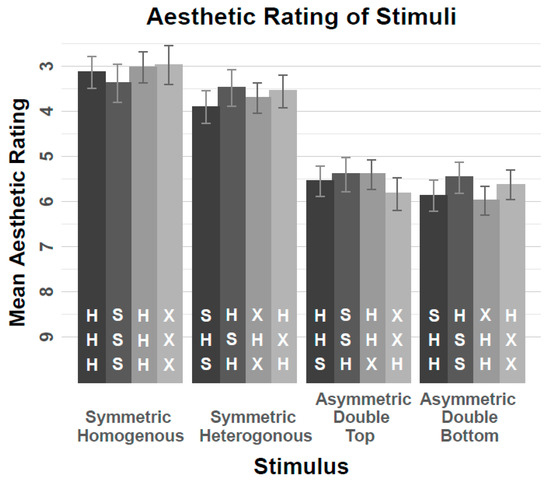
Figure 1.
Mean aesthetic ratings of the stimuli used in the experiments of the current study as a function of Stimulus Structure, Stimulus Set and Central Letter. Error bars indicate 95% within-confidence-intervals. Note, that lower values refer to higher aesthetic value, therefore the y-axis was inverted for purposes of illustration.
The results were clear-cut. Letter strings that involved a symmetrical sequence of elements received higher aesthetic ratings than letter strings involving an asymmetrical sequence of elements. In addition, among the letter strings involving a symmetrical sequence of elements, homogeneous letter strings received somewhat higher aesthetic ratings than heterogenous letter strings, although this effect was much smaller than the difference of letter strings that involved a symmetrical sequence of elements and letter strings involving an asymmetrical sequence of elements. Interestingly, the (mirror) symmetry of individual letters did not yield any effect, which is, the ratings for strings involving Ss and corresponding strings involving Xs were virtually identical.
Concerning the experiments of the current study, these results are noteworthy for at least two reasons. First, there seems so far, no reason to assume that aesthetic appreciation plays a substantial role for the flanker compatibility effect, as typical compatible and incompatible Eriksen task stimuli (presented in the first and second column of Table 1, respectively) hardly differed concerning their rated aesthetic value. On the other hand, second, there was a very close correspondence of aesthetic judgements and stimulus symmetry/asymmetry, which adds to a large number of observations of preference for symmetry, as introduced above. The correspondence is so close in this case, one could almost feel tempted to consider the symmetry judgement task as a proxy of an aesthetic judgment task [13].
3. Experiment 1
Experiment 1 conceptually replicated Experiment 2 of Wendt et al. [47] and extended it in various ways. First, whereas all of the letter strings administered in the Eriksen task and the homogeneous/heterogeneous judgment task in the study of Wendt et al. [47] were symmetrical concerning the sequence of letters, Experiment 1 of the current study involved an additional (same-sized) set of asymmetrical letter strings (i.e., HHS, SSH, HSS, SHH, top right quarter of Table 1). Adding these stimuli allowed for us to replace the homogeneous/heterogeneous judgment task by a symmetrical/asymmetrical judgment task (symmetry referring to the sequence of elements of the letter strings, rather than to the perceptual symmetry of individual letters or the strings), which was our second modification. We, again, intermixed trials of a visual search task after cues predicting the Eriksen task or the symmetry task to assess the distribution of visual attention across the three letter positions as a function of cue-based preparation. In contrast to using digit strings including a target digit at one of the three positions of that string, the search task in the current study involved geometrical shapes, presented in black or white color as stimulus elements, and target stimuli were defined by specific shape-color conjunctions (i.e., white circle and black square). Assuming that preparing for the Eriksen task involves focusing of visual attention to the expected target location, whereas preparing for a symmetry/asymmetry judgment involves broadening the attentional focus across the area of the expected stimulus constellation, we expected to replicate Wendt et al.’s [47] findings of a more pronounced center-to-periphery gradient of the search times in search task trials after an Eriksen task cue than after a symmetry task cue. Wendt et al. [47] observed no corresponding effect of the task previously executed, contrasting with the task cueing effect on the search time gradient. That is, the search time gradient was unaffected by whether the directly preceding trial involved the Eriksen task, assumed to be associated with focusing attention on the central position, or the homogeneous/heterogeneous judgment task, assumed to be associated with defocusing visual attention, that is, more evenly distributing the attentional weights across the positions of the elements that make up the stimuli for the two tasks.
Although analyzing the search time patterns in the probe task was the main method of assessing attentional processing, assessment of performance in the Eriksen task and in the symmetry task (henceforth context tasks) as a function of the structure of the stimulus was interesting, also, for number of reasons. For one thing, the design of our study allowed for us to explore a possible influence of the aesthetic value detached of an explicit contemplation on preferences or the beauty of a given stimulus, as well as possible evaluative judgements replacing descriptive judgements of the symmetrical property in speeded forced choice tasks while bearing in mind that the beauty of the letter strings is closely related to the symmetrical properties. With regard to the Eriksen task, a non-replication of the flanker compatibility effect (i.e., worse performance for symmetrical-heterogeneous stimuli than for symmetrical-homogeneous stimuli) would be surprising, given the vast number of previous studies that yielded the effect. By contrast, no clear prediction seems to converge from the literature concerning responding to the asymmetric letter strings. A previous study, attempting to gradually vary flanker compatibility, manipulated the number of flanker stimuli that were identical with the target letter [52]. In that study, symmetrical seven-letter strings were made up of a central target letter H or S and a varying number of incompatible flankers or redundant targets (e.g., SSSHSSS, SSHHHSS, SHHHHHS, or HHHHHHH). Consistent with the idea that the flanker compatibility effect is mainly driven by the competition of responses that are associated with the target and the flankers and that flanker-based response activation accumulates over corresponding stimulus letters, RT, and error rate increased with the number of incompatible flankers in the string. Transferring these results to the case of asymmetric letter strings that were used in the current study leads to the prediction of worse performance than for homogeneous strings, but better performance than for symmetrical-heterogeneous strings. On the other hand, Friedenberg [53], presenting a vertical and a horizontal line as targets and flankers in three-element strings, presented in horizontal or vertical format, observed the advantageous processing of homogeneous strings when compared to both symmetrical-heterogeneous and asymmetrical strings, but no difference of response performance between the latter two conditions. Friedenberg [53] attributed the disadvantage of asymmetric strings as compared to homogeneous strings to pop-out of the incompatible flanker, which likely diverts attention away from the target stimulus.
Contrasting attentional sets adopted in the context of Eriksen and symmetry judgment tasks also allowed for us to analyze effects of covert execution of a currently irrelevant task on performance in the relevant task. To this end, the same set of motor responses was used for both tasks, allowing for us to assess the effects of response congruency between these tasks. Faster and less error-prone responses when a current stimulus calls for the same response in a currently irrelevant task are a common finding in task switching protocols [54,55]. Thus, such effects reflect some form of translation of the current stimulus into its associated response according to the rules of the irrelevant task.
3.1. Method
3.1.1. Participants
Sixteen female and eight male students of the Medical School Hamburg, ranging in age from 19 to 30 years, participated in a single-session experiment in exchange for partial fulfillment of course requirements.
3.1.2. Apparatus and Stimuli
The experiment took place in a dimly lit and sound-attenuated room. The stimuli were presented on a 22”-LCD-Display screen. The responses were detected by the Serial Response Box, Model 200a (Psychology Software Tools, Inc). Stimulus presentation and recording of response times were controlled by E-Prime 2.0 (Psychology Software Tools, Inc.). The participants were seated approximately 60 cm in front of the screen. They responded by pressing a left-sided and a right-sided response key, while using the index fingers or the middle fingers of their left and right hand, respectively. The background color was medium gray. The stimuli used for the Eriksen flanker task and for the symmetry judgment task were the three-letter strings displayed in the top half of Table 1. Each letter extended 0.5 cm horizontally and 0.6 cm vertically and was presented inside a square frame extending 1.9 cm in width and height. The three squares were arranged to a vertical structure (all squares horizontally aligned), extending 5.6 cm in height, and were presented in the center of the screen. Letters and frames were presented in red color. In the Eriksen flanker task, the participants were instructed to press the left-sided key if the central letter was an H and the right-sided key if the central letter was an S. In the symmetry judgment task, the participants were instructed to press the left-sided key if the sequence of letters was symmetrical and the right-sided key if the sequence of letters was asymmetrical. Stimuli used for the search task were presented in the same three-square frame structure used for the Eriksen task and the symmetry task. Instead of the stimulus letters, three-element strings of disks and squares, being presented in black or white color and extending 1.7 cm horizontally and vertically each, were presented inside the three-square frame. Each search task stimulus included either a white circle or a black square (never both), which acted as target stimuli in the search task. The participants were instructed to press the left-sided key if they detected a white circle and to press the right-sided key if they detected a black square. The target stimulus (i.e., white circle or black square) was presented in either the top, central, or bottom square frame. The other two square frames of a search task stimulus were filled with a white square and a black circle, which acted as non-target stimuli. By this arrangement, 12 different strings of elements were constructed that comprised the complete set of stimuli of the search task (see Figure 2. for exemplary stimuli). The words “Mitte” (“center”), indicating the Eriksen task, and “Gesamt” (“entire”), indicating the symmetry task, presented in red color, were used as cues. They extended 4.7 cm and 6.8 cm horizontally 1.9 cm vertically.
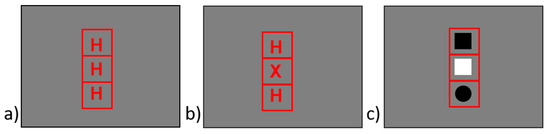
Figure 2.
Examples of (a) a homogenous symmetrical stimulus, (b) a heterogeneous symmetrical stimulus in Experiment 2 (“X” replacing the “S” of Experiment 1), and (c) a stimulus for a probe task trial with the target (black square) presented in the top location.
3.1.3. Procedure
Each trial started with the presentation of a task cue for 100 ms. After a blank screen of 700 ms, the imperative stimulus (letter string or string of geometrical shapes, being presented inside the three-square shape), was presented for 100 ms. A block of trials consisted of 20 trials of the Eriksen task, 20 trials of the symmetry task, and 24 trials of the search task. Eriksen task trials and symmetry task trials were always validly indicated by the preceding cue. Each letter string was presented in four trials of the Eriksen task and in four trials of the symmetry task. Regarding the search task, each of the 12 stimulus strings was presented twice, occurring once in a trial that was associated with a cue indicating the Eriksen task and once in a trial that was associated with a cue indicating the symmetry task. Apart from these constraints, the selection of task and stimulus per trial was random. After the presentation of the imperative stimulus, the screen remained blank until a response was given or 3,000 ms elapsed. If the response was correct, the cue of the following trial was presented after an interval of 500 ms. If the response was incorrect, the German word “Falsch” (Incorrect) was presented in yellow color in the screen center for 500 ms and the cue of the following trial was presented after another blank screen interval of 500 ms. Similarly, the message “Keine Antwort gegeben” (No response given) was presented in yellow color in the screen center for 500 ms and the cue of the following trial was presented after another blank screen interval of 500 ms if no response was given during the 3,000 ms interval. Between blocks of trials, the participants were free to take a rest. The participants worked through 10 blocks of 64 trials, each. Before these experimental trials, the participants received some practice with the three tasks.
3.2. Results
Data of the practice blocks, as well as the first three trials of each experimental block, were treated as warm-up trials and were excluded from the analyses. We also excluded all data of trials following a trial associated with an erroneous response. The analyses of probe task performance were confined to trials that were preceded by a letter task trial. The RT analyses were confined to data from trials associated with a correct response and to data not deviating more than 2.5 SDs from the participant’s mean of the respective condition.
3.2.1. Probe Task
The data of one participant were excluded from the analysis of the probe task due to two empty cells in the probe data RT. A 2x2x3 ANOVA with repeated measures on the factors Cue (Eriksen, Symmetry), Task on Preceding Trial (Eriksen, Symmetry), and Probe Position (top, center, bottom) run on the mean RTs obtained in search task trials. Figure 3, left panel, displays the mean RTs and error rates. The main effect of the Probe Position reached significance (F(2, 44) = 11.59, p < 0.001, MSE = 10284.64, ηp2 = 0.35), as well as the two-way interaction of Probe Position and Previous Cue (F(2, 44) = 4.33, p = 0.02, MSE = 10140.87, ηp2 = 0.16) and of Probe Position and Cue (F(2, 44) = 3.76, p = 0.03, MSE = 11785.82, ηp2 = 0.15) reached the level of significance. Planned comparison contrasting quadratic trends for trials that were associated with an Eriksen task cue and trials associated with a symmetry task cue reached the level of significance (F(1, 22) = 6.76, p = 0.02, MSE = 11566.61, ηp2 = 0.24), demonstrating a more pronounced center-to-periphery gradient of search time across the three probe positions after an Eriksen task cue as compared to after a symmetry task cue. No further main or interaction effects reached statistical significance.
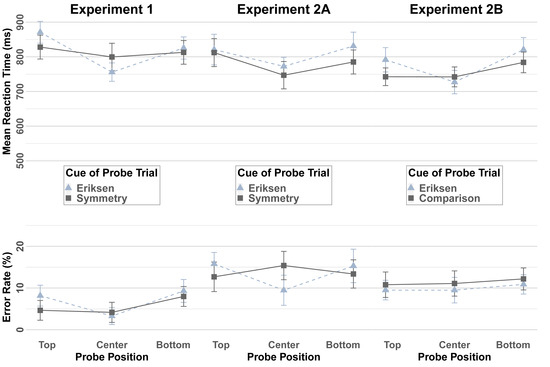
Figure 3.
Reaction times (top panel) and error rates in the probe task of Experiments 1, 2A, and 2B as a function of Probe Position and Cue. Error bars indicate 95% within-confidence intervals.
The analogous ANOVA for the error rates yielded a significant main effect of Probe Position (F(1.6, 34.12) = 6.00, p = 0.01, MSE = 0.01, ηp2 = 0.21), reflecting that fewer errors were made for probe task targets that were presented in the central when compared to in the peripheral positions. The interaction of Probe Position and Cue failed to reach statistical significance, the planned comparison of the quadratic trends mirrored the RT results descriptively, demonstrating a steeper center-to-periphery gradient in trials associated with an Eriksen task cue than in trials associated with a symmetry task cue, slightly failing to reach the level of significance (F(2, 44) = 2.41, p = 0.1, MSE = 0.005, ηp2 = 0.1, and F(1, 22) = 3.88, p = 0.06, MSE = 0.004, ηp2 = 0.15), respectively. No other main or interaction effect reached the level of significance.
3.2.2. Context Tasks
A 4x2x2 ANOVA with repeated measures on the factors Stimulus Structure (HHH/SSS, HHS/SSH, HSS/SHH, SHS/HSH), Response Congruency (Congruent, Incongruent), and Task (Eriksen, Symmetry) was run on the mean RT and the error data. Figure 4, left panel, displays the mean RTs and error rates. In the RT analysis, the main effects of Stimulus Structure, Response Congruency, and Task reached significance (F(1.99, 45.76) = 8.54, p = 0.001, MSE = 17411.96, ηp2 = 0.27; F(1, 23) = 8.64, p = 0.007, MSE = 8596.48, ηp2 = 0.27; and F(1, 23) = 32.07, p < 0.001, MSE = 45814.28, ηp2 = 0.58, respectively). Congruent trials yielded faster responses than incongruent trials, and the Eriksen flanker task was, overall, associated with faster responses than the symmetry task, as can be seen in Figure 4. Furthermore, the interaction effect of Stimulus Structure and Task reached the level of significance (F(2.44, 56.04) = 16.86, p < 0.001, MSE = 10687.90, ηp2 = 0.42), reflecting that RTs in the Eriksen task were similar for the different stimuli, whereas there appeared to be a three-level pattern of RTs in the symmetry task (i.e., fastest responses for symmetrical-homogeneous, slowest responses for symmetrical-heterogeneous, and responses of intermediate speed for asymmetrical stimuli). No other main or interaction effects reached the level of significance.
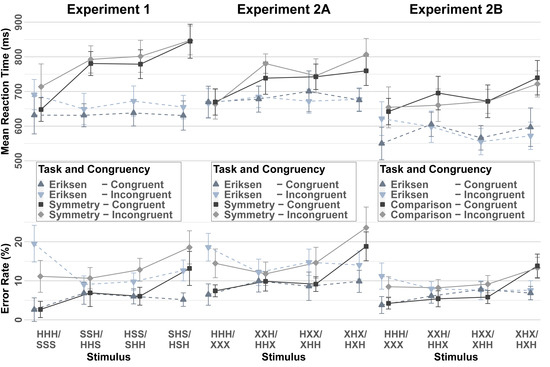
Figure 4.
Reaction times (top panel) and error rates in the context tasks of Experiments 1, 2A, and 2B as a function of Stimulus Structure, Task, and Congruency. Error bars indicate 95% within-confidence intervals.
In the analogous ANOVA for the error rates the main effects of Stimulus Structure and Congruency reached significance (F(3, 69) = 10.19, p < 0.001, MSE = 0.003, ηp2 = 0.31; F(1, 23) = 47.45, p < 0.001, MSE = 0.009, ηp2 = 0.68; respectively). In addition, the interaction effects of Stimulus and Congruency (F(3, 69) = 11.90, p < 0.001, MSE = 0.004, ηp2 = 0.34) as the Congruency Effect appeared to be larger for symmetrical-homogenous stimuli than for all other three stimulus types. In addition, the interaction of Stimulus and Task (F(3, 69) = 9.18, p < 0.001, MSE = 0.004, ηp2 = 0.29) reached significance as, in the Eriksen task trials, the error rate seemed to be similar for the four stimulus types, whereas in the symmetry task the pattern of error rates resembled the RT results (best performance for symmetrical-homogeneous, worst performance for symmetrical-heterogeneous, and intermediate performance for asymmetrical stimuli). Finally, the three-way interaction reached the level of significance (F(1.9, 43.94) = 6.35, p < 0.001, MSE = 0.005, ηp2 = 0.22), reflecting that the congruency effect seemed to be comparable across the stimulus conditions in the symmetry task, whereas, in the Eriksen task, it was markedly larger for symmetrical-homogeneous stimuli than for the other stimulus conditions (see Figure 4).
We repeated the ANOVAs on data from trials of the context tasks, separately for the Eriksen task and for the symmetry task, to further analyze the effects of the different types of stimuli. To control for alpha accumulation, we raised the alpha level in these additional analyses to 0.05/3 (i.e., we divided our original 5% level by the number of comparisons made in each additional ANOVA). Therefore, only results with a p-value below 0.05/3 were reported. The RT analysis of the Eriksen task trials yielded neither a significant main effect, nor a significant interaction effect. In the analogous ANOVA of the error rates both the main effect of Congruency and the main effect of the Stimulus Structure were significant (F(1, 23) = 41.12, p < 0.001, MSE = 0.007, ηp2 = 0.64, and F(3, 69) = 5.43, p = 0.002, MSE = 0.002, ηp2 = 0.19, respectively). Furthermore, the interaction of these two factors reached significance (F(1.71, 39.27) = 15.33, p < 0.001, MSE = 0.008, ηp2 = 0.40). As can be seen in Figure 4, this was mainly driven by the fact than the congruency effect was particularly large for the symmetrical-homogeneous stimuli.
In the RT ANOVA for the symmetry task, solely the main effects of Stimulus Structure reached significance (F(3, 69) = 20.84, p < 0.001, MSE = 10846.41, ηp2 = 0.48). The interaction of Congruency and Stimulus Structure did not reach the level of significance (F(2.43, 55.96) = 1.51, p = 0.22). In the corresponding analysis of error rates, both of the main effects reached significance, again (F(1, 23) = 15.32, p = 0.001, MSE = 0.01, ηp2 = 0.40, and F(2.18, 50.15) = 10.86, p < 0.001, MSE = 0.01, ηp2 = 0.32, respectively). The interaction of Congruency and Stimulus Structure did not reach the level of significance (F(3, 69) = 1.41, p = 0.25).
3.3. Discussion
Experiment 1 yielded several interesting results. First, the search times of the probe task were associated with markedly different patterns in trials associated with the two different context task cues, thus replicating and extending Wendt et al.’s [47] findings. Specifically, the center-to-periphery gradient of search times was more pronounced after an Eriksen task cue than after a symmetry task cue, which suggested cue-based focusing of visual attention to the position of the central stimulus element in the former case and more evenly distributed attentional weights across the three element positions in the latter. By contrast, no comparable difference was found after execution of the Eriksen task versus after execution of the symmetry task on the preceding trial. This lack of persistence of the attentional set adopted on the previous trial might be explained by efficient “overwriting” of the attentional set during preparation for the upcoming task [47].
The performance in the symmetry task differed strongly from performance in the Eriksen task. Whereas in the Eriksen task, RTs were overall substantially faster and comparable across the four types of stimulus structure (thus showing no sign of the classical flanker compatibility effect), RTs in the symmetry task were slower by more than 100 ms, except for the symmetrical-homogeneous condition. Response congruency effects between tasks were of comparable size for the two tasks, despite this asymmetry in task-specific response speed. That is, in Eriksen task trials irrelevant symmetry judgment concerning the complete letter string occurred to a certain degree despite focusing of attention on the central letter sufficiently strong to prevent incompatible flankers from interfering. Moreover, response activation brought about by such irrelevant symmetry judgment occurred quickly enough to affect responding in the (faster) Eriksen task, much like irrelevant Eriksen task’s identification of the central letter affected responding in the slower symmetry task. Although these results may dismiss a simple race model of the congruency effect, the fact that in the Eriksen task the congruency effect was largest in the symmetrical-homogeneous condition, that is, in the condition that is associated with particularly fast symmetry task responses, suggests that the speed of translating a current stimulus into its response according to the rules of the irrelevant task might be a relevant determinant of the size of the congruency effect.
Whereas performance in the Eriksen task hardly differed across the stimulus conditions, performance in the symmetry task displayed a remarkable three-level pattern. More precisely, performance was best for symmetrical-homogenous stimuli and the worst for symmetrical-heterogeneous stimuli, performance for asymmetrical stimuli falling at an intermediate level. This pattern of results strongly deviates from the pattern of aesthetical ratings of the stimuli, lending no support to the notion that participants could have substituted the instructed symmetry judgment by an aesthetic judgment.
Although facilitated processing of symmetry judgment of homogeneous stimuli might not be surprising, the impairment of detecting the symmetry of heterogeneous letters strings deserves discussion. At least two straightforward explanations for this impairment should be considered. The first of these accounts relates to the fact that in our Experiment 1 symmetry referred to the sequence of elements of the letter strings. By consequence, including asymmetrical stimulus elements, results in the formation of letter strings that require being categorized as symmetrical despite being perceptually asymmetrical (e.g., SSS, HSH). Although including asymmetrical elements might be helpful to categorize an asymmetrical string correctly and may be of little consequence for homogeneous strings (whose symmetry might be quickly inferred from their homogeneity), it might cause some form of cognitive conflict [56] when present in symmetrical-heterogeneous string, interfering with the required symmetry response. This reasoning relates to a vast number of findings demonstrating congruency effects regarding global stimulus properties when responding to local stimulus feature [57]. The second account assumes conflict that is brought about by the particular set of stimuli used. Precisely, including (a substantial proportion of) homogeneous strings in the stimulus set might induce a strategy of examining homogeneity/heterogeneity, first, resulting in particularly fast responses to homogeneous stimuli and requiring further processing for heterogeneous stimuli. Assuming that a heterogeneous judgment is associated with priming of the asymmetry response, symmetrical letter strings should suffer from conflict of this response activation with the required response.
A straightforward method to come to a decision concerning the two accounts is to replicate Experiment 1, replacing the asymmetrical stimulus element S by a symmetrical one. This is what we did in Experiment 2 (replacing the letter S by the letter X).
4. Experiments 2A and 2B
We should be able to prevent such impairment by replacing this element by a symmetrical one. if the impairment of recognizing the symmetry of the letter strings SHS and HSH goes back to the inclusion of an asymmetrical constituent element, preventing perceptual symmetry of the string. However, such replacement does not seem to alter the situation possibly resulting in an “examine homogeneity first” strategy. According to the “heterogeneous primes asymmetrical” account, we would thus still expect particularly bad performance when judging the symmetry of symmetrical-heterogeneous letter strings, because such strings should prime the incorrect asymmetry response in the first place. Likewise, replacing S by X would not seem to alter the attentional requirements that are associated with the Eriksen task and with the symmetry task. Therefore, we also expected to replicate the steeper search time gradient after Eriksen task cues than after symmetry task cues. As concerns response congruency between the tasks, it will be interesting to see if, again, these effects are comparable for the two tasks, despite marked overall difference in RT and, in particular, whether increasing response speed for the symmetrical-heterogeneous letter strings in the symmetry task would be associated with an increase in the congruency effect that is yielded by these stimuli in the Eriksen task.
A major purpose of the experiments of our study is to contrast attentional strategies for tasks involving an identical set of stimuli and motor responses (i.e., Eriksen flanker task and symmetry judgment task). Experiments 2A and 2B extended this approach by allowing us to contrast, in addition, attentional strategies for tasks involving the same sets of stimuli, motor responses, and the same stimulus-response mappings, which are, however, associated with task instructions, suggesting different attentional requirements. Wendt, Garling, Luna-Rodriguez, and Jacobsen previously used this approach [58]. These authors presented asymmetrical five-letter strings (i.e., HHHSS, HHSSS, SSSHH, and SSHHH) to two different groups of participants, asking one group to identify the central letter of each string and another group of participants to identify the letter that occurred three times in the string. As for the set of letter strings used the central letter was always identical to the letter occurring three times, both tasks could, in principle, be solved by the same cognitive operations. However, RT data suggested that the two groups applied different processing strategies. Most importantly, search time gradients in intermixed trials of a visual search task displayed a more pronounced center-to-periphery gradient, indicating stronger focusing of attention to the location of the central letter, in the group that was instructed to identify the central letter than in the group instructed to identify the letter occurring three times in the string. Intermixing search task trials thus provided an unobtrusive method of assessing participants’ adherence to the task instruction.
Experiments 2A and 2B of the current study adopted an analogous approach by exploiting the fact that symmetry/asymmetry of the letter strings used corresponds to the letters presented at the top and at the bottom position being identical or different from each other. This allowed for us to use identical procedures, including the set of stimuli, responses, and stimulus-response mappings, with the only exception that participants were either instructed to judge the symmetry of the letter string (Experiment 2A) or to compare the identity of the two peripheral letters of the string (Experiment 2B). Adhering to the latter instruction might lead to an attentional strategy of inhibiting the deployment of attention to the central letter position, particularly so because the processing of this letter is liable to interfere with judging the peripheral letters as identical if itself instantiates the other letter (for evidence of splitting the focus of visual attention to non-contiguous spatial locations and for physiological evidence) [59,60,61,62]. Such a strategy might reveal itself in a reversed search time gradient (i.e., the disadvantageous processing of centrally presented search task targets). Moreover, it might reduce the disadvantage of categorizing symmetrical-heterogeneous strings if reduced allocation of attention to the (deviant) central letter is associated with a less powerful “heterogeneity detection response”, which might, in turn, decrease the assumed priming of the asymmetry response (or “different response”, according to the task instructions that were employed in Experiment 2B).
4.1. Method
4.1.1. Participants
17 female and 11 male students of the Medical School Hamburg, ranging in age from 20 to 33 years, participated in a single-session experiment in exchange for partial fulfillment of course requirements in Experiment 2A, and 21 female and five male students of the Medical School Hamburg, ranging in age from 20 to 26 years, participated in a single-session experiment in exchange for partial fulfillment of course requirements in Experiment 2B.
4.1.2. Apparatus and Stimuli
The apparatus did not differ from Experiment 1. The stimuli were also identical to the stimuli used in Experiment 1 with the following exceptions. First, all instances of the letter S in each of the letter strings was replaced by the letter X. Second, the letters “M” and “S”, abbreviating the German words “Mitte” (center) and “Symmetrie” (symmetry), presented in red color and extending 3.2 cm and 1.8 cm horizontally and 2.9 cm vertically, respectively, were used as cues to indicate the Eriksen task (M) and the symmetry/comparison task, respectively.
4.1.3. Procedure
The procedure of Experiment 2A was the same as described in Experiment 1. The procedure of Experiment 2B only differed concerning the instruction of the “symmetry task” (henceforth, comparison task). Regarding this task, the participants were instructed to press the left key if the letters presented at the top position and the bottom position were identical and to press the right key if these letters differed from each other.
4.2. Results
The same analyses as in Experiment 1 with the same criteria for data exclusion were conducted.
4.2.1. Experiment 2A
Probe Task
The data of one participant of Experiment 2A were excluded from the analysis of the probe task due to one empty cell in the probe data RT. Figure 3, as in the middle panel, displays the mean RTs and error rates. In the 2x2x3 ANOVA with repeated measures on the factors Cue (Eriksen, Symmetry), Task on Preceding Trial (Eriksen, Symmetry), and Probe Position (top, center, bottom) on the RT data, the main effect of Probe Position reached the level of significance (F(2, 52) = 5.53, p = 0.007, MSE = 18344.63, ηp2 = 0.18) due to facilitated responses to the central position when compared to the outer ones. No other main or interaction effect reached the level of significance. The analogous ANOVA on the error rates yielded a significant interaction of Probe Position and Cue (F(1.5, 39.08) = 5.14, p = 0.02, MSE = 0.02, ηp2 = 0.17). The planned contrast comparing the quadratic trends displayed a steeper gradient of search time across the three positions following an Eriksen task cue than following a symmetry task cue (F(1, 26) = 6.82, p = 0.02, MSE = 0.02, ηp2 = 0.21). No further main or interaction effect reached significance.
Context Task
Figure 4, as in middle panel, displays the mean RTs and error rates. In the 4x2x2 ANOVA with the factors Stimulus Structure (HHH/XXX, HHX/XXH, HXX/XHH, XHX/HXH), Response Congruency (Congruent, Incongruent), and Task (Eriksen, Symmetry) of the RT data, the main effects of Stimulus Structure and of Task were significant (F(3, 81) = 10.13, p < 0.001, MSE = 8595.30, ηp2 = 0.27, and F(1, 27) = 11.50, p = 0.002, MSE = 35527.36, ηp2 = 0.30, respectively). In addition, the interaction of these two factors reached significance (F(3, 81) = 6.58, p < 0.001, MSE = 8883.33, ηp2 = 0.20). No other main or interaction effects reached significance. The analogous ANOVA on the error rates yielded significant main effects of Stimulus Structure and Congruency (F(1.85, 50.1) = 8.72, p = 0.001, MSE = 0.01, ηp2 = 0.24, and F(1, 27) = 92.24, p < 0.001, MSE = 0.004, ηp2 = 0.77, respectively). The Stimulus Structure entered into significant two-way interactions with Congruency (F(3, 81) = 4.41, p = 0.006, MSE = 0.006, ηp2 = 0.14) and with Task (F(1.97, 53.28) = 10.50, p < 0.001, MSE = 0.01, ηp2 = 0.28). The pattern of this interaction closely resembled the pattern observed in Experiment 1, as can be seen in Figure 4.
The ANOVA (alpha = 0.05/3) confined to RT data from the Eriksen flanker tasks showed no significant main or interaction effect at all. The analogous ANOVA on the error rates (alpha = 0.05/3) only yielded a significant main effect of Congruency (F(1, 27) = 31.48, p < 0.001, MSE = 0.007, ηp2 = 0.54). The ANOVA confined to RT data of the symmetry task only yielded a significant main effect of Stimulus Structure (F(3, 81) = 18.87, p < 0.001, MSE = 7532.14, ηp2 = 0.41), replicating the three-level RT pattern observed in Experiment 1. In the corresponding ANOVA on the error rates, both main effects of Stimulus Structure and Congruency reached significance (F(1.52, 40.92) = 12.24, p < 0.001, MSE = 0.023, ηp2 = 0.31, and F(1, 27) = 34.42, p < 0.001, MSE = 0.004, ηp2 = 0.56, respectively). Most notably, responding to symmetrical-heterogeneous stimuli was associated with more errors than with any other stimulus condition.
4.2.2. Experiment 2B
Probe Task
Figure 3, as in right panel, displays the mean RTs and error rates. In the RT analysis, the main effect of Probe Position reached significance (F(2, 50) = 7.77, p = 0.001, MSE = 15339.59, ηp2 = 0.24), reflecting faster responding to centrally presented search task targets than to peripherally presented search task targets. This was qualified by a two-way interaction with Cue (F(1.6, 40.01) = 3.66, p = 0.04, MSE = 10225.35, ηp2 = 0.13). The center-to-periphery gradient of the search times was more pronounced after the Eriksen task cue than after the comparison task cue, as confirmed by the planned comparison of the quadratic trends (F(1, 25) = 5.39, p = 0.029, MSE = 10780.82, ηp2 = 0.18). In addition, the two-way interaction of Cue and Task on Preceding Trial reached significance (F(1, 25) = 8.36, p = 0.008, MSE = 13500.81, ηp2 = 0.25), reflecting faster responses if the cue of the current trial matched the preceding task as compared to the other letter task. No other main effect or interactions reached significance. The analogous ANOVA of the error rates did not yield any significant effects.
Context Tasks
Figure 4, as in right panel, displays the mean RTs and error rates. The 4x2x2 ANOVA with the factors Stimulus Structure (HHH/XXX, HHX/XXH, HXX/XHH, XHX/HXH), Response Congruency (Congruent, Incongruent), and Task (Eriksen, Comparison) of the RT data yielded significant main effects of Stimulus Structure and Task(F(3, 75) = 6.40, p = 0.001, MSE = 6460.14, ηp2 = 0.20, and F(1, 25) = 11.88, p = 0.002, MSE = 85602.05, ηp2 = 0.32, respectively). Both of these factors entered into a significant interaction (F(3, 75) = 4.42, p = 0.006, MSE = 8304.57, ηp2 = 0.15). The result pattern closely resembled the pattern obtained in Experiment 1 and in Experiment 2A, as can be seen in Figure 4. Finally, the interaction of Congruency and Stimulus Structure reached the level of significance (F(3, 75) = 3.87, p = 0.01, MSE = 6007.73, ηp2 = 0.13). Figure 4 suggests that this was mainly driven by the fact that in the Eriksen task the congruency effect was the largest for homogeneous stimuli whereas in the comparison task the congruency effect was largest for asymmetrical letter strings with the double letter at the top position. No other effects reached the level of significance. In the analogous ANOVA of the error rates the main effects of Stimulus Structure and Congruency reached significance (F(3, 75) = 11.99, p < 0.001, MSE = 0.002, ηp2 = 0.32, and F(1, 25) = 26.18, p < 0.001, MSE = 0.002, ηp2 = 0.51, respectively). Both of the factors entered into a significant two-way interaction (F(3, 75) = 5.16, p = 0.003, MSE = 0.005, ηp2 = 0.17). Stimulus Structure also entered into a significant two-way interaction with Task (F(2.27, 56.66) = 8.31, p < 0.001, MSE = 0.005, ηp2 = 0.25). An inspection of Figure 4 shows that this was mainly because responding to symmetrical-heterogeneous stimuli was associated with a particularly high error rate in the comparison task. No other effects reached the level of significance.
Separately repeating the former analyses for the Eriksen task and the comparison task (with alpha = 0.5/3), yielded, for the Eriksen task, neither a main, nor an interaction effect concerning RTs. For error rates, the main effect of Congruency as well as the interaction of Congruency and Stimulus Structure reached significance (F(1, 25) = 10.18, p = 0.004, MSE = 0.003, ηp2 = 0.29, and F(2.12, 52.89) = 5.14, p = 0.008, MSE = 0.004, ηp2 = 0.17, respectively). In the analyses including only trials of the comparison task, only the main effect of Stimulus Structure reached significance for RTs (F(3, 75) = 6.63, p < 0.001, MSE = 9534.67, ηp2 = 0.21). Whereas both of the main effects of Stimulus Structure and of Congruency were significant for error rates (F(1.75, 43.62) = 16.83, p < 0.001, MSE = 0.006, ηp2 = 0.40, and F(1, 25) = 13.70, p = 0.001, MSE = 0.002, ηp2 = 0.35, respectively).
4.3. Discussion
Experiments 2A and 2B broadly replicated the findings of Experiment 1. First, the performance in the search task displayed a more pronounced center-to-periphery gradient after the Eriksen task cue than after the symmetry or comparison task cue, confirming the assumption of anticipatory focusing and defocusing of attention during preparation for the Eriksen task and the symmetry/comparison task, respectively. Second, responding in the symmetry task or comparison task again took markedly longer than responding in the Eriksen task with the exception of the condition that was associated with symmetrical-homogeneous stimuli. Third, the flanker compatibility effect in the Eriksen task performance was again absent. In light of the apparent robustness of the flanker compatibility effect across a multitude of tasks and stimulus conditions, the role of adding asymmetrical letter strings and of frequently switching to a symmetry task for selective stimulus processing deserves to be investigated in more depth in future studies. Fourth, congruency effects, albeit only significant in the error analysis, were again similar in size for both tasks, lending no support to the notion that the faster task had a larger impact on the slower task than vice versa.
Of most importance, the response performance for symmetrical-heterogeneous strings in the symmetry task was worse than for the other types of stimuli despite the usage of symmetrical stimulus elements only. This finding was predicted based on the assumption of response priming brought about by an initial homogeneous/heterogeneous judgement of the letter string (i.e., priming of the asymmetry response by detecting heterogeneity). In correspondence with the lacking impact of S and X constituent letters concerning the aesthetic appreciation of the letter strings, replacing S by X did not change the pattern of results in any of our tasks in a relevant way. In addition, all of the relevant findings occurred for both the symmetry task of Experiment 2A and the comparison task of Experiment 2B, thus providing no evidence that the attentional strategies differed under the two task instructions. In particular, there was no indication of the inhibition of deployment of attention to the central letter position in the form of a reversed search time gradient after comparison task cues in Experiment 2A.
5. General Discussion
Obtaining aesthetic beauty ratings for simple letter string stimuli yielded a clear pattern of preference for symmetrical letter sequences over asymmetrical letter sequences, being hardly affected by the identity of the constituent letters. We presented these stimuli in two different tasks whose stimulus-response assignment either corresponded to the aesthetical preference (i.e., symmetry judgment task) or was unrelated to it (i.e., Eriksen flanker task).
Regarding anticipatory adoption of sets of visual attention, the experiments of the current study corroborated Wendt et al.’s [47] findings of a more pronounced search time gradient after a cue, indicating the likely occurrence of an Eriksen task (requiring the identification of a centrally presented stimulus element) than after a cue indicating the likely occurrence of a task involving a judgment regarding a global stimulus property (or the comparison of peripheral stimulus elements). These results are consistent with anticipatory adjustment to a more focused state or defocused state of visual attention, depending on the to-be-expected demands of stimulus selection. Beyond extending previous findings, which were obtained with homogeneous/heterogeneous judgment as the “defocusing task”, to another task requiring a global stimulus judgment, the defocusing of attention in anticipation of a symmetry judgment is associated with more specific aspects that are relevant to questions of attentional management, however. For instance, assuming that symmetry detection occurs independently of the attentional setting (e.g., preattentively), frequent shifting of the attentional set to a defocused state would seem a dispensable effort, unless it is assumed that maintaining the state of focused attention needed for efficient processing of the Eriksen task constitutes an even larger effort.
Another novel aspect that is revealed by our experiments relates to the context-dependency of symmetry judgments. Observing an unexpected pattern of RTs and error rates, demonstrating advantageous processing of symmetrical-homogeneous and disadvantageous processing of symmetrical-heterogeneous stimulus strings (as compared to asymmetrical strings) raises questions regarding the strategic priming of symmetry/asymmetry responses that are based on detection of stimulus homogeneity. As such a strategy should be absent under conditions lacking homogeneous stimuli, we would expect no disadvantage of symmetrical-heterogeneous strings in a symmetry judgment task without homogeneous stimuli. Viewed from a more general perspective, the symmetry judgment findings of the current study emphasize the possibility that response-related factors, possibly being related to particular processing strategies induced by contextual factors, may strongly influence simple perceptual judgments. Although our results provided no evidence for a role of aesthetical stimulus appreciation in the symmetry judgment task, we can, of course, not dismiss the possibility that such effects were masked by presumably substantially stronger priming effects suggested to account for the impairment of responding to symmetrical-heterogeneous stimuli.
The lacking flanker compatibility effect in the Eriksen task, which was consistently observed in all three experiments, also deserves further consideration. Assuming that the stimulus conditions that were realized in our experiments are not insufficient per se to yield a performance difference for the two types of symmetrical letter strings when responding to the identity of the central element, we are inclined to conclude that adding asymmetric letter strings or/and the requirement of frequent switching to the symmetry task enhances the processing selectivity in the Eriksen task to a level not achieved in Eriksen tasks involving only symmetrical stimuli. Including asymmetrical stimulus strings might have induced a strategy of particularly strong focusing of attention to counter pop-out of the incompatible flanker stimulus in asymmetrical strings (although Friedenberg [53], observed faster responses to homogeneous stimuli than to both asymmetrical and symmetrical-heterogeneous stimuli).
Although the flanker compatibility effect was absent, the effects of congruency concerning the stimulus-response assignment in the other task were not. That is, although focusing attention to the central stimulus element in the Eriksen task sufficed to prevent the observable effects of translating the peripheral letters into their corresponding responses, covert execution of the irrelevant symmetry task affected responding in the Eriksen task. In fact, the response congruency effects did not systematically differ between the two tasks, demonstrating a substantial impact of covert application of (irrelevant) symmetry judgments on responding in the Eriksen task, despite the fact that response times were, overall, much slower in the symmetry task than in the Eriksen task. On the other hand, the congruency effects in the Eriksen task were consistently largest in the stimulus condition that was associated with the fastest responses in the symmetry task (i.e., symmetrical-homogeneous), suggesting that the ease of processing the stimulus according to the response assignment of the irrelevant task constitutes a relevant determinant of the size of the congruency effect.
In conclusion, we note that, although a clear pattern of aesthetic preference can be established even for stimuli as simply structured as the ones of the current study, the results of our experiments provided no evidence for a modulatory role of aesthetic stimulus appreciation. However, they did corroborate and extend previous findings concerning anticipatory adoption of task-specific attentional sets and revealed novel aspects concerning response priming within and between tasks.
Author Contributions
Conceptualization, S.T.K., T.J., S.K., and M.W.; Methodology, T.J. and M.W.; Software, M.W.; Validation, S.T.K., T.J., S.K., and M.W.; Formal Analysis, S.T.K. and S.K.; Investigation, T.J. and M.W.; Resources, T.J. and M.W.; Data Curation, S.T.K. and S.K.; Writing – Original Draft Preparation, S.T.K., T.J., S.K., and M.W.; Writing – Review & Editing, S.T.K., T.J., S.K., and M.W.; Visualization, S.T.K.; Supervision, T.J. and M.W.; Project Administration, T.J. and M.W. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Friedenberg, J. Perceived beauty of elongated symmetric shapes: Is more better? Psychol. Aesthet. Creat. Arts 2018, 12, 157–165. [Google Scholar] [CrossRef]
- Reber, R.; Schwarz, N. Perceptual fluency, preference, and evolution. Pol. Psychol. Bull. 2006, 37, 16–22. [Google Scholar]
- Tripp, T.M.; Bies, R.J.; Aquino, K. Poetic justice or petty jealousy? The aesthetics of revenge. Organ. Behav. Hum. Decis. Process. 2002, 89, 966–984. [Google Scholar] [CrossRef]
- Fechner, G.T. Vorschule der Ästhetik [Preliminaries of Aesthetics]; Breitkopf und Härtel: Leipzig, Germany, 1876. [Google Scholar]
- Fechner, G.T. Zur experimentalen Aesthetik; Hirzel: Leipzig, Germany, 1871. [Google Scholar]
- Perrett, D.I.; Burt, D.M.; Penton-Voak, I.S.; Lee, K.J.; Rowland, D.A.; Edwards, R. Symmetry and human facial attractiveness. Evol. Hum. Behav. 1999, 20, 295–307. [Google Scholar] [CrossRef]
- Scheib, J.E.; Gangestad, S.W.; Thornhill, R. Facial attractiveness, symmetry and cues of good genes. Proc. Biol. Sci. 1999, 266, 1913–1917. [Google Scholar] [CrossRef]
- Grammer, K.; Thornhill, R. Human (Homo sapiens) facial attractiveness and sexual selection: The role of symmetry and averageness. J. Comp. Psychol. 1994, 108, 233–242. [Google Scholar] [CrossRef]
- Cárdenas, R.A.; Harris, L.J. Symmetrical decorations enhance the attractiveness of faces and abstract designs. Evol. Hum. Behav. 2006, 27, 1–18. [Google Scholar] [CrossRef]
- Tovée, M.J.; Tasker, K.; Benson, P.J. Is symmetry a visual cue to attractiveness in the human female body? Evol. Hum. Behav. 2000, 21, 191–200. [Google Scholar] [CrossRef]
- Eisenman, R.; Rappaport, J. Complexity preference and semantic differential ratings of complexity-simplicity and symmetry-asymmetry. Psychon. Sci. 1967, 7, 147–148. [Google Scholar] [CrossRef]
- Jacobsen, T.; Höfel, L. Aesthetic judgments of novel graphic patterns: Analyses of individual judgments. Percept. Mot. Ski. 2002, 95, 755–766. [Google Scholar] [CrossRef]
- Jacobsen, T.; Höfel, L. Descriptive and evaluative judgment processes: Behavioral and electrophysiological indices of processing symmetry and aesthetics. Cogn. Affect. Behav. Neurosci. 2003, 3, 289–299. [Google Scholar] [CrossRef]
- Gartus, A.; Leder, H. The small step toward asymmetry: Aesthetic judgment of broken symmetries. i-Perception 2013, 4, 361–364. [Google Scholar] [CrossRef]
- Tinio, P.P.; Leder, H. Just how stable are stable aesthetic features? Symmetry, complexity, and the jaws of massive familiarization. Acta Psychol. 2009, 130, 241–250. [Google Scholar] [CrossRef]
- Mongoven, C.; Carbon, C.C. Acoustic Gestalt: On the perceptibility of melodic symmetry. Music. Sci. 2017, 21, 41–59. [Google Scholar] [CrossRef]
- Jacobsen, T. Individual and group modelling of aesthetic judgment strategies. Br. J. Psychol. 2004, 95, 41–56. [Google Scholar] [CrossRef]
- Leder, H.; Tinio, P.P.L.; Brieber, D.; Kröner, T.; Jacobsen, T.; Rosenberg, R. Symmetry Is Not a Universal Law of Beauty. Empir. Stud. Arts 2019, 37, 104–114. [Google Scholar] [CrossRef]
- Jacobsen, T.; Klein, S.; Löw, A. The Posterior Sustained Negativity Revisited—An SPN Reanalysis of Jacobsen and Höfel (2003). Symmetry 2018, 10, 27. [Google Scholar] [CrossRef]
- Makin, A.D.; Rampone, G.; Pecchinenda, A.; Bertamini, M. Electrophysiological responses to visuospatial regularity. Psychophysiology 2013, 50, 1045–1055. [Google Scholar] [CrossRef]
- van der Helm, P.A.; Treder, M.S. Detection of (anti)symmetry and (anti)repetition: Perceptual mechanisms versus cognitive strategies. Vis. Res. 2009, 49, 2754–2763. [Google Scholar] [CrossRef]
- Møller, A. Female swallow preference for symmetrical male sexual ornaments. Nature 1992, 357, 238–240. [Google Scholar] [CrossRef]
- Reber, R.; Schwarz, N.; Winkielman, P. Processing Fluency and Aesthetic Pleasure: Is Beauty in the Perceiver’s Processing Experience? Pers. Soc. Psychol. Rev. 2004, 8, 364–382. [Google Scholar] [CrossRef]
- Reber, R.; Wurtz, P.; Zimmermann, T.D. Exploring “fringe” consciousness: The subjective experience of perceptual fluency and its objective bases. Conscious. Cogn. 2004, 13, 47–60. [Google Scholar] [CrossRef]
- Barlow, H.B.; Reeves, B.C. The versatility and absolute efficiency of detecting mirror symmetry in random dot displays. Vis. Res. 1979, 19, 783–793. [Google Scholar] [CrossRef]
- Kahn, J.I.; Foster, D.H. Horizontal–vertical structure in the visual comparison of rigidly transformed patterns. J. Exp. Psychol. Hum. Percept. Perform. 1986, 12, 422–433. [Google Scholar] [CrossRef]
- Palmer, S.E.; Hemenway, K. Orientation and symmetry: Effects of multiple, rotational, and near symmetries. J. Exp. Psychol. Hum. Percept. Perform. 1978, 4, 691–702. [Google Scholar] [CrossRef]
- Corballis, M.C.; Roldan, C.E. Detection of symmetry as a function of angular orientation. J. Exp. Psychol. Hum. Percept. Perform. 1975, 1, 221–230. [Google Scholar] [CrossRef]
- Wenderoth, P.; Welsh, S. Effects of pattern orientation and number of symmetry axes on the detection of mirror symmetry in dot and solid patterns. Perception 1998, 27, 965–976. [Google Scholar] [CrossRef]
- Bertamini, M.; Friedenberg, J.D.; Kubovy, M. Detection of symmetry and perceptual organization: The way a lock-and-key process works. Acta Psychol. 1997, 95, 119–140. [Google Scholar] [CrossRef]
- Baylis, G.C.; Driver, J. Perception of symmetry and repetition within and across visual shapes: Part-descriptions and object-based attention. Vis. Cogn. 2001, 8, 163–196. [Google Scholar] [CrossRef]
- Friedenberg, J.; Bertamini, M. Contour symmetry detection: The influence of axis orientation and number of objects. Acta Psychol. 2000, 105, 107–118. [Google Scholar] [CrossRef]
- Fox, J. The use of structural diagnostics in recognition. J. Exp. Psychol. Hum. Percept. Perform. 1975, 1, 57–67. [Google Scholar] [CrossRef]
- Hershenson, M.; Ryder, J. Perceived symmetry and name matching. Bull. Psychon. Soc. 1982, 19, 19–22. [Google Scholar] [CrossRef]
- Koriat, A.; Norman, J.; Kimchi, R. Recognition of rotated letters: Extracting invariance across successive and simultaneous stimuli. J. Exp. Psychol. Hum. Percept. Perform. 1991, 17, 444–457. [Google Scholar] [CrossRef][Green Version]
- Treder, M.S. Behind the looking-glass: A review on human symmetry perception. Symmetry 2010, 2, 1510–1543. [Google Scholar] [CrossRef]
- Carmody, D.P.; Nodine, C.F.; Locher, P.J. Global detection of symmetry. Percept. Mot. Ski. 1997, 45, 1267–1273. [Google Scholar] [CrossRef]
- Wagemans, J. Skewed symmetry: A nonaccidental property used to perceive visual forms. J. Exp. Psychol. Hum. Percept. Perform. 1993, 19, 364–380. [Google Scholar] [CrossRef]
- Olivers, C.N.L.; van der Helm, P.A. Symmetry and selective attention: A dissociation between effortless perception and serial search. Percept. Psychophys. 1998, 60, 1101–1116. [Google Scholar] [CrossRef]
- Pashler, H. Coordinate frame for symmetry detection and object recognition. J. Exp. Psychol. Hum. Percept. Perform. 1990, 16, 150–163. [Google Scholar] [CrossRef]
- Niimi, R.; Yokosawa, K.; Watanabe, K. Attentional set for axis of symmetry in symmetry-defined visual search. Percept. Psychophys. 2006, 68, 1153–1162. [Google Scholar] [CrossRef][Green Version]
- Goller, J.; Mitrovic, A.; Leder, H. Effects of liking on visual attention in faces and paintings. Acta Psychol. 2019, 197, 115–123. [Google Scholar] [CrossRef]
- Mühlenbeck, C.; Liebal, K.; Pritsch, C.; Jacobsen, T. Differences in the visual perception of symmetric patterns in orangutans (Pongo pygmaeus abelii) and two human cultural groups: A comparative eye-tracking study. Front. Psychol. 2016, 7, 408. [Google Scholar] [CrossRef]
- Eriksen, B.A.; Eriksen, C.W. Effects of noise letters upon the identification of a target letter in a nonsearch task. Percept. Psychophys. 1974, 16, 143–149. [Google Scholar] [CrossRef]
- Eriksen, C.W. The flankers task and response competition: A useful tool for investigating a variety of cognitive problems. Vis. Cogn. 1995, 2, 101–118. [Google Scholar] [CrossRef]
- Gratton, G.; Coles, M.G.H.; Donchin, E. Optimizing the use of information: Strategic control of activation of responses. J. Exp. Psychol. Gen. 1992, 121, 480–506. [Google Scholar] [CrossRef]
- Wendt, M.; Kähler, S.T.; Luna-Rodriguez, A.; Jacobsen, T. Adoption of task-specific sets of visual attention. Front. Psychol. 2017, 8, 687. [Google Scholar] [CrossRef]
- IBM Corp. Released. IBM SPSS Statistics for Windows, Version 25.0; IBM Corp: Armonk, NY, USA, 2017. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; Version 3.5.1; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Miscellaneous, R. Rmisc. R Package Version 3.5.3. 2013. Available online: https://CRAN.R-project.org/package=Rmisc (accessed on 28 February 2020).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- Forster, S.E.; Carter, C.S.; Cohen, J.D.; Cho, R.Y. Parametric manipulation of the conflict signal and control-state adaptation. J. Cogn. Neurosci. 2011, 23, 923–935. [Google Scholar] [CrossRef]
- Friedenberg, J. Coactivation, pop-out, and symmetry affect line discrimination. Percept. Mot. Ski. 2000, 90, 111–120. [Google Scholar] [CrossRef]
- Rogers, R.D.; Monsell, S. Costs of a predictible switch between simple cognitive tasks. J. Exp. Psychol. Gen. 1995, 124, 207–231. [Google Scholar] [CrossRef]
- Kiesel, A.; Steinhauser, M.; Wendt, M.; Falkenstein, M.; Jost, K.; Philipp, A.M.; Koch, I. Control and interference in task switching—A review. Psychol. Bull. 2010, 136, 849–874. [Google Scholar] [CrossRef]
- Botvinick, M.M.; Braver, T.S.; Barch, D.M.; Carter, C.S.; Cohen, J.D. Conflict monitoring and cognitive control. Psychol. Rev. 2001, 108, 624–652. [Google Scholar] [CrossRef]
- Navon, D. Forest before trees: The precedence of global features in visual perception. Cogn. Psychol. 1977, 9, 353–385. [Google Scholar] [CrossRef]
- Wendt, M.; Garling, M.; Luna-Rodriguez, A.; Jacobsen, T. Exploring conflict- and target-related movement of visual attention. Q. J. Exp. Psychol. 2014, 67, 1053–1073. [Google Scholar] [CrossRef]
- Awh, E.; Pashler, H. Evidence for split attentional foci. J. Exp. Psychol. Hum. Percept. Perform. 2000, 26, 834–846. [Google Scholar] [CrossRef]
- Kramer, S.H.A.F. Further evidence for the division of attention among non-contiguous locations. Vis. Cogn. 1998, 5, 217–256. [Google Scholar] [CrossRef]
- Yamada, Y.; Kawahara, J.I. Dividing attention between two different categories and locations in rapid serial visual presentations. Percept. Psychophys. 2007, 69, 1218–1229. [Google Scholar] [CrossRef][Green Version]
- Rihs, T.A.; Micheal, C.M.; Thut, G. Mechanisms of selective inhibition in visual spatial attention are indexed by a-band EEG synchronization. Eur. J. Neurosci. 2007, 25, 603–610. [Google Scholar] [CrossRef]
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).