Abstract
Grasslands differ in many types depending on the diversity of climatic conditions and substrates. Due to the great wealth of species found in semi-natural grasslands, they conserve an essential part of the biodiversity of the European Union (EEC 43/92), defined as habitats of community interest. Using the region of Calabria (southern Italy) as a case study, this study aims to evaluate how species assemblages and a set of indicators applied to them can be used to analyse and characterize the diversity, ecological features and conservation status of grassland habitats. Vegetation analysis was carried out using the phytosociological method, and habitat ecological characteristics were defined by Ellenberg’s ecological indicator adapted to the Italian flora. Multivariate analysis of the surveys by means of cluster analysis and Principal Components Analysis (PCA) made it possible to define eight habitat groups according to EEC Directive 43/92: 6110 Rocky or basophilous calcareous grasslands of the Alysso-Sedion albi, 6170 Alpine and subalpine calcareous grasslands, 6220*a Pseudo-steppe with grasses and annuals of the Thero-Brachypodietea dominated by a Lygeum spartum, 6220*b Pseudo-steppe with grasses and annuals of the Thero-Brachypodietea dominated by Hyparrhenia hirta, 6210* Semi-natural dry grasslands and scrub facies on calcareous substrates (Festuco-Bromometalia) (*important orchid sites), 6230* Species-rich Nardus grasslands, on siliceous substrates in mountain areas (and submountain areas, in Continental Europe), 6410 Molinia meadows on calcareous, peaty or clayey-siltladen soils (Molinion caeruleae), 6420 Mediterranean tall humid herb grasslands of the Molinio-Holoschoenion, 6430 Hydrophilous tall herb fringe communities of plains and of the montane to alpine levels. Temperature (T) and oisture (U) are the most statistically significant ecological factors in differentiating different habitat types. Analysis of diversity, assessed through Shannon and Evenness indices, showed that it is strongly correlated with habitat diversity as soil moisture increases. The study of ecological characteristics, correlated with biodiversity indices, helps to characterize grassland habitats, providing guidance for the implementation of active conservation policies for these habitats.
1. Introduction
Grasslands contain a high percentage of the world’s biodiversity, despite only a part of the continents being covered by natural or semi-natural grasslands [1,2,3]. These ecosystems account for 28% of natural and semi-natural habitats, ranking after forests in terms of importance [4]. In the Italian territory, grasslands cover about 6.2% of the surface, and about 3.6% is mapped as open grasslands by APAT [5].
Generally, semi-natural grassland habitats embedded in agricultural landscapes support floristic diversity and a variety of plant communities [6,7,8], being characterized by a high number of species, many of which are endemics, and are important ecosystems for maintaining biodiversity [9,10,11]. These habitats, therefore, have a high conservation value, particularly due to their high biodiversity and related ecosystem services [12,13]. The maintenance of grassland habitats and their associated species has been an important issue in nature conservation for decades [14]. Today, numerous studies analyse the ecological and floristic features of natural and semi-natural Palearctic grasslands (accessed on 6 November 2023). Understanding and analysing the ecological factors that influence biodiversity is essential for designing biodiversity management and conservation plans that are consistent with the floristic and ecological characteristics of habitats [15,16,17].
The remarkable species richness of grasslands is threatened across Europe due to global changes, fire and biological invasions [18,19,20,21,22,23,24]. The decline in the diversity and connectivity of grasslands in Europe has increasingly become a major issue for their conservation. Changes in land use have the greatest impact on the degradation and loss of different habitat types [25,26,27]. Over the last century, a continuous loss of species-rich grassland habitats, influenced by humans with conversion to intensive arable or grassland, or abandonment to evolve into forests, has been recognized in Europe [28,29]. In this perspective, beta-diversity, defined as the amount of turnover in species composition [30], is important as a measure of species diversity on a regional scale, to be used as a basis for conservation [31]. The conservation and sustainable use of a given territory must take into account models that consider biodiversity as a factor in ecosystem stability and functioning [32,33,34].
Some studies show that the species composition of grasslands varies to a different extent depending on environmental characteristics [35,36,37]. Various ecological factors affect the species diversity of grasslands habitats [38]; among them, the most important are climate [39,40], altitude and, above all, soil moisture conditions [41,42]. In particular, the humidity is one of the most important ecological factors that influence plant species, the diversity and the floristic composition of grasslands habitats, as highlighted by Zelnik et al. [43]; there have also been numerous research studies carried out in the last decade in Europe on wetlands and the role of ecological factors [44,45,46,47,48,49,50,51,52,53]. Some studies state that nutrient availability within the soil influences species richness in various plant communities [54,55].
Grazing management is another factor that influences the structure and function of these ecosystems [56,57,58], changing their plant diversity. Lozano et al. [59] suggest that grazing pressure has a negative effect on the presence of endemic species, whereas in alpine environments, grasslands with higher floristic diversity are generally associated with environments with forms of management that involve the removal biomass, such as mowing and grazing [60,61].
The use of ecological indices can point out anthropogenic impact and thus allow the degree of naturalness or degradation of a habitat to be assessed based on species composition [62].
This study aims to assess the conservation status of grassland habitats by analysing the diversity, naturalness, and ecological characteristics of the plant communities. The grassland spread in the Calabria region was used as a case study. In particular, the study aims to (1) identify the different habitat types through the species assemblage; (2) define the main ecological factors responsible for variation in the floristic composition of grassland habitats vegetation; (3) assess the biodiversity and naturalness of the different habitat types; (4) evaluate the relationship between species diversity and ecological factors. The results of this study can be useful and applied in the management of grassland habitats for biodiversity conservation purposes.
2. Materials and Methods
The study area was the Calabria region, which, as indicated in the Carta Natura [63], occupies 24,375 ha, or 2.4% of the regional territory. For the photointerpretation of the habitats, arc map 10.7 was used, with reference system WGS84 UTM Zone 33, (False_Easting: 500000,00000 False_Northing: 0.00000; Central_Meridian: 15.00000; Scale_Factor: 0.99960; Latitude_Of_Origin: 0.00000; Linear Unit: Kilometres). The analysis has affected all grassland habitats depicted on the nature map.
These habitats are distributed throughout the Calabrian territory, from sea level to the mountain range, affecting different environmental contexts characterized by different ecological conditions [27].
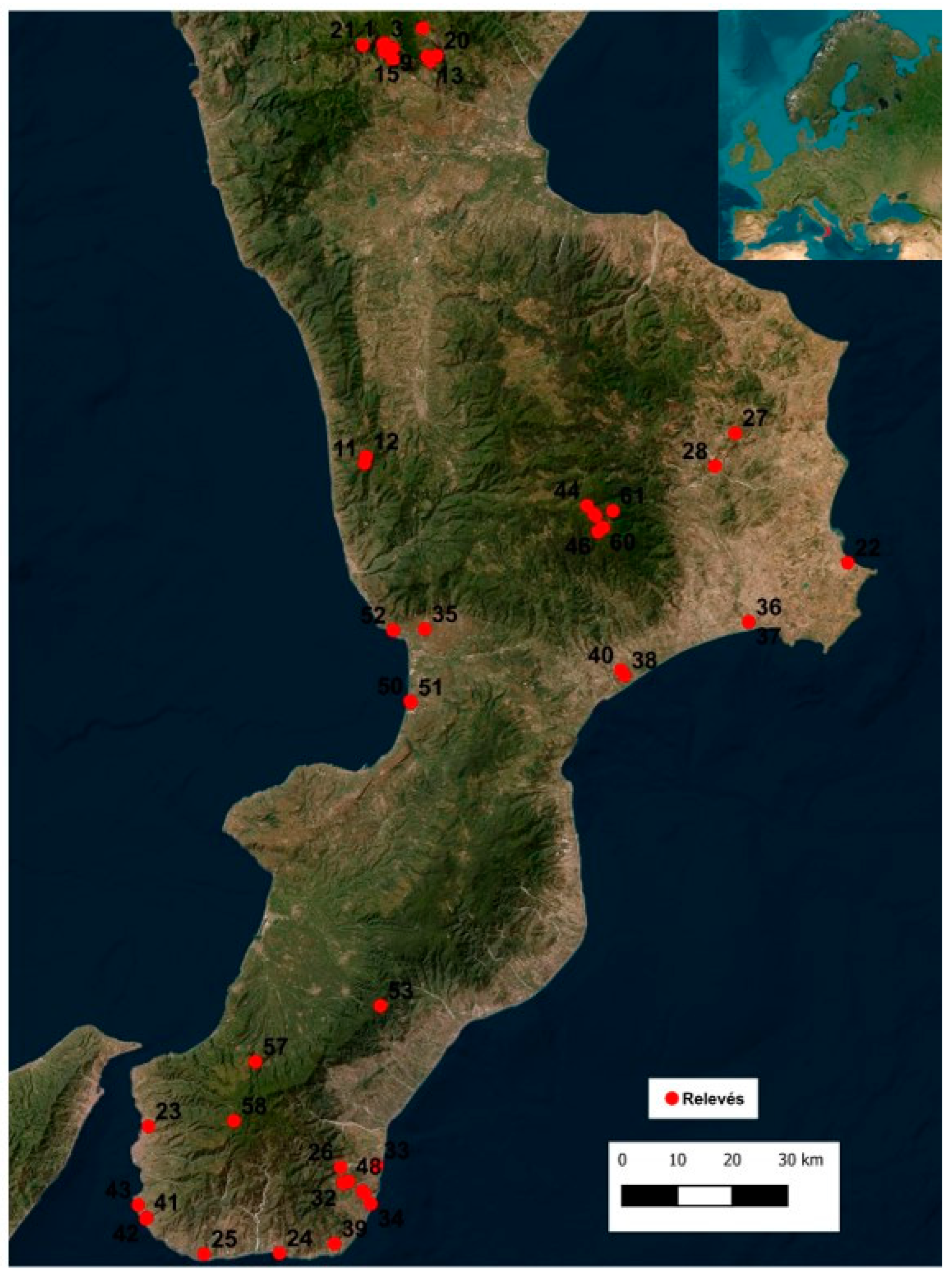
The analysis of grassland habitats was based on the vegetation relevés carried out in the period 2018–2021, using the phytosociological method of the Zurich-Montpellier school, considered to date the most suitable method for analysing vegetation [15,64,65]. A total of 61 unpublished relevés with 490 species were carried out (Figure 1) (Supplementary Materials Table S1). The matrix of phytosociological relevés was subjected to multivariate analysis to define coenological groups on an actual statistical basis. To this end, the abundance-dominance data of the individual species reported with the Braun-Blanquet scale were converted with the scale of numerical values proposed by Van der Mareel. The software used for the organization of raw data and subsequent statistical analyses was Microsoft Excel 2010; PAST version 4.13; R 4.1.1 R Core Team 2021.

Figure 1.
Study area: Calabria region. The red dots and numbers indicate the location of the vegetation relevés.
The cluster analysis of the relief matrix used the average bond criterion (UPGMA) and the Chord distance algorithm to identify homogeneous floristic compositions.
To highlight the ecological characteristics of plant species, the Pignatti–Ellenberg indicator value was used in the version modified by Pignatti [66,67,68] for the Italian territory. The values of the ecological indicator of continentality (C), substrate reaction (R), temperature (T), humidity/water availability (U), light (L) and nutrients (N) were taken into account.
The PCA was performed taking into account the Ellenberg ecological indicator attributed to each species in the survey matrix to show the distribution of values for ecological characteristics.
The nomenclature of the species follows the “Portal to the flora of Italia” [69], while syntaxonomic references of plant communities take into account several studies [70]. Manuals for the interpretation and monitoring of habitats of community interest in Italy were used for the interpretation of community habitats according to EEC Directive 43/92 [71,72].
Grassland biodiversity was assessed using the Shannon–Wiener H⁺ [73] and Evenness (J) indices.
where pi is the relative presence value of the ith species.
where Hmax = ln S and S = species richness.
H+ = −Σpi ln pi
J = H/Hmax,
This index depends on the abundance and uniform distribution of species [74]. J’ varies between 1 when the species of the community are equally distributed and 0 when only one species is present.
The Shannon index (H⁺) has also made it possible to evaluate Naturality (Na) by taking into account not only the presence of alien species [74,75,76,77] but also the indicators of disturbance, which emphasise the anthropic impact on the community.
Na = H⁺ (without alien and disturbing plant species)/H⁺).
The Na index varies from 0 to 1, where 0 indicates that plant diversity consists entirely of alien and disturbance species, while 1 indicates the absence of the latter in phytocoenosis [74,75,76,77,78]. The alien species were extrapolated from the “Portal of the flora of Italy” [69], while for the disturbance species the syntaxonomic references of the plant communities [70] belonging to the anthropic vegetation classes Stellarietea mediae Tüxen, Lohmeyer & Preising ex Von Rochow 1951 and Galio aparines-Urticetea dioicae Passarge ex Kopecký 1969 were taken. Most disturbance species characterize various alliances: Securigero securidacae-Dasypyrion villosi Cano-ortiz, Biondi & Canoex Cano-ortiz, Biondi & Canoin Biondi et al., 2015 [Bromopsis erecta (Huds.), Dasypyrum villosum (L.) P.Candargy), Fedio graciliflorae-Convolvulion cupaniani Brullo & Spampinato 1986 [Avena barbata Pott ex Link, Galactites tomentosus Moench, Sonchus oleraceus L., Geranium dissectum L.]; Fumarion wirtgenii-agrariae Brullo in Brullo & Marcenò 1985 [Calendula arvensis L. (Raf.) Nyman, Rumex bucephalophorus L., Fumaria agraria Lag]; Trifolio medii-Geranietea sanguinei Müller 1962 [Senecio scopolii Hoppe & Hornsch; Asphodelus ramosus L.]; Scleranthion annui (Kruseman & Vlieger 1939) Sissingh in Westhoff, Dijk, Passchier & Sissingh 1946 [Anthemis arvensis L., Agrostis castellana Boiss. & Reut., Muscari comosum (L.) Mill.]; Convolvulo arvensis-Agropyrion repenti Görs 1966 [Picris hieracioides L., Dactylis glomerata L., Bunium bulbocastanum L.]; Inulo viscosae-Agropyrion repentis Biondi & Allegrezza 1996 [Reichardia picroides (L.) Roth, Pallenis spinosa (L.) Cass., Daucus carota L., Verbena officinalis L.]; Cerastio arvensis-Cynosurenion cristati Blasi, Tilia, Rosati, Del Vico, Copiz, Ciaschetti, Burrascano 2012 [Anthoxanthum odoratum L., Lotus corniculatus L., Plantago lanceolata L.]; Resedo albae–chrysanthemenion coronarii Cano-Ortiz, Biondi & Cano In Biondi, Allegrezza, Casavecchia, Galdenzi, Gasparri, Pesaresi, Poldini, Sburlino, Vagge & Venanzoni 2015 [Reseda alba L.]; Echio plantaginei-Galactition tomentosae O. Bolòs & Molinier 1969 [Sherardia arvensis L., Plantago lagopus L., Lotus ornithopodioides L.]; Digitario ischaemi-Setarion viridis Sissingh in Westhoff, Dijk, Passchier & Sissingh 1946 (Digitaria sanguinalis (L.) Scop.; Hordeion leporini Br.-Bl. in Br.-Bl., Gajewski, Wraber & Walas 1936 corr. O. Bolòs 1962 [Carduus pycnocephalus L., Anacyclus clavatus (Desf.) Pers., Rostraria cristata (L.) Tzvelev, Glebionis coronaria (L.) Spach]; Hypochoeridion achyrophori Biondi & Guerra 2008 [Linum strictum L., Euphorbia exigua L.]; Taeniathero-Aegilopion geniculatae Rivas-Martínez & Izco 1977 [Trifolium stellatum L., Trifolium scabrum L.]; Holco mollis-Pteridion aquilini Passarge (1994) 2002) [Pteridium aquilinum (L.); Holcus lanatus L.]; Ridolfion segeti Negre ex Rivas-Martinez, Fernandez-Gonzalez & Loidi 1999 [Phalaris paradoxa L., Trigonella sulcata (Desf.) Coulot & Rabaute]; Caucalidion platycarpi Tüxen ex Von Rochow 1950 nom. mut. Rivas-martinez, T.E. Diaz, Fernandez-Gonzalez, Izco, Loidi, Lousã & Penas 2002 [Lysimachia arvensis (L.) U. Manns & Anderb.]; Veronico agrestis-Euphorbion peplus Sissingh ex Passaggio 1964 [Sonchus asper (L.) Hill].
To verify the significance of the differences observed between the different habitat groups and the average values of the ecological indicator, the Mann–Whitney test was used. Finally, a correlation analysis (PAST version 4.13 [79,80]) was applied between the value of Naturalness (Na) and the values of the ecological indicator of temperature (T) and humidity (U) of the identified habitat types. Person statistics were used, and, finally, the linear regression model by Durbin–Watson and Breusch–Pagan tests were applied [81,82,83].
3. Results
3.1. Cluster Analysis to Define Habitat Types
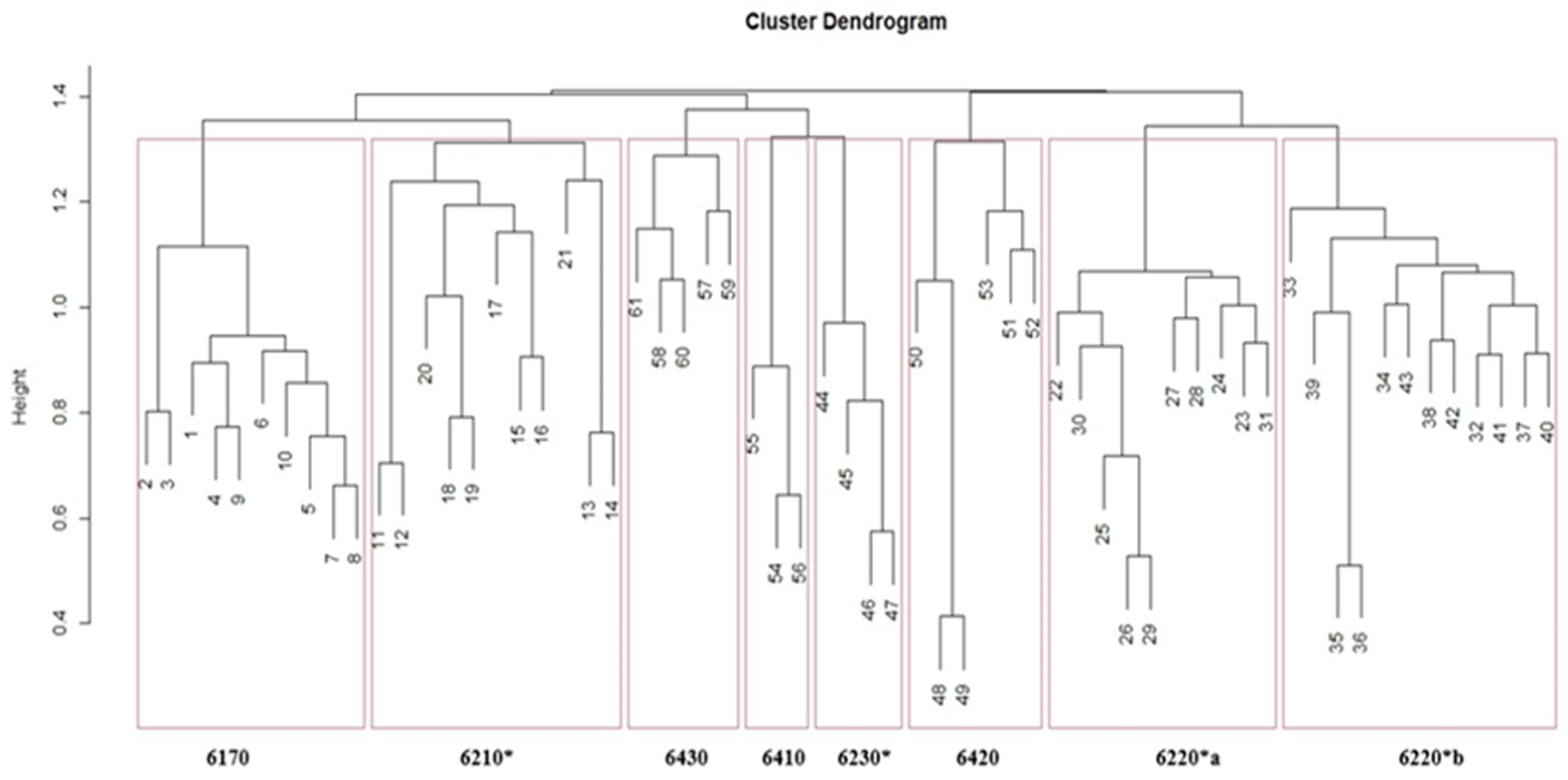
Cluster analysis of the grassland vegetation relevés matrix produced the dendrogram in Figure 2, where the abscissa shows the distinctive number of each relevés, while the ordinate shows the scale of similarity. The first subdivision is at a similarity level of 1.4; the subsequent subdivisions (similarity coefficient 1.3-1.2-1.1) show us eight groups of different grassland habitats.
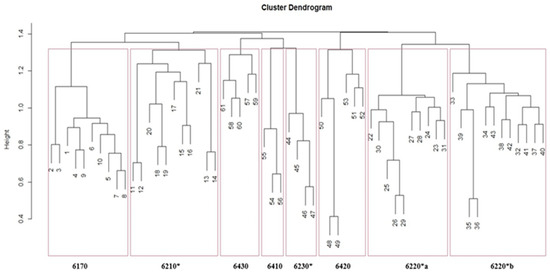
Figure 2.
Dendrogram derived from cluster analysis on grassland relevés (Chord as distance coefficient and UPGMA as clustering algorithm) showing 8 habitat groups: 6220*a: Pseudo-steppe with grasses and annuals of the Thero-Brachypodietea dominated by Lygeum spartum L., 6220*b: Pseudo-steppe with grasses and annuals of the Thero-Brachypodietea dominated a Hyparrhenia hirta (L.) Stapf, 6230*: Nardus grasslands rich in species, on siliceous substrates in mountain areas; 6210*: Semi-natural dry grasslands and scrubland facies on calcareous substrates (Festuco-Brometalia) (*important orchid sites).
The first group (Rel. 1–10), according to “Italian interpretation manual of the 92/43/EEC Habitats Directive” [71], belongs to the habitat 6170: Alpine and subalpine calcareous grasslands, the second group (Ril 11–21) belongs to habitat 6210*: Semi-natural dry grasslands and scrubland facies on calcareous substrates (Festuco-Brometalia) (*important orchid sites), the third group (Rel. 22–31) to the habitat 6220*a: Pseudo-steppe with grasses and annuals of the Thero-Brachypodietea dominated by Lygeum spartum L., the fourth group (Rel. 32–43) to habitat 6220*b: Pseudo-steppe with grasses and annuals of the Thero-Brachypodietea dominated by Hyparrhenia hirta, the fifth group (Rel. 44–47) fits to the habitat 6230*: Nardus grasslands rich in species, on siliceous substrates in mountain areas (and submountain areas, in Continental Europe), the sixth group (Rel. 48–53) belongs to habitat 6420: Mediterranean tall humid herb grasslands of the Molinio-Holoschoenion, the seventh group (Rel. 54–56) belongs to habitat 6410: Molinia meadows on calcareous, peaty or clayey-siltladen soils (Molinion caeruleae), and the eighth group (Rel. 57–61) belongs to habitat 6430: Hydrophilous tall herb fringe communities of plains and of the montane to alpine levels.
Habitat 6220* is characterized as grassland of small grasses rich in therophytes, predominantly open, thermo- and meso-Mediterranean bioclimate, xerophilous; oligotrophic soil communities are located on base-rich substrates, often calcareous. In loose soil, we find the habitat dominated by Hyparrhenia hirta [84], while on clayey soils, we find the habitat dominated by Lygeum spartum [85]; in both, it is possible to find some interesting species [86]. Habitat 6230* [87,88] includes grasslands dominated by Nardus stricta which develop on acid soils in rather flat areas from the hills to the mountain range; in acidic soils, they can derive from siliceous or carbonate rocks through leaching. The grasslands of habitat 6210* [71,89,90] are predominantly of secondary origin and characterized by a high density of species. Hemicryptophytes dominate these communities and can develop on different types of substrate. In Italy, these grasslands, excluding the North, show their greatest presence along the Apennine chain. Habitat 6170 is located on limestone substrates [91,92] and frames the grasslands present in the Alps and the Apennines above the forest limit. Habitat 6410 includes wet semi-natural grasslands characterized by Molinia caerulea [93,94] and is located from the plain to the upper limit of the forest. Habitat 6420 includes high size grasslands characterised by Mediterranean reeds and other hygrophilous species [95,96] and is present in retrodunal systems but also in indoor wetlands. Habitat 6430 is characterized by hygrophilous and nitrophilous species (megaforbes), which settle mainly along the edge of waterways [97,98].
3.2. Ecological Factors Analysis
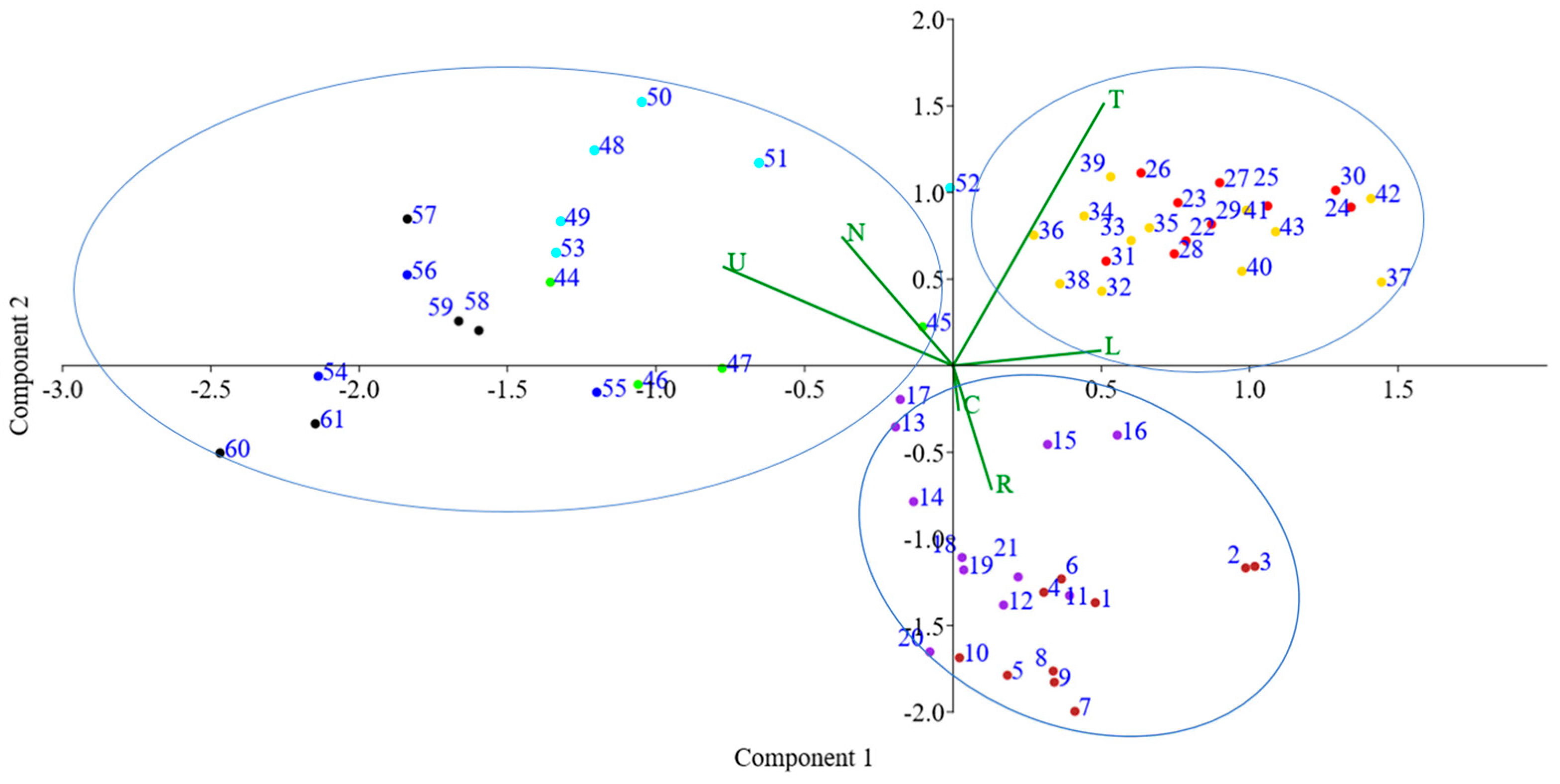
The PCA was carried out taking into account the Ellenberg ecological indicator value, which showed the distribution of relevés on the basis of ecological characteristics and explained 81.3% of total dataset variation with the first two components, with PC1: 60.9% and PC2: 20.4% (Figure 3).

Figure 3.
Biplot of relevés dispersion on PCA related to Ellenberg ecological indicator. The numbered points are the relevés of the various habitat types: in purple is the habitat 6210*, brown is the habitat 6170, light green is the habitat 6230*, red is the habitat 6220*a, yellow is the habitat 6220*b, sky blue is the habitat 6410, black is the habitat 6430, blue is the habitat 6420. The green lines indicate the direction of maximum correlation of the Ellenberg indicator values. (U = humidity; C = continentality; R = substrate reaction; T = temperature; L = light; N = nutrients).
The PCA highlights three habitat groups. The habitats 6230*, 6410, 6420, and 6430 are related to the ecological factors of moisture (U) and nutrients (N); these are in fact habitat types located in the mountainous belt, where there is a climate with greater water availability. The habitats 6220a* and 6220*b are arranged along ecological gradients of temperature (T) and light (L), confirming that these habitats are characterised by herbaceous, perennial, thermophilous vegetation that settles in the driest areas of the Mediterranean [70]. Habitats 6170 and 6210* are distributed on the basis of the ecological indicator of continentality (C) and substrate reaction (R); they are in fact habitats located in the mountainous belt on highly permeable basic pH limestone substrates.
The overall table of this assessment is Supplementary Materials Table S2.
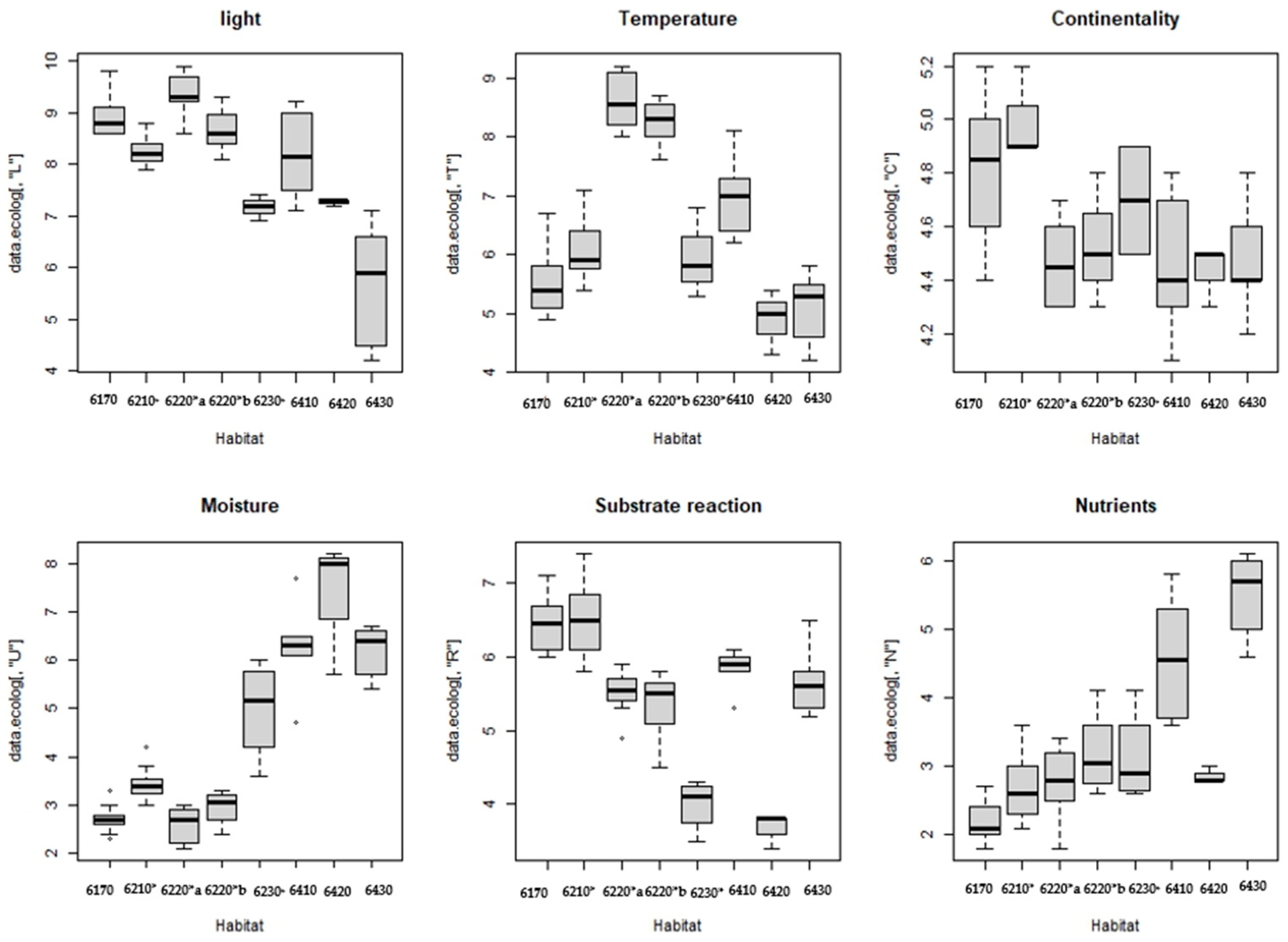
The ecological features of the eight identified habitats are shown in Figure 4 by boxplots with the values of the ecological indicator of humidity (U), temperature (T), light (L), nutrient (N), continentality (C) and substrate reaction (R). To quantify the separability of the various habitat groups considering the mean values of the ecological index, the Mann–Whitney test was applied.
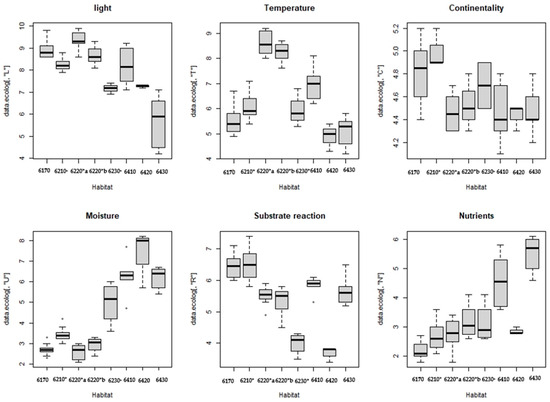
Figure 4.
Box plot representing the Ellenberg indicator values of each habitat: 6170 Alpine and subalpine calcareous grasslands; 6210* Semi-natural dry grasslands and scrubland facies on calcareous substrates (Festuco-Brometalia) (*important orchid sites); 6220*a Pseudo-steppe with grasses and annuals of the Thero-Brachypodietea dominated a Lygeum spartum L., 6220*b Pseudo-steppe with grasses and annuals of the Thero-Brachypodietea dominated by Hyparrhenia hirta; 6230* Nardus grasslands rich in species, on siliceous substrates in mountain areas; 6420 Mediterranean tall wet grass grasslands of the Molinio-Holoschoenion; 6410 Molinia meadows on calcareous, peaty clayey-siltladen soils (Molinion caeruleae); 6430 Hydrophilous tall grass fringe communities of plains and of the mountain to alpine levels.
The ecological factor light (L) is greatest regarding 6220*a and 6220b*, while lower values are observed in the habitat 6420; significant differences occur for this ecological indicator between habitat 6220*a and habitats 6170 (p-value = 0.000) and with habitat 6210* (p-value = 0.000); other significant differences are observed between habitat 6220*b and habitat 6430 (p-value = 0.04).
The ecological indicator temperature (T) is higher in habitats 6220a*and 6220*b and lower in habitats 6170, 6420 and 6430, as shown by the dissimilarity between habitat 6170 and habitats 6220*a (p-value = 0.00) and 6220*b (p-value = 0.00); other statistically significant differences are shown between habitat 6220*a and habitats 6210* (p-value = 0.00); 6420 (p-value = 0.05); and 6430 (p-value = 0.05). Finally, habitat 6220*b also shows differences with habitat 6210* (p-value = 0.00) and 6430 (p-value = 0.04).
The Ellenberg indicator values of continentality (C) are highest in habitat 6210* and 6170, while they are lower in habitats 6220*a 6220*b, 6410 and 6430. These differences are significant because habitat 6210* differs from habitats 6220*a (p value = 0.00), 6220*b (p value = 0.00), 6410 (p value = 0.01) and 6430 (p value = 0.03).
The indicator values of moisture (U) are higher in habitats 6430, 6420 and 6410, while they are lower in habitats 6220*a, 6220*b, 6170 and 6210*. Statistically significant differences are observed between habitat 6170 and 6210* (p-value = 0.0097), 6410 (p-value = 0.0314;) and 6430 (p-value = 0.0490;). Other significant differences are found between habitat 6210* and habitats 6220*a (p-value = 0.00), 6220*b (p-value = 0.03), 6410 = (p-value = 0.02) and 6430 (p-value = 0.04). Finally, the humidity factor differentiates habitat 6220*a with 6410 (p-value = 0.03), 6220*a with 6430 (p-value = 0.04), habitat 6220*b with 6410 (p-value = 0.03), and 6220*b with 6430 (p-value = 0.03).
The indicator values of substrate reaction (R) show other significant differences between habitat 6170 and habitats 6220*a (p-value = 0.00); 6220*b (p-value = 0.00). Analysis shows that habitat 6210* differs from habitats 6220*a (p-value = 0.00) and 6220*b (p-value = 0.00), confirming that the grasslands of this habitat develop on calcareous and basic or sub-acid substrates [70].
The nutrient ecological indicator (N) in wet grasslands 6430 and 6410 differs significantly from other habitats. In particular, habitat 6170 differs from habitat 6220*b (p-value = 0.00) and habitats 6410 (p-value = 0.03) and 6430 (p-value = 0.05), and also habitat 6220*a has significant differences to habitats 6410 (p-value = 0.03) and 6430 (p-value = 0.05), and habitat 6220*b differs from habitat 6430 (p-value = 0.04).
Overall, this analysis showed that the ecological indicator humidity (U) and nutrients (N) are the most important that influence the composition of the species in the analysed dataset, while the factors of light (L) and substrate reaction (R) are less important. This confirms the clear difference between dry and wet grasslands. The overall table of this assessment is a Supplementary Materials Table S3.
3.3. Plant Diversity and Naturalness
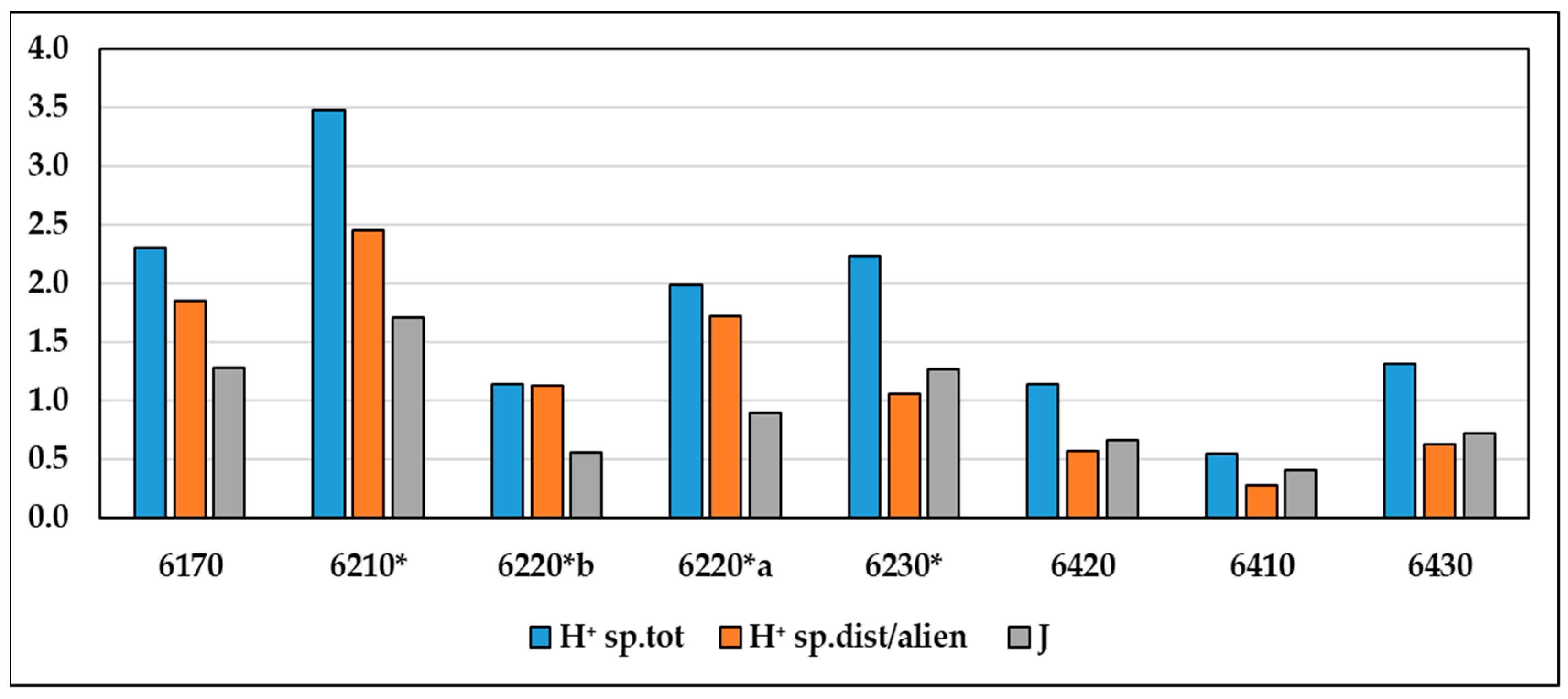
The biodiversity values of the grassland habitats compared (Figure 5) show that the total Shannon index (H⁺) is higher in grassland habitats characterized by species with higher humidity values (U), including 6230*, 6430, 6420 and 6210*, while the Shannon index (H⁺) calculated on disturbance and alien species has higher values for the driest grassland habitats, such as 6220*a and 6220*b, characterized by species with higher Ellemberg indicator value of temperature (T) and light (L). Equipartition (J) is higher in habitats with higher humidity index (U) (see Supplementary Materials Table S4).

Figure 5.
Values of biodiversity indices for each habitat type (H+ = Shannon total species, J = Evenness, H+ sp. dist/alien = Shannon alien and disturbance species).
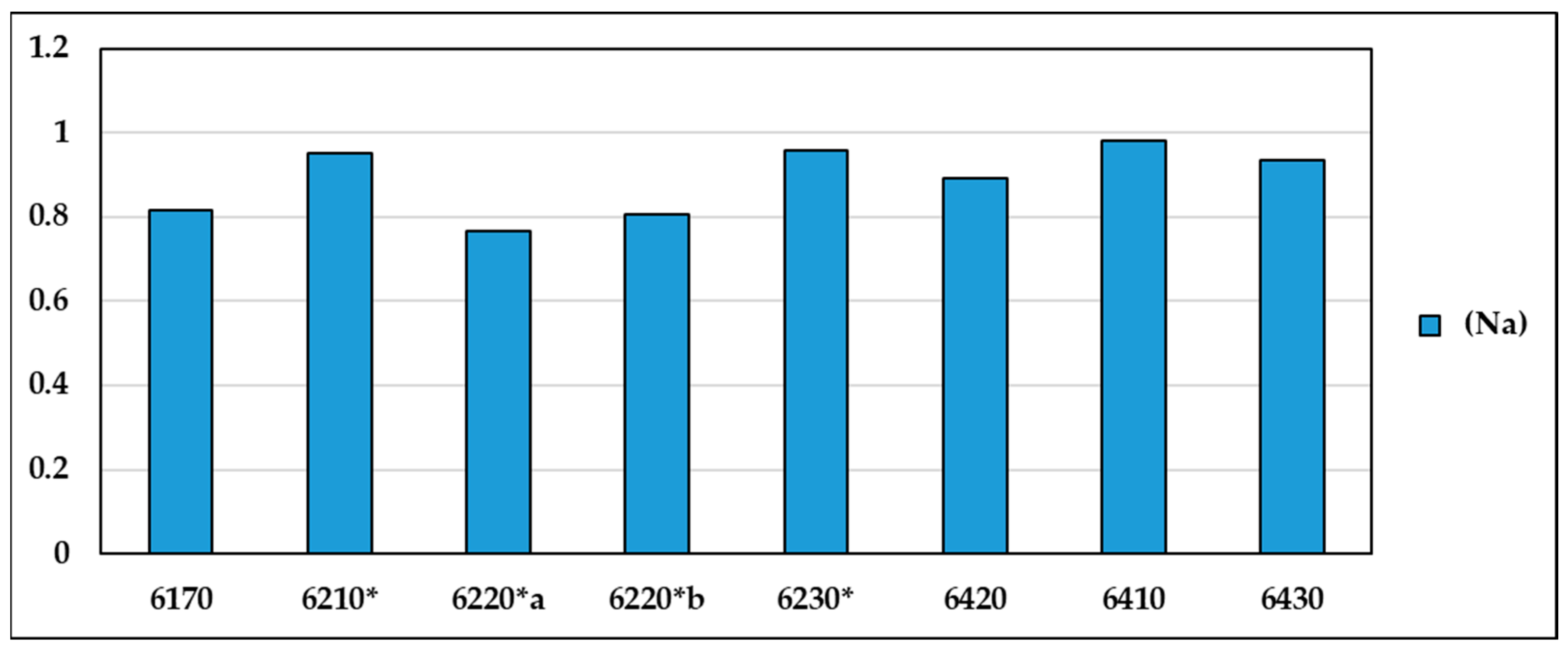
The naturalness values (Na) for the different habitat types (Figure 6), which take into account the ratio between the values of total H⁺ and H⁺ without disturbance and alien species, are found to be at a maximum level in habitats 6230*, 6410, 6430 and 6420, while habitats 6220*b, 6220* and 6170 show lower values of naturalness (Supplementary Materials Table S5).
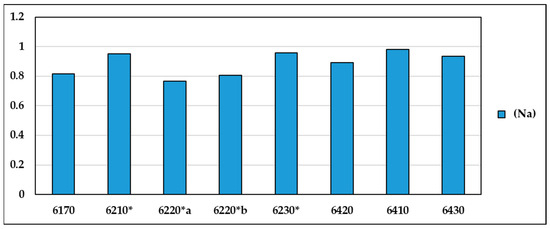
Figure 6.
Values of the naturalness index (Na) for the various habitat types.
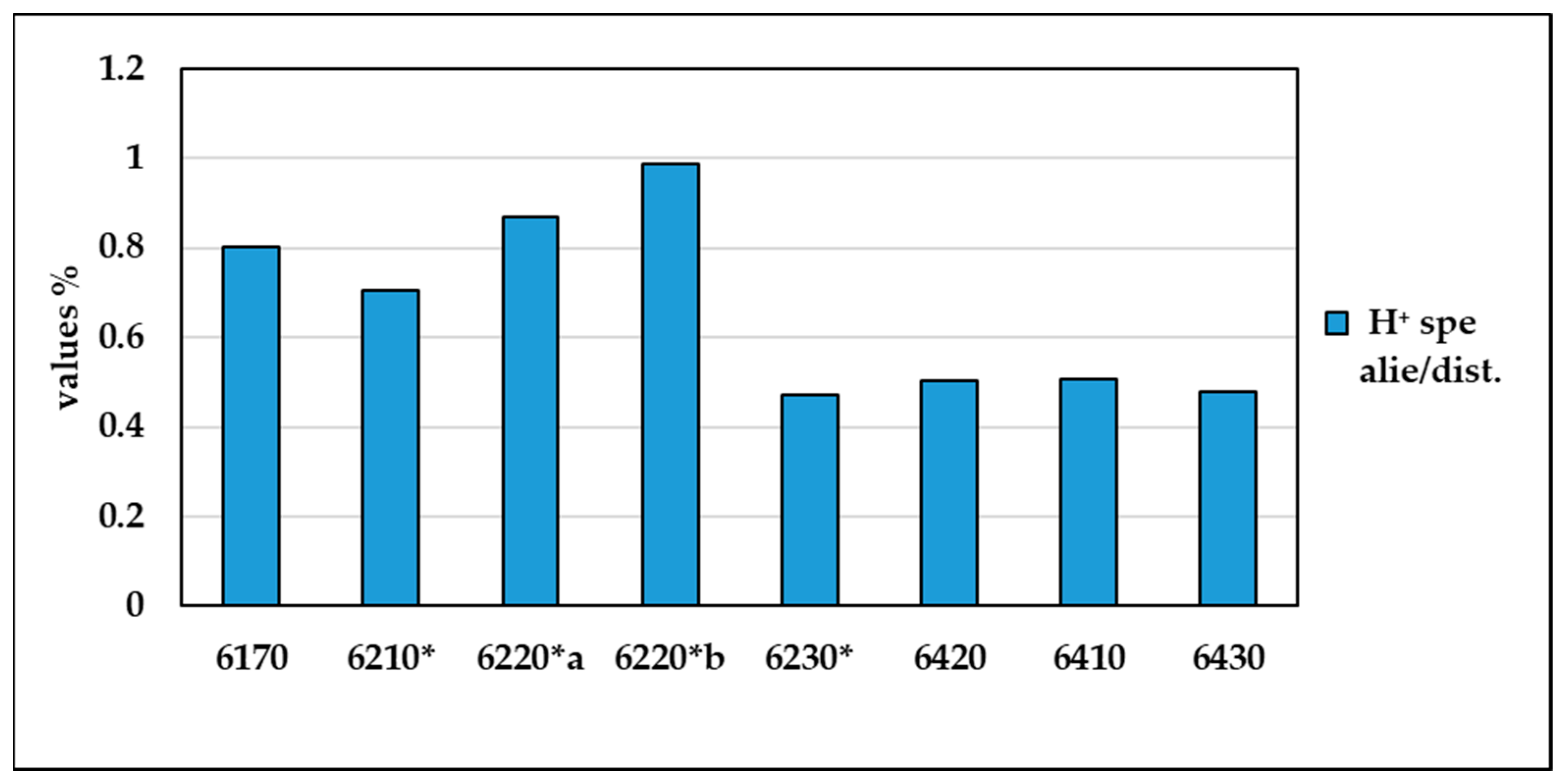
Figure 7 shows the contribution to the biodiversity of alien and disturbance species in the different habitat types, taking into account the relationship between the total H⁺ value and the H⁺ value of the alien and disturbance species. The graph shows that maximum values are found in habitats 6220*b, 6220* and 6170, confirming that in these habitats disturbance and alien species contribute more to the value of biodiversity than habitats 6230*, 6410, 6430 and 6420, which show lower values and are therefore characterised by greater biodiversity (Supplementary Materials Table S6).
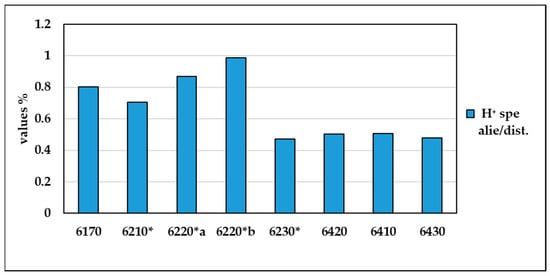
Figure 7.
Percentage values of the H⁺ of alien and disturbance species on total biodiversity in the various types of habitat.
3.4. Relationship between Diversity and Ellenberg Indicator Values
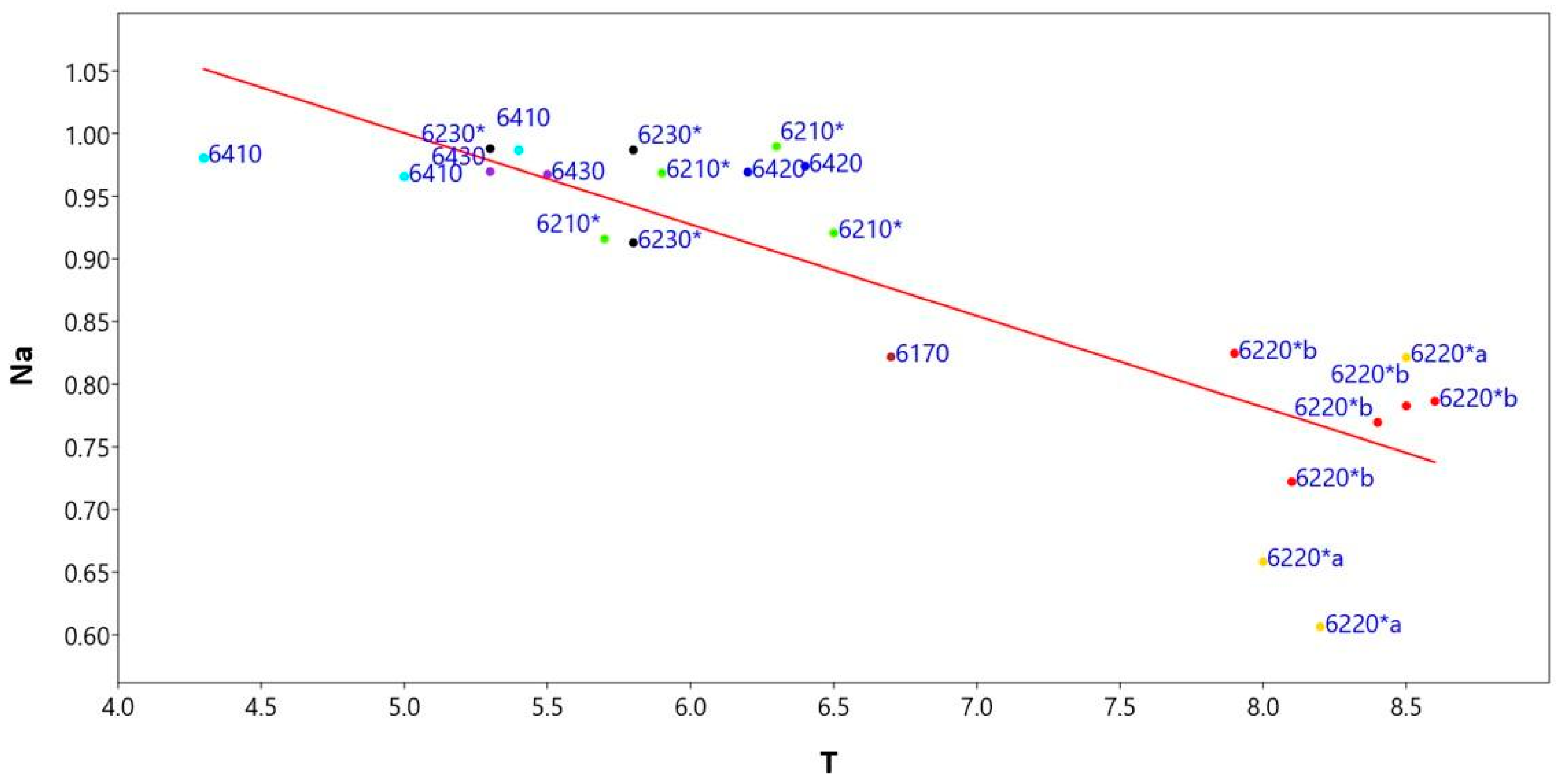
The linear regression model (Figure 8) between the naturalness index (Na) and the ecological temperature indicator (T) shows a significant slope value < 0.05 (0.000046817), correlation index r = −0.84 and regression index r2 = 0.71 (Supplementary Materials Table S7).
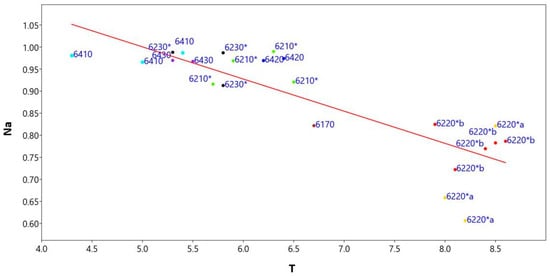
Figure 8.
Linear regression model between the naturalness index (Na) and the Ellenberg indicator values of temperature (T) of the representative relevès for each habitat type: in yellow 6220*a, in red 6220*b, in black 6230*, in green 6210*, in brown 6170, in dark blue 6420, in blue 6410, in violet 6230*.
To check the autocorrelation analyses, the Durbin-Walson statistical test was applied, which assumes a value of 1.82 and a probability of autocorrelation of 32%, while the variance of the residuals and thus the homeschedasticity was calculated using the Breusch–Pagan test, with a value of 3.01 and a probability of homeschedasticity of 0.08%. This analysis shows that habitats’ naturalness decreases with increasing temperature, as low-lying habitats are most impacted by anthropogenic activity and are often close to urbanized areas.
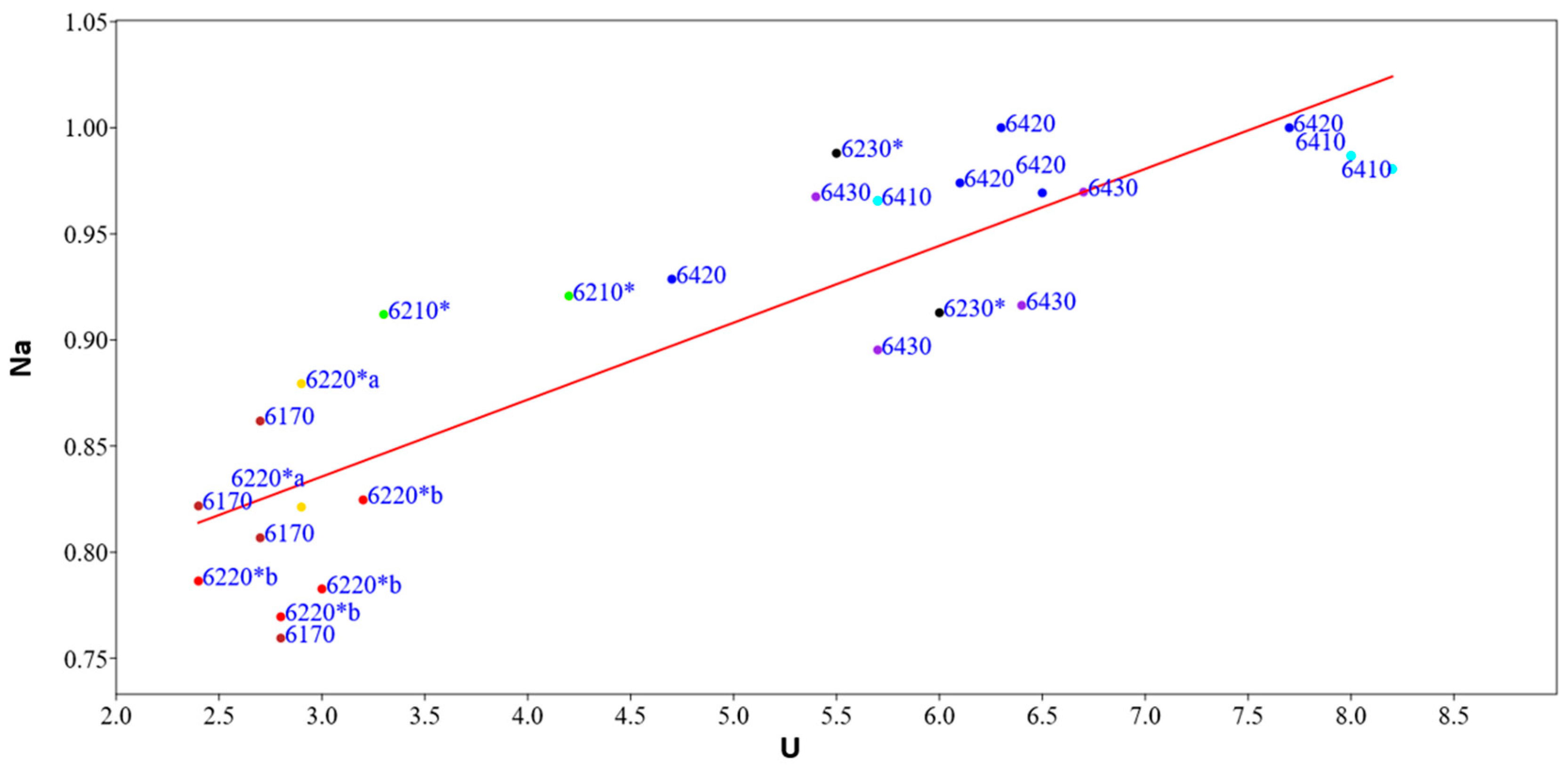
The linear regression model between the naturalness index (Na) and the Ellenberg indicator values of moisture (U) (Figure 9) shows a significant slope value < 0.05 (0.000000010994), correlation index r = −0.87 and regression index r2 = 0.75 (Supplementary Materials Table S8). In order to verify the autocorrelation analyses, the Durbin–Watson statistical test was applied, which assumes a value of 1.51 and a probability of autocorrelation of 0.09%, while the variance of the residuals and thus the homeschedasticity was calculated using the Breusch–Pagan test, with a value of 1.40 and a probability of homeschedasticity of 23%. This analysis agrees with the previous one; in fact, it shows that the naturalness of habitats increases with increasing humidity, an ecological indicator that characterizes habitats at higher altitudes with low values of the temperature index, located in the mountain belt where most of the protected areas of the region are located and where the anthropic pressure is lower.
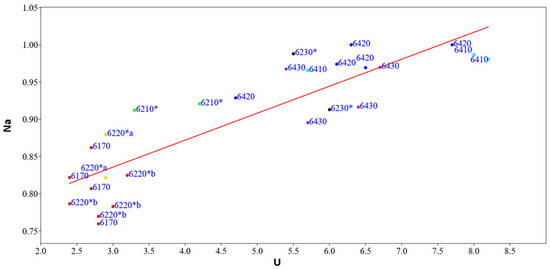
Figure 9.
Linear regression model between the naturalness index (Na) and the Ellenberg indicator values of humidity (U) of the representative relevés for each habitat type: in yellow 6220*a, in red 6220*b, in black 6230*, in green 6210*, in brown 6170, in dark blue 6420, in blue 6410, in violet 6230*.
4. Discussion
Cluster analysis on grasslands vegetation allowed us to identify eight different groups attributed to as many habitats of EEC Directive 43/92 in accordance with the interpretation manual [71]. These groups are in agreement with the alliances recognized in the phytosociological literature [70,99].
Principal component analysis (PCA) allowed us to recognize three groups in the matrix of phytosociological relevés based on ecological factors: habitats 6210* and 6170 are characterized by species that prefer base-rich substrates, also showing high values of continentality; habitats 6410, 6430 and 6420 are those with the greatest availability of moisture and nutrients in the soil; and habitats 6220*a and 6220*b are the most thermophilous ones as they are located at low altitudes in purely Mediterranean bioclimatic conditions [86].
Box plots of Ellengerg indicator values confirm ecological differences between habitats. In fact, the box plots show a significant difference for the continentality indicator between habitat 6210* and the other habitats, which is related to the fact that the typical vegetation of this habitat, referring to the class Festuco valesiacae-Brometea erecti Br.-Bl. & Tüxen ex Br.-Bl. 1949, characterizes the continental grasslands of Central and Eastern Europe, which develop in areas with low annual rainfall, in agreement with Biondi et al. [70]. This habitat, in the analysis carried out, is similar to 6170, characterized by the vegetation of the Ranunculo pollinensis-Nardion strictae Bonin 1972 alliance distributed in the mountain range on calcareous or dolomitic substrates and compacted soils rich in organic substances, often in stations characterized by prolonged snowfalls which guarantee greater soil humidity [100].
The high biodiversity values of habitats 6230*, 6430, 6420 and 6210* are associated with the high value of the Ellember indicator moisture value (U) and this is consistent with [101], which emphasises high biodiversity values for semi-natural wet grassland habitats.
The ecological factors that contribute to the differentiation of habitat types are mainly temperature (T) and humidity (U), which play an important role in defining the floristic composition of the identified grassland habitats. Indeed, grassland habitats are widespread in the region from sea level to the mountain belt and occupy a wide altitudinal range, where temperature and humidity factors play a fundamental role in the ecological characterisation of the different habitat types.
The nutrient factor (N) showed significant differences between the findings belonging to habitat type 6430, characterized by nitrophilous communities, and those belonging to groups 6220a and 6220b; this could be due, in agreement with the evidence of other studies [102,103], to a higher soil moisture influencing nutrient availability. In fact, habitat 6430 presents higher humidity values, but, above all, high values of naturalness and diversity are characterized by a variable phytocoenosis consisting also of species from other communities [104], some of which are rare and threatened, which, in agreement with other studies, guarantees a greater conservation value for the habitat [105].
Overall, the analysis performed, in agreement with some studies [106], shows that the most important ecological gradients influencing the diversity of grassland vegetation are moisture and nutrients. Furthermore, the results show that grassland habitats characterised by species with higher moisture values are more natural.
This is evident in habitat 6410, of grasslands with high floristic diversity and a very broad ecology developing on mesotrophic, oligotrophic, moist and cool soils [70,107]. The vegetation of this habitat is linked to the water table, which determines the differentiation of species in these environments [101].
A Mediterranean, thermo-xerophilous vegetation characterizes the 6220*a and 6220*b habitat types; they grow at low altitudes in the driest stations where ecological factors such as temperature and brightness are greater and therefore reduce water availability within the soil [86]. These habitats are the least natural, as intensive land use at low altitudes, associated with fires, soil disturbance and anthropisation, plays a fundamental role in structuring grassland plant communities [86,108], affecting diversity and naturalness and causing changes in biodiversity, both in terms of a reduction in species richness and the replacement of typical species by disturbed ones [109,110].
Many of the species typical of grasslands are threatened with extinction [110], and the decrease in surface area and quality shows that grasslands are now among the most endangered European habitats [18,111].
5. Conclusions
Our study shows that the plant assemblage composition is the best tool to characterize grassland habitats from an ecological and typological point of view, indeed the analysis of grassland vegetation can provide information on ecological factors that influence the species composition and distribution of plant communities. In particular, the species diversity indices (Shannon, Evenness) applied in this study show that naturalness is greater in wet than dry grasslands. However, knowledge of land use and the environmental context of ecosystems varies due to the interaction between the effects of human activity and other environmental factors. Ecological indicators, such as those used here, can therefore be equally effective to define patterns and processes of change and are a useful tool for identifying areas of high natural value. To counter the continuing loss of grasslands and their typical species, it is important to understand the causes of vegetation change, which are likely to differ between different habitat groups. Therefore, the results of this study showed that the diversity and naturalness indices applied, together with the assessment of ecological factors, are useful tools for implementing both an appropriate interpretation and a management and conservation strategy under EEC Directive 43/92. Understanding the ecological features and assessing habitat biodiversity allows us to interpret evolutionary dynamics and predict future scenarios due to climate change [112,113].
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/land13060719/s1, Table S1: matrix of phytosociological relevés; Table S2: Relevés’ dispersion biplot on PCA in relation to ecological factor; Table S3: The mean values of the ecological index and the non-parametric Kruskal–Wallis test; Table S4: Values of biodiversity indices for each habitat type (H⁺ = Shannon total species, J = Evenness, H⁺ sp. dist/alien = Shannon index of alien and disturbance species); Table S5: Values of the naturalness index (Na) of habitat types; Table S6: Percentage values of the H⁺ index of alien and disturbance species on total biodiversity in the various types of habitat; Table S7: Linear regression model between the naturalness index (Na) and the values of the ecological temperature factor (T); Table S8: Linear regression model between the naturalness index (Na) and the values of the ecological factors of humidity (U) of the representative surveys for each type of habitat.
Author Contributions
Conceptualization, A.M., C.M.M. and G.S.; methodology, A.M.; software, A.M.; analysis, A.M.; data curation, C.M.M.; writing—original draft preparation, A.M. and C.M.M.; writing—review and editing, A.M., C.M.M. and G.S.; visualization, G.S.; supervision, G.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was partially supported by “Nature Map System” project signed between the AGRARIA Department—Mediterranean University of Reggio Calabria and the Territory and Environment Department of Calabria Region, Sector 5, Parks and Protected Natural Areas, under the Regional Operational Program (ROP) 2014/2020—Action 6.5.A.1—Actions provided for in the Prioritez Action Framework (PAF) and in the Natura 2000 Network Management Plans, scientific manager GiovanniSpampinato.
Data Availability Statement
The original contributions presented in the study are included in the article.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Bohn, U.; Gollub, G.; Hettwer, C.; Neuhäuslová, Z.; Raus, T.; Schlüter, H.; Weber, H.; Hennekens, S. (Eds.) Map of the Natural Vegetation of Europe. Scale 1:2,500,000. Interactive CD-ROM: Explanatory Text, Legend, Maps; Bundesamt für Naturschutz: Bonn, Germany, 2004. [Google Scholar]
- Musarella, C.M.; Brullo, S.; del Galdo, G.G. Contribution to the Orophilous Cushion-Like Vegetation of Central-Southern and Insular Greece. Plants 2020, 9, 1678. [Google Scholar] [CrossRef] [PubMed]
- Cano Ortiz, A.; Quinto Canas, R.; Pinar Fuentes, J.C.; del Rio, S.; Pinto Gomes, C.J.; Cano, E. Endemic Hemicryptophyte Grasslands of the High Mountains of the Caribbean. Res. J. Ecol. Environ. Sci. 2022, 2, 1–20. [Google Scholar] [CrossRef]
- Mücher, C.A.; Hennekens, S.M.; Bunce, R.G.; Schaminée, J.H.; Schaepman, M.E. Modelling the spatial distribution of Natura 2000 habitats across Europe. Landsc. Urban Plan. 2009, 92, 148–159. [Google Scholar] [CrossRef]
- APAT. The Implementation in Italy of the European Corine Land Cover 2000 Project; Reports 36/2005; APAT: Rome, Italy, 2005; p. 86. [Google Scholar]
- Oppermann, R.; Beaufoy, G.; Jones, G. (Eds.) High Nature Value Farming in Europe: 35 European Countries—Experiences and Perspectives; Verlag Regionalkultur: Ubstadt-Weiher, Germany, 2012. [Google Scholar]
- Van Elsen, T. Species diversity as a task for organic agriculture in Europe Agric. Ecosyst. Environ. 2000, 77, 101–109. [Google Scholar] [CrossRef]
- Cano, E.; Cano-Ortiz, A.; Quinto Canas, R.; Piñar Fuentes, J.C.; Rodrigues Meireles, C.; Raposo, M.; Pinto Gomes, C.; Laface, V.L.A.; Spampinato, G.; Musarella, C.M. Ecological and Syntaxonomic Analysis of the Communities of Glebionis coronaria and G. discolor (Malvion neglectae) in the European Mediterranean Area. Plants 2024, 13, 568. [Google Scholar] [CrossRef] [PubMed]
- Söderström, B.; Svensson, B.; Vessby, K.; Glimskär, A. Plants, insects and birds in semi-natural pastures in relation to local habitat and landscape factors. Biodivers. Conserv. 2001, 10, 1839–1863. [Google Scholar] [CrossRef]
- Pärtel, M.; Bruun, H.H.; Sammul, M. Biodiversity in temperate European grasslands: Origin and conservation. Grassl. Sci. Eur. 2005, 10, 1–14. [Google Scholar]
- Gibson, D.J. Grasses and Grassland Ecology; Oxford University Press: Oxford, UK, 2009. [Google Scholar]
- Wilson, J.; Peet, R.K.; Dengler, J.; Pärtel, M. Plant species richness: The world records. J. Veg. Sci. 2012, 23, 796–802. [Google Scholar] [CrossRef]
- Bengtsson, J.; Bullock, J.M.; Egoh, B.; Everson, C.; Everson, T.; O’Connor, T.; O’Farrell, P.J.; Smith, H.G.; Lindborg, R. Grasslands—More important for ecosystem services than you might think. Ecosphere 2019, 10, e02582. [Google Scholar] [CrossRef]
- Schrautzer, J.; Jansen, D.; Breuer, M.; Nelle, O. Succession and management of calcareous dry grasslands in the Northern Franconian Jura, Germany. Tuexenia 2009, 29, 339–351. [Google Scholar]
- Cano Carmona, E.; Quinto Canas, R.; Cano Ortiz, A.; Musarella, C.M. Introductory Chapter: Methodological Aspects for the Study of Vegetation. In Vegetation Index and Dynamics; IntechOpen: London, UK, 2022. [Google Scholar] [CrossRef]
- Dengler, J.; Janišová, M.; Török, P.; Wellstein, C. Biodiversity of Palaearctic grasslands: A synthesis. Agric. Ecosyst. Environ. 2014, 182, 1–14. [Google Scholar] [CrossRef]
- Valkó, O.; Zmihorski, M.; Biurrun, I.; Loos, J.; Labadessa, R.; Venn, S. Ecology and conservation of steppes and semi-natural grasslands. Hacquetia 2016, 15, 5. [Google Scholar] [CrossRef]
- Veen, P.; Jefferson, R.; De Smidt, J.; Van der Straaten, J. (Eds.) Grasslands in Europe of High Nature Value; KNNV Publishing: Zeist, The Netherlands, 2009. [Google Scholar]
- Reitalu, T.; Johansson, L.J.; Sykes, M.T.; Hall, K.; Prentice, H.C. History matters: Village distances, grazing and grassland species diversity. J. Appl. Ecol. 2010, 47, 1216–1224. [Google Scholar] [CrossRef]
- Reitalu, T.; Helm, A.; Pärtel, M.; Bengtsson, K.; Gerhold, P.; Rosén, E.; Takkis, K.; Znamenskiy, S.; Prentice, H.C. Determinants of fine-scale plant diversity in dry calcareous grasslands within the Baltic Sea Agricultural region. Agric. Ecosyst. Environ. 2014, 182, 59–68. [Google Scholar] [CrossRef]
- Turtureanu, P.D.; Palpurina, S.; Becker, T.; Dolnik, C.; Ruprecht, E.; Sutcliffee, L.M.E.; Szabó, A.; Dengler, J. Scale- and taxon-dependent biodiversity patterns of dry grassland vegetation in Transylvania. Agric. Ecosyst. Environ. 2014, 182, 15–24. [Google Scholar] [CrossRef]
- Mendes, P.; Meireles, C.; Vila-Viçosa, C.; Musarella, C.; Pinto-Gomes, C. Best management practices to face degraded territories occupied by Cistus ladanifer shrublands—Portugal case study. Plant Biosyst. 2015, 149, 494–502. [Google Scholar] [CrossRef]
- Roleček, J.; Čornej, I.I.; Tokarjuk, A.I. Understanding the extreme species richness of semi-dry grasslands in east-central Europe: A comparative approach. Preslia 2014, in press. [Google Scholar]
- Musarella, C.M.; Sciandrello, S.; Domina, G. Competition between alien and native species in xerothermic Steno-Mediterranean grasslands: Cenchrus setaceus and Hyparrhenia hirta in Sicily and southern Italy. Vegetos 2024, 1–8. [Google Scholar] [CrossRef]
- Hansen, A.J.; De Fries, R.S.; Turner, W. Land use change and biodiversity. In Land Change Science, Remote Sensing and Digital Image Processing; Gutman, G., Janetos, A.C., Justice, C.O., Moran, E.F., Mustard, J.F., Rindfuss, R.R., Skole, D., Turner, B.L., Cochrane, M.A., Eds.; Springer: Dordrecht, The Netherlands, 2004; pp. 277–299. [Google Scholar]
- Spampinato, G.; Crisarà, R.; Cameriere, P.; Cano-Ortiz, A.; Musarella, C.M. Analysis of the Forest Landscape and Its Transformations through Phytotoponyms: A Case Study in Calabria (Southern Italy). Land 2022, 11, 518. [Google Scholar] [CrossRef]
- Spampinato, G.; Crisarà, R.; Cannavò, S.; Musarella, C.M. Phytotoponims of southern Calabria: A tool for the analysis of the landscape and its transformations. Atti Soc. Toscana Sci. Nat. Mem. Ser. B 2017, 124, 61–72. [Google Scholar] [CrossRef]
- Kuemmerle, T.; Levers, C.; Erb, K.; Estel, S.; Jepsen, M.R.; Müller, D.; Plutzar, C.; Stürck, J.; Verkerk, P.J.; Verburg, P.H.; et al. Hotspots of land use change in Europe. Environ. Res. Lett. 2016, 11, 064020. [Google Scholar] [CrossRef]
- Watson, J.E.M.; Jones, K.R.; Fuller, R.A.; Di Marco, M.; Segan, D.B.; Butchart, S.H.M.; Allan, J.R.; McDonald-Madden, E.; Venter, O. Persistent disparities between recent rates of habitat conversion and protection and implications for future global conservation targets. Conserv. Lett. 2016, 9, 413–421. [Google Scholar] [CrossRef]
- Whittaker, R.H. Evolution and measurement of species diversity. Taxon 1972, 21, 213–251. [Google Scholar] [CrossRef]
- Margules, C.; Pressey, R. Systematic conservation planning. Nature 2000, 405, 243–253. [Google Scholar] [CrossRef] [PubMed]
- Whittaker, R.J.; Araújo, M.B.; Jepson, P.; Ladle, R.J.; Watson, J.E.M.; Willis, K.J. Conservation Biogeography: Assessment and prospect. Divers. Distrib. 2015, 11, 3–23. [Google Scholar] [CrossRef]
- Tilman, D.; Downing, J.A. Biodiversity and stability in grasslands. Nature 1994, 367, 363–365. [Google Scholar] [CrossRef]
- Hooper, D.U.; Chapin, F.S., III; Ewel, J.J.; Hector, A.; Inchausti, P.; Lavorel, S.; Lawton, J.H.; Lodge, D.M.; Loreau, M.; Naeem, S.; et al. Effects of biodiversity on ecosystem functioning: A consensus of current knowledge. Ecol. Monogr. 2005, 75, 3–35. [Google Scholar] [CrossRef]
- Auestad, I.; Rydgren, K.; Økland, R.H. Scale-dependence of vegetation–environment relationships in semi-natural grasslands. J. Veg. Sci. 2008, 19, 139–148. [Google Scholar] [CrossRef]
- De Knegt, H.J.; Van Langevelde, F.; Coughenour, M.B.; Skidmore, A.K.; De Boer, W.F.; Heitkonig, I.M.A.; Knox, N.M.; Slotow, R.; Van der Waal, C.; Prins, H.H.T. Spatial autocorrelation and the scaling of species–environment relationships. Ecology 2010, 91, 2455–2465. [Google Scholar] [CrossRef]
- Maccherini, S.; Marignani, M.; Gloria, M.; Renzi, M.; Rocchini, D.; Santi, E.; Torri, D.; Tundo, J.; Honnay, O. Determinants of plant community composition of remnant biancana badlands: A hierarchical approach to quantify species–environment relationships. Appl. Veg. Sci. 2011, 14, 378–387. [Google Scholar] [CrossRef]
- Boch, S.; Prati, D.; Schöning, I.; Fischer, M. Lichen species richness is highest in non-intensively used grasslands promoting suitable microhabitats and low vascular plant competition. Biodivers. Conserv. 2016, 25, 225–238. [Google Scholar] [CrossRef]
- Chytrý, M.; Danihelka, J.; Ermakov, N.; Hájek, M.; Hájková, P.; Kočí, M.; Kubešová, S.; Lustyk, P.; Otýpková, Z.; Popov, D.; et al. Plant species richness in continental southern Siberia: Effects of pH and climate in the context of the species pool hypothesis. Glob. Ecol. Biogeogr. 2007, 16, 668–678. [Google Scholar] [CrossRef]
- Palpurina, S.; Chytrý, M.; Tzonev, R.; Danihelka, J.; Axmanová, I.; Merunková, K.; Duchoň, M.; Karakiev, T. Patterns of fines-scale plant species richness in dry grasslands across the eastern Balkan Peninsula. Acta Oecol. 2015, 63, 36–46. [Google Scholar] [CrossRef]
- Garcia, A. Conserving the species- rich meadows of Europe. Agric. Ecosyst. Environ. 1992, 40, 219–232. [Google Scholar] [CrossRef]
- Bratli, H.; Myhre, A. Vegetation–environment relationship ship of old hay meadows in Sverveli, Telemark, S Norway. Nord. J. Bot. 1999, 19, 455–471. [Google Scholar] [CrossRef]
- Zelnik, I.; Čarni, A. Plant species diversity and composition of wet grasslands in relation to environmental factors. Biodivers. Conserv. 2013, 22, 2179–2192. [Google Scholar] [CrossRef]
- Hölzel, N.; Otte, A. Inter-annual variation in the soil seed bank of flood-meadows over two years with different flooding patterns. Plant Ecol. 2004, 174, 279–291. [Google Scholar] [CrossRef]
- Grootjans, A.P.; Hunneman, H.; Verkiel, H.; Van Andel, J. Long-term effect of drainage on species richness of a phen meadow at different spatial scales. Basic Appl. Ecol. 2005, 6, 185–193. [Google Scholar] [CrossRef]
- Zelnik, I. Vegetation of the Meadows from the Order Molinietalia W. Koch 1926 and Contact Sites in Slovenia. Ph.D. Dissertation, University of Ljubljana, Ljubljana, Slovenia, 2005. [Google Scholar]
- Havlová, M. Syntaxonomical revision of the Molinion meadows in the Czech Republic. Preslia 2006, 78, 87–101. [Google Scholar]
- Janišová, M.; Hájková, P.; Hrivnák, R.; Kliment, J.; Michálková, D.; ζezníčková, M.; Tichy, L.; Škodová, I.; Uhliarová, E.; Ujhazy, K.; et al. Grassland Vegetation of Slovak Republic: Electronic Expert System for Identification of Syntax; Botanický ústav SAV: Bratislava, Slovakia, 2007. (In Slovakian) [Google Scholar]
- Hájek, M.; Hájková, P.; Sopotlieva, D.; Apostolova, I.; Velev, N. The Balkan wet grassland vegetation: A prerequisite to better understanding of European habitat diversity. Plant Ecol. 2008, 195, 197–213. [Google Scholar] [CrossRef]
- Stančić, Z. Classification of mesic and wet grasslands in northwest Croatia. Biology 2008, 63, 1089–1103. [Google Scholar] [CrossRef]
- Zelnik, I.; Čarni, A. Wet meadows of the Molinion Koch 1926 alliance and their environmental gradients in Slovenia. Biology 2008, 63, 187–196. [Google Scholar]
- Spampinato, G.; Sciandrello, S.; del Galdo, G.G.; Puglisi, M.; Tomaselli, V.; Cannavò, S.; Musarella, C.M. Contribution to the knowledge of Mediterranean wetland biodiversity: Plant communities of the Aquila Lake (Calabria, Southern Italy). Plant Sociol. 2019, 56, 53–68. [Google Scholar]
- Perrino, E.V.; Musarella, C.M.; Magazzini, P. Management of Grazing Italian River Buffalo to Preserve Habitats Defined by Directive 92/43/EEC in a Protected Wetland Area on the Mediterranean Coast: Palude Frattarolo, Apulia, Italy. Euro-Mediterr. J. Environ. Integr. 2021, 6, 32. [Google Scholar] [CrossRef]
- Borer, E.T.; Seabloom, E.W.; Gruner, D.S.; Harpole, W.S.; Hillebrand, H.; Lind, E.M.; Yang, L.H. Herbivores and nutrients control grassland plant diversity via light limitation. Nature 2014, 508, 517–520. [Google Scholar] [CrossRef] [PubMed]
- Klaus, V.H.; Hölzel, N.; Boch, S.; Müller, J.; Socher, S.A.; Prati, D.; Kleinebecker, T. Direct and indirect associations between plant species richness and productivity in grasslands: Regional differences preclude simple generalization of productivity-biodiversity relationships. Preslia 2013, 85, 97–112. [Google Scholar]
- Roman, A.; Ursu, T.-M.; Onțel, I.; Marușca, T.; Grigore Pop, O.; Milanovici, S.; Pál Frink, J. Deviation from Grazing Optimum in the Grassland Habitats of Romania Within and Outside the Natura 2000 Network; IntechOpen: London, UK, 2019. [Google Scholar] [CrossRef]
- Li, X.L.; Png, G.K.; Li, Y.H.; Jimoh, S.O.; Ding, Y.; Li, P.; Sun, S.X. Leaf plasticity contributes to plant anti-herbivore defenses and indicates selective foraging: Implications for sustainable grazing. Ecol. Indic. 2021, 122, 107273. [Google Scholar] [CrossRef]
- Zhang, R.Y.; Wang, J.S.; Niu, S.L. Toward a sustainable grazing management based on biodiversity and ecosystem multifunctionality in drylands. Curr. Opin. Environ. Sustain. 2021, 48, 36–43. [Google Scholar] [CrossRef]
- Lozano, V.; Vacca, G.; Camarda, I.; Brundu, G.; Sanna, F.; Caredda, S.; Re, G.A. Plant Diversity in Sardinian Mountain Rangelands: Analysis of Its Relationships with Grazing, Land Management, and Pastoral Value. Ecologies 2021, 2, 164–174. [Google Scholar] [CrossRef]
- Grime, J.P.; Pierce, S. The Evolutionary Strategies that Shape Ecosystems; John Wiley & Sons: Hoboken, NJ, USA, 2012. [Google Scholar]
- Zanzottera, M.; Dalle Fratte, M.; Caccianiga, M.; Pierce, S.; Cerabolini, B.E.L. Community-level variation in plant functional traits and ecological strategies shapes habitat structure along succession gradients in alpine environment. Community Ecol. 2020, 21, 55–65. [Google Scholar] [CrossRef]
- Erdős, L.; Bede-Fazekas, Á.; Bátori, Z.; Berg, C.; Kröel-Dulay, G.; Magnes, M.; Sengl, P.; Tölgyesi, C.; Török, P.; Zinnen, J. Species-based indicators to assess habitat degradation: Comparing the conceptual, methodological, and ecological relationships between hemeroby and naturalness values. Ecol. Indic. 2022, 136, 108707. [Google Scholar] [CrossRef]
- Spampinato, G.; Angelini, P.; Bernardo, L.; Caridi, D.; Caruso, P.; Gargano, D.; Ligato, E.; Lumia, G.; Mei, G.; Manti, L.; et al. Carta della Natura della Regione Calabria; ISPRA: Rome, Italy, 2023. [Google Scholar]
- Rivas-Martinez, S. Notions on dynamic catenal phytosociology as a basis of landscape science. Plant Biosyst. 2005, 139, 135–144. [Google Scholar] [CrossRef]
- Blasi, C.; Biondi, E.; Izco, J. 100 years of plant sociology: A celebration. Plant Biosyst. 2011, 145, 1–3. [Google Scholar] [CrossRef]
- Pignatti, S.; Guarino, R.; La Rosa, M. Flora d’Italia 1, 2nd ed.; Edagricole di New Business Media: Milano, Italy, 2017; p. 1064. [Google Scholar]
- Pignatti, S.; Guarino, R.; La Rosa, M. Flora d’Italia 2, 2nd ed.; Edagricole di New Business Media: Milano, Italy, 2017; p. 1178. [Google Scholar]
- Pignatti, S.; Guarino, R.; La Rosa, M. Flora d’Italia 3, 2nd ed.; Edagricole di New Business Media: Milano, Italy, 2018; p. 1286. [Google Scholar]
- Portal to the Flora of Italy. Portale della Flora d’Italia. 2023. Available online: http://dryades.units.it/floritaly/ (accessed on 6 March 2023).
- Biondi, E.; Blasi, C. Prodromo of Italian Vegetation. MATTM, SBI. 2015. Available online: https://www.prodromo-vegetazione-italia.org/ (accessed on 6 November 2023).
- Biondi, E.; Blasi, C.; Burrascano, S.; Casavecchia, S.; Copiz, R.; Del Vico, E.; Galdenzi, D.; Gigante, D.; Lasen, C.; Spampinato, G.; et al. Italian Interpretation Manual of the 92/43/EEC Habitats Directive. 2009. Available online: http://vnr.unipg.it/habitat/index.jsp (accessed on 6 November 2023).
- Gigante, D.; Attorre, F.; Venanzoni, R.; Acosta, A.T.R.; Agrillo, E.; Aleffi, M.; Alessi, N.; Allegrezza, M.; Angelini, P.; Angiolini, C.; et al. Un protocollo metodologico per il monitoraggio degli Habitat dell’Allegato I: Il contributo della scienza della Vegetazione. Sociol. Veg. 2016, 53, 77–87. [Google Scholar]
- Shannon, C.E.; Weaver, W. A Mathematical Theory of Communication; University of Illinois Press: Urbana, IL, USA, 1949. [Google Scholar]
- Pielou, E.C. The measurement of diversity in different types of biological collections. J. Theor. Biol. 1966, 13, 131–144. [Google Scholar] [CrossRef]
- Grunewald, R.; Schubert, H. The definition of a new plant diversity index “H′dune” to assess human damage on coastal dunes, derived from the Shannon entropy index H′. Ecol. Indic. 2007, 7, 1–21. [Google Scholar] [CrossRef]
- Pinna, M.S.; Cogoni, D.; Fenu, G.; Bacchetta, G. The conservation status and anthropogenic impacts assessments of Mediterranean coastal dunes. Estuar. Coast. Shelf Sci. 2015, 167, 25–31. [Google Scholar] [CrossRef]
- Pinna, M.S.; Bacchetta, G.; Cogoni, D.; Fenu, G. Is vegetation an indicator for evaluating the impact of tourism on the conservation status of Mediterranean coastal dunes? Sci. Total Environ. 2019, 674, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Calderisi, G.; Cogoni, D.; Loni, A.; Fenu, G. Difference between invasive alien and native vegetation in trapping beach litter: A focus on a typical sandy beach of W-Mediterranean Basin. Mar. Pollut. Bull. 2023, 192, 115065. [Google Scholar] [CrossRef]
- Zar, J.H. Biostatistical Analysis, 3rd ed.; Prentice Hall: Hoboken, NJ, USA, 1996. [Google Scholar]
- Farebrother, R.W. Pan’s procedure for the tail probabilities of the Durbin-Watson statistic. Appl. Stat. 1980, 29, 224–227. [Google Scholar] [CrossRef]
- Rousseeuw, P.J.; Van Driessen, K. Computing LTS Regression for Large Data Sets; Institute of Mathematical Statistics Bulletin, Citeseer, 1999. [Google Scholar]
- Warton, D.I.; Wright, I.J.; Falster, D.S.; Westoby, M. Bivariate line-fitting methods for allometry. Biol. Rev. 2006, 81, 259–291. [Google Scholar] [CrossRef] [PubMed]
- Wooldridge, J.M. Introductory Econometrics—A Modern Approach, 5th ed.; South-Western Cengage Learning: Boston, MA, USA, 2012. [Google Scholar]
- Brullo, S.; Scelsi, F.; Spampinato, G. Aristido caerulescentis-Hyparrhenion hirtae, alleanza nuova della classe Lygeo-Stipetea a distribuzione sud mediterraneo-macaronesica. Fitosociologia 1997, 32, 189–206. [Google Scholar]
- Brullo, S.; De Marco, G.; Signorello, P. Studio fitosociologico delle praterie a Lygeum spartum dell’Italia meridionale. Boll. Acc. Gioenia Sci. Nat. Catania 1990, 23, 561–579. [Google Scholar]
- Spampinato, G.; Musarella, C.M.; Cano-Ortiz, A.; Signorino, G. Habitat, occurrence and conservation status of the Saharo-Macaronesian and Southern-Mediterranean element Fagonia cretica L. (Zygophyllaceae) in Italy. J. Arid. Land 2018, 10, 140–151. [Google Scholar] [CrossRef]
- Viciani, D.; Gabellini, A. Contributo alla conoscenza della vegetazione del Pratomagno (Toscana orientale): Le praterie di crinale ed il complesso forestale regionale del versante casentinese. Webbia 2000, 55, 297–316. [Google Scholar] [CrossRef]
- Di Pietro, R.; De Santis, A.; Fortini, P.; Blasi, C. Geobotanical investigation on acidophilous grasslands in the Abruzzo, Lazio and Molise National Park. Lazaroa 2005, 26, 115–137. [Google Scholar]
- Venanzoni, R.; Gigante, D. Contribution to the study of the summit pastures of M. Tezio (Perugia, Italy). Phytosociology 1999, 36, 157–174. [Google Scholar]
- Brullo, S.; Cormaci, A.; Giusso Del Galdo, G.; Guarino, R.; Minissale, P.; Siracusa, G.; Spampinato, G. A syntaxonomical survey of the Sicilian dwarf shrub vegetation belonging to the class Rumici-Astragaletea siculi. Ann. Bot. 2005, 5, 57–104. [Google Scholar]
- Passalacqua, N.G.; Bernardo, L. Flora relitta di altitudine dell’Appennino meridionale: Quale origine? Biogeography 1998, 19, 105–117. [Google Scholar] [CrossRef]
- Di Pietro, R. New taxonomical data about genus Sesleria Scop. from southern Italy. In Proceedings of the XI Optima Meeting, Beograd, Serbia, 5–11 September 2004. Book of Abstract: 87. [Google Scholar]
- Venanzoni, R. Contributo alla conoscenza di prati umidi della Sila (Calabria-Italia). Doc. Phytosociol. 1988, 11, 613–633. [Google Scholar]
- Buffa, G.; Ghirelli, L.; Bracco, F.; Sburlino, G. Conservazione e biodiversità nella progettazione ambientale dell’area A.M.A.G. di Villaverla (VI). Noteb. I.A.E.D. 1997, 8, 56–58. [Google Scholar]
- Acosta, A.; Blasi, C.; Esposito, S.; Stanisci, A. Analisi della vegetazione delle dune costiere del Lazio centro-meridionale. Inform. Bot. Ital. 2000, 32, 5–10. [Google Scholar]
- Maiorca, G.; Spampinato, G.; Crisafulli, A. Carta della Vegetazione Reale della Foce del Fiume Crati (CS-Calabria); Progetto PHYTOS.I.S. Monograph no. 2; ARSSA: Cosenza, Italy, 2005. [Google Scholar]
- Brullo, S.; Spampinato, G. La vegetazione dei corsi d’acqua della Sicilia. Boll. Acc. Gioenia Sci. Nat. 1990, 23, 119–252. [Google Scholar]
- Gianguzzi, L.; La Mantia, A.; Lo Presti, R.M. Distribuzione, ecologia e status conservativo delle stazioni di Petagnaea gussonei (Sprengel) Rauschert (Apiaceae) nell’area dei Monti Nebrodi (Sicilia nord-orientale). Nat. Sicil. 2004, 28, 265–326. [Google Scholar]
- Mucina, L.; Bültmann, H.; Dierßen, K.; Theurillat, J.P.; Raus, T.; Čarni, A.; Šumberová, K.; Willner, W.; Dengler, J.; Gavilán García, R.; et al. Vegetation of Europe: Hierarchical floristic classification system of plant, lichen, and algal communities. Appl. Veg. Sci. 2016, 19, 3–264. [Google Scholar] [CrossRef]
- Biondi, E.; Galdenzi, D. Phytosociological analysis of the grasslands of Montagna dei Fiori (central Italy) and syntaxonomic review of the class Festuco-Brometea in the Apennines. Plant Sociol. 2012, 49, 91–112. [Google Scholar]
- Chytrý, M.; Dražil, T.; Hájek, M.; Kalníková, V.; Preislerová, Z.; Šibík, J.; Ujházy, K.; Axmanová, I.; Bernátová, D.; Blanár, D.; et al. The most species-rich plant communities in the Czech Republic and Slovakia (with new world records). Preslia 2015, 87, 217–278. [Google Scholar]
- Loiseau, P.; Louault, F.; Le Roux, X.; Bardy, M. Does extensification of rich grasslands alter the C and N cycles, directly or via species composition? Basic Appl. Ecol. 2005, 6, 275–287. [Google Scholar] [CrossRef]
- Rodriguez-Iturbe, I.; D’odorico, P.; Porporato, A.; Ridolfi, L.J.W.R.R. On the spatial and temporal links between vegetation, climate, and soil moisture. Water Resour. Res. 1999, 35, 3709–3722. [Google Scholar] [CrossRef]
- Myśliwy, M.; Pešić, V. Tall Herb Fringe Vegetation on Banks of Montenegrin Rivers as a Habitat Type of European Importance. Water 2023, 15, 3684. [Google Scholar] [CrossRef]
- Oroian, S.; Sămărghiţan, M.; Sămărghiţan, M. Phytosociogical research on wet meadows dominated by Scirpus sylvaticus L. in călimani and gurghiului mountains (mureș county). ABMJ 2019, 2, 49–59. [Google Scholar] [CrossRef]
- Aćić, S.; Šilc, U.; Petrović, M.; Tomović, G.; Dajić Stevanović, Z. Classification, ecology and biodiversity of Central Balkan dry grasslands. Tuexenia 2015, 35, 329–353. [Google Scholar]
- Utratna, M.; Okruszko, T. The Impact of Water Quality on the Special Areas of Conservation (Natura 2000) Located in the River Narew and River Warta Valleys. In Contemporary Problems of Management and Environmental Protection; 2011; Volume 7, Chapter 12. [Google Scholar]
- Foster, C.N.; Sato, C.F.; Lindenmayer, D.B.; Barton, P.S. Integrating theory into disturbance interaction experiments to better inform ecosystem management. Glob. Change Biol. 2016, 22, 1325–1335. [Google Scholar] [CrossRef]
- Facioni, L.; Del Vico, E.; Rosati, L.; Burrascano, S.; Tilia, A.; Blasi, C. Dry grasslands database of central Italy. In Vegetation Databases for the 21st Century; Dengler, J., Oldeland, J., Jansen, F., Chytrý, M., Ewald, J., Finckh, M., Glöckler, F., Lopez-Gonzalez, G., Peet, R.K., Schaminée, J.H.J., Eds.; Biodiversity and Ecology: Hamburg, Germany, 2012; Volume 4, p. 398. [Google Scholar]
- Lotze, H.K.; Lenihan, H.S.; Bourque, B.J.; Bradbury, R.H.; Cooke, R.G.; Kay, M.C.; Kidwell, S.M.; Kirby, M.X.; Peterson, C.H.; Jackson, J.B. Depletion, degradation, and recovery potential of estuaries and coastal seas. Science 2006, 312, 1806–1809. [Google Scholar] [CrossRef]
- Willems, J.H.; Peet, R.K.; Bik, L. Changes in chalk-grassland structure and species richness resulting from selective nutrient additions. J. Veg. Sci. 1993, 4, 203–212. [Google Scholar] [CrossRef]
- Henle, K.; Bauch, B.; Auliya, M.; Külvik, M.; Pe‘er, G.; Schmeller, D.S.; Framstad, E. Priorities for biodiversity monitoring in Europe: A review of supranational policies and a novel scheme for integrative prioritization. Ecol. Indic. 2013, 33, 5–18. [Google Scholar] [CrossRef]
- Hoffmann, S.; Beierkuhnlein, C.; Field, R.; Provenzale, A.; Chiarucci, A. Uniqueness of protected areas for conservation strategies in the European Union. Sci. Rep. 2018, 8, 6445. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).