Indirect Effects of Grazing on Wind-Dispersed Elm Seeds in Sparse Woodlands of Northern China


1
School of Life Science, Liaoning University, Shenyang 110036, China
2
Leibniz Institute of Agricultural Development in Transition Economies (IAMO), 06120 Halle (Saale), Germany
*
Author to whom correspondence should be addressed.
Land 2020, 9(12), 490; https://doi.org/10.3390/land9120490
Submission received: 28 October 2020
/
Revised: 25 November 2020
/
Accepted: 2 December 2020
/
Published: 3 December 2020
Abstract
:Grazing leads to the reduction of biomass and plays a critical role in land degradation in arid and semiarid lands. However, the indirect effects of grazing on the ecosystem, e.g., the effect on seed dispersal, have not been well understood. In this study, we built an agent-based model (ABM) to simulate how grazing intensity affects the seed dispersal of elm trees, one of the native vegetation species of temperate woodlands in semiarid lands. The simulated results from the ABM and observed data from the real world were compared to assess the accuracy and validity of the ABM. The results show that elm seed densities in non-grazing, light, moderate, and heavy grazing lands were 74.97 ± 1.44, 57.63 ± 0.89, 37.73 ± 0.95, and 0.97 ± 0.05 seeds m−2, respectively—an apparently decreasing trend. Moreover, as grazing intensity increased, the values of nugget, sill, and partial sill decreased and the value of the ratio of nugget to sill increased. This study indicates that the grazing indirectly leads to the reduction of elm seed density and the increase of spatial heterogeneity of elm seed on the ground in sparse elm woodlands. Moreover, values of geostatistical indices from the ABM were not significantly different from field observation data except for the ratio of nugget to sill. It shows that ABMs can reasonably replicate the spatial pattern of elm seed densities in the field and thus are useful for simulating long-distance seed dispersal in sandy lands. This finding suggests that the indirect effects of grazing should be considered to effectively protect sparse elm woodlands.
1. Introduction
Grazing influences land use in arid and semiarid lands in various ways, ranging from changing vegetation structure to inducing land degradation [1,2]. Grazing influences vegetation by modifying individual growth, varying species abundance, and changing population dynamics [3]. Moreover, grazing regulates the vegetation spatial pattern by changing the spatial heterogeneity of soil nutrients and influencing both intra- and interspecies interaction [4,5,6,7,8,9]. However, the effects of grazing on seed dispersal have rarely been considered. It is vital to obtain a thorough understanding of the effects of grazing on seed dispersal. This could help us protect the ecosystem and combat land degradation in arid and semiarid lands.
Seed dispersal is a vital process linking the reproductive cycle of plants with the establishment of their offspring. Thus, it has various implications for species population and ecosystem biology, especially in arid and semiarid lands where winds primarily drive seeds to shape the pattern of seed dispersal and mainly determine the spatial structure of plant populations [10,11]. At the same time, the seed dispersal process is highly stochastic, and the underlying mechanism is notoriously challenging to quantify. In the current literature, field experiments and modeling approaches have been conducted to explore the influencing factors of seed dispersal [12,13].
The seed dispersal process might be influenced by grazing in direct and indirect ways. Directly, grazing promotes more seed production compared with exclosure lands [14,15]. Indirectly, the grazing animals eat and trample vegetation. Furthermore, vegetation cover regulates seed survival on the ground [16]. Thus, we hypothesize that grazing intensities regulate seed density patterns by changing vegetation coverage.
The effects of grazing on seed dispersal have not been well understood. This might be due to two reasons. One is that the effects of grazing on seed dispersal are difficult to test with field experiments. Firstly, the trace of seed dispersal, especially long-distance, is not easy to follow [12]. Secondly, heterogeneous patches of vegetation cover could cause nonuniform samples in designed experiments [17,18]. Additionally, the effects of grazing on seed dispersal in arid and semiarid lands are commonly intertwined with the wind, the main factor regulating seed dispersal [19,20,21]. This makes disentangling various driving factors extremely difficult.
Given the challenges and obstacles in field experiments, spatial simulating models have gained increasing attention in ecological modeling for their capability to incorporate spatial and temporal aspects of environmental factors and the dynamics of plants and animals [22]; in this case, the effects of grazing and wind. Agent-based models (ABMs), also known as individual-based models in the ecological modeling field, are spatially explicit models that take into account aspects of agents and their interactions, as well as heterogeneous environmental factors [23,24,25,26]. While ABMs are mostly used in simulating animal behaviors and patterns [27,28,29], recently, ABMs have been increasingly used for plant ecology; for example, in simulating species distribution [30], tree regeneration patterns [31], plant extensions [32], interspecies competition [33], and seeds dispersal [24,34]. However, the combination of ABMs and grazing has rarely been reported, especially for arid and semiarid regions, where grazing is the main driving force regulating vegetation and land-use type.
In this research, we built an ABM simulating seed dispersal that is associated with vegetation cover, grazing, and wind. In this study, the main aim is to explore the indirect effects of grazing on seed dispersal. Knowing the indirect effects of grazing on seed dispersal is vital for predicting the spatial pattern of seed distribution and evaluating the probability of recruitment in lands. The specific objectives are (1) to test our hypothesis, provided here, and (2) to verify the feasibility of the ABM on sparse woodlands. For that, firstly, we built an ABM to simulate seed dispersal under various wind frequencies and directions. Secondly, we compared the simulated results with the experimental results observed in the field. Thirdly, we used the ABM to evaluate the effects of grazing under various scenarios that consider the vegetation cover as heterogeneous environments.
2. Materials and Methods
2.1. Study Area
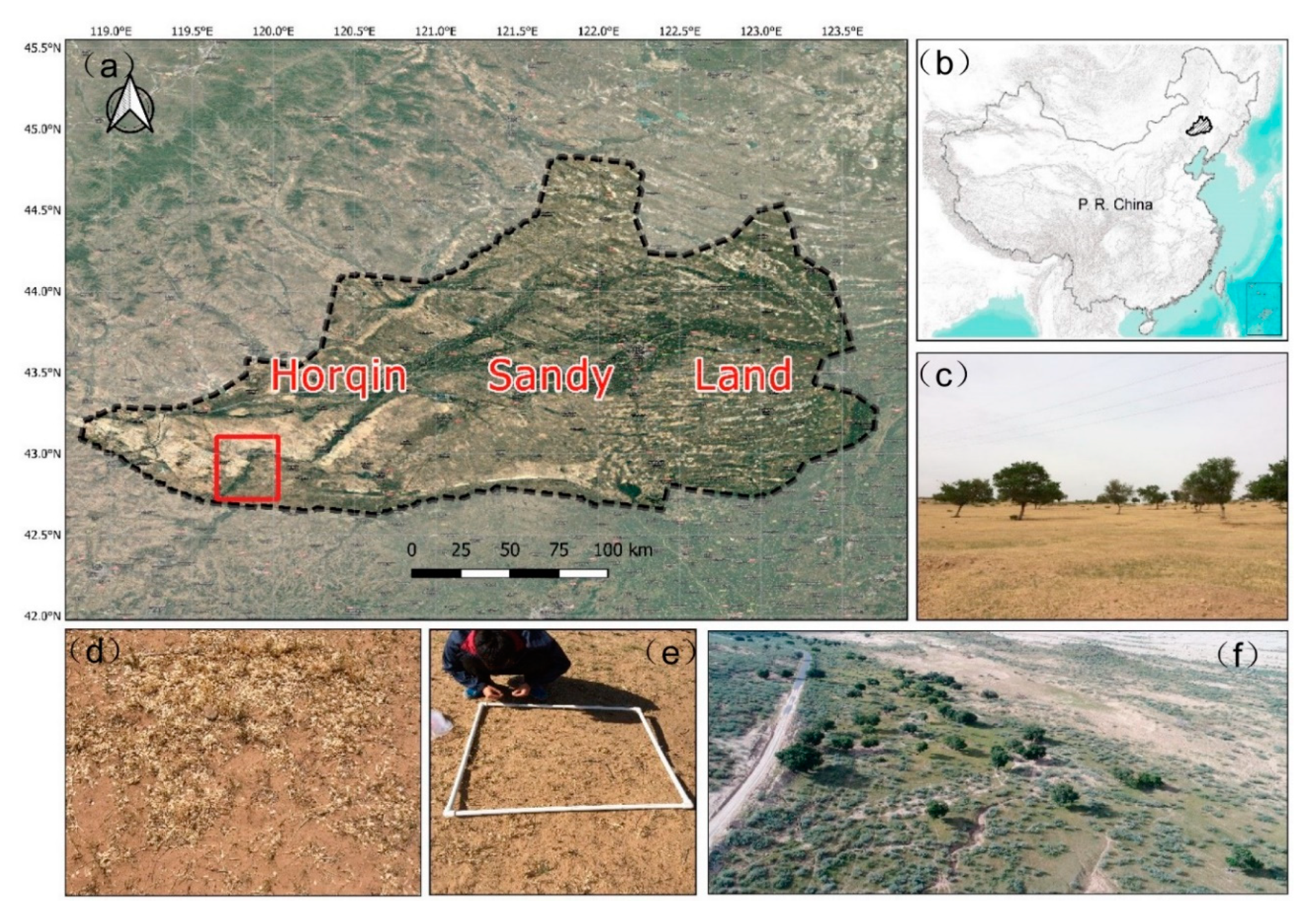
Horqin Sandy Land is located in a typical agriculture and pasture interlaced zone on the eastern edge of the Mongolian Plateau [35] (Figure 1). Horqin Sandy Land, with an area of 5.23 × 104 km2, is one of the largest sandy lands in China [36]. Due to the sandy and loose soil, sparse vegetation cover, and its semiarid climate condition, this area is highly prone to wind erosion and desertification. Exacerbated by ever-intensifying human activities such as livestock grazing and agricultural expansion and climate changes (e.g., the decreasing precipitation) in the past years, the risk of desertification and land degradation remains very high [37,38]. The continuous land degradation of Horqin Sandy Land attracts much attention from researchers and policymakers. A series of national ecological restoration projects, such as the Grain for Green Project and the Beijing and Tianjin Sandstorm Source Treatment Project, was implemented to fight for land degradation and preserve the fragile ecosystems [35].
We mainly focused on the Wulanaodu area of Horqin Sandy Land (119°39′–120°02′ E, 42°29′–43°06′ N, 480 m a.s.l.). This area is characterized by a continental semiarid monsoon climate. The average annual temperature is 6.3 °C, with July being the warmest month, averaging 23.0 °C, and January being the coldest month, averaging −14.0 °C. The mean annual precipitation is 340 mm, of which 70% falls in June, July, and August. The mean annual wind velocity is 4.4 m s−1, and the number of gale days (>16 m s−1) ranges between 21 and 80. The windy season is from March to May, and the growing season begins in late April and ends in late September. Typical plants are Caragana microphylla Lam., Setaria viridis (L.) Beauv., Bassia dasyphylla, Chenopodium glaucum Linn., Chenopodium aristatum Linn., Lespedeza daurica (Laxm.) Schindl., and Pennisetum centrasiaticum Tzvel [6].
Sparse woodland is the native vegetation type and climax community in Horqin Sandy Land [14]. It plays an essential role in reducing wind erosion, promoting ecological stability, and supplying ecological services [14,39]. Sparse woodland constitutes three layers, i.e., trees, shrubs, and herbs. Additionally, Ulmus pumila L. (elm trees) is the key and defining species in sparse woodland. Elm trees provide various ecosystem services and play critical roles in helping to maintain the stability of the ecosystem of sparse woodland. For example, elm trees provide a seed shadow for regeneration and shade for grasses and livestock [15,16].
Sparse woodlands and elm trees, in particular, are, however, threatened by ever-increasing grazing pressure. Grazing leads to a decrease in the area of sparse woodland [14]. In addition, elm trees are also adversely influenced by grazing in the aspects of seedling growth, reproduction allocation, and spatial distribution [6]. More interestingly, grazing also changes vegetation coverage and subsequently influences elm seed dispersal, which plays a vital role in the renewal of elm trees in the wind season [40].
2.2. The Agent-Based Model
The ABM here was developed using NetLogo, a free software platform for building ABMs [41,42]. The model description largely follows the ODD protocol, i.e., overview, design concepts, and details [43]. More details can be found in Supplementary Material. The purpose of the model is (1) to simulate the seed dispersal of elm trees with wind directions in Horqin Sandy Land and (2) to evaluate the effects of grazing on seed dispersal under scenarios of grazing intensities, i.e., light, moderate, and heavy. The model framework is illustrated in Figure 2.
We built the model with two entities, seeds, and vegetation cover. The seeds were modeled as agents, and their movements were driven by winds. Wind directions controlled the direction of the movement of seeds. The seed locations, quantified in a 2D coordinate system, i.e., x- and y- coordinates, changed depending on the wind, with eight directions. We recorded the locations of seeds at each moment and calculated the distance from the original point (x = 0, y = 0). The vegetation cover was modeled as patches within a regular bidimensional lattice. Vegetation coverage, defined as the percentage of the ground surface covered by herbs and shrubs, was assigned as an attribute of each patch, a grid cell. The relationship between vegetation coverage and seed density followed an experimental equation in the same study area [16], i.e., b = −0.131a2 + 9.527a − 30.53 (R2 = 0.446). Here, b indicates seed densities, and a indicates vegetation coverage.
The model was initialized with several parameters, including seed number, frequency of the eight wind directions, and vegetation coverage. The value of these parameters, except vegetation coverage, was taken from a previous study [40]. The value of vegetation coverage was randomly designed and varied within a range of 0–30%. We conducted the sensitivity analysis on the total number of patches and running time steps of the model, which might influence the output of the model.
The effects of grazing on seed dispersal were focused on changes in vegetation cover. Vegetation cover has a significant relationship with seed densities [16]. Meanwhile, grazing influences vegetation cover by various degrees, according to grazing intensities [44]. Thus, we could evaluate the effects of grazing on seed dispersal through changes in vegetation cover. The changes in vegetation cover reflect the changing grazing intensities under various scenarios.
2.3. Sensitivity Analysis
A sensitivity analysis is used to determine which parameters have more significant impacts on a model and whether the model has stability [45,46]. The high and low values of parameter sensitivity indices indicate whether a parameter change has a great or little effect on the model, respectively. The changes in the sensitivity index could be the criteria to set the optimal values of particular parameters. The sensitivity index (Syi) is expressed as ([47,48])
In Equation (1), Xt and Yt are the input parameters and output of the model at time t; dXt and dYt are the changes in Xt and Yt. The output of this model is the seed density. The average sensitivity index (S) is calculated as Equation (2), where n is the number of model repetitions.
A small value of the sensitivity index of a particular parameter indicates that a change in the values of this parameter has little effect on the model. Conversely, the relatively large value of the sensitivity index of a particular parameter indicates that a change in the values of this parameter has significant effects on the model. We select the relatively small values of the sensitivity index. Here, the running time steps are set to 40.
2.4. The Field Investigation
At the end of the seed rain period, we randomly selected four plots (10 × 10 m) in enclosed sandy grasslands in our study area. For these plots, the dominant species was Chloris virgata Swartz, Setaria viridis (L.) Beauv., and Corispermum hyssopifolium L. The vegetation coverage in the four plots was 10%, 30%, 10%, and less than 5%, respectively (the vegetation coverage was relatively low because of decreased precipitation). In each plot, we counted the number of elm seeds in 100 subplots (1 × 1 m) separately. According to seed densities in the subplots, the geostatistical indices of the spatial patterns of seed dispersal were estimated.
2.5. Model Validation
How successfully the model-simulated results replicate the system behaviors of the real world, i.e., observations on the ground, validates the ABM; this is critical to assessing the credibility of an ABM [49]. We used an ABM to model the seed dispersal process by simulating the movement of individual seeds. However, the ultimate goal of the model is to understand the macro patterns of the targeted system, in this case, the spatial distribution patterns of the elm seeds. Thus, we used the pattern-oriented modeling approach (POM) to validate the simulated outputs of our ABM [50,51]. Given the abstract nature of ecological modeling, with a loss of information during the modeling process due to simplification and abstraction, POM focuses on a few relevant patterns in the real system. Patterns are macro-level characteristics of a complex system and often emerged from the essential underlying processes. Thus, POM is widely used to validate an ABM by testing how well an ABM can replicate system-level patterns. In other words, if an ABM can reasonably reproduce structurally realistic patterns, it is more likely that the model catches the key dynamics in the modeled system. As there were many potential patterns for the seed dispersal process (e.g., the maximum or average distance a seed travels, the shape of seed distribution of a single elm tree, density distribution of seeds), we here focused on the spatial patterns of seed dispersal and chose geostatistical indices as our pattern.
The results of the spatial pattern of seed densities were calculated with geostatistical methods and presented with geostatistical indices, including nugget, partial sill, sill, and the ratio of nugget to sill (see the data analysis section). Meanwhile, we obtained these geostatistical indices from the results of the field investigation, and we compared the geostatistical indices from the field investigation to the simulated results. The values of the geostatistical indices from the simulations fell into the interval (mean ± 1 standard error) of corresponding values from the investigated results, indicating that the model’s results are consistent with the data observed on the ground.
2.6. Scenario Analysis
To further explore the effects of grazing on elm seed dispersal under intensity scenarios, we simulated the effects of grazing on seed dispersal with different intensities, including heavy grazing (HG), moderate grazing (MG), and light grazing (LG). The loss of vegetation cover caused by HG, MG, and LG was 85.90%, 45.90%, and 23.70%, respectively, when HG, MG, and LG was designed as six sheep ha−1, four sheep ha−1, and two sheep ha−1 in Horqin Sandy Land (fine wool sheep, 50 kg on average, 2-year-old) [44]. The values of loss in vegetation caused by grazing intensities were set as parameters in this model under different scenarios. The different scenarios of grazing intensities on seed dispersal could be simulated with the ABM.
2.7. Data Analysis
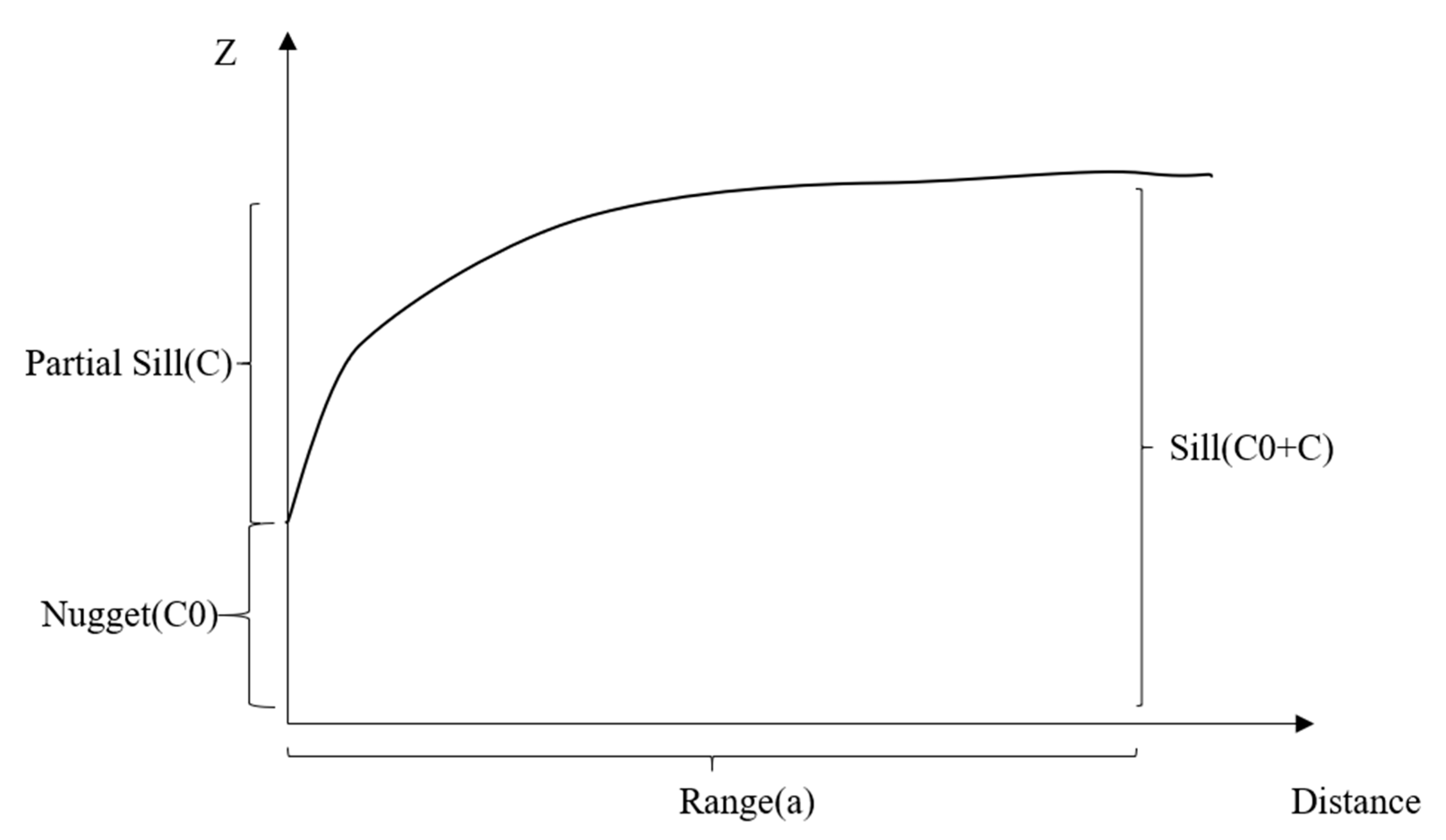
Geostatistical indices, including nugget, partial sill, sill, and the ratio of nugget to sill, were calculated with geostatistical models (Figure 2). In geostatistical models, one core concept is the variogram. A variogram characterizes the spatial continuity or roughness of a data set [52]. For each variable (Z), the variogram (γ) can be calculated from all sample pairs with lag distance h between locations (s) as
The variogram is defined by the following parameters: nugget (C0), partial sill (C), sill (C0 + C), and range (a). These parameters perform differently in the spherical model, the Gaussian model, and the exponential model. These models are determined by the equations above. We used the geoR package in R to perform the geostatistical analysis [53]. The difference of mean in the spatial indices in this model and the field investigation data was determined with a t-test. The difference between grazing intensities was investigated with ANOVA and Turkey’s HSD test. p-values less than 0.05 were considered significant in this study.
3. Results
3.1. Model Validation
According to the results of the field investigation, the values of nugget, partial sill, sill, and the ratio of nugget to sill in the plots were 127.93 ± 118.18, 241.05 ± 216.30, 368.97 ± 334.41, and 30.26 ± 10.70 m, respectively. Meanwhile, the outputs of this model showed that values of nugget, partial sill, sill, and the ratio of nugget to sill were 39.57 ± 4.03, 38.49 ± 4.46, 78.06 ± 2.56, and 50.99 ± 0.05 m. The difference of mean in nugget, partial sill, and sill was not significant between the results of the field investigation and this model (p > 0.05, Table 1). The difference of mean in the ratio of nugget to sill was significant between the results of the field investigation and this model (p < 0.05; Table 1).
3.2. Effects of Grazing on Seeds Dispersal
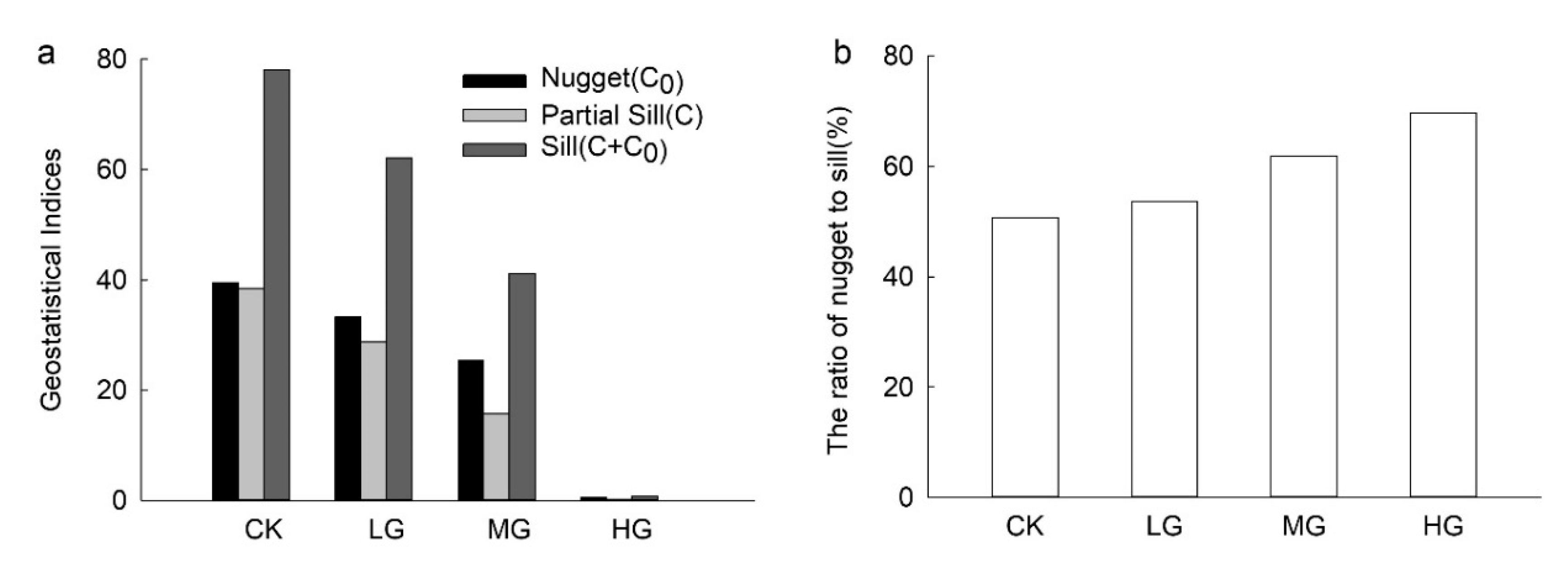
The elm seed densities in light, moderate, and heavy grazing lands were 57.63 ± 0.89, 37.73 ± 0.95, and 0.97 ± 0.05 seeds m−2, respectively. Meanwhile, the elm seed density in lands without grazing was 74.97 ± 1.44 seeds m−2. The difference in seed densities between grazing intensities was significant (p < 0.05).
The values of nugget, partial sill, and sill in the heavy grazing treatment were far less than in the other three treatments (Figure 3a). The values of nugget, partial sill, and sill decreased along with the increase in grazing intensities (Figure 3a). Meanwhile, the values of the ratio of nugget to sill increased along with the increase in grazing intensities (Figure 3b). The values of nugget, partial sill, and sill were significantly different between heavy grazing and other intensities (p < 0.05).
4. Discussion
With an increase in grazing intensity, seed densities decreased from 58 to 1 seed m−2, indicating that grazing is likely to reduce the seed density of elm trees in sandy lands. This finding supports our hypothesis that grazing intensities regulate seed densities. However, this result is not consistent with a previous study, where the seed density in grazing lands was more abundant than in exclosure lands [14]. In the previous study, only the direct effects of grazing on elm seed production were considered, as seed density was investigated in the seed rain period; however, the indirect effects of grazing on seed production were not considered. Different from the previous study, we explored the indirect effects of grazing on seed production, i.e., the effects of grazing on seed dispersal through the change in vegetation coverage. This study found that the indirect effects of grazing play a vital role in seed dispersal. Vegetation coverage influences elm seed densities because it regulates seed dispersal by wind [16]. Grazing mainly reduces vegetation cover through excessive foraging and trampling. Thus, these studies suggest the effects of grazing on the elm seed dispersal are more complex than what we thought and have an impact through direct and indirect ways.
With an increase in grazing intensity, values of nugget, sill, and partial sill decreased, but the value of the ratio of nugget to sill increased, indicating that grazing causes an increase in the spatial heterogeneity of seed densities. This finding confirms our hypothesis that grazing indirectly influences seed density patterns by regulating grass vegetation coverage on the ground. Moreover, this is also in line with the findings from previous studies [8,16,54]. This study emphasizes the intermediary role of vegetation cover in linking grazing and seed dispersal. Moreover, the spatial heterogeneity of seed densities changed by the grazing may intensify the spatial heterogeneity of the vegetation as seed dispersal provides seed sources for vegetation restoration [55].
The values of geostatistical indices from the ABM were not significantly different from the investigated results, except for one index (the ratio of nugget to sill). It is suggested that the model represents some of the key processes of the seed dispersal, and, consequently, the results of the simulation are consistent with the field observations. It shows that ABMs are useful for simulating the long-distance seed dispersal in sandy lands—a daunting task faced by researchers due to its highly stochastic nature. Long-distance dispersal is relatively difficult to trace in experimental studies [12]. Meanwhile, existing simulated models depend on various environmental and vegetation factors (including wind velocity, seed release height, and vegetation height) and their prior distributions, which can be difficult to parameterize [13]. Therefore, ABMs have their advantages when simulating seed dispersal as seed movements are modeled with simple rules [34].
The study also shows that ABMs can reasonably replicate the observations on the ground, that is, the spatial pattern of elm seed densities, which has profound implications on elm seed germination, seedling growth, and sapling growth. In this ABM, seed dispersal is mainly driven by winds and is influenced by vegetation cover. To keep the parsimony of the model, only a few key factors such as the direction and frequency of wind, vegetation cover, and grazing intensities are included. Factors not considered in this research, such as wind power, slope, and animal predation, may be considered in future research [56]. However, we believe the agent-based models should not include unnecessary complications, which hinder model output analysis and the complexity represented in the model [57]. We believe that the model designed in this study strikes a balance with its parsimony and its capability to replicate empirical observations.
5. Conclusions
The results show that a decrease of vegetation cover (mainly grass cover), as an indirect effect of grazing, has a negative effect on the elm seed dispersal; in other words, grazing causes a decrease of seed density on the ground. Alternatively, grazing leads to an increase in spatial heterogeneity in sparse elm woodlands. This finding supports our hypotheses and suggests that the indirect effects of grazing should be fully considered to protect sparse elm woodlands, which is the original vegetation type of temperate woodlands in semiarid lands in this region.
This research demonstrates that ABMs are a promising approach in studying the seed dispersal process, e.g., with their capability to fully consider plant and environmental factors at the same time. Such ABM-based seed dispersal models can help us reveal the key dynamics in the complex process of vegetation cover and land degradation in sandy lands.
Supplementary Materials
The following are available online at https://www.mdpi.com/2073-445X/9/12/490/s1, Table S1: The state variables of entities, Table S2: Input data of this seed dispersal model.
Author Contributions
Conceptualization, Y.T. and M.L.; methodology, Z.S., Y.T., and M.L.; software, M.L.; validation, M.L. and Y.T.; investigation, M.L. and Y.T.; writing—original draft preparation, Y.T. and M.L.; writing—review and editing, Z.S. and Y.T.; visualization, M.L.; supervision, Z.S.; All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Nature Science Foundation of China (31870709).
Acknowledgments
We thank Peter Diggle for guiding us in the use of the geoR package.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Miri, A.; Dragovich, D.; Dong, Z. Wind-borne sand mass flux in vegetated surfaces—Wind tunnel experiments with live plants. Catena 2019, 172, 421–434. [Google Scholar] [CrossRef]
- Van de Koppel, J.; Rietkerk, M.; van Langevelde, F.; Kumar, L.; Klausmeier, C.A.; Fryxell, J.M.; Hearne, J.W.; van Andel, J.; de Ridder, N.; Skidmore, A.; et al. Spatial heterogeneity and irreversible vegetation change in semiarid grazing systems. Am. Nat. 2002, 159, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Pande, T.N.; Yamamoto, H. Cattle treading effects on plant growth and soil stability in the mountain grassland of Japan. Land Degrad. Dev. 2006, 17, 419–428. [Google Scholar] [CrossRef]
- Saiz, H.; Alados, C.L. Changes in Semi-arid plant species associations along a livestock grazing gradient. PLoS ONE 2012, 7, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, H.W.; Bai, Y.F.; Hooper, D.U.; Schonbach, P.; Gierus, M.; Schiborra, A.; Taube, F. Selective grazing and seasonal precipitation play key roles in shaping plant community structure of semi-arid grasslands. Landsc. Ecol. 2015, 30, 1767–1782. [Google Scholar] [CrossRef]
- Zhang, M.; Wu, J.; Tang, Y. The effects of grazing on the spatial pattern of elm (Ulmus pumila L.) in the sparse woodland steppe of Horqin Sandy Land in northeastern China. Solid Earth 2016, 7, 631–637. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Song, X.X.; Wang, L.; Wang, D.L.; Zhou, X.M.; Liu, J.; Zhao, X.; Li, J.; Lin, H.J. Effects of grazing on soil nitrogen spatial heterogeneity depend on herbivore assemblage and pre-grazing plant diversity. J. Appl. Ecol. 2016, 53, 242–250. [Google Scholar] [CrossRef]
- Munyati, C. Spatial variations in plant nutrient concentrations in tissue of a grass species as influenced by grazing intensity in a confined savannah rangeland. J. Arid Environ. 2018, 155, 46–58. [Google Scholar] [CrossRef]
- Deleglise, C.; Loucougaray, G.; Alard, D. Spatial patterns of species and plant traits in response to 20 years of grazing exclusion in subalpine grassland communities. J. Veg. Sci. 2011, 22, 402–413. [Google Scholar] [CrossRef]
- Miri, A.; Dragovich, D.; Dong, Z. Vegetation morphologic and aerodynamic characteristics reduce aeolian erosion. Sci. Rep. 2017, 7, 1–9. [Google Scholar] [CrossRef]
- Wright, S.J.; Trakhtenbrot, A.; Bohrer, G.; Detto, M.; Katul, G.G.; Horvitz, N.; Muller-Landau, H.C.; Jones, F.A.; Nathan, R. Understanding strategies for seed dispersal by wind under contrasting atmospheric conditions. Proc. Natl. Acad. Sci. USA 2008, 105, 19084–19089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bullock, J.M.; Clarke, R.T. Long distance seed dispersal by wind: Measuring and modelling the tail of the curve. Oecologia 2000, 124, 506–521. [Google Scholar] [CrossRef] [PubMed]
- Soons, M.B.; Heil, G.W.; Nathan, R.; Katul, G.G. Determinants of long-distance seed dispersal by wind in grasslands. Ecology 2004, 85, 3056–3068. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.; Jiang, D.; Lu, X. Effects of Exclosure Management on Elm (Ulmus pumila) Recruitment in Horqin Sandy Land, Northeastern China. Arid Land Res. Manag. 2014, 28, 109–117. [Google Scholar] [CrossRef]
- Tang, Y. Impact of grazing intensities on reproduction patterns of elm trees (Ulmus pumila) in degraded sandy lands in China. PeerJ 2020, 8. [Google Scholar] [CrossRef] [Green Version]
- Jiang, D.; Tang, Y.; Busso, C.A. Effects of vegetation cover on recruitment of Ulmus pumila L. in Horqin Sandy Land, northeastern China. J. Arid Land 2014, 6, 343–351. [Google Scholar] [CrossRef] [Green Version]
- Zuo, X.; Zhao, H.; Zhao, X.; Zhang, T.; Guo, Y.; Wang, S.; Drake, S. Spatial pattern and heterogeneity of soil properties in sand dunes under grazing and restoration in Horqin Sandy Land, Northern China. Soil Till. Res. 2008, 99, 202–212. [Google Scholar] [CrossRef]
- Tayyebi, A.; Jenerette, G.D. Increases in the climate change adaption effectiveness and availability of vegetation across a coastal to desert climate gradient inmetropolitan Los Angeles, CA, USA. Sci. Total Environ. 2016, 548, 60–71. [Google Scholar] [CrossRef]
- Nathan, R.; Safriel, U.N.; Noy-Meir, I.; Schiller, G. Spatiotemporal variation in seed dispersal and recruitment near and far from Pinus halepensis trees. Ecology 2000, 81, 2156–2169. [Google Scholar] [CrossRef]
- Xiao, Z.S.; Zhang, Z.B.; Wang, Y.S. Effects of seed size on dispersal distance in five rodent-dispersed fagaceous species. Acta Oecol. 2005, 28, 221–229. [Google Scholar] [CrossRef]
- Yang, Y.; Bai, Y.; Li, J. Seed dispersal and seedling recruitment of Ulmus pumila woodland in the Keerqin Sandy Land, China. Acta Ecol. Sin. 2013, 33, 374–381. [Google Scholar] [CrossRef] [Green Version]
- Wallentin, G. Spatial simulation: A spatial perspective on individual-based ecology—A review. Ecol. Model. 2017, 350, 30–41. [Google Scholar] [CrossRef]
- David, V.; Joachim, S.; Tebby, C.; Porcher, J.-M.; Beaudouin, R. Modelling population dynamics in mesocosms using an individual-based model coupled to a bioenergetics model. Ecol. Model. 2019, 398, 55–66. [Google Scholar] [CrossRef]
- Fedriani, J.M.; Wiegand, T.; Ayllon, D.; Palomares, F.; Suarez-Esteban, A.; Grimm, V. Assisting seed dispersers to restore oldfields: An individual-based model of the interactions among badgers, foxes and Iberian pear trees. J. Appl. Ecol. 2018, 55, 600–611. [Google Scholar] [CrossRef]
- Grimm, V. Ten years of individual-based modelling in ecology: What have we learned and what could we learn in the future? Ecol. Model. 1999, 115, 129–148. [Google Scholar] [CrossRef]
- Grimm, V.; Railsback., S.F. Individual-Based Modeling and Ecology; Princeton University Press: Princeton, NJ, USA, 2005. [Google Scholar]
- Tang, W.; Bennett, D.A. Agent-based Modeling of Animal Movement: A Review. Geogr. Compass 2010, 4, 682–700. [Google Scholar] [CrossRef]
- McLane, A.J.; Semeniuk, C.; McDermid, G.J.; Marceau, D.J. The role of agent-based models in wildlife ecology and management. Ecol. Model. 2011, 222, 1544–1556. [Google Scholar] [CrossRef]
- Loraamm, R.W. Incorporating behavior into animal movement modeling: A constrained agent-based model for estimating visit probabilities in space-time prisms. Int. J. Geogr. Inf. Sci. 2020, 34, 1607–1627. [Google Scholar] [CrossRef]
- Lin, S.; DeVisser, M.H.; Messina, J.P. An agent-based model to simulate tsetse fly distribution and control techniques: A case study in Nguruman, Kenya. Ecol. Model. 2015, 314, 80–89. [Google Scholar] [CrossRef] [Green Version]
- Wallentin, G.; Tappeiner, U.; Strobl, J.; Tasser, E. Understanding alpine tree line dynamics: An individual-based model. Ecol. Model. 2008, 218, 235–246. [Google Scholar] [CrossRef]
- Wong, S.; Anand, M.; Bauch, C.T. Agent-based modelling of clonal plant propagation across space: Recapturing fairy rings, power laws and other phenomena. Ecol. Inform. 2011, 6, 127–135. [Google Scholar] [CrossRef] [Green Version]
- Bittebiere, A.K.; Mony, C.; Clement, B.; Garbey, M. Modeling competition between plants using an Individual Based Model: Methods and effects on the growth of two species with contrasted growth forms. Ecol. Model. 2012, 234, 38–50. [Google Scholar] [CrossRef]
- Bialozyt, R.; Flinkerbusch, S.; Niggemann, M.; Heymann, E.W. Predicting the seed shadows of a Neotropical tree species dispersed by primates using an agent-based model with internal decision making for movements. Ecol. Model. 2014, 278, 74–84. [Google Scholar] [CrossRef]
- Li, J.; Xu, B.; Yang, X.; Qin, Z.; Zhao, L.; Jin, Y.; Zhao, F.; Guo, J. Historical grassland desertification changes in the Horqin Sandy Land, Northern China (1985–2013). Sci. Rep. 2017, 7, 3009. [Google Scholar] [CrossRef] [Green Version]
- Yan, Y.; Zhu, J.; Yan, Q.; Zheng, X.; Song, L. Demarcation of the Horqin Sandy Land Boundary Based on Remote Sensing and GIS Technique. Sci. Geogr. Sin. 2014, 34, 122–128. [Google Scholar]
- Ge, X.; Dong, K.; Luloff, A.; Wang, L.; Xiao, J. Impact of land use intensity on sandy desertification: An evidence from Horqin Sandy Land, China. Ecol. Indic. 2016, 61, 346–358. [Google Scholar] [CrossRef]
- Liu, X.; He, Y.; Zhang, T.; Zhao, X.; Li, Y.; Zhang, L.; Wei, S.; Yun, J.; Yue, X. The response of infiltration depth, evaporation, and soil water replenishment to rainfall in mobile dunes in the Horqin Sandy Land, Northern China. Environ. Earth Sci. 2015, 73, 8699–8708. [Google Scholar] [CrossRef]
- Yang, J.; Tang, Y. The increase in ecosystem services values of the sand dune succession in northeastern China. Heliyon 2019, 5, e02243. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Tang, Y. The simulation in wind-driven seed dispersal of Ulmus pumila in sparse woodland steppe. Chin. J. Ecol. 2018, 37, 2524–2531. [Google Scholar]
- Railsback, S.; Ayllon, D.; Berger, U.; Grimm, V.; Lytinen, S.; Sheppard, C.; Thiele, J. Improving Execution Speed of Models Implemented in NetLogo. JASSS J. Artif. Soc. Simul. 2017, 20. [Google Scholar] [CrossRef] [Green Version]
- Wilensky, U. NetLogo. Center for Connected Learning and Computer-Based Modeling; Northwestern University: Evanston, IL, USA, 1999. [Google Scholar]
- Grimm, V.; Berger, U.; DeAngelis, D.L.; Polhill, J.G.; Giske, J.; Railsback, S.F. The ODD protocol A review and first update. Ecol. Model. 2010, 221, 2760–2768. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Zhang, T.; Toshiya, O.; Yasuhito, S. Soil Characteristics and Spatial Pattern of Vegetation after Successive Grazing in Horqin Sandy Land, Inner Mongolia. J. Arid Land Resour. Environ. 2003, 17, 117–121. [Google Scholar]
- Thiele, J.C.; Kurth, W.; Grimm, V. Facilitating Parameter Estimation and Sensitivity Analysis of Agent-Based Models: A Cookbook Using Net Logo and R. JASSS J. Artif. Soc. Simul. 2014, 17. [Google Scholar] [CrossRef]
- Snell, R.S. Simulating long-distance seed dispersal in a dynamic vegetation model. Glob. Ecol. Biogeogr. 2014, 23, 89–98. [Google Scholar] [CrossRef]
- Tang, Y.; Li, X. Simulating effects of precipitation and initial planting density on population size of Mongolian pine in the Horqin Sandy Land, China. Agrofor. Syst. 2018, 92, 1–9. [Google Scholar] [CrossRef]
- Wei, S.K.; Yang, H.; Song, J.X.; Abbaspour, K.C.; Xu, Z.X. System dynamics simulation model for assessing socio-economic impacts of different levels of environmental flow allocation in the Weihe River Basin, China. Eur. J. Oper. Res. 2012, 221, 248–262. [Google Scholar] [CrossRef]
- North, M.J.; Macal, C.M. Managing Business Complexity: Discovering Strategic Solutions with Agent-Based Modeling and Simulation; Oxford University Press: Oxford, UK, 2007. [Google Scholar]
- Grimm, V.; Revilla, E.; Berger, U.; Jeltsch, F.; Mooij, W.M.; Railsback, S.F.; Thulke, H.H.; Weiner, J.; Wiegand, T.; DeAngelis, D.L. Pattern-Oriented Modeling of Agent-Based Complex Systems: Lessons from Ecology. Science 2005, 210, 987–991. [Google Scholar] [CrossRef] [Green Version]
- Wiegand, T.; Jeltsch, F.; Hanski, I.; Grimm, V. Using pattern-oriented modeling for revealing hidden information: A key for reconciling ecological theory and application. Oikos 2003, 100, 209–222. [Google Scholar] [CrossRef] [Green Version]
- Rossi, R.E.; Mulla, D.J.; Journel, A.G.; Franz, E.H. Geostatistical tools for modeling and interpreting ecological spatial dependence. Ecol. Monogr. 1992, 62, 277–314. [Google Scholar] [CrossRef]
- Ribeiro, P.; Diggle, P. geoR: A Package for Geostatistical Analysis. R News 2001, 1, 15–18. [Google Scholar]
- Adler, P.; Raff, D.; Lauenroth, W. The effect of grazing on the spatial heterogeneity of vegetation. Oecologia 2001, 128, 465–479. [Google Scholar] [CrossRef] [PubMed]
- Nathan, R.; Muller-Landau, H.C. Spatial patterns of seed dispersal, their determinants and consequences for recruitment. Trends Ecol. Evol. 2000, 15, 278–285. [Google Scholar] [CrossRef]
- Florencia Miguel, M.; Tabeni, S.; Cona, M.I.; Campos, C.M. Secondary seed dispersal by mammals between protected and grazed semiarid woodland. For. Ecol. Manag. 2018, 422, 41–48. [Google Scholar] [CrossRef]
- Sun, Z.L.; Lorscheid, I.; Millington, J.D.; Lauf, S.; Magliocca, N.R.; Groeneveld, J.; Balbi, S.; Nolzen, H.; Muller, B.; Schulze, J.; et al. Simple or complicated agent-based models? A complicated issue. Environ. Model. Softw. 2016, 86, 56–67. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
The location of the study area (source: authors). (a) Horqin Sandy Land and the field investigation sites (marked with the red rectangle); (b) location of Horqin Sandy Land in China; (c) landscape of sparse elm woodlands; (d) elm seed shadow on the ground; (e) field experiments on the spatial pattern of seed densities; (f) aerial overview of the landscape of sparse elm woodlands.
Figure 1.
The location of the study area (source: authors). (a) Horqin Sandy Land and the field investigation sites (marked with the red rectangle); (b) location of Horqin Sandy Land in China; (c) landscape of sparse elm woodlands; (d) elm seed shadow on the ground; (e) field experiments on the spatial pattern of seed densities; (f) aerial overview of the landscape of sparse elm woodlands.

Figure 2.
The diagram of geostatistical indices.
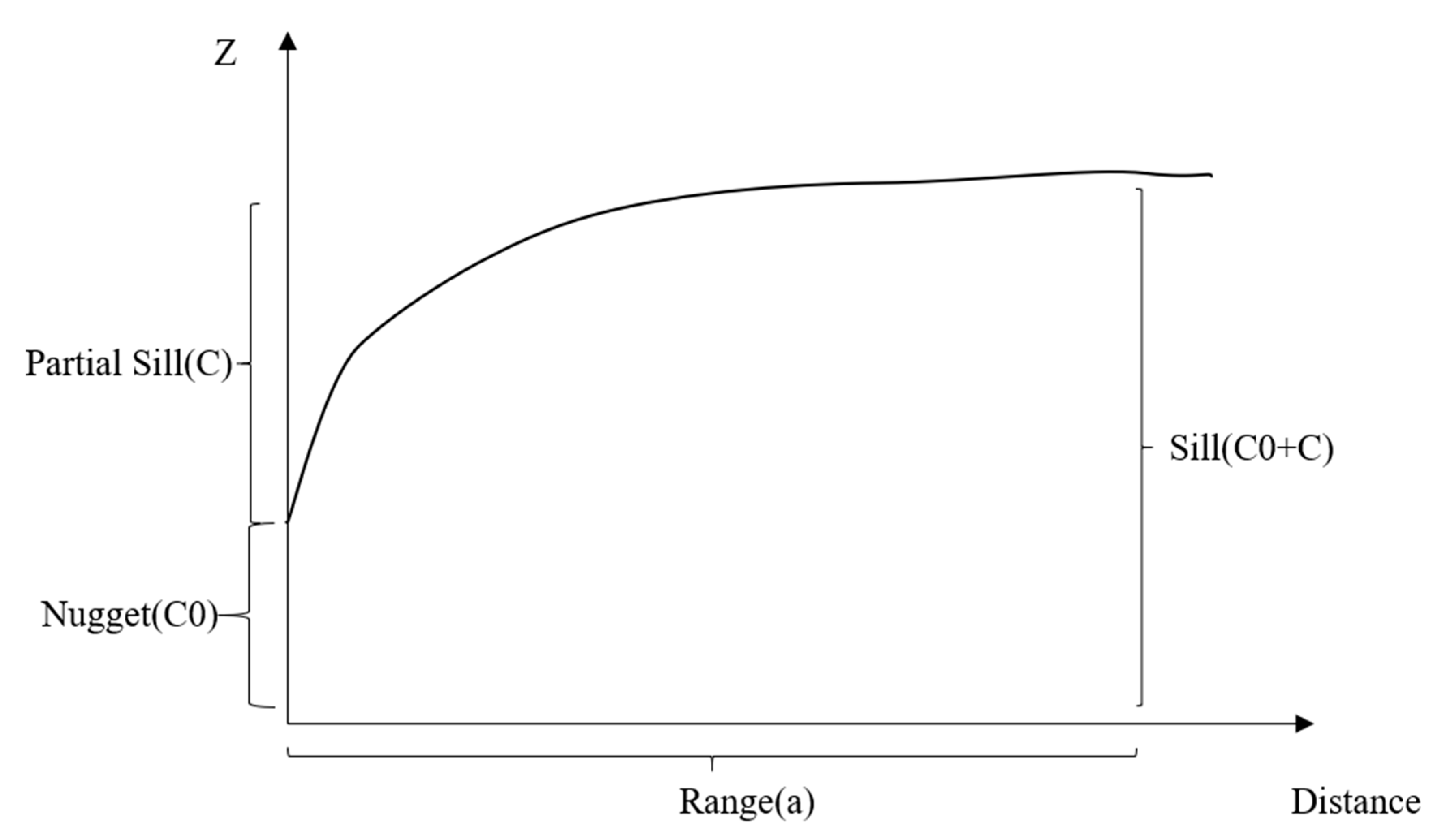
Figure 3.
The geostatistical indices of seed density under different grazing intensities. (a) The values of nugget, partial still, and still under different grazing intensities; (b) the ratio of nugget to sill under different grazing intensities. CK: without grazing; LG: light grazing; MG: moderate grazing; HG: heavy grazing.
Figure 3.
The geostatistical indices of seed density under different grazing intensities. (a) The values of nugget, partial still, and still under different grazing intensities; (b) the ratio of nugget to sill under different grazing intensities. CK: without grazing; LG: light grazing; MG: moderate grazing; HG: heavy grazing.

{kind=link}
{kind=link}
{kind=link}
Table 1.
The comparison between the results of the field investigation (n = 4) and model outputs (n = 10).
Table 1.
The comparison between the results of the field investigation (n = 4) and model outputs (n = 10).
| Indices | Field Investigation | Model | t-Statistics | p-Values | ||
|---|---|---|---|---|---|---|
| Mean | S.E. | Mean | S.E. | |||
| Nugget (C0) | 127.93 | 118.18 | 39.57 | 4.03 | 1.258 | 0.240 |
| Partial Sill (C) | 241.05 | 216.3 | 38.49 | 4.46 | 1.580 | 0.149 |
| Sill (C + C0) | 368.97 | 334.41 | 78.06 | 2.56 | 1.470 | 0.176 |
| Nugget/Sill (C0/C0 + C) | 30.26 | 10.7 | 50.69 | 0.05 | 3.227 | 0.010 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tang, Y.; Liu, M.; Sun, Z. Indirect Effects of Grazing on Wind-Dispersed Elm Seeds in Sparse Woodlands of Northern China. Land 2020, 9, 490. https://doi.org/10.3390/land9120490
AMA Style
Tang Y, Liu M, Sun Z. Indirect Effects of Grazing on Wind-Dispersed Elm Seeds in Sparse Woodlands of Northern China. Land. 2020; 9(12):490. https://doi.org/10.3390/land9120490
Chicago/Turabian StyleTang, Yi, Mingyu Liu, and Zhanli Sun. 2020. "Indirect Effects of Grazing on Wind-Dispersed Elm Seeds in Sparse Woodlands of Northern China" Land 9, no. 12: 490. https://doi.org/10.3390/land9120490
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.