
Potentials and Limitations of Fluviomarine Pollen Records to Reconstruct Spatiotemporal Changes in Coastal Ecosystems During the Holocene: A Case of Study from Ría de Vigo (NW Iberia)

 , , , , and
, , , , and
Abstract
1. Introduction
2. Regional Setting
3. Materials and Methods
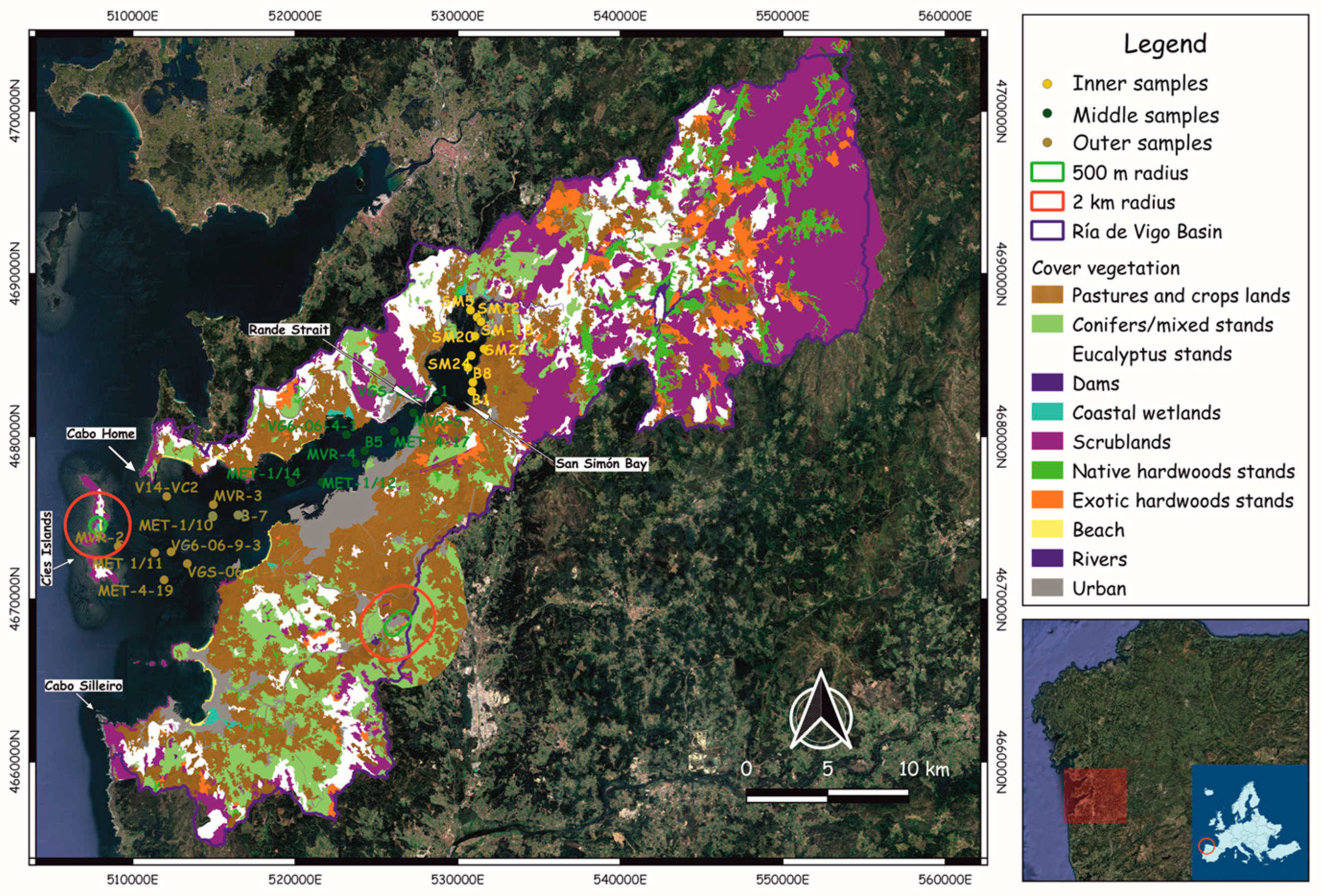
3.1. Pollen Samples
3.2. Chemical Treatments and Pollen Identification
3.3. Pollen Representation
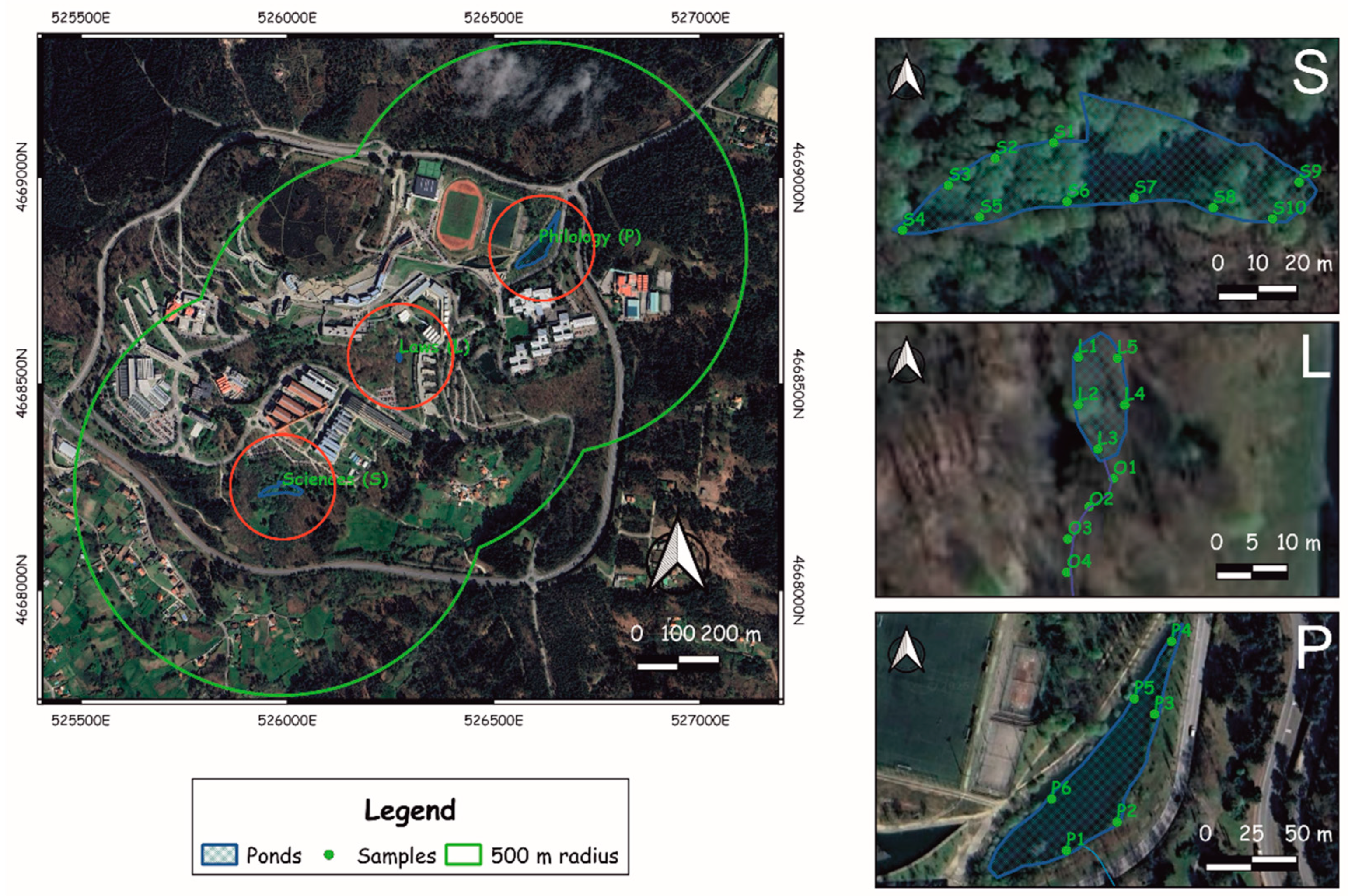
3.4. Hydrological Model of Lagoa dos Nenos
3.5. Modern Vegetation Cover
3.6. Numerical Analyses
4. Results
4.1. Vegetation Cover in the Ría de Vigo Basin
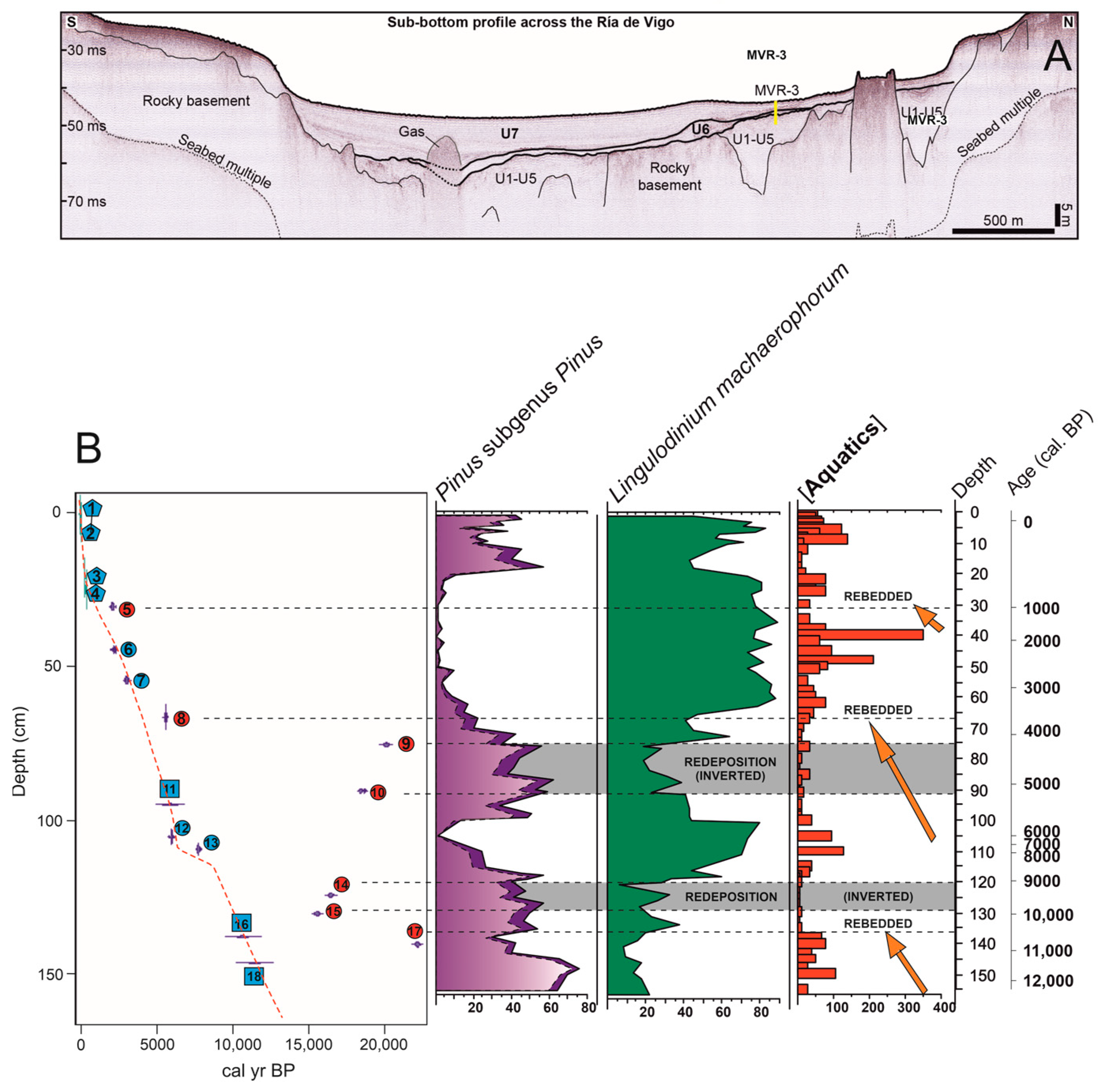
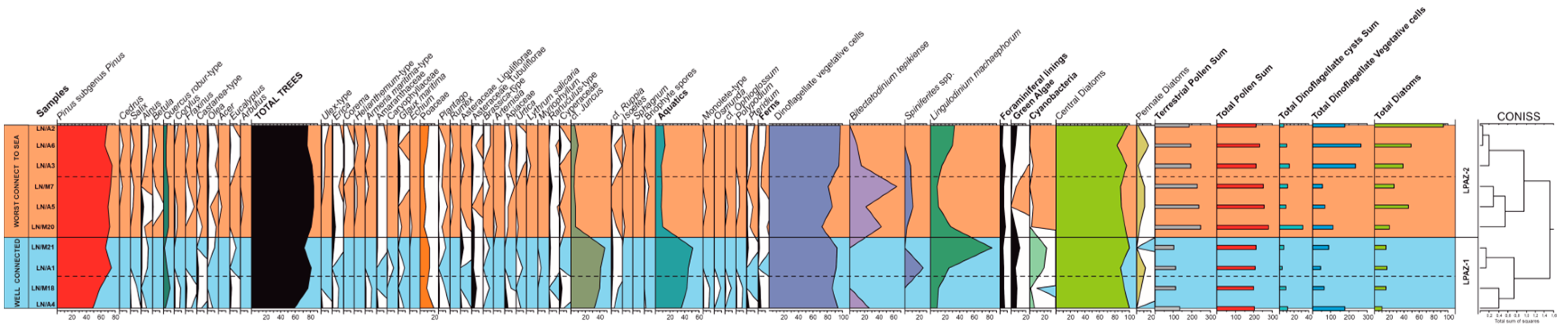
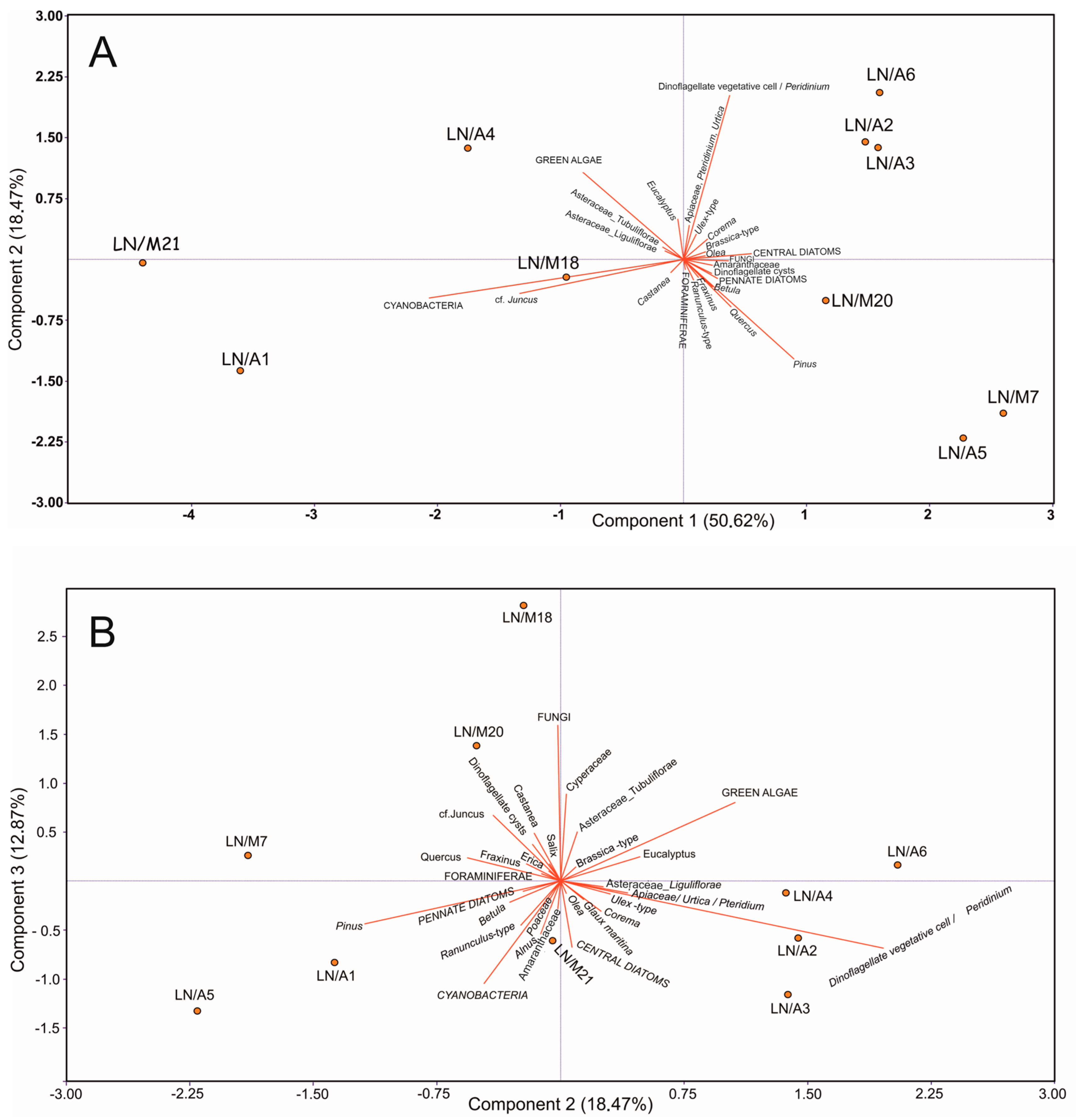
4.2. Pollen Sedimentation in the Open Coastal Lagoon System
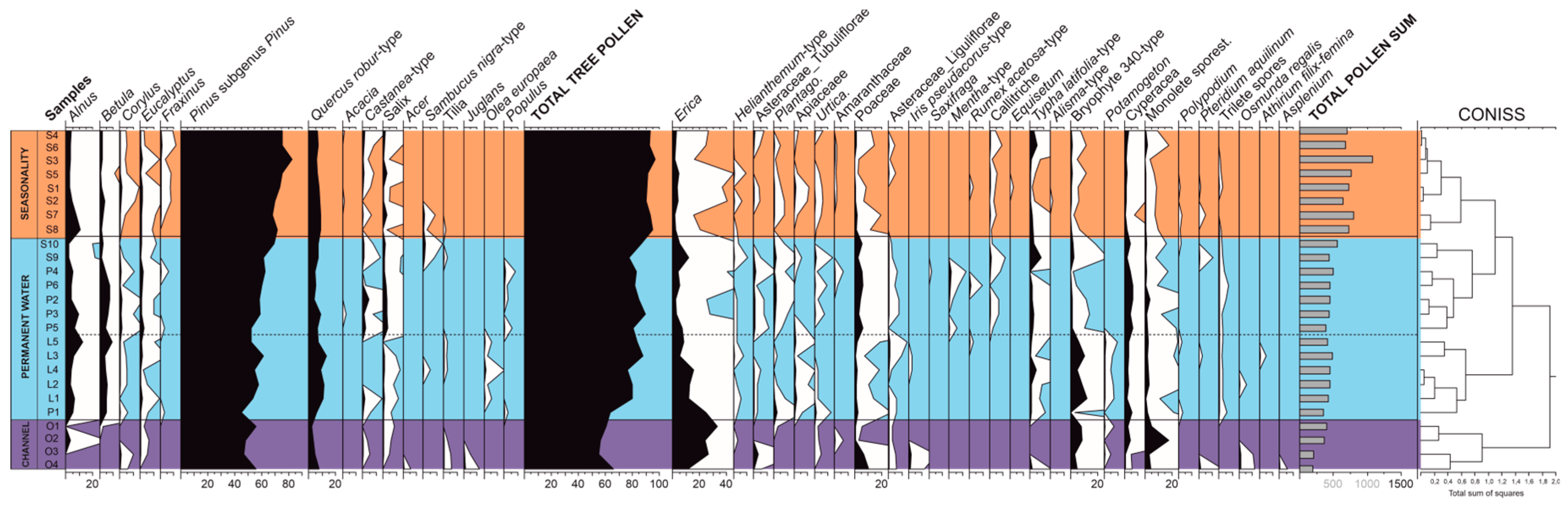
4.3. Pollen Sedimentation in the Upland Ponds
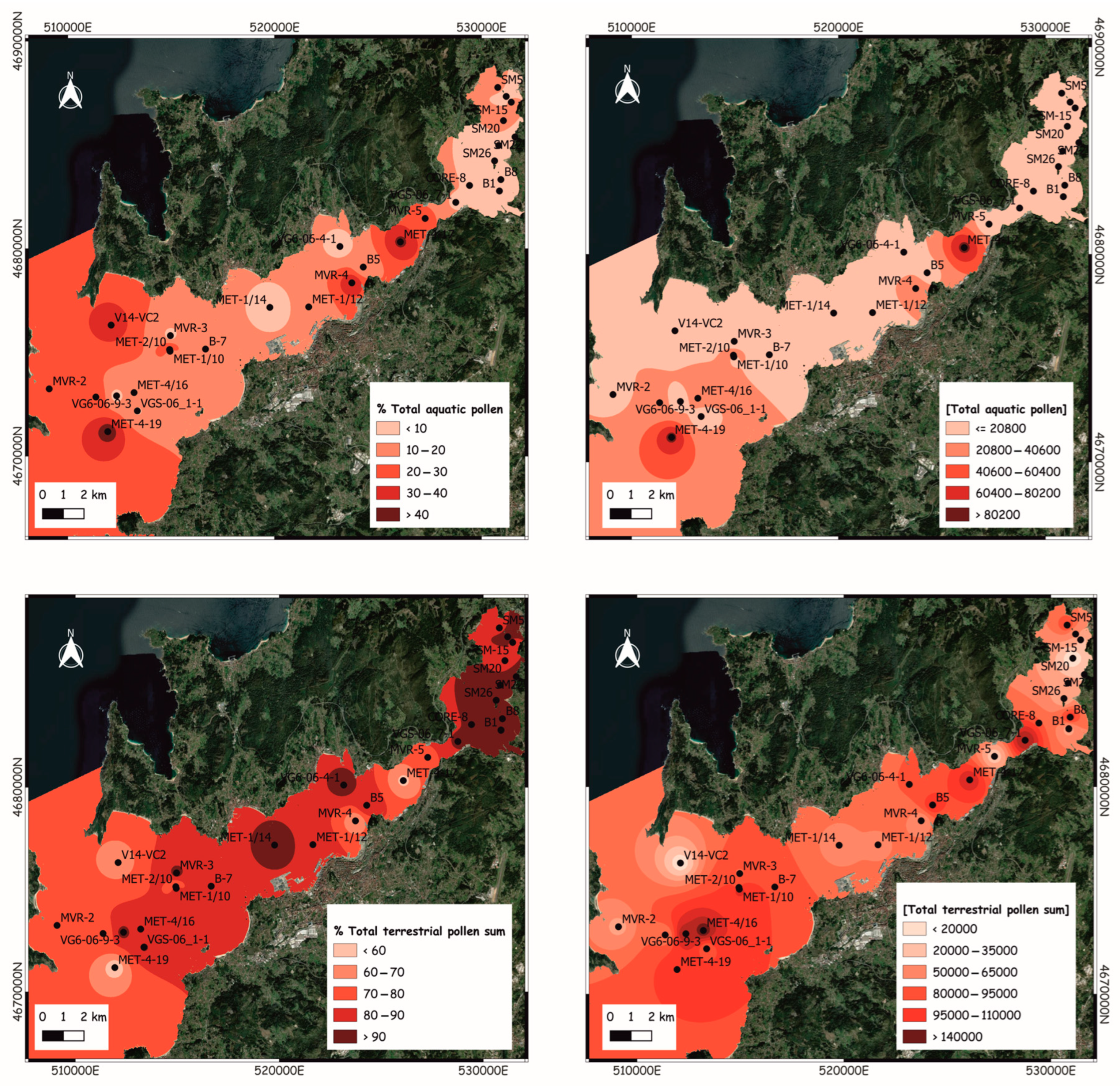
4.4. Pollen Sedimentation in the Modern Subtidal Seabed
5. Discussion
5.1. Pollen Evidence in the Ría de Vigo Basin: Differences Across Sedimentary Environments
5.2. Pollen Samples vs. Vegetation Types: Comparison Between Different Sedimentary Systems
5.3. Pollen Representation in the Coastal Lagoon vs. Vegetation Cover
5.4. Pollen Representation in Upland Ponds vs. Vegetation Cover
5.5. Pollen Representation in Modern Seabed Sediments vs. Vegetation Cover
5.6. Lessons for the Interpretation of Holocene Pollen Records from Shallow Seabed Sediments
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Johnsen, S.J.; Clausen, H.B.; Dansgaard, W.; Fuhrer, K.; Gundestrup, N.; Hammer, C.U.; Iversen, P.; Jouzel, J.; Stauffer, B.; Steffensen, J.P. Irregular Glacial Interstadials Recorded in a New Greenland Ice Core. Nature 1992, 359, 311–313. [Google Scholar] [CrossRef]
- deMenocal, P.; Ortiz, J.; Guilderson, T.; Sarnthein, M. Coherent High- and Low-Latitude Climate Variability During the Holocene Warm Period. Science 2000, 288, 2198–2202. [Google Scholar] [CrossRef]
- Muñoz Sobrino, C.; Ramil-Rego, P.; Gómez-Orellana, L.; Varela, R.A.D. Palynological Data on Major Holocene Climatic Events in NW Iberia. Boreas 2005, 34, 381–400. [Google Scholar] [CrossRef]
- Roucoux, K.H.; Shackleton, N.J.; de Abreu, L.; Schönfeld, J.; Tzedakis, P.C. Combined Marine Proxy and Pollen Analyses Reveal Rapid Iberian Vegetation Response to North Atlantic Millennial-Scale Climate Oscillations. Quat. Res. 2001, 56, 128–132. [Google Scholar] [CrossRef]
- Muñoz Sobrino, C.; Heiri, O.; Hazekamp, M.; Van der Velden, D.; Kirilova, E.P.; García-Moreiras, I.; Lotter, A.F. New Data on the Lateglacial Period of SW Europe: A High Resolution Multiproxy Record from Laguna de La Roya (NW Iberia). Quat. Sci. Rev. 2013, 80, 58–77. [Google Scholar] [CrossRef]
- Morales-Molino, C.; García-Antón, M.; Postigo-Mijarra, J.M.; Morla, C. Holocene Vegetation, Fire and Climate Interactions on the Westernmost Fringe of the Mediterranean Basin. Quat. Sci. Rev. 2013, 59, 5–17. [Google Scholar] [CrossRef]
- Iriarte-Chiapusso, M.J.; Sobrino, C.M.; Gómez-Orellana, L.; Hernández-Beloqui, B.; García-Moreiras, I.; Rodriguez, C.F.; Heiri, O.; Lotter, A.F.; Ramil-Rego, P. Reviewing the Lateglacial–Holocene Transition in NW Iberia: A Palaeoecological Approach Based on the Comparison between Dissimilar Regions. Quat. Int. 2016, 403, 211–236. [Google Scholar] [CrossRef]
- van der Horst, A.; Tinner, W.; Ezquerra, F.J.; Gobet, E.; Lotter, A.F.; Morellón, M.; Sobrino, C.M.; Niffenegger, C.; Schwörer, C.; Szidat, S. Late-Glacial and Holocene Shifts in the Mountain Landscapes of the Cantabrian Range (Northern Spain) in Response to Changing Climate, Fire Occurrence and Land Use. Quat. Sci. Rev. 2024, 342, 108899. [Google Scholar] [CrossRef]
- Gomez-Orellana, L.; Ramil-Rego, P.; Muñoz Sobrino, C. Una Nueva Secuencia Polínica y Cronológica Para El Depósito Pleistoceno de Mougás (NW de La Península Ibérica). Rev. Paléobiol. 1998, 17, 35–47. [Google Scholar]
- Gómez-Orellana, L.; Ramil-Rego, P.; Muñoz Sobrino, C. The Response of Vegetation at the End of the Last Glacial Period (MIS 3 and MIS 2) in Littoral Areas of NW I Beria. Boreas 2013, 42, 729–744. [Google Scholar] [CrossRef]
- Gómez-Orellana, L.; Ramil-Rego, P.; Ferreiro Da Costa, J.; Muñoz Sobrino, C. Holocene Environmental Change on the Atlantic Coast of NW Iberia as Inferred from the Ponzos Wetland Sequence. Boreas 2021, 50, 1131–1145. [Google Scholar] [CrossRef]
- Santos, L.; Bao, R.; Goñi, M.S. Pollen Record of the Last 500 Years from the Doninos Coastal Lagoon (NW Iberian Peninsula): Changes in the Pollinic Catchment Size versus Paleoecological Interpretation. J. Coast. Res. 2001, 17, 705–713. [Google Scholar]
- Danielsen, R. Palaeoecologial Development of the Quiaios–Mira Dunes, Northern-Central Littoral Portugal. Rev. Palaeobot. Palynol. 2008, 152, 74–99. [Google Scholar] [CrossRef]
- Costas, S.; Muñoz Sobrino, C.; Alejo, I.; Pérez-Arlucea, M. Holocene evolution of a rock-bounded barrier lagoon system, Cíes Islands, northwest Iberia. Earth Surfuface Process. Landf. 2009, 34, 1575–1586. [Google Scholar] [CrossRef]
- Galaz-Samaniego, C.A.; Peñalba, M.C.; Gardoki, J.; Cearreta, A.; Gómez-Arozamena, J.; Montoya-Laos, J.A.; Paz-Moreno, F.A.; Meling-López, A.E. Human-Induced Vegetation Dynamics Reconstructed through Pollen Analysis of a Recent Salt Marsh Sedimentary Sequence (Nalón Estuary, N Spain). Wetlands 2025, 45, 8. [Google Scholar] [CrossRef]
- Cartelle, V.; García-Gil, S. From a River Valley to a Ria: Evolution of an Incised Valley (Ría de Ferrol, North-west Spain) since the Last Glacial Maximum. Sedimentology 2019, 66, 1930–1966. [Google Scholar] [CrossRef]
- Muñoz Sobrino, C.; García-Gil, S.; Diez, J.B.; Iglesias, J. Palynological Characterization of Gassy Sediments in the Inner Part of Ría de Vigo (NW Spain). New Chronological and Environmental Data. Geo-Mar. Lett. 2007, 27, 289–302. [Google Scholar] [CrossRef]
- Muñoz Sobrino, C.; García-Gil, S.; Iglesias, J.; Martínez Carreño, N.; Ferreiro Da Costa, J.; Díaz Varela, R.A.; Judd, A. Environmental Change in the RÍa de Vigo, NW Iberia, since the Mid- HOlocene: New Palaeoecological and Seismic Evidence. Boreas 2012, 41, 578–601. [Google Scholar] [CrossRef]
- Muñoz Sobrino, C.; García-Moreiras, I.; Martínez-Carreño, N.; Cartelle, V.; Insua, T.L.; Ferreiro Da Costa, J.; Ramil-Rego, P.; Fernández Rodríguez, C.; Alejo, I.; García-Gil, S. Reconstruction of the Environmental History of a Coastal Insular System Using Shallow Marine Records: The Last Three Millennia of the Cíes Islands (Ría de Vigo, NW Iberia). Boreas 2016, 45, 729–753. [Google Scholar] [CrossRef]
- Valero, C.; Penaud, A.; Lambert, C.; Muriel, V.; David, O.; Leroux, E.; Stéphan, P.; Siano, R.; Ehrhold, A. Holocene paleoenvironmental reconstructions in western Brittany Bay of Brest: Part I—Understanding the spatial distribution of palynological records. Holocene 2025. [Google Scholar] [CrossRef]
- Traut, B.H. The Role of Coastal Ecotones: A Case Study of the Salt Marsh/Upland Transition Zone in California. J. Ecol. 2005, 93, 279–290. [Google Scholar] [CrossRef]
- Anderson, C.P.; Carter, G.A.; Waldron, M.C.B. Precise Elevation Thresholds Associated with Salt Marsh–Upland Ecotones along the Mississippi Gulf Coast. Ann. Am. Assoc. Geogr. 2022, 112, 1850–1865. [Google Scholar] [CrossRef]
- Sánchez-Goñi, M.F.; Desprat, S.; Fletcher, W.J.; Morales-Molino, C.; Naughton, F.; Oliveira, D.; Urrego, D.H.; Zorzi, C. Pollen from the Deep-Sea: A Breakthrough in the Mystery of the Ice Ages. Front. Plant Sci. 2018, 9, 38. [Google Scholar] [CrossRef]
- Ouyang, X.; Hao, X.; Culligan, N.; Dai, L.; Cheng, Z.; Li, S. Distribution of Suspended Sediments and Pollen in the Northern South China Sea: Implications for Pollen Source, Transport, and Deposition in Surface Ocean Waters. Cont. Shelf Res. 2021, 231, 104600. [Google Scholar] [CrossRef]
- Muñoz Sobrino, C.; Castro-Parada, A.; Cartelle, V.; Martínez-Carreño, N.; Delgado, C.; Cazás, N.; Lázaro, I.; García-Gil, S. Sediment Recycling during the Holocene Marine Transgression in Ría de Vigo (NW Iberia): Multiproxy Evidence and Environmental Implications. Quat. Sci. Rev. 2024, 344, 109006. [Google Scholar] [CrossRef]
- Muñoz Sobrino, C.; Cartelle, V.; Martínez-Carreño, N.; Ramil-Rego, P.; Gil, S.G. The Timing of the Postglacial Marine Transgression in the Ría de Ferrol (NW Iberia): A New Multiproxy Approach from Its Sedimentary Infill. Catena 2022, 209, 105847. [Google Scholar] [CrossRef]
- Martínez-Carreño, N.; García-Gil, S. Reinterpretation of the Quaternary Sedimentary Infill of the Ría de Vigo, NW Iberian Peninsula, as a Compound Incised Valley. Quat. Sci. Rev. 2017, 173, 124–144. [Google Scholar] [CrossRef]
- Guo, J.; Costa Jr, O.S.; Wang, Y.; Lin, W.; Wang, S.; Zhang, B.; Cui, Y.; Fu, H.; Zhang, L. Accumulation Rates and Chronologies from Depth Profiles of 210Pbex and 137Cs in Sediments of Northern Beibu Gulf, South China Sea. J. Environ. Radioact. 2020, 213, 106136. [Google Scholar] [CrossRef]
- Ferreira Cordeiro, N.G.; Dubert, J.; Nolasco, R.; Desmond Barton, E. Correction: Transient Response of the Northwestern Iberian Upwelling Regime. PLoS ONE 2018, 13, e0199806. [Google Scholar] [CrossRef]
- Picado, A.; Vaz, N.; Alvarez, I.; Dias, J.M. Modelling Coastal Upwelling off NW Iberian Peninsula: New Insights on the Fate of Phytoplankton Blooms. Sci. Total Environ. 2023, 874, 162416. [Google Scholar] [CrossRef]
- Martins, V.; Jouanneau, J.-M.; Weber, O.; Rocha, F. Tracing the Late Holocene Evolution of the NW Iberian Upwelling System. Mar. Micropaleontol. 2006, 59, 35–55. [Google Scholar] [CrossRef]
- Stuiver, M.; Braziunas, T.F. Modeling Atmospheric 14C Influences and 14C Ages of Marine Samples to 10,000 BC. Radiocarbon 1993, 35, 137–189. [Google Scholar] [CrossRef]
- Russell, N.; Cook, G.T.; Ascough, P.L.; Dugmore, A.J. Spatial variation in the MRE throughout the Scottish Post-Roman to late medieval period: North Sea values (500–1350 BP). Radiocarbon 2010, 52, 1166–1181. [Google Scholar] [CrossRef]
- Cook, G.T.; Ascough, P.L.; Bonsall, C.; Hamilton, W.D.; Russell, N.; Sayle, K.L.; Scott, E.M.; Bownes, J.M. Best Practice Methodology for 14C Calibration of Marine and Mixed Terrestrial/Marine Samples. Quat. Geochronol. 2015, 27, 164–171. [Google Scholar] [CrossRef]
- Álvarez-Iglesias, P.; Rubio, B.; Pérez-Arlucea, M. Reliability of Subtidal Sediments as “Geochemical Recorders” of Pollution Input: San Simón Bay (Ría de Vigo, NW Spain). Estuar. Coast. Shelf Sci. 2006, 70, 507–521. [Google Scholar] [CrossRef]
- Muñoz Sobrino, C.; Garcia-Moreiras, I.; Castro, Y.; Carreño, N.M.; de Blas, E.; Rodríguez, C.F.; Judd, A.; Garcia-Gil, S. Climate and Anthropogenic Factors Influencing an Estuarine Ecosystem from NW Iberia: New High Resolution Multiproxy Analyses from San Simón Bay (Ría de Vigo). Quat. Sci. Rev. 2014, 93, 11–33. [Google Scholar] [CrossRef]
- Kessler, J.D.; Reeburgh, W.S.; Valentine, D.L.; Kinnaman, F.S.; Peltzer, E.T.; Brewer, P.G.; Southon, J.; Tyler, S.C. A Survey of Methane Isotope Abundance (14 C,13 C,2 H) from Five Nearshore Marine Basins That Reveals Unusual Radiocarbon Levels in Subsurface Waters. J. Geophys. Res. 2008, 113, 2008JC004822. [Google Scholar] [CrossRef]
- Alperin, M.; Hoehler, T. The Ongoing Mystery of Sea-Floor Methane. Science 2010, 329, 288–289. [Google Scholar] [CrossRef]
- Tunno, I.; Zimmerman, S.R.; Brown, T.A.; Hassel, C.A. An Improved Method for Extracting, Sorting, and AMS Dating of Pollen Concentrates from Lake Sediment. Front. Ecol. Evol. 2021, 9, 668676. [Google Scholar] [CrossRef]
- Omori, T.; Yamada, K.; Kitaba, I.; Hori, T.; Nakagawa, T. Reliable Radiocarbon Dating of Fossil Pollen Grains: It Is Truly Possible. Quat. Geochronol. 2023, 77, 101456. [Google Scholar] [CrossRef]
- Cazás, N.; Castro-Parada, A.; Cartelle, V.; Muñoz Sobrino, C. Analyses of vegetation, pollen distribution and transportation patterns in surface sediments of fluviomarine systems (Ría de Vigo, NW Iberia): Implications for palaeoenvironmental studies. Veg. Hist. Archaeobotany 2025, Submitted. [Google Scholar]
- Faegri, K.; Iversen, J.; Krzywinski, K. Textbook of Pollen Analysis, 4th ed.; John Wiley and Sons Ltd.: London, UK, 1989; p. 328. [Google Scholar]
- Keil, R.G.; Hu, F.S.; Tsamakis, E.C.; Hedges, J.I. Pollen in marine sediments as an indicator of oxidation of organic matter. Nature 2014, 369, 639–641. [Google Scholar] [CrossRef]
- Traverse, A. Paleopalynology, 2nd ed.; Springer: Dordrecht, The Nethelands, 2008; p. 813. ISBN 978-1-4020-5609-3. [Google Scholar]
- Hopkins, J.A.; McCarthy, F.M.G. Oxidation and the palynological record. Palynology 2002, 26, 266–267. [Google Scholar]
- Li, Y.; Wang, N.; Li, Z.; Zhang, C.; Zhou, X. Reworking Effects in the Holocene Zhuye Lake Sediments: A Case Study by Pollen Concentrates AMS 14C Dating. Sci. China Earth Sci. 2012, 55, 1669–1678. [Google Scholar] [CrossRef]
- Krüger, S.; Damrath, M. In Search of the Bølling-Oscillation: A New High Resolution Pollen Record from the Locus Classicus Lake Bølling, Denmark. Veget Hist Archaeobot 2020, 29, 189–211. [Google Scholar] [CrossRef]
- Pawłowska, J.; Zajączkowski, M.; Łącka, M.; Lejzerowicz, F.; Esling, P.; Pawlowski, J. Palaeoceanographic Changes in Hornsund Fjord (Spitsbergen, Svalbard) over the Last Millennium: New Insights from Ancient DNA. Clim. Past 2016, 12, 1459–1472. [Google Scholar] [CrossRef]
- Serge, M.-A.; Mazier, F.; Fyfe, R.; Gaillard, M.-J.; Klein, T.; Lagnoux, A.; Galop, D.; Githumbi, E.; Mindrescu, M.; Nielsen, A.B. Testing the Effect of Relative Pollen Productivity on the REVEALS Model: A Validated Reconstruction of Europe-Wide Holocene Vegetation. Land 2023, 12, 986. [Google Scholar] [CrossRef]
- Castro-Parada, A.; Sobrino, C.M. Variations in Modern Pollen Distribution in Sediments from Nearby Upland Lakes: Implications for the Interpretation of Paleoecological Data. Rev. Palaeobot. Palynol. 2022, 306, 104765. [Google Scholar] [CrossRef]
- Dai, L.; Weng, C.; Lu, J.; Mao, L. Pollen Quantitative Distribution in Marine and Fluvial Surface Sediments from the Northern South China Sea: New Insights into Pollen Transportation and Deposition Mechanisms. Quat. Int. 2014, 325, 136–149. [Google Scholar] [CrossRef]
- Luo, C.; Chen, M.; Xiang, R.; Liu, J.; Zhang, L.; Lu, J.; Yang, M. Modern Pollen Distribution in Marine Sediments from the Northern Part of the South China Sea. Mar. Micropaleontol. 2014, 108, 41–56. [Google Scholar] [CrossRef]
- Yedema, Y.W.; Donders, T.; Peterse, F.; Sangiorgi, F. Dinoflagellate Cyst and Pollen Assemblages as Tracers for Marine Productivity and River Input in the Northern Gulf of Mexico. J. Micropalaeontol. 2023, 42, 257–276. [Google Scholar] [CrossRef]
- Rodríguez Guitián, M.; Ramil-Rego, P. Fitogeografía de Galicia (NW Ibérico): Análisis Histórico y Nueva Propuesta Corológica. Recur. Rurais 2008, 4, 19–50. [Google Scholar] [CrossRef]
- Meteogalicia. Unidade de Observación e Predicción Meteorolóxica. 2021. Available online: https://www.meteogalicia.gal/observacion/estacionshistorico/historico.action?idEst=10161 (accessed on 9 January 2025).
- Rodríguez Guitián, M.; Ramil-Rego, P. Clasificaciones Climáticas Aplicadas a Galicia: Revisión Desde Una Perspectiva Biogeográfica. Recur. Rurais 2007, 3, 31–53. [Google Scholar] [CrossRef]
- García-Moreiras, I.; Pospelova, V.; García-Gil, S.; Sobrino, C.M. Climatic and Anthropogenic Impacts on the Ría de Vigo (NW Iberia) over the Last Two Centuries: A High-Resolution Dinoflagellate Cyst Sedimentary Record. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2018, 504, 201–218. [Google Scholar] [CrossRef]
- Moore, P.D.; Webb, J.A.; Collinson, M.E. Pollen Analysis, 2nd ed.; Blackwell Scientific Publications: Oxford, UK, 1991. [Google Scholar]
- Valdés, B.; Díez, M.J.; Fernandez, I. Atlas Polínico de Andalucía Occidental; Instituto de Desarrollo Regional de la Universidad de Sevilla, Excma; Diputación de Cádiz: Sevilla, España, 1987. [Google Scholar]
- Reille, M. Pollen et Spores d’Europe et d’Afrique du Nord, Supplément 1, 1st ed.; Laboratoire de Botanique Historique et Palynologie: Marseille, France, 1995. [Google Scholar]
- Reille, M. Pollen et Spores d’Europe et d’Afrique du Nord, Supplément, 2nd ed.; Laboratoire de Botanique Historique et Palynologie: Marseille, France, 1998. [Google Scholar]
- Reille, M. Pollen et Spores d’Europe et d’Afrique du Nord, 2nd ed.; Laboratoire de Botanique Historique et Palynologie: Marseille, France, 1999. [Google Scholar]
- Grimm, E. TILIA and TILIA graph: PC spreadsheets and graphics software for pollen data. INQUA Comm. Study Holocene Work. Group Data Handl. Methods Newsl. 1990, 4, 5–7. [Google Scholar]
- López, F.; Vigo, M.A. Estudio Básico. Gestión de la Laguna Costera “Lago dos Nenos” en el Archipiélago de Cíes; Xunta de Galicia: Vigo, Pontevedra, 2016. [Google Scholar]
- PNOA-LIDAR. Plan Nacional de Ortofotografía Aérea. Plan Nacional de Observación del Territorio. Instituto Geográfico Nacional 2015–2021. Available online: https://pnoa.ign.es/web/portal/pnoa-lidar/estado-del-proyecto (accessed on 14 February 2025).
- QGIS.org. QGIS 3.40.2 ’Bratislava’. GIS Geographic Information System. QGIS Association 2024. Available online: http://www.qgis.org (accessed on 25 November 2024).
- IET. Mapas de Usos e Coberturas do Solo 2017. Available online: http://descargas.xunta.es/fb5738cd-788f-4b8d-ac66-96abbacccede1495189290462 (accessed on 7 December 2021).
- Hammer, Ǿ.C.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing 2021, Vienna, Austria. Available online: https://www.R-project.org/ (accessed on 10 December 2024).
- Brown, A.G.; Carpenter, R.G.; Walling, D.E. Monitoring Fluvial Pollen Transport, Its Relationship to Catchment Vegetation and Implications for Palaeoenvironmental Studies. Rev. Palaeobot. Palynol. 2007, 147, 60–76. [Google Scholar] [CrossRef]
- Quamar, M.F.; Mishra, A.K.; Mohanty, R.B.; Kar, R. Implications of Pinus L. Pollen Abundance for Reconstructing the Holocene Palaeoclimate from the Himalayas, India. Rev. Palaeobot. Palynol. 2024, 326, 105130. [Google Scholar] [CrossRef]
- Havinga, A.J. A 20-Year Experimental Investigation into the Differential Corrosion Susceptibility of Pollen and Spores in Various Soil Types. Pollen Spores 1984, 26, 541–557. [Google Scholar]
- Rodríguez-Rajo, F.J.; Jato, M.V.; Seijo, M.C. El Polen de Eucalyptus y su incidencia en la atmósfera de Vigo (NO España). Acta Botánica Malacit. 2001, 26, 99–110. [Google Scholar] [CrossRef]
- Ollerton, J.; Alarcón, R.; Waser, N.M.; Price, M.V.; Watts, S.; Cranmer, L.; Hingston, A.; Peter, C.I.; Rotenberry, J. A Global Test of the Pollination Syndrome Hypothesis. Ann. Bot. 2009, 103, 1471–1480. [Google Scholar] [CrossRef] [PubMed]
- Nunes Morgado, L.; Gonçalves-Esteves, V.; Resendes, R.; Mateus Ventura, M.A. A pollen inventory of endemic species from the Azores archipelago, Portugal. Palynology 2018, 42, 273–289. [Google Scholar] [CrossRef]
- Solomon, S.; Mudie, P.J.; Cranston, R.; Hamilton, T.; Thibaudeau, S.A.; Collins, E.S. Characterisation of marine and lacustrine sediments in a drowned thermokarst embayment, Richards Island, Beaufort Sea, Canada. Int. Jounal Earth Sci. 2000, 89, 503–521. [Google Scholar] [CrossRef]
- Mudie, P.J.; Yanko-Hombach, V.V.; Mudryk, I. Palynomorphs in surface sediments of the North-Western Black Sea as indicators of environmental conditions. Quat. Int. 2021, 590, 122–145. [Google Scholar] [CrossRef]
- Lambert, C.; Vidal, M.; Penaud, A.; Combourieu-Nebout, N.; Lebreton, V.; Ragueneau, O.; Gregoire, G. Modern palynological record in the Bay of Brest (NW France): Signal calibration for palaeo-reconstructions. Rev. Palaeobot. Palynol. 2017, 244, 13–25. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Altitude m asl/ Depth m NMMA | Environment of Sedimentation | Group of Samples | Subgroup of Samples | Subgroup of Samples | Sampling Year | References |
|---|---|---|---|---|---|---|---|
| LN/A1 | 0 | Lagoon | Lagoon | 2016 | This study | ||
| LN/A2 | 0 | Lagoon | Lagoon | 2016 | This study | ||
| LN/A3 | −0.5 | Lagoon | Lagoon | 2016 | This study | ||
| LN/A4 | −0.5 | Lagoon | Lagoon | 2016 | This study | ||
| LN/A5 | 0.5 | Lagoon | Lagoon | 2016 | This study | ||
| LN/A6 | 0 | Lagoon | Lagoon | 2016 | This study | ||
| LN/M7 | 0.5 | Lagoon | Lagoon | 2016 | This study | ||
| LN/M18 | 0.5 | Lagoon | Lagoon | 2016 | This study | ||
| LN/M20 | −0.25 | Lagoon | Lagoon | 2016 | This study | ||
| LN/M21 | 0 | Lagoon | Lagoon | 2016 | This study | ||
| P1 | 458 | Upland Pond | Upland | Pond P | 2019 | 49 | |
| P2 | 458 | Upland Pond | Upland | Pond P | 2019 | 49 | |
| P3 | 458 | Upland Pond | Upland | Pond P | 2019 | 49 | |
| P4 | 458 | Upland Pond | Upland | Pond P | 2019 | 49 | |
| P5 | 458 | Upland Pond | Upland | Pond P | 2019 | 49 | |
| P6 | 458 | Upland Pond | Upland | Pond P | 2019 | 49 | |
| L1 | 433 | Upland Pond | Upland | Pond L | 2019 | 49 | |
| L2 | 433 | Upland Pond | Upland | Pond L | 2019 | 49 | |
| L3 | 433 | Upland Pond | Upland | Pond L | 2019 | 49 | |
| L4 | 433 | Upland Pond | Upland | Pond L | 2019 | 49 | |
| L5 | 433 | Upland Pond | Upland | Pond L | 2019 | 49 | |
| O1 | 432 | Drainage channel | Upland | Channel | 2019 | 49 | |
| O2 | 432 | Drainage channe | Upland | Channel | 2019 | 49 | |
| O3 | 431 | Drainage channe | Upland | Channel | 2019 | 49 | |
| O4 | 431 | Drainage channe | Upland | Channel | 2019 | 49 | |
| S1 | 387 | Upland Pond | Upland | Pond S | 2019 | 49 | |
| S2 | 387 | Upland Pond | Upland | Pond S | 2019 | 49 | |
| S3 | 387 | Upland Pond | Upland | Pond S | 2019 | 49 | |
| S4 | 387 | Upland Pond | Upland | Pond S | 2019 | 49 | |
| S5 | 387 | Upland Pond | Upland | Pond S | 2019 | 49 | |
| S6 | 387 | Upland Pond | Upland | Pond S | 2019 | 49 | |
| S7 | 387 | Upland Pond | Upland | Pond S | 2019 | 49 | |
| S8 | 387 | Upland Pond | Upland | Pond S | 2019 | 49 | |
| S9 | 387 | Upland Pond | Upland | Pond S | 2019 | 49 | |
| S10 | 387 | Upland Pond | Upland | Pond S | 2019 | 49 | |
| SM12 | 3 | Seabed sediment | Subtidal | Inner | 2012 | 40 | |
| SM-15 | 3 | Seabed sediment | Subtidal | Inner | 2012 | 40 | |
| SM5 | 3 | Seabed sediment | Subtidal | Inner | 2012 | 40 | |
| SM20 | 2 | Seabed sediment | Subtidal | Inner | 2012 | 40 | |
| SM22 | 2 | Seabed sediment | Subtidal | Inner | 2012 | 40 | |
| SM26 | −2 | Seabed sediment | Subtidal | Inner | 2012 | 40 | |
| B-8 (0-1) | −3 | Seabed sediment | Subtidal | Inner | 2012 | 40 | |
| B-1 (0-1) | −4 | Seabed sediment | Subtidal | Inner | 2012 | 40 | |
| VG6-06-4-1 (0-2) | −13 | Seabed sediment | Subtidal | Middle | Outer Rande Strait | 2006 | 40 |
| B-5 (0-1) | −18 | Seabed sediment | Subtidal | Middle | Outer Rande Strait | 2012 | 40 |
| MET4/17 (0-1) | −20 | Seabed sediment | Subtidal | Middle | Outer Rande Strait | 2011 | 40 |
| MVR-4 (0-1) | −22 | Seabed sediment | Subtidal | Middle | Outer Rande Strait | 2011 | 40 |
| VGS-06_7-1 (0-2) | −24 | Seabed sediment | Subtidal | Middle | Outer Rande Strait | 2006 | 40 |
| MET-1-14 (0-2) | −26 | Seabed sediment | Subtidal | Middle | Outer Rande Strait | 2010 | 40 |
| MET-1/12 (0-1) | −28 | Seabed sediment | Subtidal | Middle | Outer Rande Strait | 2010 | 40 |
| MVR-5 (0-1) | −28 | Seabed sediment | Subtidal | Middle | Outer Rande Strait | 2011 | 40 |
| MVR-3 (0-1) | −30 | Seabed sediment | Subtidal | Outer | Outer Rande Strait | 2011 | 24 |
| MVR-2 (0-1) | −33 | Seabed sediment | Subtidal | Outer | Outer Rande Strait | 2011 | 40 |
| V14-VC2 (0-1) | −35 | Seabed sediment | Subtidal | Outer | Outer Rande Strait | 2014 | 40 |
| MET-1/11 (0-1) | −38 | Seabed sediment | Subtidal | Outer | Outer Rande Strait | 2010 | 40 |
| MET-2/10 (0-1) | −38 | Seabed sediment | Subtidal | Outer | Outer Rande Strait | 2010 | 40 |
| VGS-06_1-1 (0-2) | −38 | Seabed sediment | Subtidal | Outer | Outer Rande Strait | 2006 | 40 |
| VGS-06-9-3 (0-2) | −38 | Seabed sediment | Subtidal | Outer | Outer Rande Strait | 2006 | 40 |
| MET-1-10 (0-1) | −40 | Seabed sediment | Subtidal | Outer | Outer Rande Strait | 2010 | 40 |
| MET-4/19 (0-1) | −40 | Seabed sediment | Subtidal | Outer | Outer Rande Strait | 2010 | 40 |
| MET-4/16 (0-1) | −42 | Seabed sediment | Subtidal | Outer | Outer Rande Strait | 2011 | 40 |
| Pond S | Pond L | Pond P | |
|---|---|---|---|
| Surface covered by tree-canopy (%) | 80 | 100 | 5 |
| Altitude (m a.s.l.) | 387 | 433 | 458 |
| Shape | Elongated | Elliptic | Irregular |
| Total Surface (m2) | 2500 | 100 | 4500 |
| Position in the sub-basin | Low | Intermediate | High |
| Seasonality | Marked | None | Little |
| Active Outflow | Winter and spring | All the year | None |
| Local vegetation | Typha angustifolia L., Lemna minor L., Callitriche stagnalis Scop., Alisma plantago-aquatica L., Juncus effusus L., Iris pseudacorus L., Alnus glutinosa (L.) Gaertn., Betula pubescens Ehrh., Fraxinus angustifolia Vahl, Salix atrocinerea Brot. | Bryophytes, Osmunda regalis L., Dryopteris spp., Athyrium filix-femina (L.) Roth, Typha angustifolia L., Juncus effusus L. Potamogeton natans L. | Salix alba L., Salix atrocinerea Brot., Betula alba L., Fraxinus excelsior L., Alnus glutinosa (L.) Gaertn., Typha angustifolia L., Phragmites australis (Cav.) Trin. Former Steud., Juncus effusus L., Taxodium distichum (L.) |
| Vegetation Units | Pollen Types Included |
|---|---|
| Scrublands | Calluna vulgaris; Erica; Corema; Helianthemum-type; Hedera; Ulex-type |
| Pastures and crops lands | Vitis; Alchemilla; Anchusa; Armeria maritima-type; Artemisia; Asphodelus; Asteraceae_Liguliflorae; Asteraceae_Tubuliflorae; Brassicaceae; Caryophyllaceae; Centaurea scabiosa; Campanula-type; Amaranthaceae; Echium; Glaux maritima; Geranium-type; Humulus lupulus-type; Allium-type; Lotus-type; Mentha-type; Pentaglotis sempervirens; Plantago; Polygonum amphibium; Potentilla-type; Rumex acetosa-type; Urtica; Poaceae; Cerealia-type; Zea; Sedum; Apiaceae; Umbilicus; Callitriche, Lythrum salicara |
| Eucalyptus stands | Eucalyptus |
| Coastal wetlands | Cyperaceae; Isoetes; Myriophyllum; Ranunculus-type; Typha latifolia; Sphagnum; Iris pseudacorus-type; cf. Juncus; Anthoceros; cf. Ruppia |
| Conifers/mixed stands | Pinus subgenus Pinus |
| Exotic hardwoods stands | Acacia |
| Native hardwoods stands | Alnus; Betula; Castanea; Corylus; Platanus; Fraxinus; Ilex-type; Juniperus –type; Arbutus; Olea; Populus; Quercus robur-type; Salix; Sambucus nigra-type; Tilia |
| Lagoa dos Nenos | Upland Ponds | Seabed Samples | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Pollen | Coverage | Pollen | Coverage | Pollen | Coverage | |||||||||||
| Vegetation Units | % | <500 m | <2 km | % S | % L | % P | % O | % ALL | <500 m | <2 km | % All | % Inner | % Middle | % Outer | % Out R. Strait | RdV Basin |
| Scrublands | 2.02 | 40.49 | 56.91 | 4.41 | 11.44 | 8.10 | 26.12 | 12.52 | 15.25 | 2.66 | 8.82 | 7.31 | 9.29 | 9.85 | 9.61 | 29.4 |
| Pastures and crops | 13.3 | 22.06 | 10.64 | 4,13 | 4.59 | 9.68 | 8.06 | 6.62 | 45.10 | 50.41 | 25.18 | 27.68 | 26.73 | 21.12 | 23.53 | 30.5 |
| Eucalyptus stands | 0.58 | 8.88 | 20.18 | 0.98 | 1.07 | 1.63 | 0.40 | 1.02 | 2.45 | 2.19 | 0.49 | 0.65 | 0.59 | 0.22 | 0.38 | 17.5 |
| Coastal wetlands | 22.79 | 15.43 | 3.10 | --- | --- | --- | --- | --- | --- | 16.40 | 8.11 | 18.23 | 22.87 | 20.88 | 0.3 | |
| Conifers/mixed stands | 54.11 | 13.14 | 9.16 | 72.89 | 57.66 | 57.52 | 57.31 | 61.35 | 31.20 | 43.31 | 37.08 | 39.24 | 34.19 | 37.82 | 36.26 | 12.6 |
| Native hardwood stands | 7.19 | --- | --- | 17.56 | 25.23 | 23.04 | 8.10 | 18.48 | 0.38 | 0.73 | 11.97 | 16.82 | 10.97 | 8.11 | 9.34 | 5.5 |
| Exotic hardwood stands | --- | --- | --- | 0.03 | --- | 0.04 | --- | 0.02 | 5.61 | 0.70 | 0.07 | 0.20 | --- | --- | --- | 4.3 |
| % Pollen | LN | Upland | P Pond | S Pond | L Pond | O Channel | All RdV | Inner | Middle | Outer | Out R. Strait | All 61 Samples |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Scrublands | <0.05 | <0.05 | 0.03 | <0.05 | <0.05 | 0.01 | <0.05 | <0.05 | <0.05 | <0.05 | <0.05 | <0.05 |
| Pastures and crops | <0.05 | <0.05 | <0.05 | <0.05 | <0.05 | 0.125 | <0.05 | <0.05 | <0.05 | <0.05 | <0.05 | <0.05 |
| Eucalyptus stands | 0.02 | <0.05 | <0.05 | <0.05 | 0.03 | 0.09 | <0.05 | <0.05 | 0.05 | 0.125 | <0.05 | <0.05 |
| Coastal wetlands | <0.05 | --- | --- | --- | --- | --- | <0.05 | <0.05 | 0.01 | <0.05 | <0.05 | <0.05 |
| Conifers/mixed stands | <0.05 | <0.05 | <0.05 | <0.05 | <0.05 | <0.05 | <0.05 | <0.05 | <0.05 | <0.05 | <0.05 | <0.05 |
| Exotic hardwood stands | <0.05 | 0.25 | 1.00 | 0.05 | --- | --- | 0.05 | 0.25 | --- | --- | --- | <0.05 |
| Native hardwood stands | <0.05 | <0.05 | <0.05 | <0.05 | <0.05 | 0.01 | <0.05 | 0.01 | <0.05 | <0.05 | <0.05 | <0.05 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castro-Parada, A.; Cazás, N.; Cartelle, V.; Ferreiro da Costa, J.; Martínez-Carreño, N.; García-Gil, S.; Muñoz Sobrino, C. Potentials and Limitations of Fluviomarine Pollen Records to Reconstruct Spatiotemporal Changes in Coastal Ecosystems During the Holocene: A Case of Study from Ría de Vigo (NW Iberia). Land 2025, 14, 540. https://doi.org/10.3390/land14030540
Castro-Parada A, Cazás N, Cartelle V, Ferreiro da Costa J, Martínez-Carreño N, García-Gil S, Muñoz Sobrino C. Potentials and Limitations of Fluviomarine Pollen Records to Reconstruct Spatiotemporal Changes in Coastal Ecosystems During the Holocene: A Case of Study from Ría de Vigo (NW Iberia). Land. 2025; 14(3):540. https://doi.org/10.3390/land14030540
Chicago/Turabian StyleCastro-Parada, Alberto, Nerea Cazás, Víctor Cartelle, Javier Ferreiro da Costa, Natalia Martínez-Carreño, Soledad García-Gil, and Castor Muñoz Sobrino. 2025. "Potentials and Limitations of Fluviomarine Pollen Records to Reconstruct Spatiotemporal Changes in Coastal Ecosystems During the Holocene: A Case of Study from Ría de Vigo (NW Iberia)" Land 14, no. 3: 540. https://doi.org/10.3390/land14030540
APA StyleCastro-Parada, A., Cazás, N., Cartelle, V., Ferreiro da Costa, J., Martínez-Carreño, N., García-Gil, S., & Muñoz Sobrino, C. (2025). Potentials and Limitations of Fluviomarine Pollen Records to Reconstruct Spatiotemporal Changes in Coastal Ecosystems During the Holocene: A Case of Study from Ría de Vigo (NW Iberia). Land, 14(3), 540. https://doi.org/10.3390/land14030540