How Do Invasive Species Influence Biotic and Abiotic Factors Drive Vegetation Success in Salt Marsh Ecosystems?

 ,
,
Abstract
1. Introduction
2. Materials and Methods
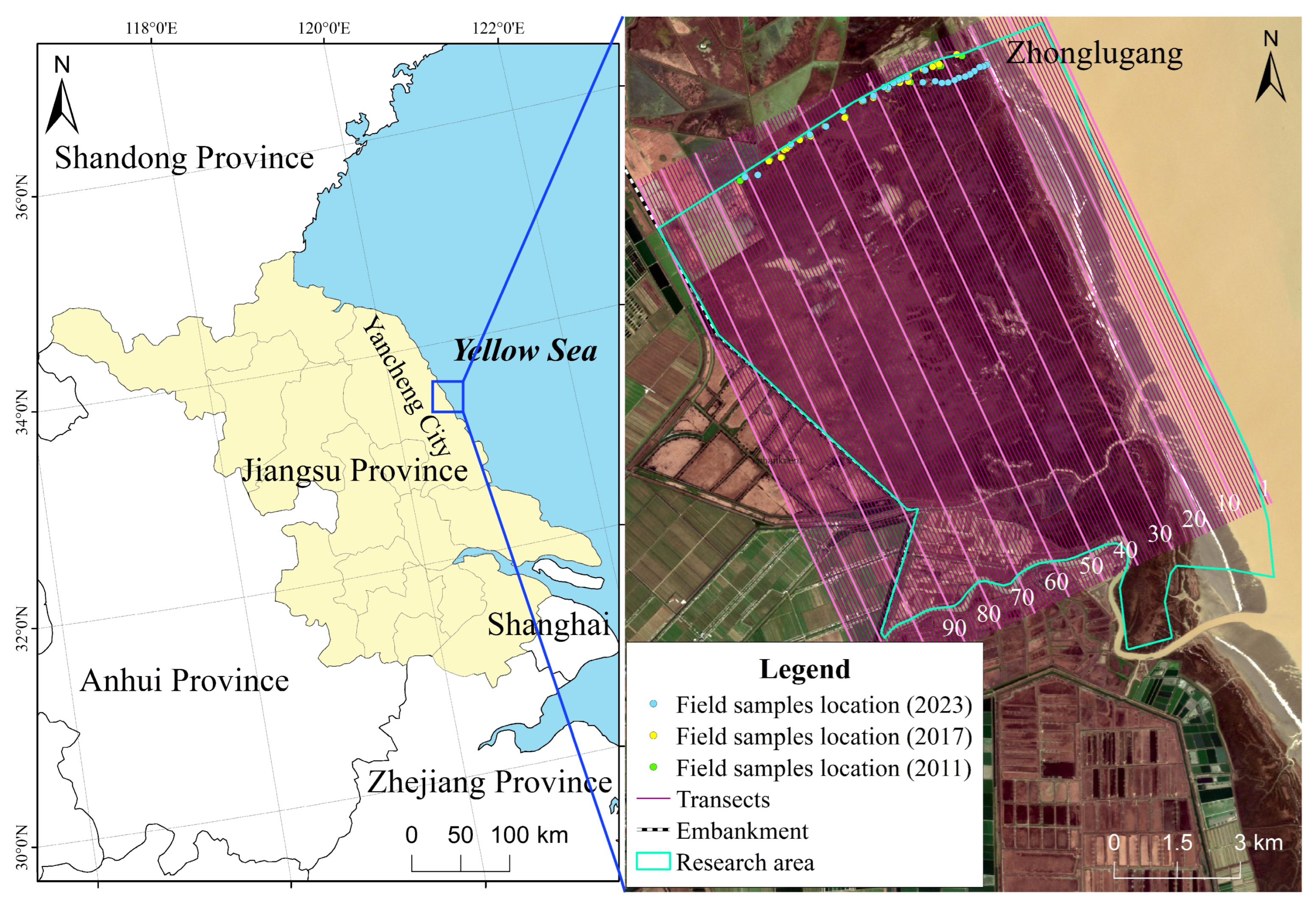
2.1. Study Sites
2.2. Data Collection
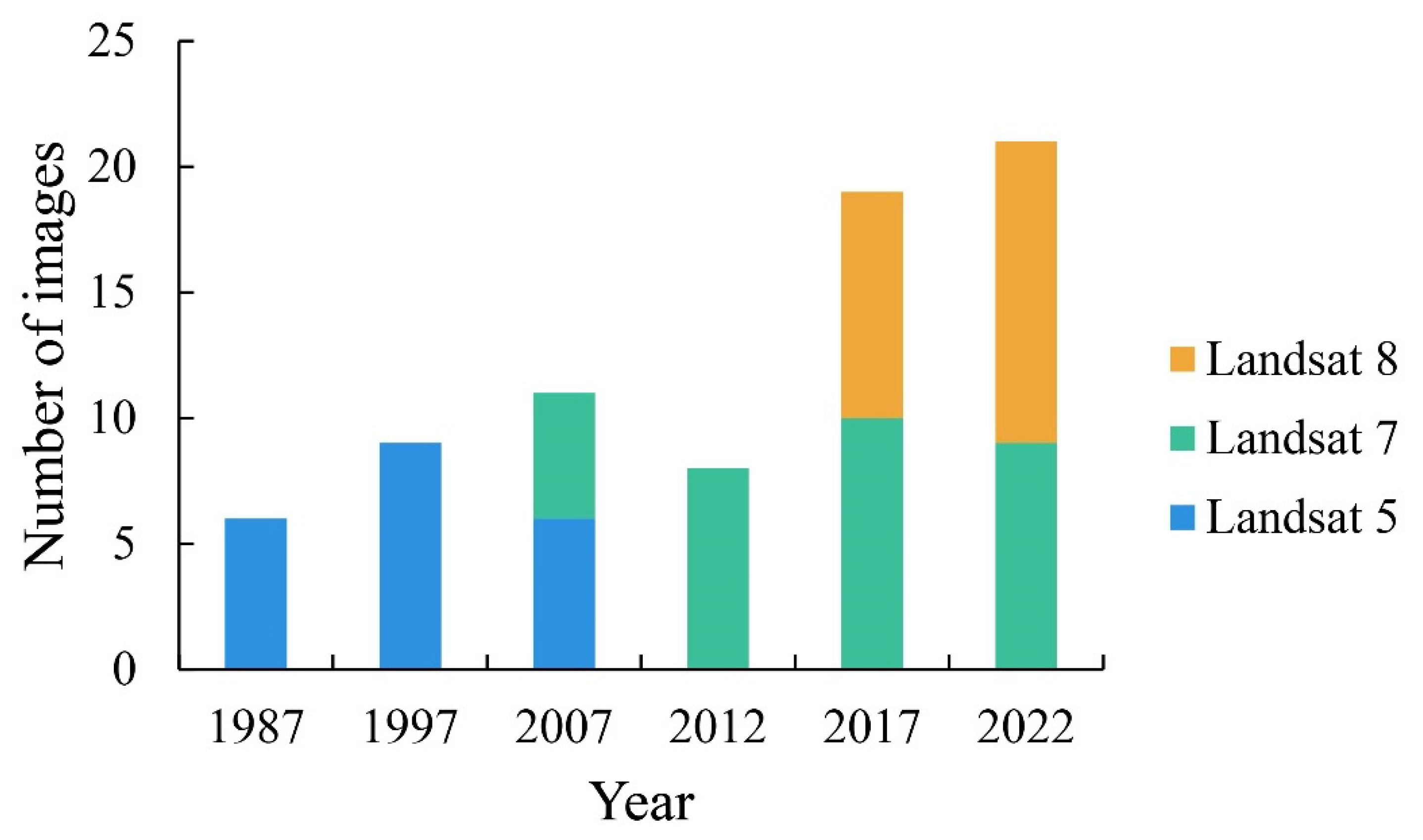
2.2.1. Satellite Remote Sensing Images
2.2.2. Soil Salinity
2.3. Methods
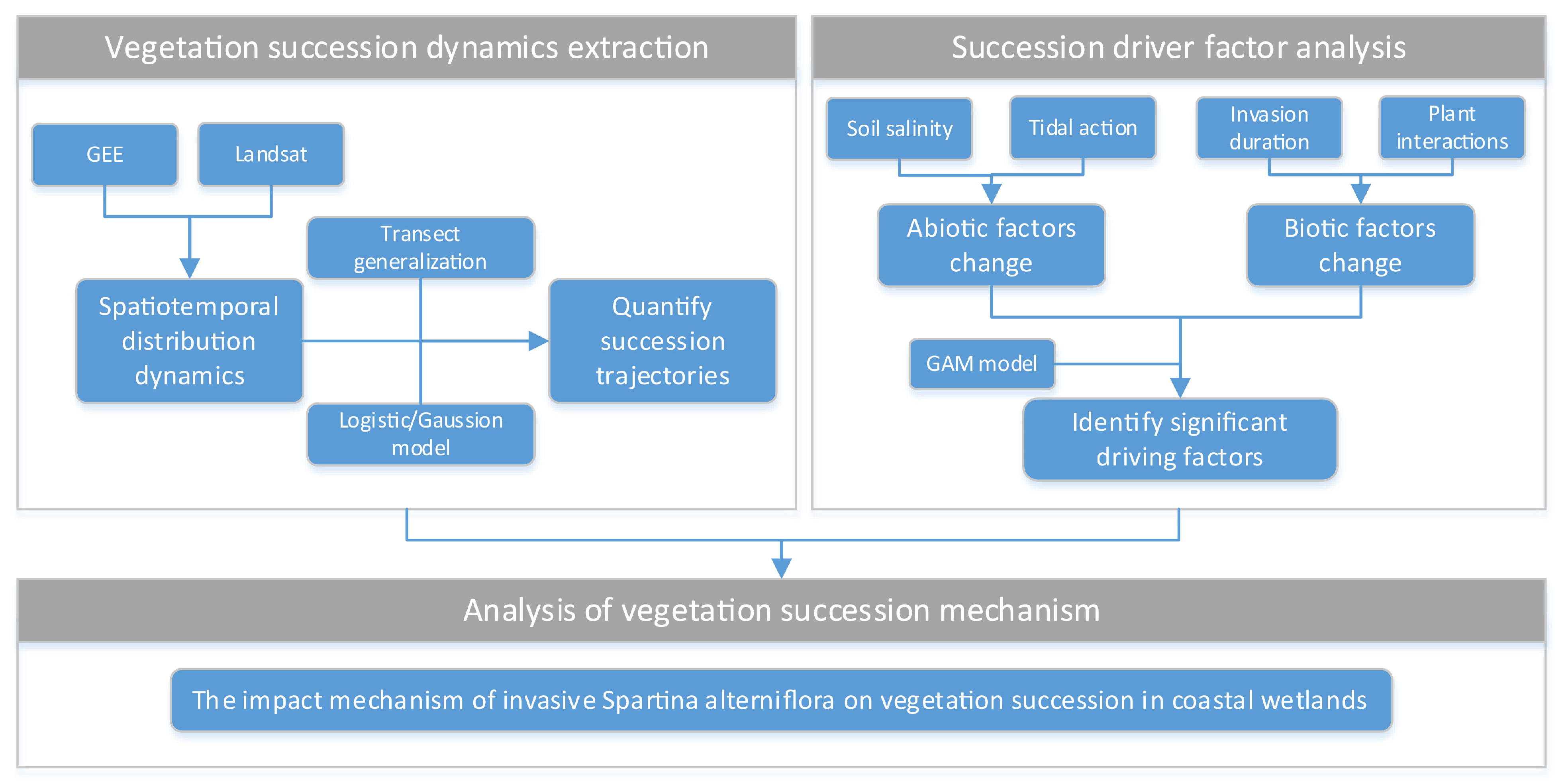
2.3.1. Overall Analysis Framework
2.3.2. Classification and Interspecific Interaction Extraction of Plant Communities via the Google Earth Engine
- (1)
- Classification of typical plant community types
- (2)
- Extraction of changes in interspecific interactions based on fractional vegetation cover
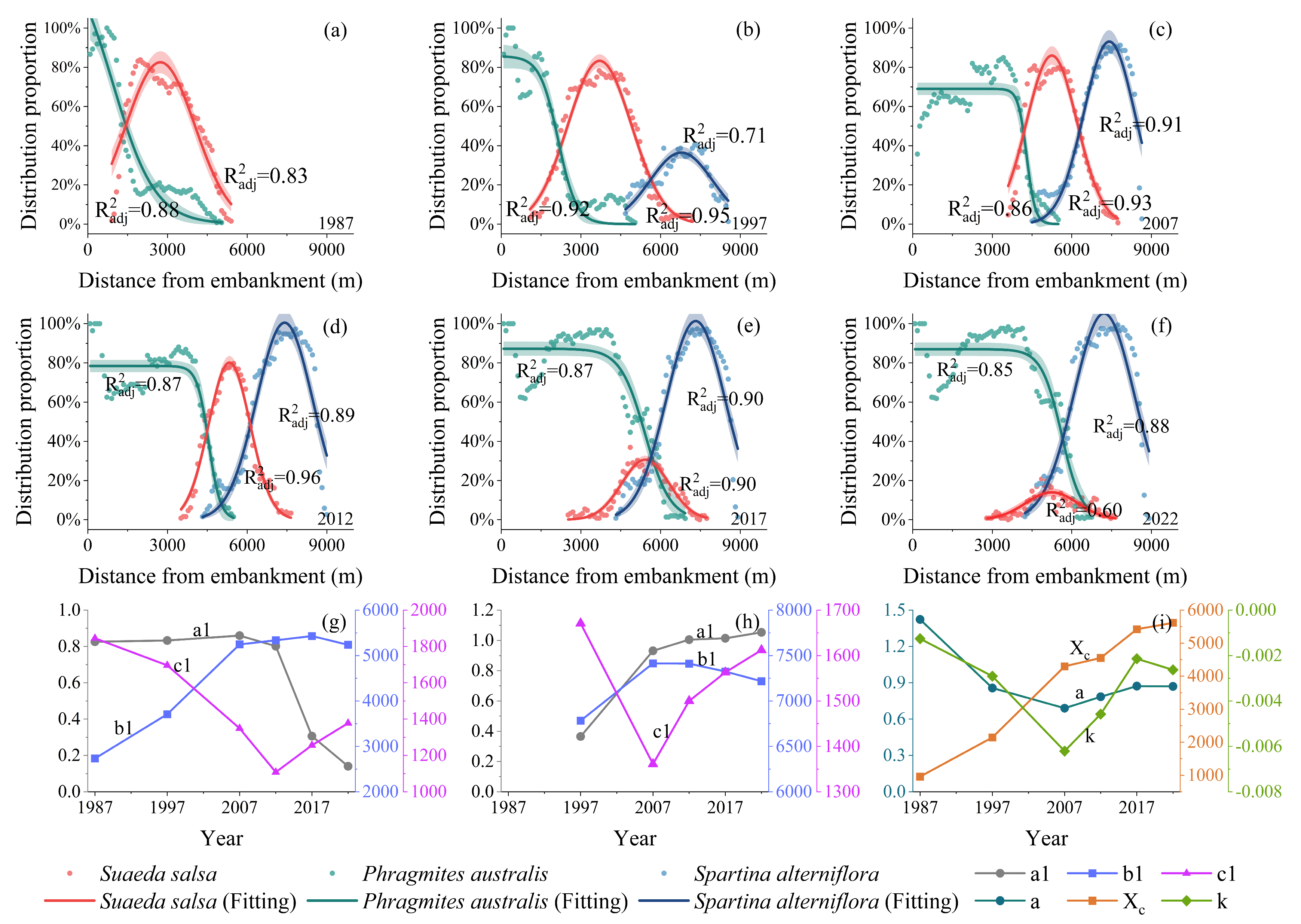
2.3.3. Modeling the Relationship Between the Sea–Land Distribution Positions and the Distribution Proportions of Typical Plant Communities
2.3.4. Modeling the Driving Factors and Mechanisms of Vegetation Succession
2.3.5. Statistical Analysis
3. Results
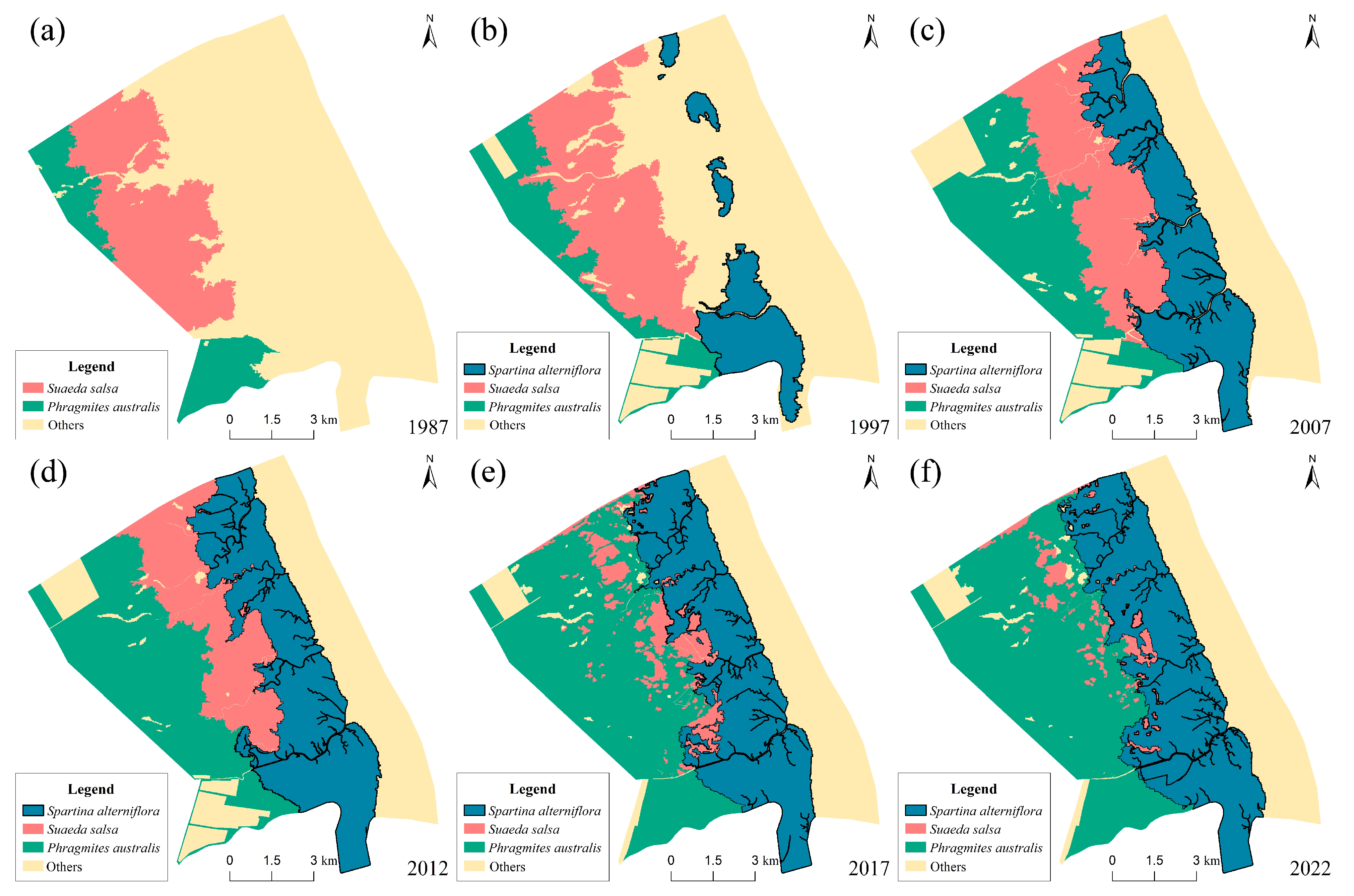
3.1. Changes in Vegetation Succession Within Salt Marsh Ecosystems
- (1)
- Changes in the type and area of plant communities
- (2)
- Changes in the spatial distribution pattern of plant communities
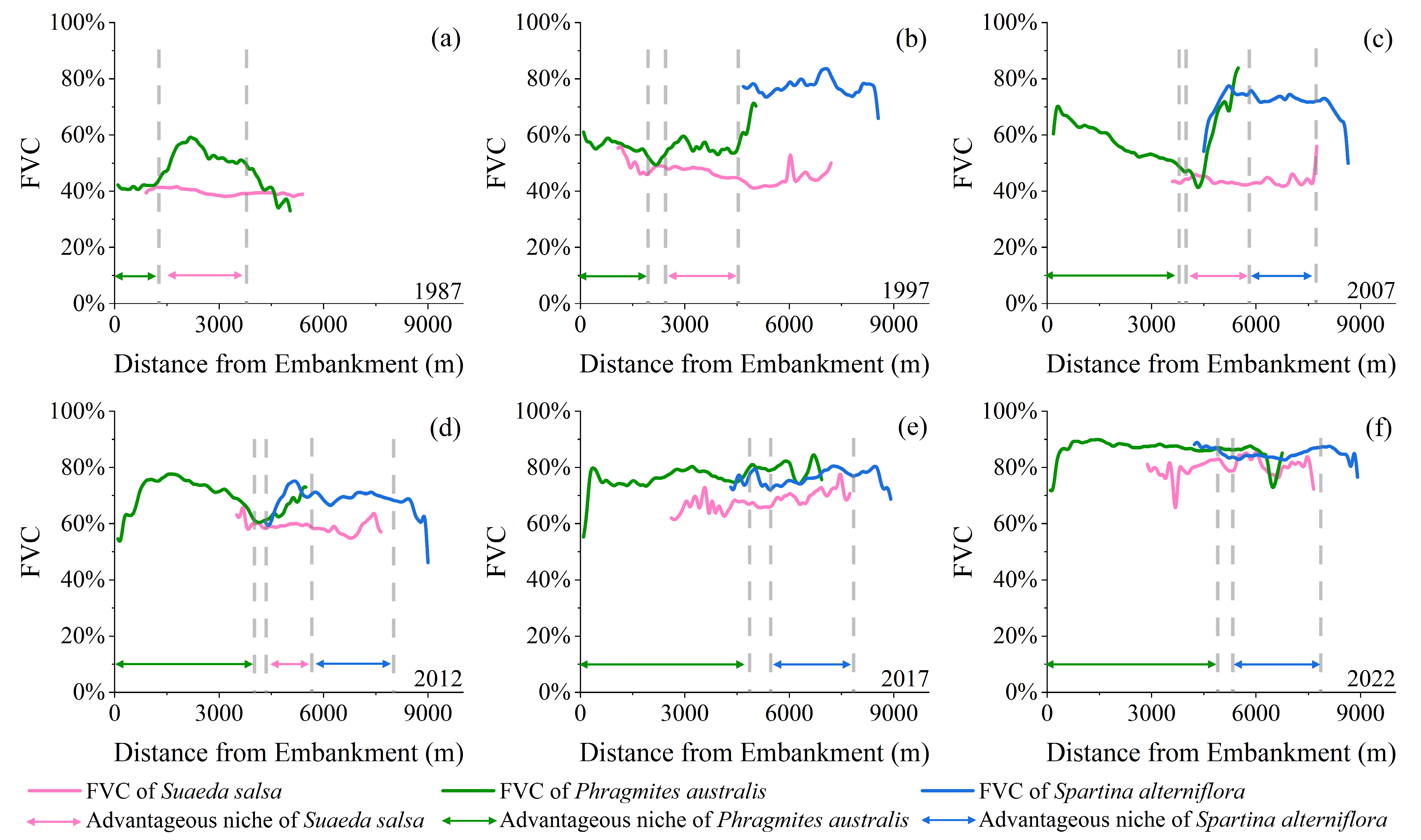
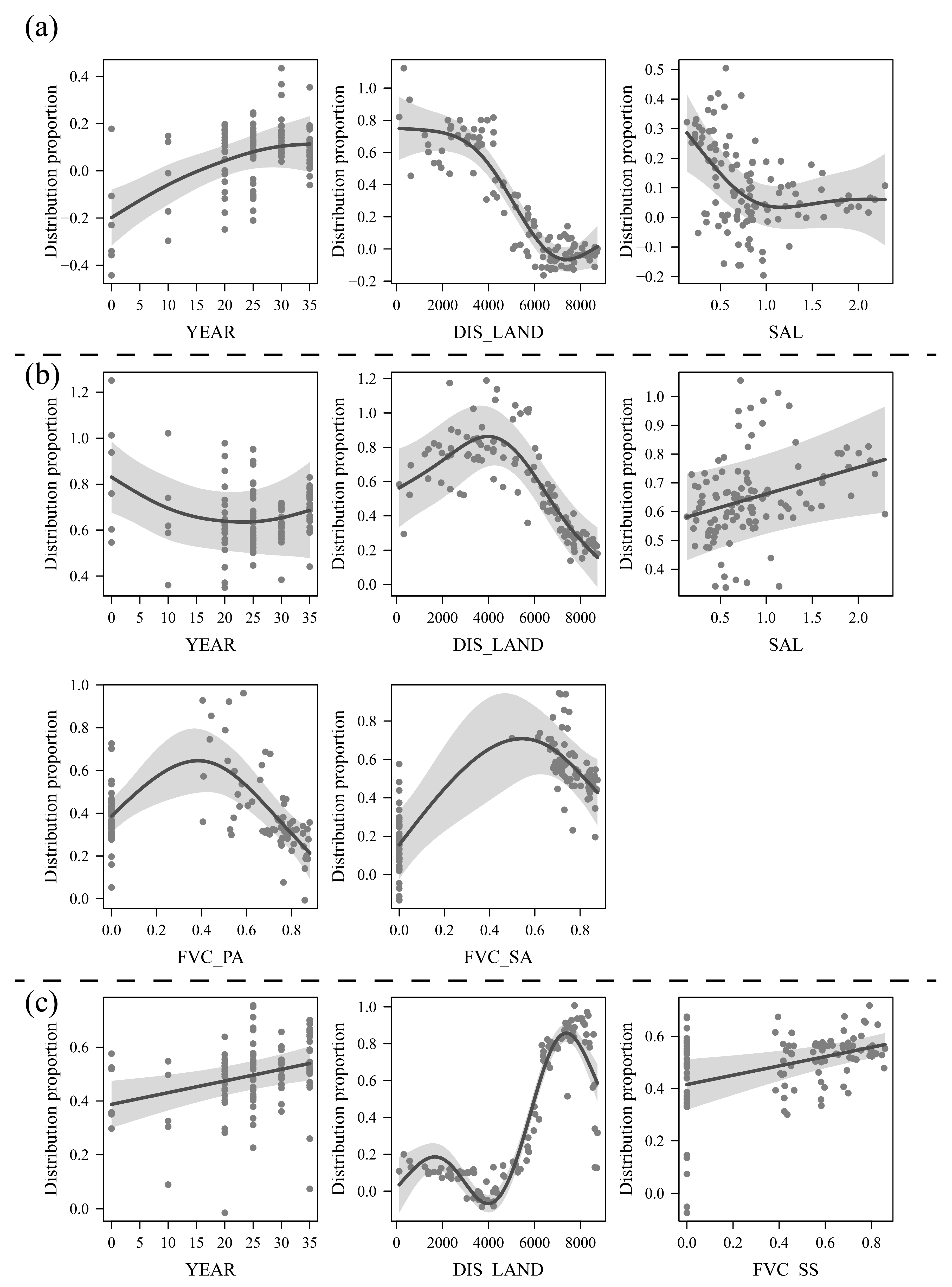
3.2. Analysis of Changes in the Driving Factors and Driving Effects of Vegetation Succession
- (1)
- Spatial and temporal changes in soil salinity
- (2)
- Changes in interspecific interactions based on fractional vegetation cover
- (3)
- Analysis of the driving effects of different factors on vegetation succession
4. Discussion
4.1. Coupling of Major Biotic and Abiotic Factors with Salt Marsh Vegetation Succession
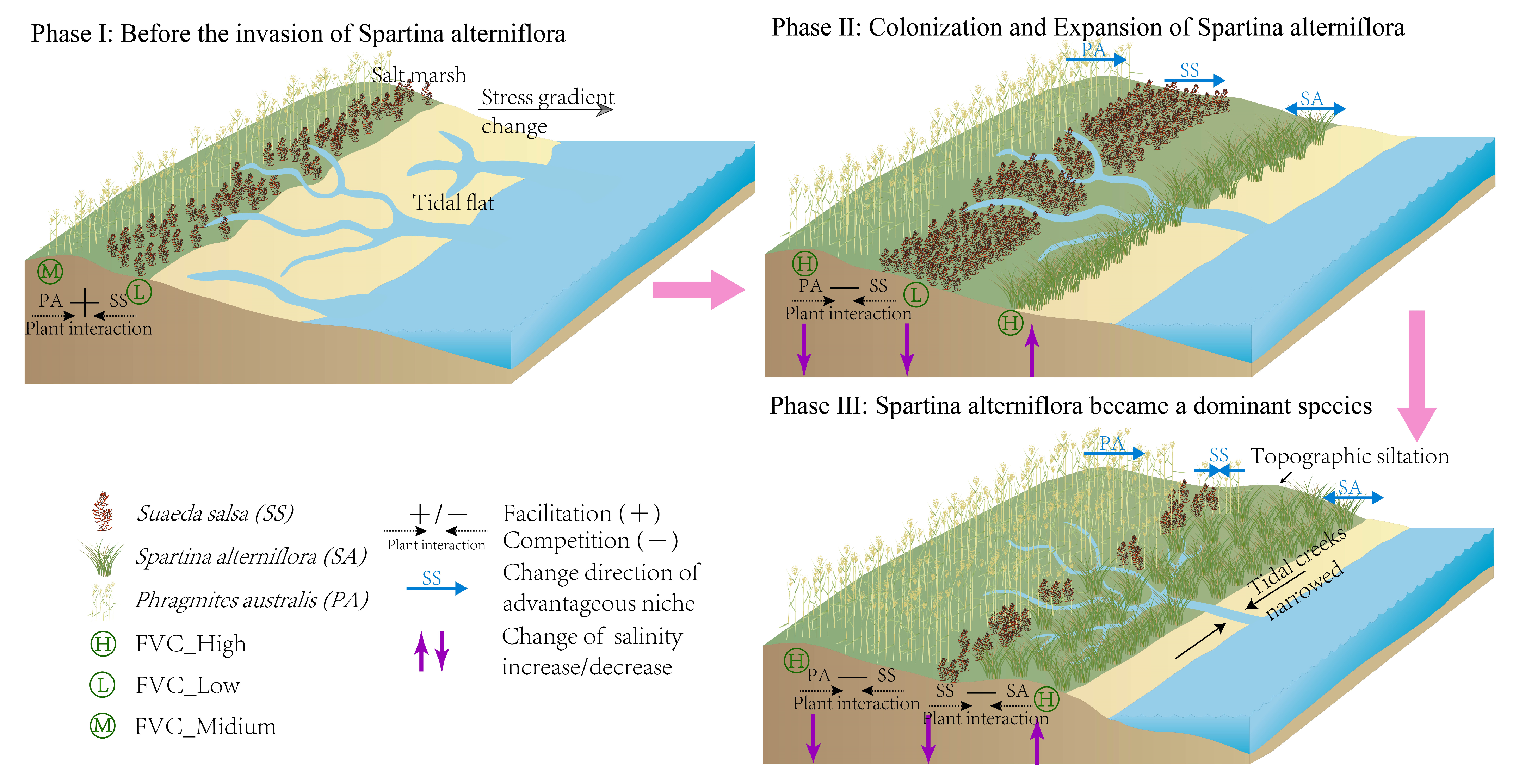
4.2. Mechanisms of the Impact of Biological Invasions on Changes in Vegetation Succession
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Poorter, L.; Amissah, L.; Bongers, F.; Hordijk, I.; Kok, J.; Laurance, S.; Lohbeck, M.; Martínez-Ramos, M.; Matsuo, T.; Meave, J.; et al. Successional theories. Biol. Rev. 2023, 98, 2049–2077. [Google Scholar] [CrossRef] [PubMed]
- Shugart, H.H. Ecological succession and community dynamics. In Encyclopedia of Sustainability Science and Technology; Springer: New York, NY, USA, 2012; pp. 3278–3296. [Google Scholar]
- Li, T.; Yang, H.; Yang, X.; Guo, Z.; Fu, D.; Liu, C.; Li, S.; Pan, Y.; Zhao, Y.; Xu, F.; et al. Community assembly during vegetation succession after metal mining is driven by multiple processes with temporal variation. Ecol. Evol. 2022, 12, e8882. [Google Scholar] [CrossRef] [PubMed]
- Purschke, O.; Schmid, B.C.; Sykes, M.T.; Poschlod, P.; Michalski, S.G.; Durka, W.; Kühn, I.; Winter, M.; Prentice, H.C. Contrasting changes in taxonomic, phylogenetic and functional diversity during a long-term succession: Insights into assembly processes. J. Ecol. 2013, 101, 857–866. [Google Scholar] [CrossRef]
- Måren, I.E.; Kapfer, J.; Aarrestad, P.A.; Grytnes, J.; Vandvik, V. Changing contributions of stochastic and deterministic processes in community assembly over a successional gradient. Ecology 2017, 99, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Das Gupta, S.; Pinno, B.D. Spatial patterns and competition in trees in early successional reclaimed and natural boreal forests. Acta Oecol. 2018, 92, 138–147. [Google Scholar] [CrossRef]
- Han, T.; Ren, H.; Hui, D.; Zhu, Y.; Lu, H.; Guo, Q.; Wang, J. Dominant ecological processes and plant functional strategies change during the succession of a subtropical forest. Ecol. Indic. 2023, 146, 109885. [Google Scholar] [CrossRef]
- Crooks, J.A. Characterizing ecosystem-level consequences of biological invasions: The role of ecosystem engineers. Oikos 2002, 97, 153–166. [Google Scholar] [CrossRef]
- Walsh, J.R.; Carpenter, S.R.; Vander Zanden, M.J. Invasive species triggers a massive loss of ecosystem services through a trophic cascade. Proc. Natl. Acad. Sci. USA 2016, 113, 4081–4085. [Google Scholar] [CrossRef] [PubMed]
- Meiners, S.J.; Rye, T.A.; Klass, J.R. On a level field: The utility of studying native and non-native species in successional systems. Appl. Veg. Sci. 2009, 12, 45–53. [Google Scholar] [CrossRef]
- Muñoz-Rodríguez, A.F.; Infante-Izquierdo, M.D.; Polo-Ávila, A.; Hermoso-López, V.; Nieva, F.J.J.; Gallego-Tévar, B.; Castillo, J.M. Recruitment niche segregation of halophytes along the tidal gradient. Estuar. Coast. Shelf Sci. 2024, 305, 108859. [Google Scholar] [CrossRef]
- Prach, K.; Walker, L.R. Four opportunities for studies of ecological succession. Trends Ecol. Evol. 2011, 26, 119–123. [Google Scholar] [CrossRef] [PubMed]
- Ning, Z.; Chen, C.; Xie, T.; Li, S.; Zhu, Z.; Wang, Q.; Cai, Y.; Bai, J.; Cui, B. Invasive plant indirectly affects its self-expansion and native species via bio-geomorphic feedbacks: Implications for salt marsh restoration. CATENA 2023, 226, 107056. [Google Scholar] [CrossRef]
- Zhu, K.-H.; Zeng, J.; Ge, Z.-M.; Zuo, Y.; Li, S.-H.; Zhao, L.-H.; Han, Y.; Cheng, H.-F.; Xin, P. A model coupling ecological and hydrodynamic processes for simulating the biogeomorphology of a coastal salt marsh. Ecol. Modell. 2024, 493, 110758. [Google Scholar] [CrossRef]
- Yan, D.; Li, J.; Yao, X.; Luan, Z. Integrating UAV data for assessing the ecological response of Spartina alterniflora towards inundation and salinity gradients in coastal wetland. Sci. Total Environ. 2022, 814, 152631. [Google Scholar] [CrossRef] [PubMed]
- Ivajnšič, D.; Šajna, N.; Kaligarič, M. Primary succession on re-created coastal wetland leads to successful restoration of coastal halophyte vegetation. Landsc. Urban Plan. 2016, 150, 79–86. [Google Scholar] [CrossRef]
- Zhou, Z.; Hua, J.; Xue, J. Salinity drives shifts in soil microbial community composition and network complexity along vegetation community succession in coastal tidal flats. Estuar. Coast. Shelf Sci. 2022, 276, 108005. [Google Scholar] [CrossRef]
- Veldhuis, E.R.; Schrama, M.; Staal, M.; Elzenga, J.T.M. Plant stress-tolerance traits predict salt marsh vegetation patterning. Front. Mar. Sci. 2019, 5, 501. [Google Scholar] [CrossRef]
- Wang, A.; Gao, S.; Jia, J. Impact of Spartina alterniflora on sedimentary and morphological evolution of tidal salt marshes of Jiangsu, China. Hai Yang Xue Bao 2006, 28, 92–99. (In Chinese) [Google Scholar]
- Seo, J.Y.; Choi, S.M.; Ha, H.K. Assessment of potential impact of invasive vegetation on cohesive sediment erodibility in intertidal flats. Sci. Total Environ. 2021, 766, 144493. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Wang, C.; Li, Y.; Huang, W.; Jia, Y.; Wang, Y.; Xu, W.; Qiu, C.; Liu, H. Study on spatio-temporal variation and hydrological connectivity of tidal creek evolution in Yancheng coastal wetlands. Environ. Sci. Pollut. Res. 2022, 30, 37143–37156. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Han, G.; Wang, X.; Zhang, B. Effects of Spartina alterniflora invasion on morphological characteristics of tidal creeks and plant community distribution in the Yellow River Estuary. Chin. J. Ecol. 2022, 41, 42–49. (In Chinese) [Google Scholar]
- Lorrain-Soligon, L.; Robin, F.; Bertin, X.; Jankovic, M.; Rousseau, P.; Lelong, V.; Brischoux, F. Long-term trends of salinity in coastal wetlands: Effects of climate, extreme weather events, and sea water level. Environ. Res. 2023, 237, 116937. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Liang, Z.; Li, X.; Wang, X.; Chu, X.; Zhao, M.; Zhang, X.; Li, P.; Song, W.; Huang, W.; et al. Precipitation changes alter plant dominant species and functional groups by changing soil salinity in a coastal salt marsh. J. Environ. Manag. 2024, 368, 122235. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Ge, Z.-M.; Tan, L.-S.; Xie, L.-N.; Li, Y.-L. Multiple competitive superiority made a great successful invasion of Spartina alterniflora in Eastern China: Hints for management. J. Environ. Manag. 2025, 389, 126287. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Liu, Z.; Zheng, G.; Zhao, C. Identification of Spartina alterniflora habitat expansion in a Suaeda salsa dominated coastal wetlands. Ecol. Indic. 2022, 145, 109704. [Google Scholar] [CrossRef]
- Kulmatiski, A. Exotic plants establish persistent communities. Plant Ecol. 2006, 187, 261–275. [Google Scholar] [CrossRef]
- Nawaz, M.; Sun, J.; Shabbir, S.; Khattak, W.A.; Ren, G.; Nie, X.; Bo, Y.; Javed, Q.; Du, D.; Sonne, C. A review of plants strategies to resist biotic and abiotic environmental stressors. Sci. Total Environ. 2023, 900, 165832. [Google Scholar] [CrossRef] [PubMed]
- Muench, A.; Elsey-Quirk, T. Competitive reversal between plant species is driven by species-specific tolerance to flooding stress and nutrient acquisition during early marsh succession. J. Appl. Ecol. 2019, 56, 2236–2247. [Google Scholar] [CrossRef]
- Ma, S.; Ren, J.; Wu, C.; He, Q. Extreme precipitation events trigger abrupt vegetation succession in emerging coastal wetlands. CATENA 2024, 241, 108066. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhang, D.; Cutler, M.E.J.; Xu, N.; Wang, X.H.; Sha, H.; Shen, Y. Estimating muddy intertidal flat slopes under varied coastal morphology using sequential satellite data and spatial analysis. Estuar. Coast. Shelf Sci. 2021, 251, 107183. [Google Scholar] [CrossRef]
- Kuai, Y.; Tao, J.; Zhou, Z.; Aarninkhof, S.; Wang, Z.B. Sediment characteristics and intertidal beach slopes along the Jiangsu Coast, China. J. Mar. Sci. Eng. 2021, 9, 347. [Google Scholar] [CrossRef]
- Fan, X.; Zhang, L.; Yuan, L.; Guo, B.; Zhang, Q.; Wang, Y.; Wu, Q. Loss of tidal creek ecosystem vitality caused by tidal flat narrowing on the central Jiangsu coast, China. Sci. Total Environ. 2023, 864, 161216. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Liu, H.; Li, Y.; Dong, B.; Qiu, C.; Yang, J.; Zong, Y.; Chen, H.; Zhao, Y.; Zhang, Y. Study on habitat suitability and environmental variable thresholds of rare waterbirds. Sci. Total Environ. 2021, 785, 147316. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Hu, Y.; Liu, M.; Chang, Y.; Yan, X.; Bu, R.; Zhao, D.; Li, Z. Introduction and spread of an exotic plant, Spartina alterniflora, along coastal marshes of China. Wetlands 2017, 37, 1181–1193. [Google Scholar] [CrossRef]
- Feng, Y. The ecological characteristics and evolution of the tidal flat in Zhonglu Port. Jiangsu Agric. Sci. 1989, 1, 18–20. (In Chinese) [Google Scholar]
- Yang, G.; Shi, Y.; Ji, Z. The ecological response of typical mud flat to sea level change in Jiangsu coastal plain. Acta Geogr. Sin. 2002, 1, 76–84. (In Chinese) [Google Scholar]
- Ren, L.; Wang, G.; He, D.; Mao, Z.; Liu, J. Spatial distributions of soil organic matter in different vegetation zones of the Yancheng tidal flat. Adv. Mar. Biol. 2011, 29, 54–62. (In Chinese) [Google Scholar]
- Mao, Z.; Gu, X.; Liu, J.; Ren, L.; Wang, G. Evolvement of soil quality in saltmarshes and reclaimed farm lands in Yancheng coastal wetland. Chin. J. Appl. Ecol. 2010, 21, 1986–1992. (In Chinese) [Google Scholar]
- Zhang, H.; Zhen, Y.; Li, Y.; Sun, X. Spatial heterogeneity of soil Salinity in Jiangsu Yancheng Wetland National Nature Reserve, Rare Bird. Wetl. Sci. 2018, 16, 152–158. (In Chinese) [Google Scholar]
- Tucker, C.J. Red and photographic infrared linear combinations for monitoring vegetation. Remote Sens. Environ. 1979, 8, 127–150. [Google Scholar] [CrossRef]
- Xu, H. Modification of normalised difference water index (NDWI) to enhance open water features in remotely sensed imagery. Int. J. Remote Sens. 2006, 27, 3025–3033. [Google Scholar] [CrossRef]
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, L.G. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Diek, S.; Fornallaz, F.; Schaepman, M.E.; De Jong, R. Barest pixel composite for agricultural areas using Landsat time series. Remote Sens. 2017, 9, 1245. [Google Scholar] [CrossRef]
- Breiman, L. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Lou, Y.; Dai, Z.; Long, C.; Dong, H.; Wei, W.; Ge, Z. Image-based machine learning for monitoring the dynamics of the largest salt marsh in the Yangtze River Delta. J. Hydrol. 2022, 608, 127681. [Google Scholar] [CrossRef]
- Liancourt, P.; Callaway, R.M.; Michalet, R. Stress tolerance and competitive-response ability determine the outcome of biotic interactions. Ecology 2005, 86, 1611–1618. [Google Scholar] [CrossRef]
- Carlson, T.N.; Ripley, D.A. On the relation between NDVI, fractional vegetation cover, and leaf area index. Remote Sens. Environ. 1997, 62, 241–252. [Google Scholar] [CrossRef]
- Wang, B.; Zhang, K.; Liu, Q.-X.; He, Q.; van de Koppel, J.; Teng, S.N.; Miao, X.; Liu, M.; Bertness, M.D.; Xu, C. Long-distance facilitation of coastal ecosystem structure and resilience. Proc. Natl. Acad. Sci. USA 2022, 119, e2123274119. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J.; Minchin, P.R. Continuum theory revisited: What shape are species responses along ecological gradients? Ecol. Modell. 2002, 157, 119–129. [Google Scholar] [CrossRef]
- Coudun, C.; Gégout, J.-C. The derivation of species response curves with Gaussian logistic regression is sensitive to sampling intensity and curve characteristics. Ecol. Modell. 2006, 199, 164–175. [Google Scholar] [CrossRef]
- Ma, S.; Ren, J.; Wu, C.; Cheng, F.; Wang, X.; Li, B.; He, Q. Hydrological control of threshold transitions in vegetation over early-period wetland development. J. Hydrol. 2022, 610, 127931. [Google Scholar] [CrossRef]
- Sun, C.; Li, J.; Liu, Y.; Zhao, S.; Zheng, J.; Zhang, S. Tracking annual changes in the distribution and composition of saltmarsh vegetation on the Jiangsu coast of China using Landsat time series–based phenological parameters. Remote Sens. Environ. 2023, 284, 113370. [Google Scholar] [CrossRef]
- Li, X.; Yang, W.; Li, S.; Sun, T.; Bai, J.; Pei, J.; Xie, T.; Cui, B. Asymmetric responses of spatial variation of different communities to a salinity gradient in coastal wetlands. Mar. Environ. Res. 2020, 158, 105008. [Google Scholar] [CrossRef] [PubMed]
- Lissner, J.; Schierup, H.-H. Effects of salinity on the growth of Phragmites australis. Aquat. Bot. 1997, 55, 247–260. [Google Scholar] [CrossRef]
- Liu, Y.; Ding, Z.; Bachofen, C.; Lou, Y.; Jiang, M.; Tang, X.; Lu, X.; Buchmann, N. The effect of saline-alkaline and water stresses on water use efficiency and standing biomass of Phragmites australis and Bolboschoenus planiculmis. Sci. Total Environ. 2018, 644, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Gao, F.; Pang, J.; Wang, H.; Wang, H.; Wang, Y.; Whitt, A.A.; Ma, C. Plant-plant interactions of Phragmites australis and Suaeda salsa as mediated by combined influences of salinity and tidal level changes. Plant Soil 2022, 474, 141–161. [Google Scholar] [CrossRef]
- Matsuda, R.; Yamada, K.; Hayasaka, D.; Henmi, Y. Effects of salinity, temperature, and immersion conditions on seed germination of invasive Spartina alterniflora Loisel (smooth cordgrass) in Japan. Reg. Stud. Mar. Sci. 2023, 57, 102738. [Google Scholar] [CrossRef]
- Shi, F.; Bao, F. Effects of salinity and temperature stress on ecophysiological characteristics of exotic cordgrass, Spartina alterniflora. Sheng Tai Xue Bao 2007, 27, 2733–2741. [Google Scholar] [CrossRef]
- Elsey-Quirk, T.; Middleton, B.A.; Proffitt, C.E. Seed flotation and germination of salt marsh plants: The effects of stratification, salinity, and/or inundation regime. Aquat. Bot. 2009, 91, 40–46. [Google Scholar] [CrossRef]
- Schwarz, C.; Bouma, T.J.; Zhang, L.Q.; Temmerman, S.; Ysebaert, T.; Herman, P.M.J. Interactions between plant traits and sediment characteristics influencing species establishment and scale-dependent feedbacks in salt marsh ecosystems. Geomorphology 2015, 250, 298–307. [Google Scholar] [CrossRef]
- Zhou, Y.; Qiu, C.; Li, Y.; Wang, C.; Zhang, Y.; Huang, W.; Li, L.; Liu, H.; Zhang, D. Integrating UAV data to explore the relationship between microtopographic variation and Spartina alterniflora expansion during its early invasion. Ecol. Indic. 2023, 154, 110633. [Google Scholar] [CrossRef]
- Ma, X.; Yan, J.; Wang, F.; Qiu, D.; Jiang, X.; Liu, Z.; Sui, H.; Bai, J.; Cui, B. Trait and density responses of Spartina alterniflora to inundation in the Yellow River Delta, China. Mar. Pollut. Bull. 2019, 146, 857–864. [Google Scholar] [CrossRef] [PubMed]
- Pang, B.; Xie, T.; Ning, Z.; Cui, B.; Zhang, H.; Wang, X.; Gao, F.; Zhang, S.; Lu, Y. Invasion patterns of Spartina alterniflora: Response of clones and seedlings to flooding and salinity—A case study in the Yellow River Delta, China. Sci. Total Environ. 2023, 877, 162803. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Guo, X.; Yu, X.; Liu, L.; Wang, N.; Eller, F.; Guo, W. Is there evidence of local adaptation of Phragmites australis to water level gradients and fluctuation frequencies? Sci. Total Environ. 2021, 756, 144065. [Google Scholar] [CrossRef] [PubMed]
- Soliveres, S.; Smit, C.; Maestre, F.T. Moving forward on facilitation research: Response to changing environments and effects on the diversity, functioning and evolution of plant communities. Biol. Rev. 2014, 90, 297–313. [Google Scholar] [CrossRef] [PubMed]
- Kuebbing, S.E.; Nuñez, M.A. Negative, neutral, and positive interactions among nonnative plants: Patterns, processes, and management implications. Glob. Chang. Biol. 2014, 21, 926–934. [Google Scholar] [CrossRef] [PubMed]
- Carboni, M.; Livingstone, S.W.; Isaac, M.E.; Cadotte, M.W. Invasion drives plant diversity loss through competition and ecosystem modification. J. Ecol. 2021, 109, 3587–3601. [Google Scholar] [CrossRef]
- Reijers, V.C.; van den Akker, M.; Cruijsen, P.M.J.M.; Lamers, L.P.M.; van der Heide, T. Intraspecific facilitation explains the persistence of Phragmites australis in modified coastal wetlands. Ecosphere 2019, 10, e02842. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, J.; Zhang, R.; Shen, Y. Analysis of remote sensing images for vegetation succession on tidal salt marsh in Jiangsu. J. Ecol. Rural. Environ. 2001, 17, 39–41. (In Chinese) [Google Scholar]
- He, Q.; Bertness, M.D. Extreme stresses, niches, and positive species interactions along stress gradients. Ecology 2014, 95, 1437–1443. [Google Scholar] [CrossRef] [PubMed]
- Berthon, K. How do native species respond to invaders? Mechanistic and trait-based perspectives. Biol. Invasions 2015, 17, 2199–2211. [Google Scholar] [CrossRef]
- Larsen, L.G. Multiscale flow-vegetation-sediment feedbacks in low-gradient landscapes. Geomorphology 2019, 334, 165–193. [Google Scholar] [CrossRef]
- Kim, S.; Yu, C.; Ruesink, J.; Hong, J.-S. Vertical distribution of the salt marsh invader Spartina alterniflora and native halophytes on the west coast of Korea in relation to tidal regimes. Aquat. Invasions 2023, 18, 331–349. [Google Scholar] [CrossRef]
- Brusati, E.D.; Grosholz, E.D. Native and Introduced Ecosystem engineers produce contrasting effects on estuarine infaunal communities. Biol. Invasions 2006, 8, 683–695. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Represented Year | Method | Number of Samples | Actual Sampling Years | Cited Sources |
|---|---|---|---|---|
| 1987 | Literature search | 6 | 1986 | [36] |
| 1997 | Literature search | 5 | 1996 | [37] |
| 2007 | Literature search | 22 | 2006–2008 | [38,39] |
| 2012 | Literature search and field sampling | 34 | 2011–2012 | [40] |
| 2017 | Field sampling | 21 | 2017 | / |
| 2022 | Field sampling | 27 | 2023 | / |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, Y.; Qiu, C.; Liu, H.; Li, Y.; Wang, C.; Wang, G.; Su, M.; He, C. How Do Invasive Species Influence Biotic and Abiotic Factors Drive Vegetation Success in Salt Marsh Ecosystems? Land 2025, 14, 1523. https://doi.org/10.3390/land14081523
Zhou Y, Qiu C, Liu H, Li Y, Wang C, Wang G, Su M, He C. How Do Invasive Species Influence Biotic and Abiotic Factors Drive Vegetation Success in Salt Marsh Ecosystems? Land. 2025; 14(8):1523. https://doi.org/10.3390/land14081523
Chicago/Turabian StyleZhou, Yong, Chunqi Qiu, Hongyu Liu, Yufeng Li, Cheng Wang, Gang Wang, Mengyuan Su, and Chen He. 2025. "How Do Invasive Species Influence Biotic and Abiotic Factors Drive Vegetation Success in Salt Marsh Ecosystems?" Land 14, no. 8: 1523. https://doi.org/10.3390/land14081523
APA StyleZhou, Y., Qiu, C., Liu, H., Li, Y., Wang, C., Wang, G., Su, M., & He, C. (2025). How Do Invasive Species Influence Biotic and Abiotic Factors Drive Vegetation Success in Salt Marsh Ecosystems? Land, 14(8), 1523. https://doi.org/10.3390/land14081523