


The Role of Vegetation Monitoring in the Conservation of Coastal Habitats N2000: A Case Study of a Wetland Area in Southeast Sicily (Italy)



Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Phytosociological Data and Taxonomy
2.3. Statistical Analysis and Classification
2.4. Landscape and Diachronic Analysis
3. Results and Discussions
3.1. The Vascular Flora: Trait Analysis
3.2. Taxa of Relevant Interest
3.3. Plant Community Description
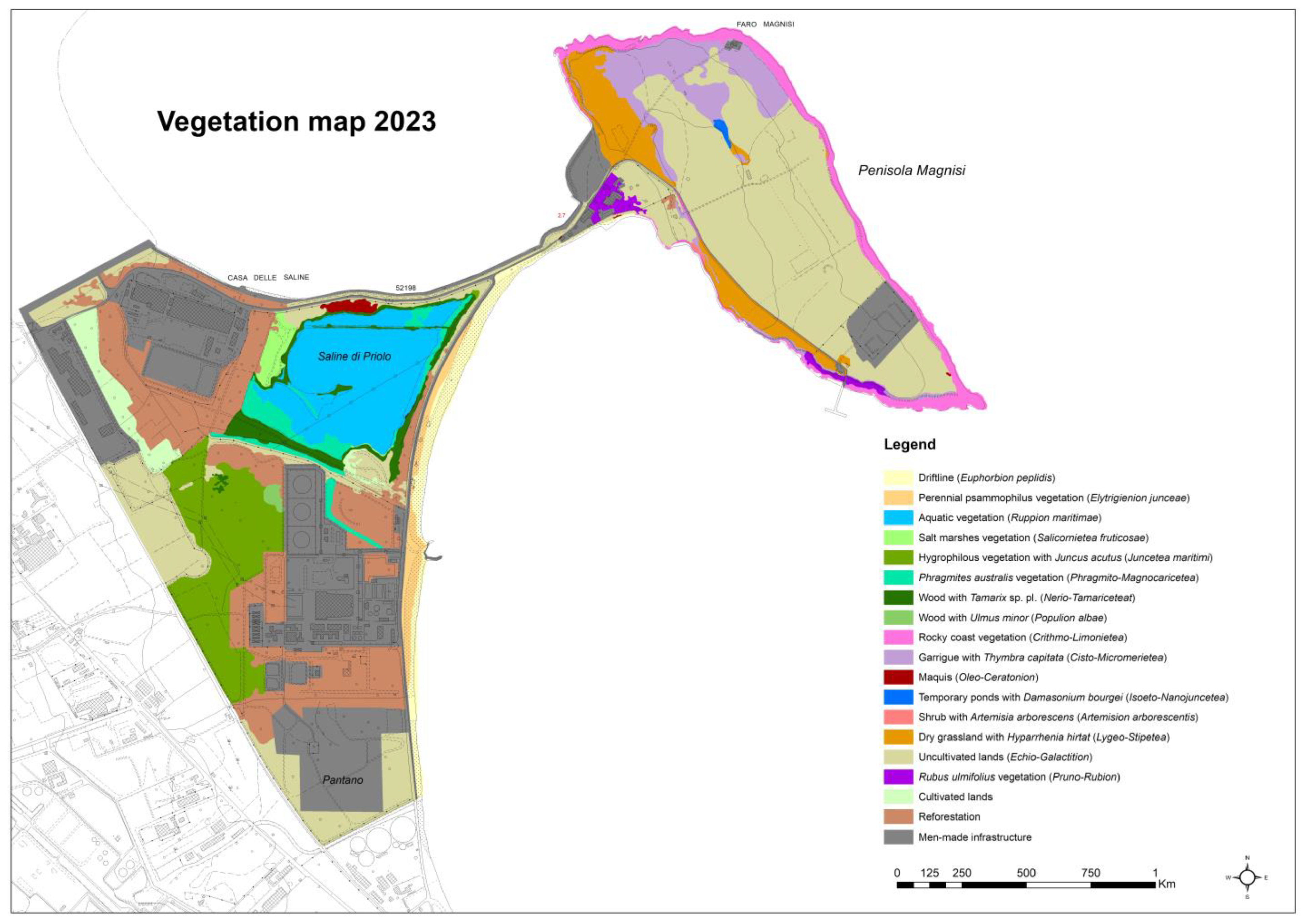
3.3.1. Coastal Dune Vegetation (Figure 2; Table S1)

3.3.2. Rocky Coast Vegetation (Figure 3 and Figure 4; Table S2)


3.3.3. Coastal Wetland Vegetation (Figure 5; Table S3)
Helophytic and Herbaceous Perennial Communities of Fresh and Brackish Waters

Temporary Ponds
3.3.4. Dry Grasslands and Garrigues/Shrubs (Figure 6 and Figure 7; Table S4)


3.4. Diacronic Analysis—Landscape Composition and Configuration
3.5. Conservation Status
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Species List of the Vascular Plants Recorded from the “Saline di Priolo” SAC (SE Sicily)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N. | Family | Corology | Life form | Species | Saline Priolo | Penisola Magnisi |
|---|---|---|---|---|---|---|
| 1 | Asteraceae | Medit.-Atl. | Ch | Achillea maritima (L.) Ehrend. and Y.P. Guo | 1 | |
| 2 | Amaranthaceae | SW-Medit. | Ch | Achyranthes sicula (L.) All. | 1 | 1 |
| 3 | Rosaceae | Eurasiat. | H | Agrimonia eupatoria L. | 1 | |
| 4 | Lamiaceae | Euri-Medit. | T | Ajuga chamaepitys (L.) Schreb. | 1 | 1 |
| 5 | Alismataceae | Subcosmop. | I | Alisma lanceolatum With. | 1 | |
| 6 | Amaryllidaceae | Europ. | G | Allium sphaerocephalon subsp. arvense (Guss.) Arcang. | 1 | |
| 7 | Amaryllidaceae | Steno-Medit. | G | Allium roseum L. | 1 | |
| 8 | Amaryllidaceae | Steno-Medit. | G | Allium commutatum Guss. | 1 | |
| 9 | Amaryllidaceae | Steno-Medit. | G | Allium subhirsutum L. | 1 | |
| 10 | Asteraceae | Steno-Medit. | T | Anacyclus clavatus (Desf.) Pers. | 1 | |
| 11 | Poaceae | Paleotrop. | H | Andropogon distachyos L. | 1 | |
| 12 | Apiaceae | Euri-Medit. | H | Anethum piperitum Ucria | 1 | 1 |
| 13 | Poaceae | Paleosubtrop. | T | Anisantha rigida (Roth) Hyl. | 1 | |
| 14 | Poaceae | Medit.-Turan. | T | Anisantha rubens (L.) Nevski | 1 | |
| 15 | Poaceae | Medit.-Turan. | T | Anisantha sterilis (L.) Nevski | 1 | |
| 16 | Poaceae | Paleotemp. | T | Anisantha tectorum (L.) Nevski | 1 | 1 |
| 17 | Asteraceae | Steno-Medit. | T | Anthemis arvensis L. | 1 | 1 |
| 18 | Apiaceae | Subcosmop. | T | Torilis arvensis (Huds.) Link | 1 | |
| 19 | Plantaginaceae | Endem. Ital. | Ch | Antirrhinum siculum Mill. | 1 | |
| 20 | Asteraceae | S-Medit. | NP | Artemisia arborescens (Vaill.) L. | 1 | |
| 21 | Asteraceae | Steno-Medit. | H | Carlina corymbosa L. | ||
| 22 | Lamiaceae | Steno-Medit. | T | Stachys major (L.) Bartolucci and Peru | ||
| 23 | Amaranthaceae | Medit. | Ch | Arthrocaulon meridionale Es.Ramírez, Rufo, Sánchez Mata, V.Fuente | 1 | 1 |
| 24 | Araceae | Steno-Medit. | G | Arum italicum Mill. | 1 | |
| 25 | Poaceae | Subcosmop. | G | Arundo donax L. | 1 | |
| 26 | Asparagaceae | Steno-Medit. | G | Asparagus acutifolius L. | 1 | 1 |
| 27 | Asphodelaceae | Subtrop. | H | Asphodelus fistulosus L. | 1 | 1 |
| 28 | Asphodelaceae | Steno-Medit. | G | Asphodelus ramosus L. | 1 | 1 |
| 29 | Asteraceae | Steno-Medit. | T | Asteriscus aquaticus (L.) Less. | 1 | |
| 30 | Fabaceae | S-Medit. | T | Astragalus epiglottis L. | 1 | |
| 31 | Fabaceae | S-Medit. | T | Astragalus boeticus L. | 1 | 1 |
| 32 | Fabaceae | Medit.-Turan. | T | Astragalus hamosus L. | 1 | |
| 33 | Fabaceae | Steno-Medit. | T | Astragalus pelecinus (L.) Barneby | 1 | |
| 34 | Amaranthaceae | Circumbor. | T | Atriplex prostrata Boucher ex DC. | 1 | |
| 35 | Poaceae | Medit.-Turan. | T | Avena barbata Pott ex Link | 1 | 1 |
| 36 | Poaceae | Medit.-Turan. | T | Avena sterilis L. | 1 | |
| 37 | Orobanchaceae | Euri-Medit. | T | Bellardia trixago (L.) All. | 1 | |
| 38 | Asparagaceae | Endem. Ital. | G | Bellevalia dubia (Guss.) Rchb. | 1 | |
| 39 | Asteraceae | Steno-Medit. | T | Bellis annua L. | 1 | |
| 40 | Amaranthaceae | Euri-Medit. | H | Beta vulgaris subsp. maritima (L.) Arcang. | 1 | 1 |
| 41 | Brassicaceae | Medit.-Turan. | T | Biscutella didyma L. | 1 | |
| 42 | Cyperaceae | Cosmop. | G | Bolboschoenus maritimus (L.) Palla | 1 | |
| 43 | Poaceae | Medit.-Turan. | T | Brachypodium distachyon (L.) P. Beauv. | 1 | 1 |
| 44 | Poaceae | Europ.-Caucas. | T | Bromus racemosus L. | 1 | |
| 45 | Crassulaceae | Subatl. | T | Bulliarda vaillantii (Willd.) DC. | 1 | |
| 46 | Apiaceae | Euri-Medit. | T | Bupleurum tenuissimum L. | 1 | |
| 47 | Brassicaceae | Medit.-Atl. | T | Cakile maritima Scop. | 1 | |
| 48 | Capparaceae | Steno-Medit. | NP | Capparis orientalis Veill. | 1 | 1 |
| 49 | Brassicaceae | Cosmop. | T | Cardamine hirsuta L. | 1 | |
| 50 | Asteraceae | Steno-Medit. | T | Carduus argyroa Biv. | 1 | |
| 51 | Asteraceae | Medit.-Turan. | H | Carduus pycnocephalus L. | 1 | |
| 52 | Cyperaceae | Steno-Medit. | G | Carex hispida Willd. ex Schkuhr | 1 | |
| 53 | Cyperaceae | Medit.-Atl. | H | Carex extensa Gooden. | 1 | |
| 54 | Cyperaceae | Atl. | H | Carex otrubae Podp. | 1 | |
| 55 | Cyperaceae | Eurosiber. | H | Carex vulpina L. | 1 | |
| 56 | Cyperaceae | Euri-Medit. | H | Carex distans L. | 1 | |
| 57 | Asteraceae | Endem. Ital. | H | Carlina hispanica Lam. | 1 | 1 |
| 58 | Asteraceae | Euri-Medit. | T | Carthamus lanatus L. | 1 | 1 |
| 59 | Poaceae | Medit.-Atl. | T | Catapodium balearicum (Willk.) H. Scholz | 1 | |
| 60 | Asteraceae | SW-Medit. | H | Centaurea sicula L. | 1 | |
| 61 | Asteraceae | Steno-Medit. | H | Centaurea sphaerocephala L. | 1 | |
| 62 | Gentianaceae | Paleotemp. | T | Centaurium tenuiflorum (Hoffmanns. and Link) Fritsch | 1 | |
| 63 | Characeae | I | Chara sp. | 1 | ||
| 64 | Asparagaceae | Steno-Medit. | G | Squilla maritima (L.) Steinh. | 1 | 1 |
| 65 | Asteraceae | Cosmop. | H | Cichorium intybus L. | 1 | |
| 66 | Asteraceae | Orof. NE-Medit. | H | Cirsium creticum subsp. triumfettii (Lacaita) K. Werner | 1 | |
| 67 | Boraginaceae | Euri-Medit. | H | Cynoglossum creticum Mill. | 1 | 1 |
| 68 | Lamiaceae | Medit.-Mont. | Ch | Clinopodium nepeta (L.) Kuntze | 1 | |
| 69 | Convolvulaceae | Steno-Medit. | H | Convolvulus althaeoides L. | 1 | 1 |
| 70 | Convolvulaceae | Cosmop. | G | Convolvulus soldanella L. | 1 | |
| 71 | Asteraceae | Medit. | H | Crepis bursifolia L. | 1 | |
| 72 | Convolvulaceae | Cosmop. | Ch | Cressa cretica L. | 1 | |
| 73 | Apiaceae | Euri-Medit. | Ch | Crithmum maritimum L. | 1 | 1 |
| 74 | Convolvulaceae | Eurasiat. | T | Cuscuta epithymum (L.) L. | 1 | |
| 75 | Poaceae | Steno-Medit. | T | Cutandia maritima (L.) Benth. ex Barbey | 1 | |
| 76 | Poaceae | Steno-Medit. | T | Cutandia divaricata (Desf.) Barbey | 1 | |
| 77 | Poaceae | Cosmop. | G | Cynodon dactylon (L.) Pers. | 1 | 1 |
| 78 | Cyperaceae | Steno-Medit. | G | Cyperus capitatus Vand. | 1 | |
| 79 | Poaceae | Steno-Medit. | H | Dactylis glomerata subsp. hispanica (Roth) Nyman | 1 | |
| 80 | Poaceae | Paleotrop. | T | Dactyloctenium aegyptium (L.) Willd. | 1 | |
| 81 | Alismataceae | Atl. | I | Damasonium bourgaei Coss. | 1 | |
| 82 | Apiaceae | Paleotemp. | H | Daucus carota L. | 1 | 1 |
| 83 | Apiaceae | Cosmop. | H | Daucus carota L. subsp. carota | 1 | |
| 84 | Apiaceae | W-Medit. | H | Daucus carota L. subsp. maritimus (Lam.) Batt. | 1 | |
| 85 | Brassicaceae | W-Medit. | T | Diplotaxis erucoides (L.) DC. | 1 | |
| 86 | Dipsacaceae | Euri-Medit. | H | Dipsacus fullonum L. | 1 | |
| 87 | Asteraceae | Euri-Medit. | H | Dittrichia viscosa (L.) Greuter | 1 | 1 |
| 88 | Apiaceae | Euri-Medit. | H | Echinophora spinosa L. | 1 | |
| 89 | Boraginaceae | Steno-Medit. | H | Echium arenarium Guss. | 1 | |
| 90 | Boraginaceae | Endem. Sic. | H | Echium italicum subsp. siculum (Lacaita) Greuter and Burdet | 1 | |
| 91 | Boraginaceae | Steno-Medit. | H | Echium parviflorum Moench | 1 | |
| 92 | Cyperaceae | Subcosmop. | G | Eleocharis palustris (L.) Roem. and Schult. | 1 | |
| 93 | Geraniaceae | Subcosmop. | T | Erodium cicutarium (L.) L’Hér. | 1 | |
| 94 | Geraniaceae | Steno-Medit. | T | Erodium laciniatum (Cav.) Willd. | 1 | |
| 95 | Apiaceae | Euri-Medit. | H | Eryngium campestre L. | 1 | |
| 96 | Apiaceae | SW-Medit. | H | Eryngium dicothomum Desf. | 1 | |
| 97 | Apiaceae | Medit.-Atl. | G | Eryngium maritimum L. | 1 | |
| 98 | Euphorbiaceae | Cosmop. | T | Euphorbia helioscopia L. | 1 | |
| 99 | Euphorbiaceae | Medit. | G | Euphorbia hirsuta L. | 1 | |
| 100 | Euphorbiaceae | Cosmop. | T | Euphorbia peplus L. | 1 | |
| 101 | Euphorbiaceae | W-Medit. | H | Euphorbia segetalis L. | 1 | |
| 102 | Euphorbiaceae | Steno-Medit. | H | Euphorbia terracina L. | 1 | |
| 103 | Apiaceae | Euri-Medit. | H | Ferula communis L. | 1 | |
| 104 | Poaceae | Subcosmop. | T | Festuca danthonii Asch. and Graebn. | 1 | 1 |
| 105 | Poaceae | Subcosmop. | T | Festuca myuros L. | 1 | |
| 106 | Poaceae | Medit.-Atl. | T | Festuca fasciculata Forssk. | 1 | |
| 107 | Moraceae | Medit.-Turan. | P | Ficus carica L. | 1 | 1 |
| 108 | Asteraceae | Steno-Medit. | T | Filago pygmaea L. | 1 | |
| 109 | Frankeniaceae | Steno-Medit. | Ch | Frankenia hirsuta L. | 1 | |
| 110 | Frankeniaceae | Steno-Medit. | T | Frankenia pulverulenta L. | 1 | |
| 111 | Asteraceae | Steno-Medit. | H | Galactites tomentosus Moench | 1 | 1 |
| 112 | Rubiaceae | Eurasiat. | T | Galium aparine L. | 1 | |
| 113 | Geraniaceae | Cosmop. | T | Geranium dissectum L. | 1 | |
| 114 | Geraniaceae | Paleotemp. | T | Geranium rotundifolium L. | 1 | 1 |
| 115 | Iridaceae | Euri-Medit. | G | Gladiolus italicus Mill. | 1 | 1 |
| 116 | Papaveraceae | Euri-Medit. | H | Glaucium flavum Crantz | 1 | |
| 117 | Asteraceae | Steno-Medit. | T | Glebionis coronaria (L.) Spach | ||
| 118 | Amaranthaceae | Circumbor. | Ch | Halimione portulacoides (L.) Aellen | 1 | |
| 119 | Asteraceae | Steno-Medit. | T | Hedypnois rhagadioloides (L.) F.W. Schmidt | 1 | 1 |
| 120 | Asteraceae | Euri-Medit. | T | Helminthotheca echioides (L.) Holub | 1 | |
| 121 | Brassicaceae | Subatl. | H | Hirschfeldia incana (L.) Lagr.-Foss. | 1 | 1 |
| 122 | Poaceae | Circumbor. | H | Holcus lanatus L. | 1 | |
| 123 | Poaceae | Euri-Medit. | T | Hordeum murinum subsp. leporinum (Link) Arcang. | 1 | 1 |
| 124 | Asteraceae | Steno-Medit. | H | Hyoseris radiata L. | 1 | |
| 125 | Poaceae | Paleotrop. | H | Hyparrhenia hirta (L.) Stapf | 1 | |
| 126 | Hypericaceae | Steno-Medit. | H | Hypericum triquetrifolium Turra | 1 | |
| 127 | Asteraceae | Steno-Medit. | T | Hypochaeris achyrophorus L. | 1 | 1 |
| 128 | Cyperaceae | Subcosmop. | H | Isolepis cernua (Vahl) Roem. and Schult. | 1 | |
| 129 | Juncaceae | Euri-Medit. | H | Juncus acutus L. | 1 | 1 |
| 130 | Juncaceae | Circumbor. | G | Juncus articulatus L. | 1 | |
| 131 | Juncaceae | Cosmop. | T | Juncus bufonius L. | 1 | |
| 132 | Juncaceae | Paleosubtrop. | G | Juncus fontanesii J.Gay | 1 | |
| 133 | Juncaceae | Euri-Medit. | T | Juncus hybridus Brot. | 1 | 1 |
| 134 | Juncaceae | Subcosmop. | G | Juncus maritimus Lam. | 1 | |
| 135 | Juncaceae | S-Medit. | G | Juncus subulatus Forssk. | 1 | |
| 136 | Plantaginaceae | Steno-Medit. | H | Kickxia commutata (Bernh. ex Rchb.) Fritsch | 1 | |
| 137 | Poaceae | Euri-Medit. | T | Lagurus ovatus L. | 1 | |
| 138 | Fabaceae | Euri-Medit. | T | Lathyrus annuus L. | 1 | |
| 139 | Fabaceae | Euri-Medit. | T | Lathyrus aphaca L. | 1 | 1 |
| 140 | Fabaceae | Euri-Medit. | T | Lathyrus cicera L. | 1 | |
| 141 | Fabaceae | Steno-Medit. | T | Lathyrus clymenum L. | 1 | |
| 142 | Fabaceae | Steno-Medit. | T | Lathyrus ochrus (L.) DC. | 1 | 1 |
| 143 | Malvaceae | Steno-Medit. | H | Malva arborea (L.) Webb and Berthel. | ||
| 144 | Brassicaceae | Euri-Medit. | T | Lepidium coronopus (L.) Al-Shehbaz | 1 | |
| 145 | Asteraceae | Medit.Atl. | Ch | Limbarda crithmoides (L.) Dumort. | 1 | 1 |
| 146 | Plumbacinaceae | Euri-Medit. | H | Limonium narbonense Mill. | 1 | |
| 147 | Plumbaginaceae | Endem. Sic. | Ch | Limonium syracusanum Brullo | 1 | 1 |
| 148 | Plantaginaceae | W-Medit. | T | Linaria triphylla (L.) Mill. | 1 | |
| 149 | Linaceae | Euri-Medit. | H | Linum usitatissimum subsp. angustifolium (Huds.) Thell. | 1 | |
| 150 | Brassicaceae | Steno-Medit. | Ch | Lobularia maritima (L.) Desv. | 1 | 1 |
| 151 | Poaceae | Paleotemp. | H | Lolium arundinaceum (Schreb.) Darbysh. | 1 | |
| 152 | Fabaceae | Euri-Medit. | H | Lotus corniculatus L. subsp. preslii (Ten.) P.Fourn. | 1 | |
| 153 | Fabaceae | Steno-Medit. | Ch | Lotus cytisoides L. | 1 | 1 |
| 154 | Fabaceae | Steno-Medit. | T | Lotus edulis L. | 1 | |
| 155 | Fabaceae | Steno-Medit. | T | Lotus ornithopodioides L. | 1 | |
| 156 | Primulaceae | Cosmop. | T | Lysimachia arvensis (L.) U. Manns and Anderb. | 1 | 1 |
| 157 | Lythraceae | Subcosmop. | T | Lythrum hyssopifolia L. | 1 | |
| 158 | Lythraceae | Steno-Medit. | H | Lythrum junceum Banks and Sol. | 1 | |
| 159 | Malvaceae | Subcosmop. | H | Malva sylvestris L. | 1 | |
| 160 | Solanaceae | Steno-Medit. | H | Mandragora autumnalis Bertol. | 1 | |
| 161 | Lamiaceae | Euri-Medit. | H | Marrubium vulgare L. | 1 | 1 |
| 162 | Brassicaceae | Steno-Medit. | T | Matthiola tricuspidata (L.) W.T. Aiton | 1 | 1 |
| 163 | Fabaceae | Euri-Medit. | T | Medicago littoralis Rohde ex Loisel. | 1 | 1 |
| 164 | Fabaceae | Euri-Medit. | Ch | Medicago marina L. | 1 | |
| 165 | Fabaceae | Euri-Medit. | T | Medicago minima (L.) L. | 1 | 1 |
| 166 | Fabaceae | Euri-Medit. | T | Medicago polymorpha L. | 1 | |
| 167 | Fabaceae | Steno-Medit. | T | Medicago truncatula Gaertn. | 1 | |
| 168 | Lamiaceae | Euri-Medit. | H | Mentha pulegium L. | 1 | 1 |
| 169 | Euphorbiaceae | Paleotemp. | T | Mercurialis annua L. | 1 | 1 |
| 170 | Aizoaceae | S-Medit. | T | Mesembryanthemum nodiflorum L. | 1 | |
| 171 | Lamiaceae | Endem. Ital. | Ch | Micromeria graeca subsp. tenuifolia (Ten.) Nyman | 1 | 1 |
| 172 | Lamiaceae | S-Medit. | Ch | Micromeria nervosa (Desf.) Benth. | 1 | |
| 173 | Plantaginaceae | Euri-Medit. | T | Misopates orontium (L.) Raf. | 1 | |
| 174 | Iridaceae | Steno-Medit. | G | Moraea sisyrinchium (L.) Ker Gawl. | 1 | |
| 175 | Asparagaceae | Euri-Medit. | G | Muscari comosum (L.) Mill. | 1 | |
| 176 | Asparagaceae | Steno-Medit. | G | Muscari parviflorum Desf. | 1 | |
| 177 | Myrtaceae | Steno-Medit. | P | Myrtus communis L. | 1 | |
| 178 | Ranunculaceae | Euri-Medit. | T | Nigella damascena L. | 1 | |
| 179 | Asteraceae | Steno-Medit. | T | Notobasis syriaca (L.) Cass. | 1 | |
| 180 | Apiaceae | Steno-Medit. | H | Oenanthe globulosa L. | 1 | |
| 181 | Oleaceae | Steno-Medit. | P | Olea europaea L. var. sylvestris (Mill.) Lehr | 1 | 1 |
| 182 | Poaceae | Medit.-Turan. | H | Oloptum miliaceum (L.) Röser and H.R. Hamasha | 1 | |
| 183 | Fabaceae | Steno-Medit. | T | Onobrychis caput-galli (L.) Lam. | 1 | 1 |
| 184 | Fabaceae | Euri-Medit. | H | Ononis natrix subsp. ramosissima (Desf.) Batt. | 1 | 1 |
| 185 | Fabaceae | Medit.-Turan. | T | Ononis reclinata L. | 1 | 1 |
| 186 | Fabaceae | Steno-Medit. | T | Ononis variegata L. | 1 | |
| 187 | Asteraceae | Steno-Medit. | H | Onopordum illyricum L. | 1 | |
| 188 | Orchidaceae | Steno-Medit | G | Ophrys bertolonii Moretti | 1 | |
| 189 | Cactaceae | Americ. | P | Opuntia dillenii (Ker Gawl.) Haw. | 1 | |
| 190 | Cactaceae | Neotrop. | P | Opuntia ficus-indica (L.) Mill. | 1 | |
| 191 | Asparagaceae | Steno-Medit. | G | Ornithogalum gussonei Ten. | 1 | |
| 192 | Oxalidaceae | Africana | G | Oxalis pes-caprae L. | 1 | 1 |
| 193 | Asteraceae | Euri-Medit. | H | Pallenis spinosa (L.) Cass. | 1 | |
| 194 | Amaryllidaceae | Steno-Medit. | G | Pancratium maritimum L. | 1 | |
| 195 | Poaceae | Euri-Medit. | T | Parapholis cylindrica (Willd.) Romero Zarco | 1 | |
| 196 | Poaceae | Medit.-Atl. | T | Parapholis filiformis (Roth) C.E. Hubb. | 1 | |
| 197 | Poaceae | Medit.-Atl. | T | Parapholis incurva (L.) C.E. Hubb. | 1 | 1 |
| 198 | Urticaceae | Euri-Medit. | H | Parietaria judaica L. | 1 | |
| 199 | Asteraceae | Steno-Medit. | Ch | Phagnalon saxatile (L.) Cass. | 1 | |
| 200 | Poaceae | Steno-Medit. | H | Phalaris coerulescens Desf. | 1 | |
| 201 | Poaceae | Subcosmop. | G | Phragmites australis (Cav.) Trin. ex Steud. | 1 | |
| 202 | Verbenaceae | Pantrop. | H | Phyla nodiflora (L.) Greene | 1 | |
| 203 | Anacardiaceae | S-Medit. | P | Pistacia lentiscus L. | 1 | |
| 204 | Plantaginaceae | Euri-Medit. | H | Plantago coronopus L. | 1 | |
| 205 | Plantaginaceae | Steno-Medit. | T | Plantago lagopus L. | 1 | 1 |
| 206 | Plantaginaceae | Steno-Medit. | H | Plantago macrorhiza Poir. | 1 | |
| 207 | Plantaginaceae | Eurasiat. | H | Plantago media L. | 1 | |
| 208 | Plantaginaceae | Steno-Medit. | H | Plantago serraria L. | 1 | |
| 209 | Poaceae | Euri-Medit. | T | Poa infirma Kunth | 1 | 1 |
| 210 | Poaceae | Paleosubtrop. | T | Polypogon monspeliensis (L.) Desf. | 1 | |
| 211 | Rosaceae | Paleotemp. | H | Potentilla reptans L. | 1 | |
| 212 | Rosaceae | Steno-Medit. | NP | Poterium spinosum L. | 1 | |
| 213 | Asparagaceae | Steno-Medit. | G | Prospero autumnale (L.) Speta | 1 | |
| 214 | Asteraceae | Steno-Medit. | T | Pulicaria sicula (L.) Moris | 1 | |
| 215 | Asteraceae | Steno-Medit. | H | Pulicaria dysenterica (L.) Bernh. | 1 | |
| 216 | Rosaceae | Eurasiat. | P | Pyrus spinosa Forssk. | 1 | |
| 217 | Ranunculaceae | W-Medit. | T | Ranunculus trilobus Desf. | 1 | |
| 218 | Asteraceae | Steno-Medit. | H | Reichardia picroides (L.) Roth | 1 | |
| 219 | Resedaceae | Steno-Medit. | H | Reseda alba L. | 1 | |
| 220 | Rhamnaceae | Steno-Medit. | P | Rhamnus alaternus L. | 1 | 1 |
| 221 | Iridaceae | Steno-Medit. | G | Romulea ramiflora Ten. | 1 | |
| 222 | Rosaceae | Steno-Medit. | NP | Rosa sempervirens L. | 1 | |
| 223 | Poaceae | Paleotemp. | T | Rostraria cristata (L.) Tzvelev | 1 | |
| 224 | Rubiaceae | Steno-Medit. | P | Rubia peregrina L. | 1 | |
| 225 | Rosaceae | Euri-Medit. | NP | Rubus ulmifolius Schott | 1 | |
| 226 | Polygonaceae | Euri-Medit. | H | Rumex pulcher L. | 1 | 1 |
| 227 | Polygonaceae | Eurasiat. | H | Rumex conglomeratus Murray | ||
| 228 | Polygonaceae | Subcosmop. | H | Rumex crispus L. | 1 | |
| 229 | Polygonaceae | W-Medit. | H | Rumex thyrsoides Desf. | 1 | |
| 230 | Ruppiaceae | Cosmop. | I | Ruppia maritima L. | 1 | |
| 231 | Amaranthaceae | Euri-Medit. | Ch | Salicornia fruticosa (L.) L. | ||
| 232 | Amaranthaceae | W-Europ. | T | Salicornia perennans Willd. | 1 | |
| 233 | Amaranthaceae | Steno-Medit. | Ch | Salicornia perennis subsp. alpini (Lag.) Castrov. | 1 | |
| 234 | Salicaceae | Steno-Medit. | P | Salix pedicellata Desf. | 1 | |
| 235 | Amaranthaceae | Paleotemp. | T | Salsola tragus L. | 1 | |
| 236 | Lamiaceae | Euri-Medit. | H | Salvia verbenaca L. | 1 | |
| 237 | Gentianaceae | Euri-Medit. | T | Schenkia spicata (L.) G. Mans. | 1 | |
| 238 | Cyperaceae | Euri-Medit. | G | Scirpoides holoschoenus (L.) Soják | 1 | |
| 239 | Asteraceae | SW-Medit. | H | Scolymus grandiflorus Desf. | 1 | |
| 240 | Asteraceae | Euri-Medit. | H | Scolymus hispanicus L. | 1 | |
| 241 | Fabaceae | S-Medit. | T | Scorpiurus vermiculatus L. | 1 | |
| 242 | Crassulaceae | SW-Medit. | T | Sedum caeruleum L. | 1 | 1 |
| 243 | Asteraceae | Cosmop. | T | Senecio vulgaris L. | 1 | |
| 244 | Caryophyllaceae | S-Medit. | T | Silene bellidifolia Jacq. | 1 | |
| 245 | Caryophyllaceae | Steno-Medit. | T | Silene colorata Poir. | 1 | 1 |
| 246 | Caryophyllaceae | Steno-Medit. | T | Silene niceensis All. | 1 | |
| 247 | Caryophyllaceae | Steno-Medit. | T | Silene sedoides Poir. | 1 | |
| 248 | Caryophyllaceae | Paleotemp. | H | Silene vulgaris (Moench) Garcke | 1 | |
| 249 | Asteraceae | Medit.-Turan. | H | Silybum marianum (L.) Gaertn. | 1 | 1 |
| 250 | Brassicaceae | E-Medit. | T | Sinapis alba L. | 1 | |
| 251 | Dipsacaceae | Steno-Medit. | H | Sixalix atropurpurea (L.) Greuter and Burdet | 1 | 1 |
| 252 | Apiaceae | Medit.-Atl. | H | Smyrnium olusatrum L. | 1 | |
| 253 | Asteraceae | Eurasiat. | H | Sonchus asper (L.) Hill | 1 | |
| 254 | Asteraceae | Steno-Medit. | G | Sonchus bulbosus (L.) Kilian and Greuter | 1 | |
| 255 | Asteraceae | Cosmop. | H | Sonchus oleraceus L. | 1 | 1 |
| 256 | Asteraceae | Steno-Medit. | H | Sonchus tenerrimus L. | 1 | |
| 257 | Caryophyllaceae | Subcosmop. | T | Spergularia marina (L.) Besser | 1 | 1 |
| 258 | Poaceae | Subtrop. | G | Sporobolus virginicus (L.) Kunth | 1 | |
| 259 | Lamiaceae | Steno-Medit. | Ch | Stachys major (L.) Bartolucci and Peruzzi | 1 | |
| 260 | Lamiaceae | Steno-Medit. | T | Stachys romana (L.) E.H.L. Krause | 1 | 1 |
| 261 | Poaceae | Steno-Medit. | T | Stipellula capensis (Thunb.) Röser and H.R. Hamasha | 1 | 1 |
| 262 | Amaranthaceae | Cosmop. | T | Suaeda maritima (L.) Dumort. | 1 | |
| 263 | Amaranthaceae | Cosmop. | NP | Suaeda vera J. F. Gmelin | 1 | 1 |
| 264 | Asteraceae | Neotrop. | H | Symphyotrichum squamatum (Spreng.) G. L. Nesom | 1 | |
| 265 | Tamaricaceae | W-Medit. | P | Tamarix africana Poir. | 1 | 1 |
| 266 | Tamaricaceae | S-Medit. | P | Tamarix arborea (Ehrenb.) Bunge | 1 | |
| 267 | Tamaricaceae | W-Medit. | P | Tamarix gallica L. | 1 | |
| 268 | Lamiaceae | Europ.-Caucas. | H | Teucrium scordium L. | 1 | |
| 269 | Apiaceae | S-Medit. | H | Thapsia garganica L. | 1 | |
| 270 | Poaceae | Submedit. | H | Thinopyrum flaccidifolium (Boiss. and Heldr.) Moustakas | 1 | |
| 271 | Poaceae | Euri-Medit. | G | Thinopyrum junceum (L.) Á. Löve | 1 | |
| 272 | Lamiaceae | Steno-Medit. | Ch | Thymbra capitata (L.) Cav. | 1 | |
| 273 | Thymelaeaceae | S-Medit. | Ch | Thymelaea hirsuta (L.) Endl. | 1 | |
| 274 | Apiaceae | Steno-Medit. | T | Tordylium apulum L. | 1 | |
| 275 | Apiaceae | Subcosmop. | T | Torilis arvensis (Huds.) Link | 1 | |
| 276 | Asteraceae | Euri-Medit. | H | Tragopogon porrifolius L. | 1 | |
| 277 | Fabaceae | Paleotemp. | T | Trifolium campestre Schreb. | 1 | 1 |
| 278 | Fabaceae | Euri-Medit. | T | Trifolium cherleri L. | 1 | 1 |
| 279 | Fabaceae | Paleotemp. | H | Trifolium fragiferum L. | 1 | |
| 280 | Fabaceae | Euri-Medit. | T | Trifolium nigrescens Viv. | 1 | 1 |
| 281 | Fabaceae | Paleotemp. | H | Trifolium resupinatum L. | 1 | |
| 282 | Fabaceae | Euri-Medit. | T | Trifolium scabrum L. | 1 | 1 |
| 283 | Fabaceae | Euri-Medit. | T | Trifolium stellatum L. | 1 | |
| 284 | Fabaceae | Paleotemp. | T | Trifolium tomentosum L. | 1 | |
| 285 | Fabaceae | S-Medit. | T | Trigonella sulcata (Desf.) Coulot and Rabaute | 1 | |
| 286 | Poaceae | Steno-Medit. | T | Trisetaria aurea (Ten.) Banfi and Galasso | 1 | |
| 287 | Poaceae | Medit.-Turan. | T | Triticum vagans (Jord. and Fourr.) Greuter | 1 | |
| 288 | Typhaceae | Pantrop. | G | Typha domingensis (Pers.) Steud. | 1 | |
| 289 | Typhaceae | Circumbor. | G | Typha angustifolia L. | 1 | |
| 290 | Ulmaceae | Europ.-Caucas. | P | Ulmus minor Mill. | 1 | |
| 291 | Asteraceae | Euri-Medit. | T | Urospermum picroides (L.) Scop. ex F.W. Schmidt | 1 | 1 |
| 292 | Urticaceae | S-Medit. | T | Urtica membranacea Poir. | 1 | |
| 293 | Asteraceae | Neotrop. | T | Symphyotrichum squamatum (Spreng.) G.L. Nesom | ||
| 294 | Rubiaceae | Steno-Medit. | T | Valantia muralis L. | 1 | |
| 295 | Scrophulariaceae | Euri-Medit. | H | Verbascum sinuatum L. | 1 | 1 |
| 296 | Plantaginaceae | Cosmop. | H | Veronica anagallis-aquatica L. | 1 | |
| 297 | Fabaceae | Euri-Medit. | T | Vicia hybrida L. | 1 | |
| 298 | Fabaceae | S-Europ. | T | Vicia melanops Sm. | 1 | 1 |
| 299 | Fabaceae | Steno-Medit. | T | Vicia sativa L. | 1 | |
| 300 | Fabaceae | Steno-Medit. | H | Vicia villosa Roth | 1 | 1 |
| 301 | Fabaceae | Euri-Medit. | T | Ervum gracile DC. (=Vicia tenuissima (Bieb.) Sch. and Th.) | 1 | 1 |
| 302 | Lamiaceae | Medit.-Turan. | P | Vitex agnus-castus L. | 1 | |
| 303 | Asteraceae | S-Europ. | T | Xanthium italicum Moretti | 1 | |
| 304 | Rhamnaceae | S-Medit. | P | Ziziphus lotus (L.) Lam. | 1 |
Appendix B. Syntaxonomical Scheme of the Vegetation Units Recorded from the “Saline di Priolo” SAC (SE Sicily)
| CAKILETEA MARITIMAE Tüxen and Preising ex Br.-Bl. and Tüxen 1952 |
| EUPHORBIETALIA PEPLIS Tüxen 1950 |
| EUPHORBION PEPLIS Tüxen 1950 |
| 1. Salsolo-Cakiletum maritimae Costa and Mansanet 1981 corr. Rivas-Martínez et al., 1992 |
| EUPHORBIO PARALIAE-AMMOPHILETEA AUSTRALIS Géhu and Rivas-Martínez in Rivas |
| Martínez, Asensi, Díez-Garretas, Molero, Valle, Cano, Costa and Díaz 2011 |
| AMMOPHILETALIA AUSTRALIS Br.-Bl. 1933 |
| AMMOPHILION AUSTRALIS Br.-Bl. 1921 corr. Rivas-Martìnez, Costa Izco in Rivas-Martìnez, Lousa, T. |
| E.Diaz, Fernandez-Gonzalez and J.C.Costa 1990 |
| 2. Cypero capitati-Agropyretum juncei Kühnholtz-Lordat (1923) Br.-Bl. 1933 |
| ONONIDION RAMOSISSIMAE Pignatti 1952 |
| 3. Centaureo sphaerocephalae-Ononidetum ramosissimae Br.-Bl. e Frei in Frei, 193 |
| HELIANTHEMETEA GUTTATI (Br.-Bl. in Br.-Bl., Roussine and Nègre 1952) Rivas Goday and Rivas-Martìnez 1963 em. Rivas-Martìnez 1978 |
| CUTANDIETALIA MARITIMAE Rivas- Martìnez, Díez-Garretas, and Asensi 2002 |
| ALKANNO-MARESION NANAE Rivas Goday ex Rivas Goday and Rivas-Martínez 1963 corr. Diaz-Garretas et al., 2001 |
| 4. Sileno coloratae-Ononidetum variegatae Gèhu and Gèhu-Franck 1986 |
| RUPPIETEA MARITIMAE Tuxen ex Den Hartog and Segal 1964 |
| RUPPIETALIA MARITIMAE Tuxen ex Den Hartog and Segal 1964 |
| RUPPION MARITIMAE Br.-Bl. ex Br.-Bl., Roussine and Nègre 1952 |
| 5. Enteromorpho intestinalidis-Ruppietum maritimae Westhoff ex R.Tx. and Böckelmann 1957 |
| PHRAGMITO-MAGNOCARICETEA Klika in Klika and Novák 1941 |
| PHRAGMITETALIA Koch 1926 |
| PHRAGMITION Koch 1926 |
| 6. Phragmitetum communis (Koch 1926) Schmale 1939 |
| 7. Typhetum domingensis Brullo, Minissale and Spamp. 1994 |
| NASTURTIO-GLYCERIETALIA Pign. 1954 |
| GLYCERIO-SPARGANION Br.-Bl. and Sissing in Boer 1942 |
| 8. Eleocharido-Alismetum lanceolati Minissale and Spampinato 1987 |
| 9. Bolboschoeno maritimi-Alismetum lanceolati ass. nov. hoc loco |
| MOLINIO-ARRHENATHERETEA R.Tx.1937 |
| PASPALO-AGROSTION SEMIVERTICILLATI Br.-Bl. in Br.-Bl. Roussine and Negre 1952 |
| PASPALO-HELEOCHLOETALIA Br.-Bl. ex Rivas Goday 1956 |
| 10. Euphorbio hirsutae-Lotetum preslii ass. nov. hoc loco |
| SALICORNIETEA FRUTICOSAE Br.-Bl. et Tx. ex A. Bolòs y Vayreda et. O. de Bolòs in A. Bolòs et Vayreda 1950 |
| SALICORNIETALIA FRUTICOSAE Br.-Bl. 1933 |
| SALICORNION FRUTICOSAE Br.-Bl. 1933 |
| 11. Junco subulati-Sarcocornietum alpini Brullo et Sciandrello in Giusso et al., 2008 |
| ARTHROCNEMION GLAUCI Rivas-Mart. et Costa M. 1984 |
| 12. Arthrocaulo meridionalis-Juncetum subulati Brullo et Furnari 1976 nom. corr. Sciandrello et al., 2019 |
| 13. Limonio virgati-Arthrocnemetum macrostachyi Biondi, Casavecchia and Guerra 2006 |
| SUAEDION BREVIFOLIAE Br.-Bl. et O. de Bolòs 1958 (= Suaedion verae Brullo et Furnari 1988) |
| 14. Halimiono-Suaedetum verae Molinier et Tallon 1970 corr. Géhu 1984 |
| INULION CRITHMOIDIS Brullo et Furnari 1988 |
| 15. Agropyro scirpei-Inuletum crithmoidis Brullo in Brullo et al., 1988 |
| THERO-SUAEDETEA SPLENDENTIS Rivas-Martínez 1972 |
| THERO-SALICORNIETALIA Tüxen in Tüxen et Oberdorfer ex Géhu et Géhu-Franck 1984 |
| Salicornion patulae Géhu et Géhu-Franck ex Rivas-Martínez 1990 |
| 16. Suaedo-Salicornietum patulae Brullo et Furnari ex Géhu et Géhu-Franck 1984 |
| JUNCETEA MARITIMI Br.Bl. in Br.-Bl., Roussine and Nègre 1952 |
| JUNCETALIA MARITIMI Br.Bl. ex Horvatic 1934 |
| JUNCION MARITIMI Br.Bl. ex Horvatic 1934 |
| 17. Juncetum maritimo-acuti Horvatic 1934 (Juncus acutus comm.) |
| HALO-ARTEMISION COERULESCENTIS Pignatti 1953 |
| 18. Elymetum atherici Pellizzari, Merloni et Piccoli 1998 (=Thinopyrum acutum (DC.) Banfi) |
| SAGINETEA MARITIMAE Westhoff, Van Leeuwen et Adriani 1962 |
| SAGINETALIA MARITIMAE Westhoff, Van Leeuwen et Adriani 1962 |
| SILENO SEDOIDIS-CATAPODION BALEARICI de Foucault and Bioret 2010 corr. Tomaselli et al., 2020 |
| 19. Parapholido incurvae-Spergularietum marinae ass. nov. hoc loco |
| ISOËTO-NANOJUNCETEA Br.-Bl. and R. Tx. ex Westhoff, Dijk and Passchier 1946 |
| Isoëtetalia Br.-Bl. 1936 |
| Isoëtion Br.-Bl. 1936 |
| 20. Lythro hyssopifoliae-Crassuletum vaillantii Bagella et al., 2009 |
| NERIO-TAMARICETEA Br.-Bl. et O.Bolòs 1958 |
| TAMARICETALIA AFRICANAE Br.-Bl. et O. Bolòs 1958 |
| TAMARICION AFRICANAE Br.-Bl. et O.Bolòs 1958 |
| 21. Inulo crithmoidis-Tamaricetum africanae Gamisans 1992 (=Limbardo crithmoidis-Tamaricetum africanae Sciandrello et al., 2019) |
| CRITHMO-LIMONIETEA Br.-Bl. in Br- Bl., Roussine and Nègre1952 |
| CRITHMO-LIMONIETALIA Molinier 1934 |
| CRITHMO-LIMONION MOLINIER 1934 |
| 22. Limonietum syracusani Bartolo, Brullo and Marcenò 1982 |
| PEGANO HARMALAE-SALSOLETEA VERMICULATAE Br-Bl and O.Bolòs 1958 |
| SALSOLO VERMICULATAE-PEGANETALIA HARMALAE Br.-Bl. and O. Bolòs 1954 |
| Artemision arborescentis Géhu et al., 1986 |
| 23. Atriplici halimi-Artemisietum arborescentis Biondi 1988 (Artemisia arborescens comm.) |
| CISTO-MICROMERIETEA Oberd. 1954 |
| CISTO-ERICETALIA Horvatic 1958 |
| CISTO-ERICION Horvatic 1958 |
| 24. Thymbra capitata comm. |
| LYGEO SPARTI-STIPETEA TENACISSIMAE Rivas-Martinez 1978 |
| CYMBOPOGONO-BRACHYPODIETALIA RAMOSI Horvatić 1963 |
| HYPARRHENENION HIRTAE Brullo, Minissale and Spamp. in C. Brullo et al., 2010 |
| 25. Hyparrhenietum hirto-pubescentis A.and O. Bolòs and Br.-Bl. in A.and O. Bolòs 1950 |
| BROMO-ORYZOPSION MILIACEAE O.Bolòs 1970 |
| 26. Oloptum miliaceum comm. |
| STIPO-TRACHYNIETEA DISTACHYAE Brullo in Brullo, Scelsi and Spampinato 2001 |
| TRACHYNIETALIA DISTACHYAE Rivas-Martínez 1978 |
| TRACHYNION DISTACHYAE Rivas-Martínez 1978 |
| 27. Thero-Sedetum caerulei Brullo 1975 |
| CHENOPODIETEA Br.-Bl. in Br.-Bl. et al., 1952 |
| BROMETALIA RUBENTI-TECTORUM (Rivas Goday et Rivas-Martinez 1973) Rivas-Martinez and Izco 1977 |
| ECHIO PLANTAGINEI-GALACTITION TOMENTOSAE O. Bolòs and Molinier 1969 |
| 28. Stipellula capensis comm. |
Appendix C. Localities and Dates of Phytosociological Relevés
References
- Leberger, R.; Geijzendorffer, I.R.; Gaget, E.; Gwelmami, A.; Galewski, T.; Pereira, H.M.; Guerra, C.A. Mediterranean wetland conservation in the context of climate and land cover change. Reg. Environ. Chang. 2020, 20, 67. [Google Scholar] [CrossRef]
- Taylor, N.G.; Grillas, P.; Al Hreisha, H.; Balkız, Ö.; Borie, M.; Boutron, O.; Catita, A.; Champagnon, J.; Cherif, S.; Çiçek, K.; et al. The future for Mediterranean wetlands: 50 key issues and 50 important conservation research questions. Reg. Environ. Chang. 2021, 21, 33. [Google Scholar] [CrossRef] [PubMed]
- Scott, D.B.; Frail-Gauthier, J.; Mudie, P.J. Coastal Wetlands of the World: Geology, Ecology, Distribution and Applications; Cambridge University Press: New York, NY, USA, 2014. [Google Scholar]
- Garbutt, A.; De Groot, A.; Smit, C.; Pétillon, J. European salt marshes: Ecology and conservation in a changing world. J. Coast. Conserv. 2017, 21, 405–408. [Google Scholar] [CrossRef]
- Sciandrello, S.; Musarella, C.M.; Puglisi, M.; Spampinato, G.; Tomaselli, V.; Minissale, P. Updated and new insights on the coastal halophilous vegetation of southeastern Sicily (Italy). Plant Sociol. 2019, 56, 81–98. [Google Scholar]
- De Pietro, R.; Ientile, R.; Puccia, S.; Sabella, G. Birds of Gelsari and Lentini marshes, special protection area for the protection and maintenance of aquatic avifauna in central Mediterranean. Ocean Coast. Manag. 2019, 169, 96–103. [Google Scholar] [CrossRef]
- Frei, M. Studi fitosociologici su alcune associazioni litorali in Sicilia (Ammophiletalia e Salicornietalia). Nuovo G. Bot. Ital. 1937, 44, 273–294. [Google Scholar] [CrossRef]
- Brullo, S.; Di Martino, A. Vegetazione dell’Isola Grande dello Stagnone (Marsala). Boll. Studi Inf. Giard. Colon. Palermo 1974, 26, 15–62. [Google Scholar]
- Brullo, S.; Furnari, F. Le associazioni vegetali degli ambienti palustri costieri della Sicilia. Not. Soc. Ital. Fitosociol. 1976, 11, 1–43. [Google Scholar]
- Brullo, S.; Furnari, F. La vegetazione costiera della Cirenaica. Boll. Dell’accademia Gioenia Sci. Nat. 1988, 21, 31–117. [Google Scholar]
- Minissale, P.; Sciandrello, S. Flora e vegetazione terrestre della Riserva Naturale di Vendicari (Sicilia sud-orientale). Ente Fauna Sicil. 2010, 12, 145–208. [Google Scholar]
- Sciandrello, S. Coastal saltmarsh vegetation in Sicily (Italy): Phytosociological insights and plant diversity. Plant Biosyst. 2020, 154, 860–876. [Google Scholar] [CrossRef]
- Benadusi, M. Oil in Sicily: Petrocapitalist imaginaries in the shadow of old smokestacks. Econ. Anthropol. 2018, 5, 45–58. [Google Scholar] [CrossRef]
- Brullo, S.; Furnari, F. Vegetazione dei Pantani Litoranei della Sicilia Sud-Orientale e Problema della Conservazione Dell’ambiente; Istituto di Botanica dell’Università di Catania: Catania, Italy, 1971. [Google Scholar]
- Fagotto, F.; Baglieri, S. Ornitofauna e vegetazione delle saline di Siracusa (un luogo umido costiero della Sicilia orientale). Animalia 1976, 3, 81–103. [Google Scholar]
- La Mantia, A.; Scuderi, L. Distribuzione presente e passata di Ziziphus lotus (L.) Lam. subsp. lotus (Rhamnaceae) in Sicilia. Nat. Sicil. 2021, 45, 7–28. [Google Scholar]
- Turner, M.G.; Gardner, R.H.; O’Neill, R.V. Landscape Ecology in Theory and Practice: Pattern and Process; Springer: New York, NY, USA, 2001. [Google Scholar]
- Uuemaa, E.; Mander, Ü.; Marja, R. Trends in the use of landscape spatial metrics as landscape indicators: A review. Ecol. Indic. 2013, 28, 100–106. [Google Scholar] [CrossRef]
- Tomaselli, V.; Mantino, F.; Tarantino, C.; Albanese, G.; Adamo, P. Changing landscapes: Habitat monitoring and land transformation in a long-time used Mediterranean coastal wetland. Wetl. Ecol. Manag. 2022, 31, 1–28. [Google Scholar] [CrossRef]
- Ben-Avraham, Z.; Grasso, M. Collisional zone segmentation in Sicily and surrounding areas in the Central Mediterranean. Ann. Tectonicae 1990, 4, 131–139. [Google Scholar]
- Garfì, G.; Carimi, F.; Pasta, S.; Rühl, J.; Trigila, S. Additional insights on the ecology of the relic tree Zelkova sicula Di Pasquale, Garfì & Quézel (Ulmaceae) after the finding of a new population. Flora 2011, 206, 407–417. [Google Scholar]
- Guarino, R.; Raimondo, F.M.; Domina, G. A new species of Anthemis sect. Hiorthia (Asteraceae) SE Sicily. Plant Biosyst. 2013, 147, 821–825. [Google Scholar] [CrossRef]
- Zampino, D.; Duro, A.; Piccione, V.; Scalia, C. Termoudogrammi secondo Walter & Lieth [Thermoudogram according to Walter & Lieth]. In Atti 5° Workshop Progetto Strategico Clima Ambiente e Territorio nel Mezzogiorno; CNR: Rome, Italy, 1997; Volume 2, pp. 7–54. [Google Scholar]
- Pesaresi, S.; Biondi, E.; Casavecchia, S. Bioclimate of Italy. J. Maps 2017, 13, 955–960. [Google Scholar] [CrossRef]
- Braun-Blanquet, J. Pflanzensoziologie. Grundzüge der Vegetationskunde [Plant Sociology. Basic Course of Vegetation Science]; Springer: New York, NY, USA, 1964. [Google Scholar]
- Pignatti, S. Volume 1: Flora d’Italia & Flora Digitale. In Flora d’Italia: In 4 Volumi, 2nd ed.; Edagricole-Edizioni Agricole di New Business Media: Milano, Italy, 2017. [Google Scholar]
- Pignatti, S. Volume 2: Flora d’Italia & Flora Digitale. In Flora d’Italia: In 4 Volumi, 2nd ed.; Edagricole-Edizioni Agricole di New Business Media: Milano, Italy, 2017. [Google Scholar]
- Pignatti, S. Volume 3: Flora d’Italia & Flora Digitale. In Flora d’Italia: In 4 Volumi, 2nd ed.; Edagricole-Edizioni Agricole di New Business Media: Milano, Italy, 2018. [Google Scholar]
- Pignatti, S.; Guarino, R.; La Rosa, M. Volume 4: Flora d’Italia & Flora Digitale. In Flora d’Italia: In 4 Volumi, 2nd ed.; Edagricole-Edizioni Agricole di New Business Media: Milano, Italy, 2019. [Google Scholar]
- Portal to the Flora of Italy. Available online: http:/dryades.units.it/floritaly (accessed on 5 April 2023).
- Martellos, S.; Bartolucci, F.; Conti, F.; Galasso, G.; Moro, A.; Pennesi, R.; Peruzzi, L.; Pittao, E.; Nimis, P.L. FlorItaly—The portal to the Flora of Italy. PhytoKeys 2020, 156, 5571. [Google Scholar] [CrossRef] [PubMed]
- Giardina, G.; Raimondo, F.M.; Spadaro, V. Checklist of the vascular plants growing in Sicily. Bocconea 2007, 20, 5–582. [Google Scholar]
- McCune, B.; Mefford, M.J. PC-ORD. Multivariate Analysis of Ecological Data, Version 4.0; MjM Software Design: Gleneden Beach, OR, USA, 1999.
- Lengyel, A.; Podani, J. 12 Assessing the relative importance of methodological decisions in classifications of 13 vegetation data. J. Veg. Sci. 2015, 26, 804–815. [Google Scholar] [CrossRef]
- Biondi, E.; Blasi, C.; Allegrezza, M.; Anzellotti, I.; Azzella, M.M.; Carli, E.; Casavecchia, S.; Copiz, R.; Del Vico, E.; Facioni, L.; et al. Plant communities of Italy: The Vegetation Prodrome. Plant Biosyst. 2014, 148, 728–814. [Google Scholar] [CrossRef]
- Mucina, L.; Bültmann, H.; Dierßen, K.; Theurillat, J.P.; Raus, T.; Čarni, A.; Šumberová, K.; Willner, W.; Dengler, J.; García, R.G.; et al. Vegetation of Europe: Hierarchical floristic classification system of vascular plant, bryophyte, lichen, and algal communities. Appl. Veg. Sci. 2016, 19, 3–264. [Google Scholar] [CrossRef]
- Biondi, E.; Blasi, C. Italian Interpretation Manual of the Habitats (92/43/EEC Directive); Ministero dell’Ambiente e della Tutela del Territorio e del Mare: Roma, Italy, 2009. [Google Scholar]
- Cushman, A.; McGarigal, K.; Neel, M.C. Parsimony in landscape metrics: Strength, universality and consistency. Ecol. Indic. 2008, 8, 691–703. [Google Scholar] [CrossRef]
- Brullo, S. Taxonomic and nomenclatural notes on the genus Limonum in Sicily. Bot. Not. 1980, 133, 281–293. [Google Scholar]
- Rossi, G.; Orsenigo, S.; Gargano, D.; Montagnani, C.; Peruzzi, L.; Fenu, G.; Abel, T.; Alessandrini, A.; Astuti, G.; Bacchetta, G.; et al. Lista Rossa della Flora Italiana. 2 Endemiti e Altre Specie Minacciate; Ministero dell’Ambiente e della Tutela del Territorio e del Mare: Roma, Italy, 2020. [Google Scholar]
- Conti, F.; Manzi, A.; Pedrotti, F. Liste Rosse Regionali delle Piante D’italia; WWF–SBI: Camerino, Italy, 1997. [Google Scholar]
- Orsenigo, S.; Fenu, G.; Gargano, D.; Montagnani, C.; Abeli, T.; Alessandrini, A.; Bacchetta, G.; Bartolucci, F.; Carta, A.; Castello, M.; et al. Red list of threatened vascular plants in Italy. Plant Biosyst. 2021, 155, 310–335. [Google Scholar] [CrossRef]
- Zodda, G. Nuove aggiunte alla flora di Siracusa. Ann. R. Liceo Sc. “O. M. Corbino” Siracusa 1930, 4, 69–74. [Google Scholar]
- Martinoli, G. Poterium spinosum L. elemento del Mediterraneo Orientale a Calamosca (Capo S. Elia, Sardegna). Plant Biosyst. 1969, 103, 325–340. [Google Scholar]
- Gargano, D.; Fenu, G.; Medagli, P.; Sciandrello, S.; Bernardo, L. The status of Sarcopoterium spinosum (Rosaceae) at the western periphery of its range: Ecological constraints lead to conservation concerns. Isr. J. Plant Sci. 2007, 55, 1–13. [Google Scholar] [CrossRef]
- Sciandrello, S.; Tomaselli, G.; Minissale, P. The role of natural vegetation in the analysis of the spatio-temporal changes of coastal dune system: A case study in Sicily. J. Coast. Conserv. 2015, 19, 199–212. [Google Scholar] [CrossRef]
- Biondi, E.; Casavecchia, S.; Guerra, V. Analysis of vegetation diversity in relation to the geomorphological characteristics in the Salento coasts (Apulia-Italy). Fitosociologia 2006, 43, 25–38. [Google Scholar]
- Biondi, E.; Allegrezza, M.; Filigheddu, R. Su alcune formazioni ad Artemisia arborescens L. della Sard. Settentrionale. Boll. Soc. Sarda Sci. Nat. 1988, 26, 177–185. [Google Scholar]
- McGarigal, K.; Marks, B.J. Fragstats: Spatial Pattern Analysis Program for Quantifying Landscape Structure; Reference Manual; Forest Science Department Oregon State University: Corvallis, OR, USA, 1994; p. 60. [Google Scholar]
- Prisco, I.; Angiolini, C.; Assini, S.; Buffa, G.; Gigante, D.; Marcenò, C.; Sciandrello, S.; Villani, M.C.; Acosta, A.T.R. Conservation status of Italian coastal dune habitats in the light of the 4th Monitoring Report (92/43/EEC Habitats Directive). Plant Sociol. 2020, 57, 55–64. [Google Scholar] [CrossRef]





| Relevé number | 1 | 2 * | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | |
| Original relevé number | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | 25 | 26 | 27 | |
| Number Cluster | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | |
| Surface (mq) | 2 | 2 | 3 | 3 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | |
| Coverage (%) | 60 | 70 | 80 | 60 | 100 | 80 | 70 | 85 | 85 | 85 | 85 | |
| Altitude (m a.s.l.) | 2 | 3 | 2 | 3 | 2 | 2 | 8 | 6 | 6 | 6 | 6 | |
| Floristic richness | 6 | 8 | 10 | 4 | 11 | 14 | 4 | 7 | 6 | 7 | 7 | presence |
| Char. Ass. | ||||||||||||
| Parapholis incurva (L.) C.E. Hubb. | 2 | 2 | 3 | 3 | 3 | 2 | 2 | 4 | 3 | 4 | 3 | 11 |
| Char. Sileno sedoidis-Catapodion balearici de Foucault and Bioret 2010 corr. Tomaselli et al. 2020 | ||||||||||||
| Catapodium balearicum (Willk.) H. Scholz | + | 1 | . | . | . | . | . | . | . | . | . | 2 |
| Silene sedoides Poir. | . | + | . | . | . | . | . | . | . | . | + | 2 |
| Char. Saginetea maritimae Westhoff, Van Leeuwen and Adriani 1962, Saginetalia maritimae Westhoff, Van Leeuwen & Adriani 1962 | ||||||||||||
| Spergularia marina (L.) Besser | 3 | 4 | + | 1 | + | 3 | 1 | 1 | + | 1 | 2 | 11 |
| Beta vulgaris subsp. maritima (L.) Arcang. | + | + | 1 | . | + | 1 | . | 1 | + | 1 | . | 8 |
| Medicago littoralis Rohde ex Loisel. | + | + | + | . | 3 | 1 | . | . | . | + | + | 7 |
| Plantago coronopus L. | + | 1 | 1 | 1 | . | . | . | . | . | . | . | 4 |
| Mesembryanthemum nodiflorum L. | . | . | 1 | . | . | . | . | + | . | . | . | 2 |
| Matthiola tricuspidata (L.) W.T. Aiton | . | . | . | . | . | + | . | . | . | . | . | 1 |
| Other species | ||||||||||||
| Lotus cytisoides L. | . | . | + | 1 | + | . | + | + | 1 | + | . | 7 |
| Frankenia hirsuta L. | . | . | + | . | + | . | 2 | 1 | 1 | + | 1 | 7 |
| Trifolium resupinatum L. | . | 1 | . | . | + | + | . | + | + | + | + | 7 |
| Anthemis arvensis L. | . | . | 1 | . | 1 | + | . | . | . | . | . | 3 |
| Silene colorata Poir. | . | . | . | . | + | + | . | . | . | . | . | 2 |
| Glaucium flavum Crantz | . | . | . | . | + | + | . | . | . | . | . | 2 |
| Crithmum maritimum L. | . | . | . | . | . | + | . | . | . | . | . | 1 |
| Plantago macrorhiza Poir. | . | . | . | . | . | + | . | . | . | . | . | 1 |
| Trifolium nigrescens Viv. | . | . | . | . | . | + | . | . | . | . | . | 1 |
| Medicago truncatula Gaertn. | . | . | + | . | . | . | . | . | . | . | . | 1 |
| Lotus ornithopodioides L. | . | . | . | . | . | + | . | . | . | . | . | 1 |
| Reichardia picroides (L.) Roth | . | . | . | . | + | . | . | . | . | . | . | 1 |
| Sonchus asper (L.) Hill | . | . | . | . | . | + | . | . | . | . | . | 1 |
| Tamarix africana Poir. | . | . | . | . | . | . | . | . | . | . | + | 1 |
| Relevé number | 1 | 2 | 3 | 4 | 5 | 6 * | 7 | 8 | ||
| Original relevé number | 22 | 23 | 24 | 26 | 27 | 30 | 31 | 32 | ||
| Number Cluster | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | ||
| Surface (mq) | 16 | 16 | 16 | 16 | 16 | 16 | 16 | 16 | ||
| Coverage (%) | 80 | 75 | 85 | 90 | 100 | 90 | 90 | 90 | ||
| Altitude (m a.s.l.) | 5 | 5 | 5 | 5 | 3 | 2 | 2 | 2 | ||
| Floristic richness | 8 | 17 | 11 | 9 | 10 | 12 | 11 | 7 | presence | |
| Char. Ass. | ||||||||||
| Bolboschoenus maritimus (L.) Palla | 4 | 1 | 2 | 1 | 4 | 2 | 4 | 2 | 8 | |
| Char. Glycerio-Sparganion neglecti Br.-Bl. and Sissing in Boer 1942 | ||||||||||
| Alisma lanceolatum With. | 2 | 4 | 4 | 4 | 3 | 4 | 4 | 4 | 8 | |
| Char. Phragmito-Magnocaricetea Klika in Klika and Novák 1941 | ||||||||||
| Phragmites australis (Cav.) Trin. ex Steud. | + | . | . | 1 | . | + | 1 | 1 | 5 | |
| Veronica anagallis-aquatica L. | + | 1 | + | . | . | + | . | + | 5 | |
| Eleocharis palustris (L.) Roem. and Schult. | . | + | . | . | 1 | 1 | . | . | 3 | |
| Typha domingensis (Pers.) Steud. | . | . | . | . | . | . | 1 | 1 | 2 | |
| Carex otrubae Podp. | . | + | . | . | . | . | . | . | 1 | |
| Transgr. Molinio-Arrhenatheretea R.Tx.1937 | ||||||||||
| Potentilla reptans L. | . | + | + | + | + | 1 | + | . | 6 | |
| Lotus corniculatus L. subsp. preslii (Ten.) P.Fourn. | . | 1 | + | + | . | 1 | 1 | . | 5 | |
| Lythrum junceum Banks and Sol. | + | 1 | 1 | + | . | . | . | . | 4 | |
| Phyla nodiflora (L.) Greene | . | . | . | . | . | 2 | 1 | 1 | 3 | |
| Juncus articulatus L. | . | . | . | . | . | 2 | + | 1 | 3 | |
| Trifolium fragiferum L. | . | + | + | . | . | . | . | . | 2 | |
| Other species | 8 | |||||||||
| Symphyotrichum squamatum (Spreng.) G.L. Nesom | . | + | + | + | + | + | + | . | 6 | |
| Mentha pulegium L. | + | 1 | + | + | + | . | . | . | 5 | |
| Ranunculus trilobus Desf. | . | + | 1 | + | + | . | . | . | 4 | |
| Tamarix africana Poir. | 1 | . | . | . | 1 | . | 1 | . | 3 | |
| Plantago media L. | . | + | . | . | . | + | + | . | 3 | |
| Holcus lanatus L. | . | 1 | + | . | . | . | . | . | 2 | |
| Juncus hybridus Brot. | . | + | . | . | . | . | . | . | 1 | |
| Isolepis cernua (Vahl) Roem. and Schult. | . | . | . | . | . | + | . | . | 1 | |
| Juncus acutus L. | . | . | . | . | + | . | . | . | 1 | |
| Carex extensa Gooden. | . | + | . | . | . | . | . | . | 1 | |
| Vitex agnus-castus L. | . | . | . | . | + | . | . | . | 1 | |
| Rumex pulcher L. | . | + | . | . | . | . | . | . | 1 | |
| Chara sp. | 1 | . | . | . | . | . | . | . | 1 | |
| Relevé number | 1 | 2 * | |
| Original relevé number | 40 | 41 | |
| Number Cluster | 5 | 5 | |
| Surface (mq) | 16 | 16 | |
| Coverage (%) | 100 | 90 | |
| Altitude (m a.s.l.) | 2 | 6 | |
| Floristic richness | 18 | 20 | presence |
| Char. Ass. | |||
| Lotus corniculatus L. subsp. preslii (Ten.) P.Fourn. | 3 | 3 | 2 |
| Euphorbia hirsuta L. | 2 | 2 | 2 |
| Char. Paspalo-Agrostion semiverticillati Br.-Bl. in Br.-Bl. Roussine and Negre 1952 and Paspalo-Heleochloetalia Br.-Bl. ex Rivas Goday 1956 | |||
| Symphyotrichum squamatum (Spreng.) G. L. Nesom | 1 | + | 2 |
| Char. Molinio-Arrhenatheretea R.Tx.1937 | |||
| Phyla nodiflora (L.) Greene | 1 | 3 | 2 |
| Lythrum junceum Banks & Sol. | 3 | 1 | 2 |
| Potentilla reptans L. | + | 2 | 2 |
| Juncus articulatus L. | 1 | + | 2 |
| Scirpoides holoschoenus (L.) Soják | 1 | + | 2 |
| Teucrium scordium L. | + | + | 2 |
| Oenanthe globulosa L. | . | + | 1 |
| Kickxia commutata (Bernh. ex Rchb.) Fritsch | . | 1 | 1 |
| Trifolium resupinatum L. | + | . | 1 |
| Other species | |||
| Bolboschoenus maritimus (L.) Palla | + | + | 2 |
| Dipsacus fullonum L. | + | . | 1 |
| Rubus ulmifolius Schott | 1 | . | 1 |
| Tamarix africana Poir. | 1 | . | 1 |
| Schenkia spicata (L.) G. Mans. | . | 2 | 1 |
| Cynodon dactylon (L.) Pers. | . | 2 | 1 |
| Daucus carota L. subsp. maritimus (Lam.) Batt. | + | . | 1 |
| Xanthium italicum Moretti | . | 1 | 1 |
| Ranunculus trilobus Desf. | 1 | . | 1 |
| Mentha pulegium L. | . | 1 | 1 |
| Plantago media L. | . | 1 | 1 |
| Carex extensa Gooden. | 1 | . | 1 |
| Juncus acutus L. | 2 | . | 1 |
| Phragmites australis (Cav.) Trin. ex Steud. | . | + | 1 |
| Alisma lanceolatum With. | . | + | 1 |
| Typha domingensis (Pers.) Steud. | . | + | 1 |
| 1955 | % | 2023 | % | |
|---|---|---|---|---|
| Driftline (Euphorbion peplidis) | 29 | 7.8 | 9 | 2.6 |
| Shifting dunes (Elytrigienion junceae) | 11 | 2.8 | 4 | 1.2 |
| Saline | 19 | 5.2 | 0 | 0.0 |
| Aquatic vegetation (Ruppion maritimae) | 85 | 22.6 | 25.9 | 7.4 |
| Salt-marsh vegetation (Salicornietea fruticosae) | 53 | 14.1 | 2.2 | 0.6 |
| Hygrophylous vegetation with Juncus acutus (Juncetea maritimi) | 40 | 10.6 | 23 | 6.5 |
| Phragmites australis vegetation (Phragmito-Magnocaricetea) | 0 | 0.0 | 5.89 | 1.7 |
| Woods with Tamarix sp. pl. (Nerio-Tamaricetea) | 0 | 0.0 | 6.6 | 1.9 |
| Woods with Ulmus minor (Populion albae) | 0 | 0.0 | 0.557 | 0.2 |
| Rocky coast vegetation (Crithmo-Limonietea) | 28 | 7.4 | 11.35 | 3.2 |
| Garrigue with Thymbra capitata (Cisto-Micromerietea) | 16.13 | 4.3 | 14.31 | 4.1 |
| Maquis with Ziziphus lotus (Oleo-Ceratonion) | 0 | 0.0 | 0.87 | 0.2 |
| Shrub with Artemisia arborescens (Artemision arborescentis) | 0 | 0.0 | 0.117 | 0.0 |
| Temporary ponds with Damasonium bourgaei (Isoeto-Nanojuncetea) | 0 | 0.0 | 0.315 | 0.1 |
| Hyparrhenia hirta dry grassland (Lygeo-Stipetea) | 0 | 0.0 | 12.6 | 3.6 |
| Uncultivated lands (Echio-Galactition) | 67 | 17.8 | 96 | 27.3 |
| Rubus ulmifolius vegetation (Pruno-Rubion) | 0 | 0.0 | 3 | 0.8 |
| Cultivated lands | 28.15 | 7.5 | 6 | 1.7 |
| Reforestation | 0 | 0.0 | 44.4 | 12.6 |
| Man-made infrastructure | 0 | 0.0 | 96 | 27.3 |
| 376,274 | 362,261 |
| Habitat Type | AWMSI | MSI | MPAR | MPS | NumP | CA | TLA | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1955 | 2015 | 1955 | 2015 | 1955 | 2015 | 1955 | 2015 | 1955 | 2015 | 1955 | 2015 | 2015 | |
| 1150 | 1.74 | 2.23 | 1.62 | 1.88 | 1854.8421 | 1289.33 | 17.3 | 8.51 | 3 | 3 | 54.9 | 25.54 | 352.55 |
| 1210 | 4.74 | 3.68 | 4.74 | 3.92 | 3083.9738 | 2176.50 | 29.21 | 1.32 | 1 | 2 | 29.21 | 2.63 | 352.55 |
| 1240 | 5.31 | 6.23 | 5.31 | 2.82 | 3540.334 | 61161.51 | 27.8 | 1.26 | 1 | 9 | 27.8 | 11.34 | 352.55 |
| 1310 | 2.10 | / | 2.10 | / | 1676.7779 | / | 19.45 | / | 1 | / | 19.45 | / | 352.55 |
| 1410 | 2.69 | 2.36 | 2.69 | 2.36 | 1495.051 | 174.30 | 40.16 | 23.04 | 1 | 1 | 40.16 | 23.04 | 352.55 |
| 1420 | 3.94 | 3.94 | 2.55 | 3.37 | 2231.8372 | 3406.60 | 26.06 | 1.09 | 2 | 2 | 52.12 | 2.19 | 352.55 |
| 1430 | / | 2.18 | 2.18 | 2264.90 | 0.12 | 1 | 0.12 | 352.55 | |||||
| 2110 | 3.67 | / | 3.67 | / | 3990.6974 | / | 10.5 | / | 1 | / | 11.7 | / | 352.55 |
| 3170 | / | 1.50 | 1.50 | 947.90 | 0.31 | 1 | 0.31 | 352.55 | |||||
| 5220 | / | 1.20 | 1.20 | 3427.80 | 0.02 | 1 | 0.02 | 352.55 | |||||
| 5420 | 2.70 | 3.33 | 1.84 | 3.19 | 3998.6185 | 3336.03 | 5.4 | 1.79 | 3 | 8 | 16.13 | 14.30 | 352.55 |
| 6220 | / | 2.57 | 1.94 | 35172.79 | 1.62 | 11 | 17.87 | 352.55 | |||||
| 9320 | / | 1.96 | 1.96 | 5281.10 | 0.02 | 1 | 0.02 | 352.55 | |||||
| Habitat | ha | % | Conservation Status and Trend |
|---|---|---|---|
| 1150 * Coastal lagoons | 25.9 | 17.7 | FV |
| 1210 Annual vegetation of drift lines | 9 | 6.1 | FV |
| 1240 Vegetated sea cliffs of the Mediterranean coasts with endemic Limonium spp. | 11.35 | 7.7 | U1 (-) |
| 1310 Salicornia and other annuals colonizing mud and sand | 0.1 | 0.1 | U1 (=) |
| 1410 Mediterranean salt meadows (Juncetalia maritimi) | 23 | 15.7 | U1 (x) |
| 0.0 | |||
| 1420 Mediterranean and thermo-Atlantic halophilous scrubs (Sarcocornetiea fruticosi) | 2.2 | 1.5 | U1 (x) |
| 1430 Halo-nitrophilous scrubs (Pegano-Salsoletea) | 0.12 | 0.1 | FV |
| 2110 Embryonic shifting dunes | 4 | 2.7 | U2 (=) |
| 2210 Crucianellion maritimae fixed beach dunes | ? | 0 | U2 (-) |
| 2230 Malcolmietalia dune grasslands | 0.2 | 0.1 | U1 (x) |
| 3170 * Mediterranean temporary ponds | 0.32 | 0.2 | U1 (=) |
| 5220 * Arborescent matorral with Zyziphus | 0.1 | 0.1 | U2 (-) |
| 5420 Sarcopoterium spinosum phryganas | 14.3 | 9.8 | FV |
| 6220 * Pseudo-steppe with grasses and annuals of the Thero-Brachypodietea | 12.6 | 8.6 | FV |
| 92D0 Southern riparian galleries and thickets (Nerio-Tamaricetea and Securinegion tinctoriae) | 6.6 | 4.5 | U1 (-) |
| 9320 Olea and Ceratonia forests | 0.8 | 0.5 | U1 (-) |
| 110.6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sciandrello, S.; Ranno, V.; Tomaselli, V. The Role of Vegetation Monitoring in the Conservation of Coastal Habitats N2000: A Case Study of a Wetland Area in Southeast Sicily (Italy). Land 2024, 13, 62. https://doi.org/10.3390/land13010062
Sciandrello S, Ranno V, Tomaselli V. The Role of Vegetation Monitoring in the Conservation of Coastal Habitats N2000: A Case Study of a Wetland Area in Southeast Sicily (Italy). Land. 2024; 13(1):62. https://doi.org/10.3390/land13010062
Chicago/Turabian StyleSciandrello, Saverio, Veronica Ranno, and Valeria Tomaselli. 2024. "The Role of Vegetation Monitoring in the Conservation of Coastal Habitats N2000: A Case Study of a Wetland Area in Southeast Sicily (Italy)" Land 13, no. 1: 62. https://doi.org/10.3390/land13010062
APA StyleSciandrello, S., Ranno, V., & Tomaselli, V. (2024). The Role of Vegetation Monitoring in the Conservation of Coastal Habitats N2000: A Case Study of a Wetland Area in Southeast Sicily (Italy). Land, 13(1), 62. https://doi.org/10.3390/land13010062