Impact of Agrochemicals on Soil Microbiota and Management: A Review

 ,
, 


 , ,
, ,  , ,
, ,
Abstract

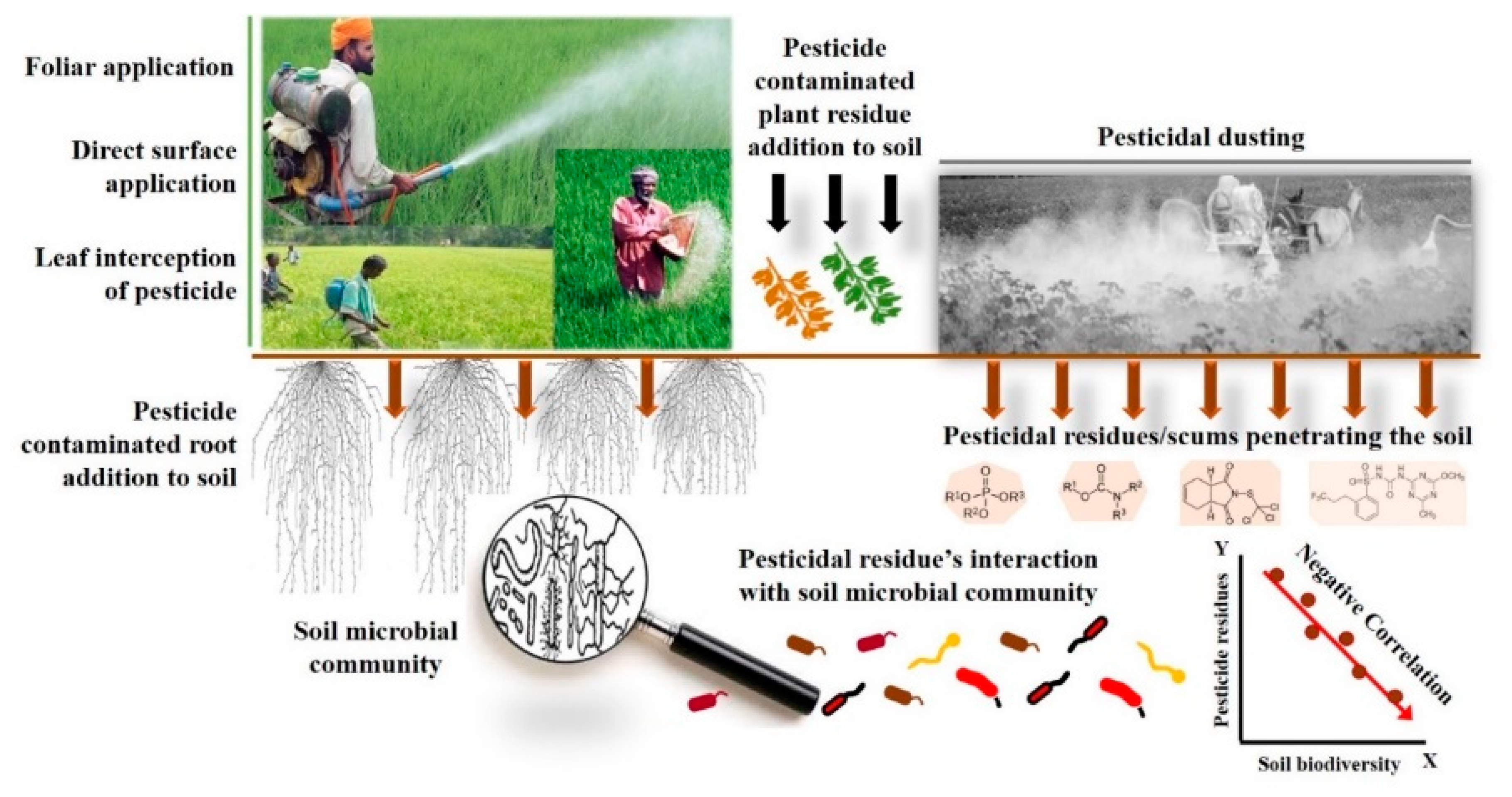
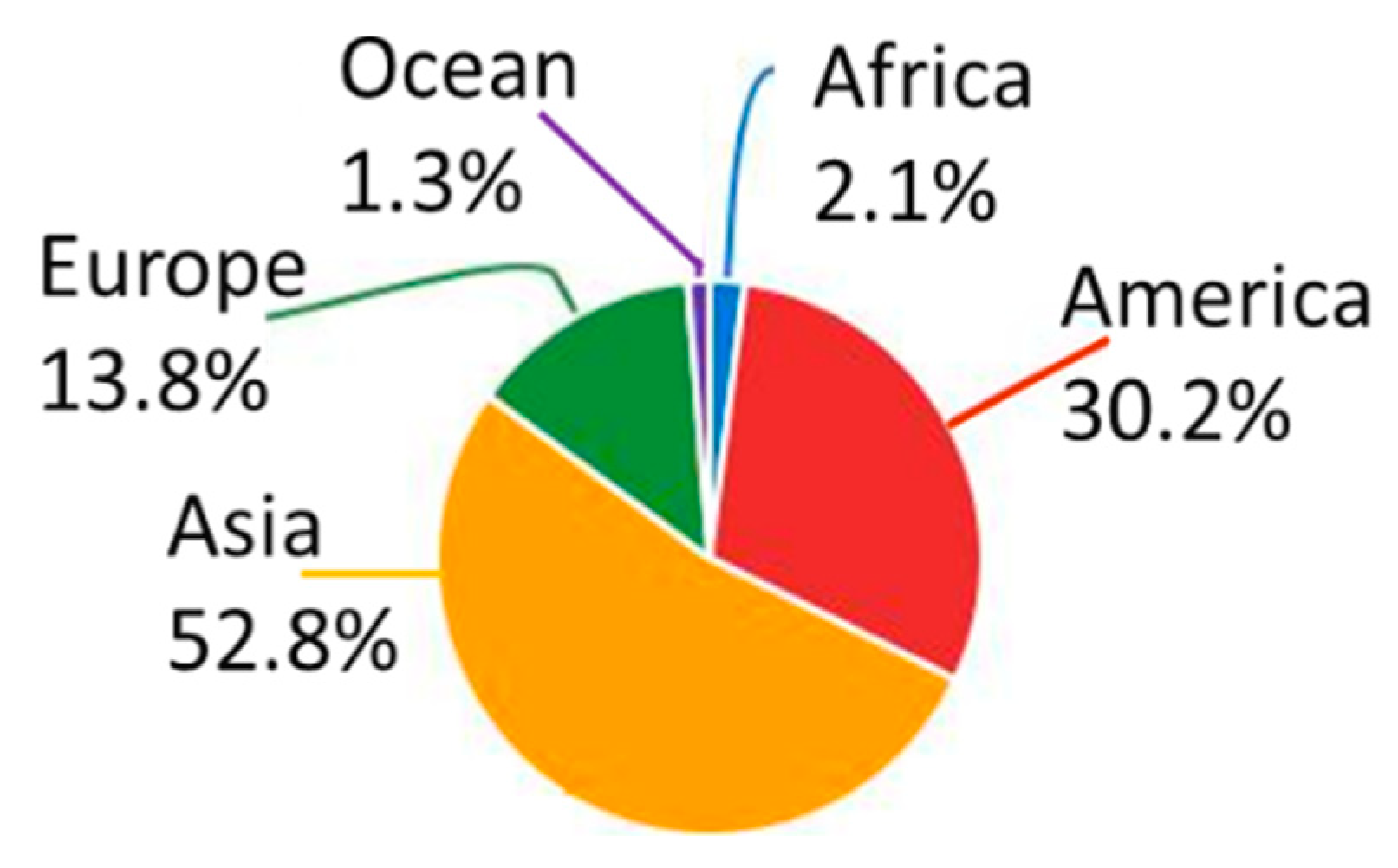
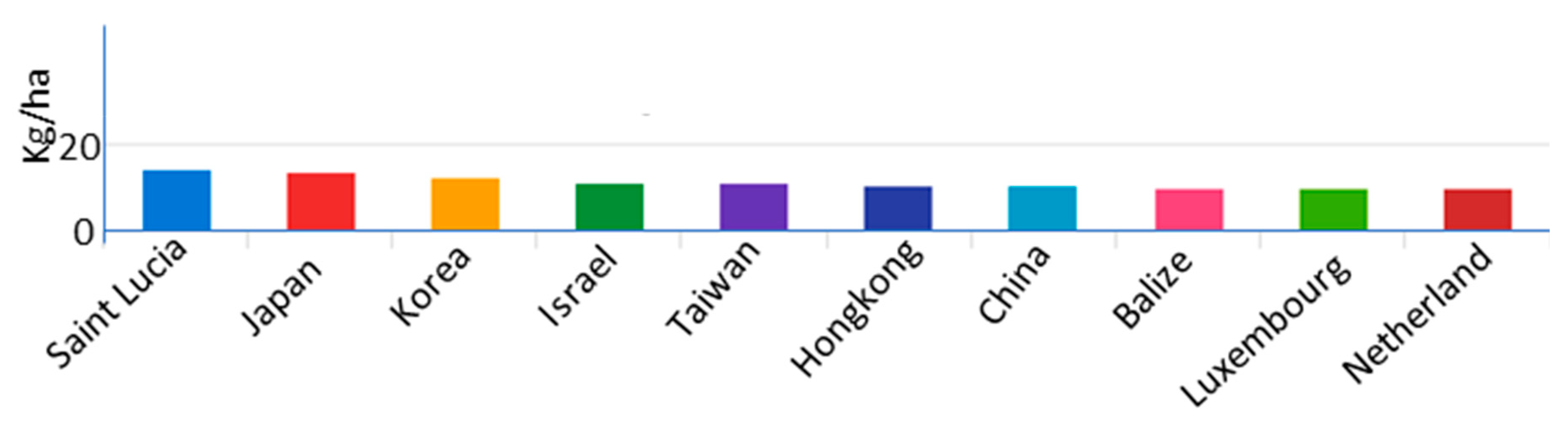
1. Introduction
2. Methodology
3. Herbicides and Soil Microbial Environment
3.1. Impacts of Herbicides on Soil Biota
3.2. Impact of Herbicides on N-Fixing Microbes
3.3. Impact of Herbicides on Arbuscular Mycorrhizal Fungi
3.4. Impact of Herbicides on Soil Biochemical and Enzymatic Environment
4. Fungicides and Soil Microbial Environment
4.1. Impact of Fungicides on N-Fixing and Growth-Promoting Microbes
4.2. Impacts of Fungicides on Soil Microbiota
4.3. Impact of Fungicides on Soil Enzymes and Biochemical Environments
5. Insecticides and Soil Microbial Environment
5.1. Impact of Insecticides on N-Fixing and Another Growth-Promoting Microorganism
5.2. Influence of Insecticides on Soil Biochemistry
5.3. Impacts of Insecticides on Agrobiology
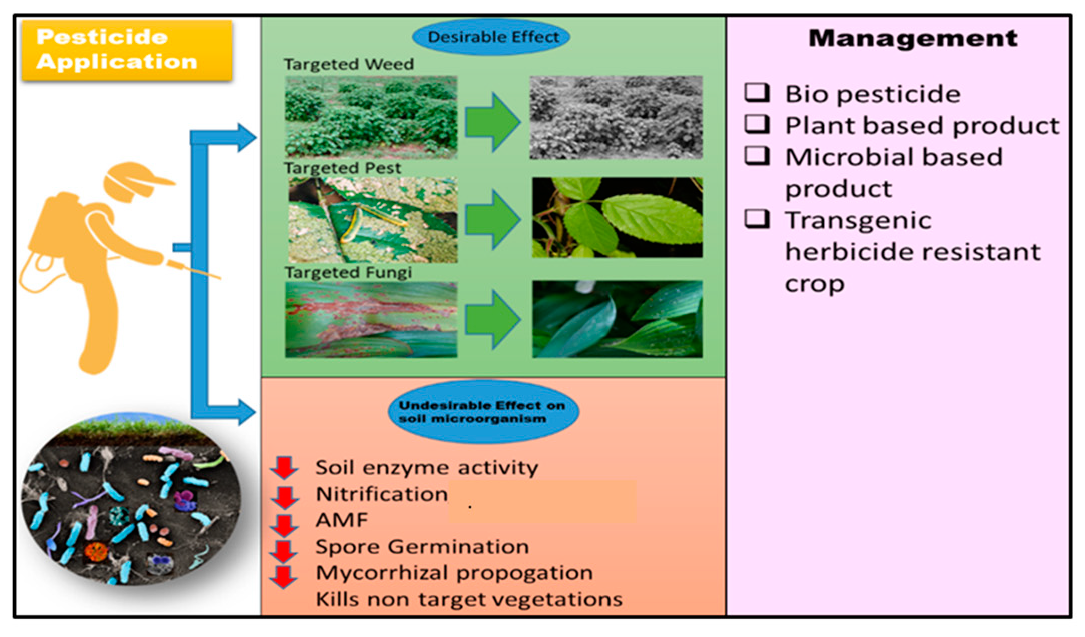
6. Management Options
6.1. Biopesticides
6.2. Plant-Based Products
6.3. Microbial-Based-Products
6.4. Transgenic Herbicide-Resistant Crops
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Yáñ ez, L.; Ortiz, D.; Calderón, J.; Batres, L.; Carrizales, L.; Mejía, J.; Martínez, L.; García-Nieto, E.; Díaz-Barriga, F. Overview of human health and chemical mixtures: Problems facing developing countries. Environ. Health Perspect. 2002, 110, 901–909. [Google Scholar] [CrossRef]
- Mishra, P.K.; Giagli, K.; Tsalagkas, D.; Mishra, H.; Talegaonkar, S.; Gryc, V.; Wimmer, R. Changing Face of Wood Science in Modern Era: Contribution of Nanotechnology. Recent Pat. Nanotechnol. 2018, 12, 13–21. [Google Scholar] [CrossRef]
- Mishra, P.K.; Gregor, T.; Wimmer, R. Utilising Brewer’s Spent Grain as a Source of Cellulose Nanofibres Following Separation of Protein-based Biomass. BioResources 2017, 12, 107–116. [Google Scholar] [CrossRef]
- Marfo, T.D.; Datta, R.; Pathan, S.I.; Vranová, V. Ecotone Dynamics and Stability from Soil Scientific Point of View. Diversity 2019, 11, 53. [Google Scholar] [CrossRef]
- Danso Marfo, T.; Datta, R.; Vranová, V.; Ekielski, A. Ecotone Dynamics and Stability from Soil Perspective: Forest-Agriculture Land Transition. Agriculture 2019, 9, 228. [Google Scholar] [CrossRef]
- Meena, H.; Meena, R.S.; Rajput, B.S.; Kumar, S. Response of bio-regulators to morphology and yield of clusterbean [Cyamopsis tetragonoloba (L.) Taub.] under different sowing environments. J. Appl. Nat. Sci. 2016, 8, 715–718. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations. FAOSTAT Statistical Database; FAO: Rome, Italy, 2019. [Google Scholar]
- Önder, M.; Ceyhan, E.; Kahraman, A. Effects of Agricultural Practices on Environment. Biol. Environ. Chem. 2011, 24, 28–32. [Google Scholar]
- Oliveira, C.M.; Auad, A.M.; Mendes, S.M.; Frizzas, M.R. Crop losses and the economic impact of insect pests on Brazilian agriculture. Crop Prot. 2014, 56, 50–54. [Google Scholar] [CrossRef]
- Meena, R.S.; Bohra, J.S.; Singh, S.P.; Meena, V.S.; Verma, J.P.; Verma, S.K.; Sihag, S.K. Towards the prime response of manure to enhance nutrient use efficiency and soil sustainability a current need: A book review. J. Clean. Prod. 2016, 112, 1258–1260. [Google Scholar] [CrossRef]
- Bahadur, S.; Verma, S.K.; Prasad, S.K.; Madane, A.J.; Maurya, S.P.; Gaurav Verma, V.K.; Sihag, S.K. Eco-friendly weed management for sustainable crop production-A review. J. Crop Weed 2015, 11, 181–189. [Google Scholar]
- Aktar, W.; Sengupta, D.; Chowdhury, A. Impact of pesticides use in agriculture: Their benefits and hazards. Interdiscip. Toxicol. 2009, 2, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.-C.; Gong, M.; Zang, H.-B.; Hua, X.-M.; Yao, J.; Pang, Y.-J.; Yang, Y.-H. Effect of Methamidophos and Urea Application on Microbial Communities in Soils as Determined by Microbial Biomass and Community Level Physiological Profiles. J. Environ. Sci. Health B 2006, 41, 399–413. [Google Scholar] [CrossRef] [PubMed]
- Miller, G.T. Sustaining the Earth; Brooks/Cole: Monterey County, CA, USA, 2004; ISBN 9780534400880. [Google Scholar]
- Lo, C.-C. Effect of pesticides on soil microbial community. J. Environ. Sci. Health Part B 2010, 45, 348–359. [Google Scholar] [CrossRef]
- Santos, A.; Flores, M. Effects of glyphosate on nitrogen fixation of free-living heterotrophic bacteria. Lett. Appl. Microbiol. 1995, 20, 349–352. [Google Scholar] [CrossRef]
- Fabra, A.; Duffard, R.; Duffard, A.E. de Toxicity of 2,4-Dichlorophenoxyacetic Acid to Rhizobium sp in Pure Culture. Bull. Environ. Contam. Toxicol. 1997, 59, 645–652. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.; Siddique, T.; Saleem, M.; Arshad, M.; Khalid, A. Chapter 5 Impact of Pesticides on Soil Microbial Diversity, Enzymes, and Biochemical Reactions. Adv. Agron. 2009, 102, 159–200. [Google Scholar]
- Monkiedje, A.; Spiteller, M. Degradation of Metalaxyl and Mefenoxam and Effects on the Microbiological Properties of Tropical and Temperate Soils. Int. J. Environ. Res. Public. Health 2005, 2, 272–285. [Google Scholar] [CrossRef]
- Bano, S.A.; Iqbal, S.M. Biological Nitrogen Fixation to Improve Plant Growth and Productivity. Int. J. Agric. Innov. Res. 2016, 4, 2319-1473. [Google Scholar]
- Reitz, M.U.; Gifford, M.L.; Schäfer, P. Hormone activities and the cell cycle machinery in immunity-triggered growth inhibition. J. Exp. Bot. 2015, 66, 2187–2197. [Google Scholar] [CrossRef]
- Milosevic, N.; Govedarica, M. Effect of herbicides on microbiological properties of soil. Matica Srp. Proc. Nat. Sci. 2002, 102, 5–21. [Google Scholar] [CrossRef]
- Kremer, R.J.; Means, N.E. Glyphosate and glyphosate-resistant crop interactions with rhizosphere microorganisms. Eur. J. Agron. 2009, 31, 153–161. [Google Scholar] [CrossRef]
- Yadav, G.; Datta, R.; Imran Pathan, S.; Lal, R.; Meena, R.; Babu, S.; Das, A.; Bhowmik, S.; Datta, M.; Saha, P. Effects of Conservation Tillage and Nutrient Management Practices on Soil Fertility and Productivity of Rice (Oryza sativa L.)–Rice System in North Eastern Region of India. Sustainability 2017, 9, 1816. [Google Scholar] [CrossRef]
- Santos, J.B.; Jakelaitis, A.; Silva, A.A.; Costa, M.D.; Manabe, A.; Silva, M.C.S. Action of two herbicides on the microbial activity of soil cultivated with common bean (Phaseolus vulgaris) in conventional-till and no-till systems. Weed Res. 2006, 46, 284–289. [Google Scholar] [CrossRef]
- Chen, F.; Dixon, R.A. Lignin modification improves fermentable sugar yields for biofuel production. Nat. Biotechnol. 2007, 25, 759–761. [Google Scholar] [CrossRef]
- Datta, R.; Kelkar, A.; Baraniya, D.; Molaei, A.; Moulick, A.; Meena, R.S.; Formanek, P. Enzymatic degradation of lignin in soil: A review. Sustainability 2017, 9, 1163. [Google Scholar] [CrossRef]
- Mishra, P.K.; Wimmer, R. Aerosol assisted self-assembly as a route to synthesize solid and hollow spherical lignin colloids and its utilization in layer by layer deposition. Ultrason. Sonochem. 2017, 35, 45–50. [Google Scholar] [CrossRef]
- Mishra, P.K.; Ekielski, A. The Self-Assembly of Lignin and Its Application in Nanoparticle Synthesis: A Short Review. Nanomaterials 2019, 9, 243. [Google Scholar] [CrossRef]
- Smith, A.R.W.; Beadle, C.A. Induction of enzymes of 2,4-dichlorophenoxyacetate degradation in Burkholderia cepacia 2a and toxicity of metabolic intermediates. Biodegradation 2008, 19, 669–681. [Google Scholar] [CrossRef]
- Tsui, M.T.K.; Chu, L.M. Aquatic toxicity of glyphosate-based formulations: Comparison between different organisms and the effects of environmental factors. Chemosphere 2003, 52, 1189–1197. [Google Scholar] [CrossRef]
- Brtnicky, M.; Dokulilova, T.; Holatko, J.; Pecina, V.; Kintl, A.; Latal, O.; Vyhnanek, T.; Prichystalova, J.; Datta, R. Long-Term Effects of Biochar-Based Organic Amendments on Soil Microbial Parameters. Agronomy 2019, 9, 747. [Google Scholar] [CrossRef]
- Khan, M.S.; Zaidi, A.; Rizvi, P.Q. Biotoxic Effects of Herbicides on Growth, Nodulation, Nitrogenase Activity, and Seed Production in Chickpeas. Commun. Soil Sci. Plant Anal. 2006, 37, 1783–1793. [Google Scholar] [CrossRef]
- Seghers, D.; Verthé, K.; Reheul, D.; Bulcke, R.; Siciliano, S.D.; Verstraete, W.; Top, E.M. Effect of long-term herbicide applications on the bacterial community structure and function in an agricultural soil. FEMS Microbiol. Ecol. 2003, 46, 139–146. [Google Scholar] [CrossRef]
- Sannino, F.; Gianfreda, L. Pesticide influence on soil enzymatic activities. Chemosphere 2001, 45, 417–425. [Google Scholar] [CrossRef]
- Chalam, A.V.; Sasikala, C.; Ramana, C.V.; Uma, N.R.; Rao, P.R. Effect of Pesticides on the Diazotrophic Growth and Nitrogenase Activity of Purple Nonsulfur Bacteria. Bull. Environ. Contam. Toxicol. 1997, 58, 463–468. [Google Scholar] [CrossRef]
- Fox, J.E.; Starcevic, M.; Kow, K.Y.; Burow, M.E.; McLachlan, J.A. Nitrogen fixation: Endocrine disrupters and flavonoid signalling. Nature 2001, 413, 128–129. [Google Scholar] [CrossRef]
- Syamsul Arif, M.A.; Houwen, F.; Verstraete, W. Agricultural factors affecting methane oxidation in arable soil. Biol. Fertil. Soils 1996, 21, 95–102. [Google Scholar] [CrossRef]
- Subhani, A.; El-ghamry, A.M.; Changyong, H.; Jianming, X. Effects of Pesticides (Herbicides) on Soil Microbial Biomass - A Review. Pak. J. Biol. Sci. 2000, 3, 705–709. [Google Scholar]
- Singh, G.; Wright, D. In vitro studies on the effects of herbicides on the growth of rhizobia. Lett. Appl. Microbiol. 2002, 35, 12–16. [Google Scholar] [CrossRef]
- Nowak, J.; Kaklewski, K.; Klódka, D. Influence of various concentrations of selenic acid (IV) on the activity of soil enzymes. Sci. Total Environ. 2002, 291, 105–110. [Google Scholar] [CrossRef]
- Makarian, H.; Poozesh, V.; Asghari, H.R.; Nazari, M. Interaction Effects of Arbuscular Mycorrhiza Fungi and Soil Applied Herbicides on Plant Growth. Commun. Soil Sci. Plant Anal. 2016, 47, 619–629. [Google Scholar] [CrossRef]
- Meena, R.S.; Meena, V.S.; Meena, S.K.; Verma, J.P. The needs of healthy soils for a healthy world. J. Clean. Prod. 2015, 102, 560–561. [Google Scholar] [CrossRef]
- Powell, J.R.; Gulden, R.H.; Hart, M.M.; Campbell, R.G.; Levy-Booth, D.J.; Dunfield, K.E.; Pauls, K.P.; Swanton, C.J.; Trevors, J.T.; Klironomos, J.N. Mycorrhizal and Rhizobial Colonization of Genetically Modified and Conventional Soybeans. Appl. Environ. Microbiol. 2007, 73, 4365–4367. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zawoznik, M.S.; Tomaro, M.L. Effect of chlorimuron-ethyl onBradyrhizobium japonicum and its symbiosis with soybean. Pest Manag. Sci. 2005, 61, 1003–1008. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, J.B.; Ferreira, E.A.; Kasuya, M.C.M.; da Silva, A.A.; de Oliveira Procópio, S. Tolerance of Bradyrhizobium strains to glyphosate formulations. Crop Prot. 2005, 24, 543–547. [Google Scholar] [CrossRef]
- Strandberg, M.; Scott-Fordsmand, J.J. Effects of pendimethalin at lower trophic levels—a review. Ecotoxicol. Environ. Saf. 2004, 57, 190–201. [Google Scholar] [CrossRef]
- Govedarica, M.; Miloševiã, N.; Konstantinoviã, B. Uticaj dimetenamida i metalahlora na mikrobiološka svojstva zemljišta pod šeãernom repom. V Jugosl. Savetov. O Zašt. Bilja Zlatibor 2001, 12, 3–8. [Google Scholar]
- Smith, S.E.; Read, D. Growth and carbon economy of arbuscular mycorrhizal symbionts. Mycorrhizal Symbiosis 2008, 117–144. [Google Scholar] [CrossRef]
- Pasaribu, A.; Mohamad, R.B.; Hashim, A.; Rahman, Z.A.; Omar, D.; Morshed, M.M.; Selangor, D.E. Effect of herbicide on sporulation and infectivity of vesicular arbuscular mycorrhizal (Glomus mosseae) symbiosis with peanut plant. J. Anim. Plant Sci. 2013, 23, 1671–1678. [Google Scholar]
- Das, A.C.; Debnath, A.; Mukherjee, D. Effect of the herbicides oxadiazon and oxyfluorfen on phosphates solubilizing microorganisms and their persistence in rice fields. Chemosphere 2003, 53, 217–221. [Google Scholar] [CrossRef]
- Zaller, J.G.; Heigl, F.; Ruess, L.; Grabmaier, A. Glyphosate herbicide affects belowground interactions between earthworms and symbiotic mycorrhizal fungi in a model ecosystem. Sci. Rep. 2014, 4, 5634. [Google Scholar] [CrossRef]
- Savin, M.C.; Purcell, L.C.; Daigh, A.; Manfredini, A. Response of Mycorrhizal Infection to Glyphosate Applications and P Fertilization in Glyphosate-Tolerant Soybean, Maize, and Cotton. J. Plant Nutr. 2009, 32, 1702–1717. [Google Scholar] [CrossRef]
- Sharma, M.P.; Buyer, J.S. Comparison of biochemical and microscopic methods for quantification of arbuscular mycorrhizal fungi in soil and roots. Appl. Soil Ecol. 2015, 95, 86–89. [Google Scholar] [CrossRef]
- Druille, M.; Omacini, M.; Golluscio, R.A.; Cabello, M.N. Arbuscular mycorrhizal fungi are directly and indirectly affected by glyphosate application. Appl. Soil Ecol. 2013, 72, 143–149. [Google Scholar] [CrossRef]
- Ipsilantis, I.; Samourelis, C.; Karpouzas, D.G. The impact of biological pesticides on arbuscular mycorrhizal fungi. Soil Biol. Biochem. 2012, 45, 147–155. [Google Scholar] [CrossRef]
- Gupta, A.; Aggarwal, A.; Chhavi, M.; Kumar, A.; Tanwar, A. Effect of herbicides Fenoxaprop-P-ethyl and 2, 4-D Ethyl-ester on soil mycoflora including VAM fungi in wheat crop. Indian J. Weed Sci. India 2011, 43, 32–40. [Google Scholar]
- Kiers, E.T.; Duhamel, M.; Beesetty, Y.; Mensah, J.A.; Franken, O.; Verbruggen, E.; Fellbaum, C.R.; Kowalchuk, G.A.; Hart, M.M.; Bago, A. Reciprocal rewards stabilize cooperation in the mycorrhizal symbiosis. Science 2011, 333, 880–882. [Google Scholar] [CrossRef] [PubMed]
- Timmusk, S.; Behers, L.; Muthoni, J.; Muraya, A.; Aronsson, A.-C. Perspectives and challenges of microbial application for crop improvement. Front. Plant Sci. 2017, 8, 49. [Google Scholar] [CrossRef]
- Kinney, C.A.; Mandernack, K.W.; Mosier, A.R. Laboratory investigations into the effects of the pesticides mancozeb, chlorothalonil, and prosulfuron on nitrous oxide and nitric oxide production in fertilized soil. Soil Biol. Biochem. 2005, 37, 837–850. [Google Scholar] [CrossRef]
- Mahía, J.; Cabaneiro, A.; Carballas, T.; Díaz-Raviña, M. Microbial biomass and C mineralization in agricultural soils as affected by atrazine addition. Biol. Fertil. Soils 2008, 45, 99–105. [Google Scholar] [CrossRef]
- Antonious, G.F. Impact of Soil Management and Two Botanical Insecticides on Urease and Invertase Activity. J. Environ. Sci. Health Part B 2003, 38, 479–488. [Google Scholar] [CrossRef]
- Virág, D.; Naár, Z.; Kiss, A. Microbial Toxicity of Pesticide Derivatives Produced with UV-photodegradation. Bull. Environ. Contam. Toxicol. 2007, 79, 356–359. [Google Scholar] [CrossRef] [PubMed]
- Sáez, F.; Pozo, C.; Gómez, M.A.; Martínez-Toledo, M.V.; Rodelas, B.; Gónzalez-López, J. Growth and denitrifying activity of Xanthobacter autotrophicus CECT 7064 in the presence of selected pesticides. Appl. Microbiol. Biotechnol. 2005, 71, 563–567. [Google Scholar] [CrossRef] [PubMed]
- Demanou, J.; Sharma, S.; Weber, A.; Wilke, B.-M.; Njine, T.; Monkiedje, A.; Munch, J.C.; Schloter, M. Shifts in microbial community functions and nitrifying communities as a result of combined application of copper and mefenoxam. FEMS Microbiol. Lett. 2006, 260, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Monkiedje, A. Soil quality changes resulting from the application of the fungicides mefenoxam and metalaxyl to a sandy loam soil. Soil Biol. Biochem. 2002, 34, 1939–1948. [Google Scholar] [CrossRef]
- Madhuri, R.J.; Rangaswamy, V. Influence of selected insecticides on phosphatase activity in groundnut (Arachis hypogeae L.) soils. J. Environ. Biol. 2002, 23, 393–397. [Google Scholar] [PubMed]
- Van Zwieten, L.; Ayres, M.R.; Morris, S.G. Influence of arsenic co-contamination on DDT breakdown and microbial activity. Environ. Pollut. 2003, 124, 331–339. [Google Scholar] [CrossRef]
- Kyei-Boahen, S.; Slinkard, A.E.; Walley, F.L. Rhizobial survival and nodulation of chickpea as influenced by fungicide seed treatment. Can. J. Microbiol. 2001, 47, 585–589. [Google Scholar] [CrossRef]
- Kostov, O.; Van Cleemput, O. Microbial Activity of Cu Contaminated Soils and Effect of Lime and Compost on Soil Resiliency. Compost Sci. Util. 2001, 9, 336–351. [Google Scholar] [CrossRef]
- Fravel, D.R.; Deahl, K.L.; Stommel, J.R. Compatibility of the biocontrol fungus Fusarium oxysporum strain CS-20 with selected fungicides. Biol. Control 2005, 34, 165–169. [Google Scholar] [CrossRef]
- Cycoń, M.; Piotrowska-Seget, Z.; Kaczyńska, A.; Kozdrój, J. Microbiological characteristics of a sandy loam soil exposed to tebuconazole and λ-cyhalothrin under laboratory conditions. Ecotoxicology 2006, 15, 639–646. [Google Scholar] [CrossRef]
- Datta, R.; Anand, S.; Moulick, A.; Baraniya, D.; Pathan, S.I.; Rejsek, K.; Vranova, V.; Sharma, M.; Sharma, D.; Kelkar, A.; et al. How enzymes are adsorbed on soil solid phase and factors limiting its activity: A Review. Int. Agrophysics 2017, 31, 287–302. [Google Scholar] [CrossRef]
- Smith, M.D.; Hartnett, D.C.; Rice, C.W. Effects of long-term fungicide applications on microbial properties in tallgrass prairie soil. Soil Biol. Biochem. 2000, 32, 935–946. [Google Scholar] [CrossRef]
- Murillo-Williams, A.; Pedersen, P. Arbuscular Mycorrhizal Colonization Response to Three Seed-Applied Fungicides. Agron. J. 2008, 100, 795. [Google Scholar] [CrossRef]
- Chen, S.-K.; Edwards, C.A.; Subler, S. Effects of the fungicides benomyl, captan and chlorothalonil on soil microbial activity and nitrogen dynamics in laboratory incubations. Soil Biol. Biochem. 2001, 33, 1971–1980. [Google Scholar] [CrossRef]
- Shukla, A.K. Impact of fungicides on soil microbial population and enzyme activities. Acta Bot. Indica 2000, 28, 85–88. [Google Scholar]
- Di Ciocco, C.A.; Rodríguez, C.E. Effect of the fungicide captan on Azospirillum brasilense Cd in pure culture and associated with Setaria italica. Rev. Argent. Microbiol. 1997, 29, 152. [Google Scholar]
- Milenkovski, S.; Bååth, E.; Lindgren, P.E.; Berglund, O. Toxicity of fungicides to natural bacterial communities in wetland water and sediment measured using leucine incorporation and potential denitrification. Ecotoxicology 2010, 19, 285–294. [Google Scholar] [CrossRef]
- Niewiadomska, A. Effect of Carbendazim, Imazetapir and Thiram on Nitrogenase Activity, the Number of Microorganisms in Soil and Yield of Red Clover (Trifolium pratense L.). Pol. J. Environ. Stud. 2004, 13, 4. [Google Scholar]
- Cycoń, M.; Piotrowska-Seget, Z.; Kozdrój, J. Responses of indigenous microorganisms to a fungicidal mixture of mancozeb and dimethomorph added to sandy soils. Int. Biodeterior. Biodegrad. 2010, 64, 316–323. [Google Scholar] [CrossRef]
- Černohlávková, J.; Jarkovský, J.; Hofman, J. Effects of fungicides mancozeb and dinocap on carbon and nitrogen mineralization in soils. Ecotoxicol. Environ. Saf. 2009, 72, 80–85. [Google Scholar] [CrossRef]
- Liebich, J.; Schäffer, A.; Burauel, P. Structural and functional approach to studying pesticide side-effects on specific soil functions. Environ. Toxicol. Chem. 2003, 22, 784–790. [Google Scholar] [CrossRef] [PubMed]
- Verdisson, S.; Couderchet, M.; Vernet, G. Effects of procymidone, fludioxonil and pyrimethanil on two non-target aquatic plants. Chemosphere 2001, 44, 467–474. [Google Scholar] [CrossRef]
- Durska, G. Fungicide effect on nitrogenase activity in methylotrophic bacteria. Pol. J. Microbiol. 2004, 53, 155–158. [Google Scholar]
- Madhuri, R.J.; Rangaswamy, V. Influence of selected fungicides on microbial population in groundnut (Arachis hypogeae L.) soils. Pollut. Res. 2003, 22, 205–212. [Google Scholar]
- Sukul, P. Enzymatic activities and microbial biomass in soil as influenced by metalaxyl residues. Soil Biol. Biochem. 2006, 38, 320–326. [Google Scholar] [CrossRef]
- Yang, Q.; Zhang, J.; Zhu, K.; Zhang, H. Influence of oxytetracycline on the structure and activity of microbial community in wheat rhizosphere soil. J. Environ. Sci. 2009, 21, 954–959. [Google Scholar] [CrossRef]
- Pal, R.; Chakrabarti, K.; Chakraborty, A.; Chowdhury, A. Pencycuron application to soils: Degradation and effect on microbiological parameters. Chemosphere 2005, 60, 1513–1522. [Google Scholar] [CrossRef]
- Pereyra, M.A.; Ballesteros, F.M.; Creus, C.M.; Sueldo, R.J.; Barassi, C.A. Seedlings growth promotion by Azospirillum brasilense under normal and drought conditions remains unaltered in Tebuconazole-treated wheat seeds. Eur. J. Soil Biol. 2009, 45, 20–27. [Google Scholar] [CrossRef]
- Yen, J.-H.; Chang, J.-S.; Huang, P.-J.; Wang, Y.-S. Effects of fungicides triadimefon and propiconazole on soil bacterial communities. J. Environ. Sci. Health Part B 2009, 44, 681–689. [Google Scholar] [CrossRef]
- Wainwright, M.; Pugh, G.J.F. Effect of fungicides on the numbers of micro-organisms and frequency of cellulolytic fungi in soils. Plant Soil 1975, 43, 561–572. [Google Scholar] [CrossRef]
- Carr, J.F.; Gregory, S.T.; Dahlberg, A.E. Severity of the Streptomycin Resistance and Streptomycin Dependence Phenotypes of Ribosomal Protein S12 of Thermus thermophilus Depends on the Identity of Highly Conserved Amino Acid Residues. J. Bacteriol. 2005, 187, 3548–3550. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Toledo, M.V.; Salmerón, V.; Rodelas, B.; Pozo, C.; González-López, J. Effects of the fungicide Captan on some functional groups of soil microflora. Appl. Soil Ecol. 1998, 7, 245–255. [Google Scholar] [CrossRef]
- Marfo, T.D.; Datta, R.; Lojkova, L.; Janous, D.; Pavelka, M.; Formanek, P. Limitation of Activity of Acid Phosphomonoesterase in Soils; Springer: Wien, Austria, 2015; Volume 47, p. 1691. [Google Scholar]
- Datta, R.; Vranová, V.; Pavelka, M.; Rejšek, K.; Formánek, P. Effect of soil sieving on respiration induced by low-molecular-weight substrates. Int. Agrophysics 2014, 28, 119–124. [Google Scholar] [CrossRef]
- Molaei, A.; Lakzian, A.; Datta, R.; Haghnia, G.; Astaraei, A.; Rasouli-Sadaghiani, M.; Ceccherini, M.T. Impact of chlortetracycline and sulfapyridine antibiotics on soil enzyme activities. Int. Agrophys. 2017, 31, 499–505. [Google Scholar] [CrossRef]
- Molaei, A.; Lakzian, A.; Haghnia, G.; Astaraei, A.; Rasouli-Sadaghiani, M.; Ceccherini, M.T.; Datta, R. Assessment of some cultural experimental methods to study the effects of antibiotics on microbial activities in a soil: An incubation study. PLoS ONE 2017, 12, e0180663. [Google Scholar] [CrossRef]
- Pandey, S.; Singh, D.K. Total bacterial and fungal population after chlorpyrifos and quinalphos treatments in groundnut (Arachis hypogaea L.) soil. Chemosphere 2004, 55, 197–205. [Google Scholar] [CrossRef]
- Survery, S.; Ahmad, S.; Subhan, S.A.; Ajaz, M.; Rasool, S.A. Hydrocarbon Degrading Bacteria from Pakistani Soil: Isolation, Identification, Screening and Genetical Studies. Pak. J. Biol. Sci. 2004, 7, 1518–1522. [Google Scholar]
- Amirkhanov, D.V.; Nikolenko, A.G.; Bagautdinov, F.Y.; Kirillova, S.S. Effect of production dosage of gamma-HCCH, foxim, cypermethrin and chlorfluazuron on soil microorganisms [grey forest soils].[Russian]. Agrokhimiya 1994, 2, 83–88. [Google Scholar]
- Ahmed, S.; Ahmad, M. Note: Toxicity of some insecticides onBracon hebetor under laboratory conditions. Phytoparasitica 2006, 34, 401–404. [Google Scholar] [CrossRef]
- Gundi, V.A.K.B.; Narasimha, G.; Reddy, B.R. Interaction Effects of Insecticides on Microbial Populations and Dehydrogenase Activity in a Black Clay Soil. J. Environ. Sci. Health Part B 2005, 40, 69–283. [Google Scholar] [CrossRef]
- Madhaiyan, M.; Poonguzhali, S.; Hari, K.; Saravanan, V.S.; Sa, T. Influence of pesticides on the growth rate and plant-growth promoting traits of Gluconacetobacter diazotrophicus. Pestic. Biochem. Physiol. 2006, 84, 143–154. [Google Scholar] [CrossRef]
- Das, A.C.; Mukherjee, D. Influence of Insecticides on Microbial Transformation of Nitrogen and Phosphorus in Typic Orchragualf Soil. J. Agric. Food Chem. 2000, 48, 3728–3732. [Google Scholar] [CrossRef]
- Zhu, G.; Wu, H.; Guo, J.; Kimaro, F.M.E. Microbial Degradation of Fipronil in Clay Loam Soil. Water. Air. Soil Pollut. 2004, 153, 35–44. [Google Scholar] [CrossRef]
- Shang, Y.; Hasan, M.; Ahammed, G.J.; Li, M.; Yin, H.; Zhou, J. Applications of nanotechnology in plant growth and crop protection: A review. Molecules 2019, 24, 2558. [Google Scholar] [CrossRef]
- Patnaik, G.K.; Kanungo, P.K.; Adhya, T.K.; Rajaramamohan Rao, V. Effect of repeated applications of gamma-hexachlorocyclohexane (γ-HCH) on nitrogenase activity and nitrogen-fixing bacteria associated with rhizosphere of tropical rice. Microbiol. Res. 1996, 151, 375–378. [Google Scholar] [CrossRef]
- Kalam, A.; Mukherjee, A.K. Influence of hexaconazole, carbofuran and ethion on soil microflora and dehydrogenase activities in soil and intact cell. Indian J. Exp. Biol. (IJEB) 2001, 39, 90–94. [Google Scholar]
- Singh, J.; Singh, D.K. Dehydrogenase and phosphomonoesterase activities in groundnut (Arachis hypogaea L.) field after diazinon, imidacloprid and lindane treatments. Chemosphere 2005, 60, 32–42. [Google Scholar] [CrossRef]
- Tu, C.M. Effect of five insecticides on microbial and enzymatic activities in sandy soil. J. Environ. Sci. Health Part B 1995, 30, 289–306. [Google Scholar] [CrossRef]
- El-Ghamry, A.M.; Xu, J.M.; Huang, C.Y.; Gan, J. Microbial response to bensulfuron-methyl treatment in soil. J. Agric. Food Chem. 2002, 50, 136–139. [Google Scholar] [CrossRef]
- Ingram, C.W.; Coyne, M.S.; Williams, D.W. Effects of Commercial Diazinon and Imidacloprid on Microbial Urease Activity in Soil and Sod. J. Environ. Qual. 2005, 34, 1573. [Google Scholar] [CrossRef]
- Colores, G.M.; Schmidt, S.K. Recovery of Microbially Mediated Processes in Soil Augmented with A Pentachlorophenol-Mineralizing Bacterium. Environ. Toxicol. Chem. 2005, 24, 1912. [Google Scholar] [CrossRef]
- Mayanglambam, T.; Vig, K.; Singh, D.K. Quinalphos Persistence and Leaching Under Field Conditions and Effects of Residues on Dehydrogenase and Alkaline Phosphomonoesterases Activities in Soil. Bull. Environ. Contam. Toxicol. 2005, 75, 1067–1076. [Google Scholar] [CrossRef]
- Qian, H.; Hu, B.; Wang, Z.; Xu, X.; Hong, T. Effects of validamycin on some enzymatic activities in soil. Environ. Monit. Assess. 2007, 125, 1–8. [Google Scholar] [CrossRef]
- Enserink, M.; Hines, P.J.; Vignieri, S.N.; Wigginton, N.S.; Yeston, J.S. The pesticide paradox. Science 2013, 341, 728–729. [Google Scholar] [CrossRef]
- Owen, M.D.K.; Beckie, H.J.; Leeson, J.Y.; Norsworthy, J.K.; Steckel, L.E. Integrated pest management and weed management in the United States and Canada. Pest Manag. Sci. 2015, 71, 357–376. [Google Scholar] [CrossRef]
- Lacey, L.A.; Georgis, R. Entomopathogenic nematodes for control of insect pests above and below ground with comments on commercial production. J. Nematol. 2012, 44, 218. [Google Scholar]
- Gupta, S.; Dikshit, A.K. Biopesticides: An ecofriendly approach for pest control. J. Biopestic. 2010, 3, 186. [Google Scholar]
- Meena, A.K.; Meena, A.K. Characterization and antagonistic effect of isolated Trichoderma sp. against pathogens under Clusterbean (Cyamopsis tetragonoloba L.). Indian J. Agric. Res. 2016, 50, 249–253. [Google Scholar] [CrossRef]
- Alam, G.; Alam, G. A Study of Biopesticides and Biofertilisers in Haryana, India; International Institute for Environment and Development: London, UK, 2000. [Google Scholar]
- Sharma, M.P.; Sharma, A.N.; Hussaini, S.S. Entomopathogenic nematodes, a potential microbial biopesticide: Mass production and commercialisation status – a mini review. Arch. Phytopathol. Plant Prot. 2011, 44, 855–870. [Google Scholar] [CrossRef]
- Neal, A.L.; Ahmad, S.; Gordon-Weeks, R.; Ton, J. Benzoxazinoids in Root Exudates of Maize Attract Pseudomonas putida to the Rhizosphere. PLoS ONE 2012, 7, e35498. [Google Scholar] [CrossRef]
- Akiyama, K.; Hayashi, H. Strigolactones: Chemical signals for fungal symbionts and parasitic weeds in plant roots. Ann. Bot. 2006, 97, 925–931. [Google Scholar] [CrossRef]
- Broughton, W.J.; Zhang, F.; Perret, X.; Staehelin, C. Signals exchanged between legumes and Rhizobium: Agricultural uses and perspectives. Plant Soil 2003, 252, 129–137. [Google Scholar] [CrossRef]
- Pathan, S.I.; Větrovský, T.; Giagnoni, L.; Datta, R.; Baldrian, P.; Nannipieri, P.; Renella, G. Microbial expression profiles in the rhizosphere of two maize lines differing in N use efficiency. Plant Soil 2018, 433, 401–413. [Google Scholar] [CrossRef]
- Gupta Sood, S. Chemotactic response of plant-growth-promoting bacteria towards roots of vesicular-arbuscular mycorrhizal tomato plants. FEMS Microbiol. Ecol. 2003, 45, 219–227. [Google Scholar] [CrossRef]
- Gopal, M.; Gupta, A.; Arunachalam, V.; Magu, S.P. Impact of azadirachtin, an insecticidal allelochemical from neem on soil microflora, enzyme and respiratory activities. Bioresour. Technol. 2007, 98, 3154–3158. [Google Scholar] [CrossRef]
- Govindachari, T.R.; Suresh, G.; Gopalakrishnan, G.; Masilamani, S.; Banumathi, B. Antifungal activity of some tetranortriterpenoids. Fitoterapia 2000, 71, 317–320. [Google Scholar] [CrossRef]
- Coventry, E.; Allan, E.J. Microbiological and Chemical Analysis of Neem (Azadirachta indica) Extracts: New Data on Antimicrobial Activity. Phytoparasitica 2001, 29, 441–450. [Google Scholar] [CrossRef]
- Elnasikh, M.H.; Osman, A.G.; Sherif, A.M. Impact of neem seed cake on soil microflora and some soil properties. J. Sc. Tech. 2011, 12, 144–150. [Google Scholar]
- Xu, X.; Chen, C.; Zhang, Z.; Sun, Z.; Chen, Y.; Jiang, J.; Shen, Z. The influence of environmental factors on communities of arbuscular mycorrhizal fungi associated with Chenopodium ambrosioides revealed by MiSeq sequencing investigation. Sci. Rep. 2017, 7, 45134. [Google Scholar] [CrossRef]
- Lu, F.C.; Lee, C.Y.; Wang, C.L. The influence of arbuscular mycorrhizal fungi inoculation on yam (Dioscorea spp.) tuber weights and secondary metabolite content. PeerJ 2015, 3, e1266. [Google Scholar] [CrossRef]
- Kiran, U.; Patra, D.D. Medicinal and aromatic plant materials as nitrification inhibitors for augmenting yield and nitrogen uptake of Japanese mint (Mentha arvensis L. Var. Piperascens). Bioresour. Technol. 2003, 86, 267–276. [Google Scholar] [CrossRef]
- Mohanty, S.; Patra, A.K.; Chhonkar, P.K. Neem (Azadirachta indica) seed kernel powder retards urease and nitrification activities in different soils at contrasting moisture and temperature regimes. Bioresour. Technol. 2008, 99, 894–899. [Google Scholar] [CrossRef]
- Medina, M.J.H.; Gagnon, H.; Piché, Y.; Ocampo, J.A.; Garrido, J.M.G.; Vierheilig, H. Root colonization by arbuscular mycorrhizal fungi is affected by the salicylic acid content of the plant. Plant Sci. 2003, 164, 993–998. [Google Scholar] [CrossRef]
- Tilak, K.; Ranganayaki, N.; Pal, K.; De, R.; Saxena, A.K.; Nautiyal, C.S.; Mittal, S.; Tripathi, A.K.; Johri, B.N. Diversity of plant growth and soil health supporting bacteria. Curr. Sci. 2005, 136–150. [Google Scholar]
- Sethi, S.; Gupta, S. Impact of Pesticides and Biopesticides on Soil Microbial Biomass Carbon. Univers. J. Environ. Res. Technol. 2013, 3, 2. [Google Scholar]
- Tripathi, A.K.; Mishra, S. Plant Monoterpenoids (Prospective Pesticides). In Ecofriendly Pest Management for Food Security; Academic Press: Cambridge, MA, USA, 2016; pp. 507–524. [Google Scholar]
- Tan, S.; Chen, X.; Li, D. Progress in the studies on Helicoverpa spp. resistance to transgenic Bt cotton and its management strategy. Kun Chong Xue Bao Acta Entomol. Sin. 2002, 45, 138–144. [Google Scholar]
- Sessitsch, A.; Kan, F.Y.; Pfeifer, U. Diversity and community structure of culturable Bacillus spp. populations in the rhizospheres of transgenic potatoes expressing the lytic peptide cecropin B. Appl. Soil Ecol. 2003, 22, 149–158. [Google Scholar] [CrossRef]
- Liu, B.; Zeng, Q.; Yan, F.; Xu, H.; Xu, C. Effects of transgenic plants on soil microorganisms. Plant Soil 2005, 271, 1–13. [Google Scholar] [CrossRef]
- Mishra, P.K.; Ekielski, A.; Mukherjee, S.; Sahu, S.; Chowdhury, S.; Mishra, M.; Talegaonkar, S.; Siddiqui, L.; Mishra, H. Wood-Based Cellulose Nanofibrils: Haemocompatibility and Impact on the Development and Behaviour of Drosophila melanogaster. Biomolecules 2019, 9, 363. [Google Scholar] [CrossRef]
- Heuer, H.; Kroppenstedt, R.M.; Lottmann, J.; Berg, G.; Smalla, K. Effects of T4 Lysozyme Release from Transgenic Potato Roots on Bacterial Rhizosphere Communities Are Negligible Relative to Natural Factors. Appl. Environ. Microbiol. 2002, 68, 1325–1335. [Google Scholar] [CrossRef]
- Schmalenberger, A.; Tebbe, C.C. Genetic profiling of noncultivated bacteria from the rhizospheres of sugar beet (Beta vulgaris) reveal field and annual variability but no effect of a transgenic herbicide resistance. Can. J. Microbiol. 2003, 49, 1–8. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Herbicides | Effects on Microorganism and Associated Process | References |
|---|---|---|
| 2,4-D | Adversely affects the activities of Rhizobium sp. | [17] |
| 2,4-D | Reduces nitrogenase, phosphatase and hydrogen photoproduction activities of purple non-sulfur bacteria | [36] |
| 2,4-D and 2,4,5-T | Adversely affects node-expression disrupting plant Rhizobium signalling. 2,4-D also reduces fixation by blue-green algae and nitrifying process impacting nitrosomonas and Nitrobacter sp. | [37] |
| 2,4-Damine, Agroxone, and Atranex | Inhibits activities of Rhizobium phaseoli and Azotobacter vinelandii (most sensitive) | [17] |
| 2,4-D, Bromoxynil, and Methomyl | Reduces CH4 oxidation to CO2 | [38] |
| Bensulfuron methyl and Metsulfuron-methyl | Decreases N-mineralization | [39] |
| Bentazone, Prometryn, Simazine, and Terbutryn | Inhibits N-fixation and decreases the number of nodules and N content overall | [40] |
| Isoproturon, Triclopyr | Adversely impacts nitrosomonas, Nitrobacter, urea hydrolyzing bacteria, nitrate reductase activity and growth of actinomycetes and fungi | [41] |
| Linuron, Terbutryn, and Methabenzthiazuron | Adversely impacts nitrogenase activity and nodulation at the pre-emergence application | [33] |
| Glyphosate | Suppresses phosphatase activity | [35] |
| Glyphosate | Reduces the growth and activity of azotobacter | [16] |
| Metribuzin | At lower doses, no effects on AM fungi in maize and barley are observed | [42] |
| Fungicides | Effects on Microorganism and Associated Process | References |
|---|---|---|
| Apron, Arrest, and Captan | Reduces viable counts of Rhizobium ciceri | [69] |
| Benomyl | Impacts mycorrhizal associations and nitrifying bacteria | [76] |
| Benomyl, Mancozeb | Arrests activity of dehydrogenase, urease, and phosphatase enzymes | [77] |
| Captan | Inhibits aerobic N-fixing, nitrifying, denitrifying bacteria, nitrogenase activity, phosphate solubilization and other fungi | [64] |
| Captan and Thiram | Decreases cell growth and nitrogenase activity in Azospirillum brasilense even at a lower dose of 10 mg/L | [78] |
| Captan and Carbendazim | Decreases nitrogenase enzyme activity | [36] |
| Captan, Carboxin, Thiram | Inhibits the activity of bacteria responsible for denitrification | [79] |
| Carbendazin and Thiram | Inhibits nodulation in legumes and thus N-fixation process | [80] |
| Chlorothalonil | Effects bacteria associated with nitrogen cycling | [76] |
| Chlorothalonil, Azoxystro | Effects biocontrol agent(s) used against Fusarium wilt | [71] |
| Copper fungicides | Decreases population of bacteria, cellulolytic fungal species and streptomycetes in sandy soil | [70] |
| Dimethomorph | Inhibits nitrification and ammonification process in sandy soils | [81] |
| Dinocap | Inhibits the activity of ammonifying bacteria | [82] |
| Dithianon | Destrucs bacterial diversity in soil | [83] |
| Fenpropimorph | Slows down bacterial activity in wetlands | [79] |
| Fludioxonil | Toxic to algal activities | [84] |
| Funaben, Baytan, Oxafun | Inhibits nitrogenase activity of methylotrophic bacteria at a higher dose | [85] |
| Hexaconazole | Impacts bacteria involved in N cycling | [86] |
| Mancozeb | Impacts on bacteria involved in N & C cycle in soil | [82] |
| Mancozeb, Chlorothalonil, Metal dithiocarbamates | Reduces nitrification process | [60] |
| Metalaxyl | Reduces urease activity continuously while phosphatase activity seems stimulated but then reduces | [87] |
| Metalaxyl | Disturbs activity of ammonifying and nitrifying bacteria | [19] |
| Oxytetracycline | Acts as bactericide | [88] |
| Pencycuron | Short-term impact on metabolically active soil bacteria | [89] |
| Propiconazole | May retard plant growth-promoting effects of Azospirillum brasilense on its host plant | [90] |
| Triadimefon | Deleterious to long-term soil bacterial community | [91] |
| Triarimol and Captan | Reduces frequency of Aspergillus sp. | [92] |
| Insecticide | Effects on Microorganism and Associated Process | References |
|---|---|---|
| Amitraz, Aztec, Cyfluthrin, Imidachlor, and Tebupirimphos | Reduces activities of urease and phosphatase enzymes for a week | [111] |
| Arsenic, DDT, and Lindane | Decreases microbial biomass and microbial and enzymatic activities as a result of longer persistence in soil | [110] |
| Bensulfuron methyl and Metsulfuron-methyl | Reduces soil microbial biomass | [112] |
| Carbamate insecticides | Inhibits several soil microorganisms, enzymes and nitrogenase activity of Azospirillum | [35,99] |
| Carbendazim, Imazetapir, Thiram | Decreases nitrogenase activity in Rhizobium leguminosarum. R. trifolii, Bradyrhizobium sp. and Sinorhizobium melilot in pot cultures as well as in field conditions | [80] |
| Carbofuran, Ethion, and Hexaconazole | Inhibits nitrogenase activity of Anabaena doliolum by 38% within 48 h of application | [109] |
| Chlorinated hydrocarbons | Inhibits methanogenesis | [61] |
| Chlorpyrifos, Dichlorvos, Phorate, Monocrotophos, Methyl parathion, Cypermethrin, Fenvalerate, Methomyl and Quinalphos | Increases phosphatase activity initially and later reduces gradually. Phorate reduces the total bacterial population and N-fixing bacteria | [67] |
| Chlorpyrifos, Profenofos, Pyrethrins, and Methylpyrimifos | Reduces the population of aerobic N-fixing, nitrifying, denitrifying bacteria and several fungi. Profenofos and Pyrethrins decreases the activity of urease enzyme and nitrate reductase | [94] |
| Chlorpyrifos, Quinalphos | Reduces ammonification process | [67] |
| Cyfluthrin, Fenpropimorph, and Imidacloprid | Decreases nitrification and denitrification process, and stimulates sulphur oxidation | [111] |
| Diazinon and Imidacloprid | Inhibits urease-producing bacterium (Proteus Vulgaris) | [113] |
| Lindane, Malathion, Diazinon, and Imidacloprid | Lindane inhibit state of nitrification, N-availability, P-solubilization and activity of phosphomonoesterase enzyme while the opposite effect is observed in the case of Diazinon and Imidacloprid | [110] |
| Metalaxyl and Mefenoxam | Decreases nitrogen-fixing bacteria and microbial biomass | [66] |
| Methamidophos | Reduces microbial biomass by 41%–83% | [13] |
| Neemix-4E | Reduces urease enzyme activity | [62] |
| Organophosphate insecticide | Impacts the activity of soil enzymes, several beneficial soil bacteria, and fungal population and reduces N-mineralization rate | [99] |
| Pentachlorophenol | Reduces nitrification | [114] |
| Quinalphos | Reduces activity of enzyme phosphomonoesterase which recovers later | [115] |
| Validamycin | Negatively effects phosphatase and urease enzyme which improves later | [116] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meena, R.S.; Kumar, S.; Datta, R.; Lal, R.; Vijayakumar, V.; Brtnicky, M.; Sharma, M.P.; Yadav, G.S.; Jhariya, M.K.; Jangir, C.K.; et al. Impact of Agrochemicals on Soil Microbiota and Management: A Review. Land 2020, 9, 34. https://doi.org/10.3390/land9020034
Meena RS, Kumar S, Datta R, Lal R, Vijayakumar V, Brtnicky M, Sharma MP, Yadav GS, Jhariya MK, Jangir CK, et al. Impact of Agrochemicals on Soil Microbiota and Management: A Review. Land. 2020; 9(2):34. https://doi.org/10.3390/land9020034
Chicago/Turabian StyleMeena, Ram Swaroop, Sandeep Kumar, Rahul Datta, Rattan Lal, Vinod Vijayakumar, Martin Brtnicky, Mahaveer Prasad Sharma, Gulab Singh Yadav, Manoj Kumar Jhariya, Chetan Kumar Jangir, and et al. 2020. "Impact of Agrochemicals on Soil Microbiota and Management: A Review" Land 9, no. 2: 34. https://doi.org/10.3390/land9020034
APA StyleMeena, R. S., Kumar, S., Datta, R., Lal, R., Vijayakumar, V., Brtnicky, M., Sharma, M. P., Yadav, G. S., Jhariya, M. K., Jangir, C. K., Pathan, S. I., Dokulilova, T., Pecina, V., & Marfo, T. D. (2020). Impact of Agrochemicals on Soil Microbiota and Management: A Review. Land, 9(2), 34. https://doi.org/10.3390/land9020034