
Quantifying Dieback in a Vulnerable Population of Eucalyptus macrorhyncha Using Remote Sensing

 ,
,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
1.1. Understanding Dieback
1.2. Dieback Monitoring through Applied Remote Sensing
1.3. Investigating Dieback Causes through Spatial Analysis
2. Methods
2.1. Study Area
2.2. Description of Imagery Data
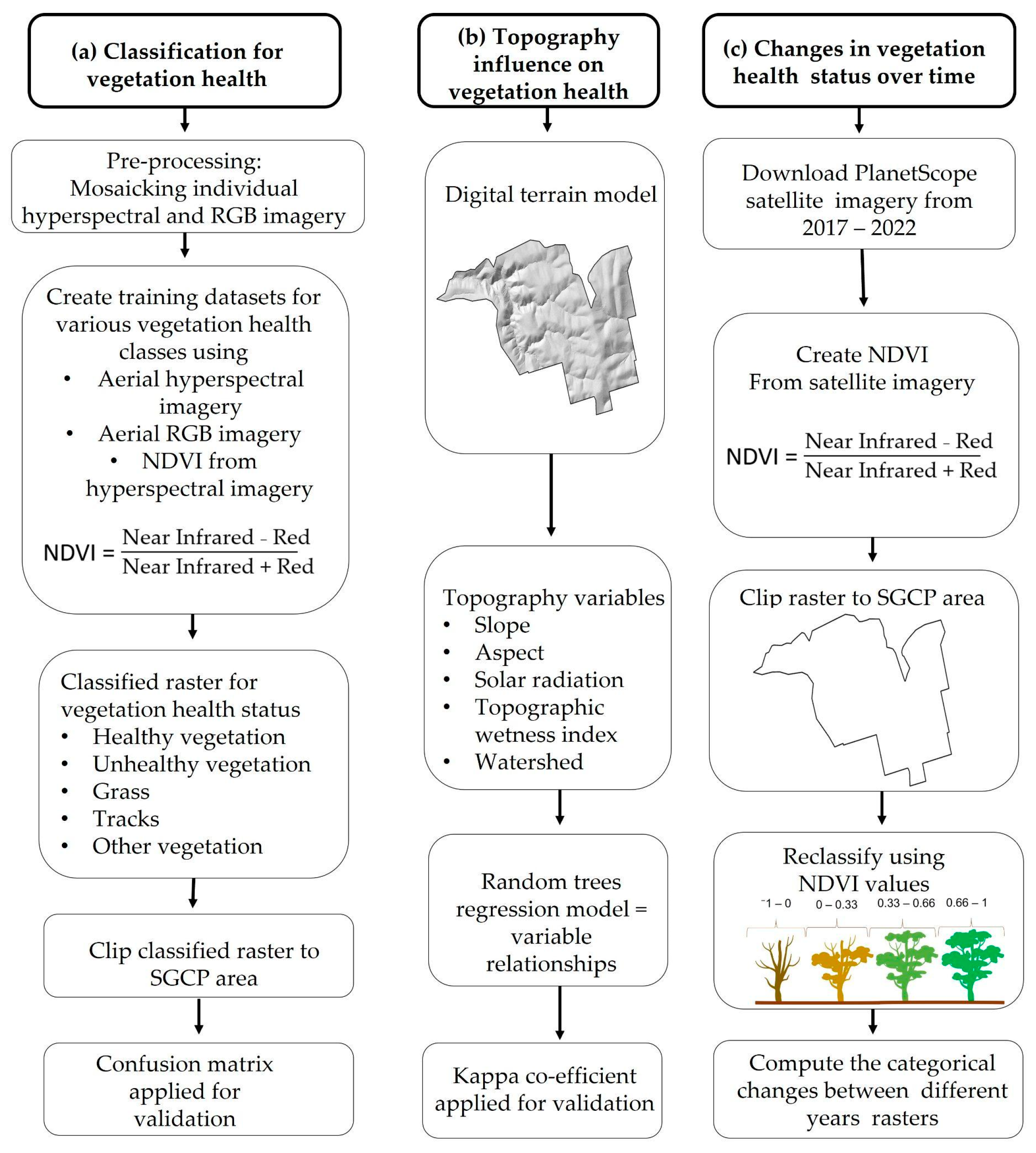
2.3. Quantifying Vegetation Health in Space and Time
2.3.1. Topography Influences in Vegetation Health Status
2.3.2. Vegetation Changes over Time Using Satellite Imagery
3. Results
3.1. Vegetation Health Status in 2022
3.2. Topography Influences on Vegetation Health Status
3.3. Vegetation Changes over Time Using Satellite Imagery
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sanchez-Salguero, R.; Camarero, J.J. Greater sensitivity to hotter droughts underlies juniper dieback and mortality in Mediterranean shrublands. Sci. Total Environ. 2020, 721, 137599. [Google Scholar] [CrossRef] [PubMed]
- Brouwers, N.; Matusick, G.; Ruthrof, K.; Lyons, T.; Hardy, G. Landscape-scale assessment of tree crown dieback following extreme drought and heat in a Mediterranean eucalypt forest ecosystem. Landsc. Ecol. 2012, 28, 69–80. [Google Scholar] [CrossRef]
- Matusick, G.; Ruthrof, K.X.; Kala, J.; Brouwers, N.C.; Breshears, D.D.; Hardy, G.E.S.J. Chronic historical drought legacy exacerbates tree mortality and crown dieback during acute heatwave-compounded drought. Environ. Res. Lett. 2018, 13, 95002. [Google Scholar] [CrossRef]
- Choat, B.; Brodribb, T.J.; Brodersen, C.R.; Duursma, R.A.; López, R.; Medlyn, B.E. Triggers of tree mortality under drought. Nature 2018, 558, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, A.A.; Rymer, P.D.; Byrne, M.; Ruthrof, K.X.; Whinam, J.; McGeoch, M.; Bergstrom, D.M.; Guerin, G.R.; Sparrow, B.; Joseph, L. Impacts of recent climate change on terrestrial flora and fauna: Some emerging Australian examples. Austral Ecol. 2019, 44, 3–27. [Google Scholar] [CrossRef]
- Jump, A.S.; Ruiz-Benito, P.; Greenwood, S.; Allen, C.D.; Kitzberger, T.; Fensham, R.; Martínez-Vilalta, J.; Lloret, F. Structural overshoot of tree growth with climate variability and the global spectrum of drought-induced forest dieback. Glob. Chang. Biol. 2017, 23, 3742–3757. [Google Scholar] [CrossRef] [PubMed]
- Eamus, D.; Boulain, N.; Cleverly, J.; Breshears, D.D. Global change-type drought-induced tree mortality: Vapor pressure deficit is more important than temperature per se in causing decline in tree health. Ecol. Evol. 2013, 3, 2711–2729. [Google Scholar] [CrossRef] [PubMed]
- Fensham, R.J.; Laffineur, B.; Allen, C.D. To what extent is drought-induced tree mortality a natural phenomenon? Glob. Ecol. Biogeogr. 2019, 28, 365–373. [Google Scholar] [CrossRef]
- Angel, A.S.; Bradley, J.S.; Davis, R. Impact of a prolonged decline in rainfall on eucalypt woodlands in southwestern Australia and its consequences for avifauna. Pac. Conserv. Biol. 2021, 28, 491–504. [Google Scholar] [CrossRef]
- Guarín, A.; Taylor, A.H. Drought triggered tree mortality in mixed conifer forests in Yosemite National Park, California, USA. For. Ecol. Manag. 2005, 218, 229–244. [Google Scholar] [CrossRef]
- Joffre, R.; Rambal, S.; Mediterranean Ecosystems. Encyclopedia of Life Sciences. 2001, p. 1. Available online: https://onlinelibrary.wiley.com/doi/10.1038/npg.els.0003196 (accessed on 25 July 2022).
- Specht, R. The effect of summer drought on vegetation structure in the Mediterranean climate region of Australia. In Plant Response to Stress: Functional Analysis in Mediterranean Ecosystems; Springer: Berlin/Heidelberg, Germany, 1987; pp. 625–639. [Google Scholar]
- Klausmeyer, K.R.; Shaw, M.R. Climate change, habitat loss, protected areas and the climate adaptation potential of species in Mediterranean ecosystems worldwide. PLoS ONE 2009, 4, e6392. [Google Scholar] [CrossRef]
- Yates, C.J.; Hobbs, R.J. Temperate eucalypt woodlands: A review of their status, processes threatening their persistence and techniques for restoration. Aust. J. Bot. 1997, 45, 949–973. [Google Scholar] [CrossRef]
- Breshears, D.D.; Cobb, N.S.; Rich, P.M.; Price, K.P.; Allen, C.D.; Balice, R.G.; Romme, W.H.; Kastens, J.H.; Floyd, M.L.; Belnap, J. Regional vegetation die-off in response to global-change-type drought. Proc. Natl. Acad. Sci. USA 2005, 102, 15144–15148. [Google Scholar] [CrossRef] [PubMed]
- Carnicer, J.; Coll, M.; Ninyerola, M.; Pons, X.; Sanchez, G.; Penuelas, J. Widespread crown condition decline, food web disruption, and amplified tree mortality with increased climate change-type drought. Proc. Natl. Acad. Sci. USA 2011, 108, 1474–1478. [Google Scholar] [CrossRef] [PubMed]
- Nicolle, D. Native Eucalypts of South Australia; Dean Nicolle: Adelaide, Australia, 2013. [Google Scholar]
- De Kauwe, M.G.; Sabot, M.E.; Medlyn, B.E.; Pitman, A.J.; Meir, P.; Cernusak, L.A.; Gallagher, R.V.; Ukkola, A.M.; Rifai, S.W.; Choat, B. Towards species-level forecasts of drought-induced tree mortality risk. New Phytol. 2022, 235, 94–110. [Google Scholar] [CrossRef]
- Pritzkow, C.; Szota, C.; Williamson, V.; Arndt, S.K. Previous drought exposure leads to greater drought resistance in eucalypts through changes in morphology rather than physiology. Tree Physiol. 2021, 41, 1186–1198. [Google Scholar] [CrossRef] [PubMed]
- Wrigley, J.; Fagg, M. Eucalypts: A Celebration; Allen & Unwin: Crows Nest, Australia, 2010. [Google Scholar]
- Saadaoui, E.; Yahia, K.B.; Dhahri, S.; Jamaa, M.L.B.; Khouja, M.L. An overview of adaptative responses to drought stress in spp. For. Stud. 2017, 67, 86–96. [Google Scholar] [CrossRef]
- Matusick, G.; Ruthrof, K.X.; Fontaine, J.B.; Hardy, G.E.S.J.; Gilliam, F. Eucalyptusforest shows low structural resistance and resilience to climate change-type drought. J. Veg. Sci. 2016, 27, 493–503. [Google Scholar] [CrossRef]
- Jurskis, V.; Turner, J. Eucalypt dieback in eastern Australia: A simple model. Aust. For. 2002, 65, 87–98. [Google Scholar] [CrossRef]
- De Kauwe, M.G.; Medlyn, B.E.; Ukkola, A.M.; Mu, M.; Sabot, M.E.; Pitman, A.J.; Meir, P.; Cernusak, L.A.; Rifai, S.W.; Choat, B. Identifying areas at risk of drought-induced tree mortality across South-Eastern Australia. Glob. Chang. Biol. 2020, 26, 5716–5733. [Google Scholar] [CrossRef]
- Matusick, G.; Ruthrof, K.X.; Brouwers, N.C.; Dell, B.; Hardy, G.S.J. Sudden forest canopy collapse corresponding with extreme drought and heat in a Mediterranean-type eucalypt forest in southwestern Australia. Eur. J. For. Res. 2013, 132, 497–510. [Google Scholar] [CrossRef]
- Jurskis, V. Eucalypt decline in Australia, and a general concept of tree decline and dieback. For. Ecol. Manag. 2005, 215, 1–20. [Google Scholar] [CrossRef]
- Mueller-Dombois, D. Perspectives for an aetiology of stand-level dieback. Annu. Rev. Ecol. Syst. 1986, 17, 221–243. [Google Scholar] [CrossRef]
- Cai, W.; Purich, A.; Cowan, T.; van Rensch, P.; Weller, E. Did climate change–induced rainfall trends contribute to the Australian Millennium Drought? J. Clim. 2014, 27, 3145–3168. [Google Scholar] [CrossRef]
- Mueller-Dombois, D. Natural Dieback in Forests. Bioscience 1987, 37, 575–583. [Google Scholar] [CrossRef]
- Fensham, R.J.; Freeman, M.E.; Laffineur, B.; Macdermott, H.; Prior, L.D.; Werner, P.A. Variable rainfall has a greater effect than fire on the demography of the dominant tree in a semi-arid Eucalyptus savanna. Austral Ecol. 2017, 42, 772–782. [Google Scholar] [CrossRef]
- Clark, D.K. Climate Change Hits Clare Valley. 2021. Available online: comagecontra.net (accessed on 10 October 2022).
- Hartmann, H.; Schuldt, B.; Sanders, T.G.M.; Macinnis-Ng, C.; Boehmer, H.J.; Allen, C.D.; Bolte, A.; Crowther, T.W.; Hansen, M.C.; Medlyn, B.E.; et al. Monitoring global tree mortality patterns and trends. Report from the VW symposium ‘Crossing scales and disciplines to identify global trends of tree mortality as indicators of forest health’. New Phytol. 2018, 217, 984–987. [Google Scholar] [CrossRef]
- DEW. Regional Species Conservation Assessment Project; Department of Environment and Water, Government of South Australia: Adelaide, Australia, 2022.
- Keppel, G.; Sarnow, U.; Biffin, E.; Peters, S.; Fitzgerald, D.; Boutsalis, E.; Waycott, M.; Guerin, G.R. Population decline in a Pleistocene refugium: Stepwise, drought-related dieback of a South Australian eucalypt. Sci. Total Environ. 2023, 876, 162697. [Google Scholar] [CrossRef]
- Sarnow, U. Red Stringybark (E. macrorhyncha) Decline in South Australia—Tree Health Status Trend and Potential Causes. Master’s Thesis, University of South Australia, Adelaide, Australia, 2021. [Google Scholar]
- Guerin, G.R.; Keppel, G.; Peters, S.; Nolan, R.H.; Sarnow, U.; Bradshaw, H.; Morgan, R.; Fitzgerald, D.; Boland, J.E.B. Dieback of Stringybark Forests in the Mount Lofty Ranges—Phase 1; University of Adelaide: Adelaide, Australia; University of South Australia Adelaide: Adelaide, Australia, 2022. [Google Scholar]
- Masson-Delmotte, V.; Zhai, P.; Pirani, S.; Connors, C.; Péan, S.; Berger, N.; Caud, Y.; Chen, L.; Goldfarb, M.; Scheel Monteiro, P.M. IPCC, 2021: Summary for policymakers. In Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Singapore, 2021. [Google Scholar]
- Congalton, R.G. Remote sensing: An overview. GISci. Remote Sens. 2010, 47, 443–459. [Google Scholar] [CrossRef]
- Sparrow, B.D.; Foulkes, J.N.; Wardle, G.M.; Leitch, E.J.; Caddy-Retalic, S.; Van Leeuwen, S.J.; Tokmakoff, A.; Thurgate, N.Y.; Guerin, G.R.; Lowe, A.J. A vegetation and soil survey method for surveillance monitoring of rangeland environments. Front. Ecol. Evol. 2020, 8, 157. [Google Scholar] [CrossRef]
- Jensen, J.R. Remote Sensing of the Environment: An Earth Resource Perspective 2/e; Pearson Education India: New Delhi, India, 2009. [Google Scholar]
- Mishra, S.; Shrivastava, P.; Dhurvey, P. Change detection techniques in remote sensing: A review. Int. J. Wirel. Mob. Commun. Ind. Syst. 2017, 4, 1–8. [Google Scholar] [CrossRef]
- Evans, B.; Lyons, T.J.; Barber, P.A.; Stone, C.; Hardy, G. Dieback classification modelling using high-resolution digital multispectral imagery and in situ assessments of crown condition. Remote Sens. Lett. 2012, 3, 541–550. [Google Scholar] [CrossRef]
- Shendryk, I.; Broich, M.; Tulbure, M.G.; McGrath, A.; Keith, D.; Alexandrov, S.V. Mapping individual tree health using full-waveform airborne laser scans and imaging spectroscopy: A case study for a floodplain eucalypt forest. Remote Sens. Environ. 2016, 187, 202–217. [Google Scholar] [CrossRef]
- Li, H.; Wang, C.; Ellis, J.T.; Cui, Y.; Miller, G.; Morris, J.T. Identifying marsh dieback events from Landsat image series (1998–2018) with an Autoencoder in the NIWB estuary, South Carolina. Int. J. Digit. Earth 2020, 13, 1467–1483. [Google Scholar] [CrossRef]
- Cunningham, S.C.; Thomson, J.R.; Read, J.; Baker, P.J.; Nally, R.M. Does stand structure influence susceptibility of eucalypt floodplain forests to dieback? Austral Ecol. 2010, 35, 348–356. [Google Scholar] [CrossRef]
- Dickson, C.R.; Baker, D.J.; Bergstrom, D.M.; Brookes, R.H.; Whinam, J.; McGeoch, M.A. Widespread dieback in a foundation species on a sub-Antarctic World Heritage Island: Fine-scale patterns and likely drivers. Austral Ecol. 2020, 46, 52–64. [Google Scholar] [CrossRef]
- Huang, C.-y.; Anderegg, W.R.L.; Asner, G.P. Remote sensing of forest die-off in the Anthropocene: From plant ecophysiology to canopy structure. Remote Sens. Environ. 2019, 231, 111233. [Google Scholar] [CrossRef]
- Evans, P.M.; Newton, A.C.; Cantarello, E.; Martin, P.; Sanderson, N.; Jones, D.L.; Barsoum, N.; Cottrell, J.E.; A’Hara, S.W.; Fuller, L. Thresholds of biodiversity and ecosystem function in a forest ecosystem undergoing dieback. Sci. Rep. 2017, 7, 6775. [Google Scholar] [CrossRef] [PubMed]
- Camarero, J.J. The drought–dieback–death conundrum in trees and forests. Plant Ecol. Divers. 2021, 14, 1–12. [Google Scholar] [CrossRef]
- Hoffmann, W.A.; Marchin, R.M.; Abit, P.; Lau, O.L. Hydraulic failure and tree dieback are associated with high wood density in a temperate forest under extreme drought. Glob. Chang. Biol. 2011, 17, 2731–2742. [Google Scholar] [CrossRef]
- Nolan, R.H.; Gauthey, A.; Losso, A.; Medlyn, B.E.; Smith, R.; Chhajed, S.S.; Fuller, K.; Song, M.; Li, X.; Beaumont, L.J. Hydraulic failure and tree size linked with canopy die-back in eucalypt forest during extreme drought. New Phytol. 2021, 230, 1354–1365. [Google Scholar] [CrossRef]
- Allen, K.J.; Verdon-Kidd, D.C.; Sippo, J.Z.; Baker, P.J. Compound climate extremes driving recent sub-continental tree mortality in northern Australia have no precedent in recent centuries. Sci. Rep. 2021, 11, 18337. [Google Scholar] [CrossRef] [PubMed]
- Seaton, S.; Matusick, G.; Ruthrof, K.X.; Hardy, G.E.S.J. Outbreak of Phoracantha semipunctata in response to severe drought in a Mediterranean Eucalyptus forest. Forests 2015, 6, 3868–3881. [Google Scholar] [CrossRef]
- da Conceição Caldeira, M.; Fernandéz, V.; Tomé, J.; Pereira, J.S. Positive effect of drought on longicorn borer larval survival and growth on Eucalyptus trunks. Ann. For. Sci. 2002, 59, 99–106. [Google Scholar] [CrossRef]
- Landsberg, J.; Wylie, F. Dieback of rural trees in Australia. GeoJournal 1988, 17, 231–237. [Google Scholar] [CrossRef]
- Stone, C. Assessment and monitoring of decline and dieback of forest eucalypts in relation to ecologically sustainable forest management: A review with a case study. Aust. For. 1999, 62, 51–58. [Google Scholar] [CrossRef]
- Chen, Y.J.; Choat, B.; Sterck, F.; Maenpuen, P.; Katabuchi, M.; Zhang, S.B.; Tomlinson, K.W.; Oliveira, R.S.; Zhang, Y.J.; Shen, J.X. Hydraulic prediction of drought-induced plant dieback and top-kill depends on leaf habit and growth form. Ecol. Lett. 2021, 24, 2350–2363. [Google Scholar] [CrossRef]
- Ruthrof, K.X.; Matusick, G.; Hardy, G.E.S.J. Early differential responses of co-dominant canopy species to sudden and severe drought in a Mediterranean-climate type forest. Forests 2015, 6, 2082–2091. [Google Scholar] [CrossRef]
- Batllori, E.; Lloret, F.; Aakala, T.; Anderegg, W.R.; Aynekulu, E.; Bendixsen, D.P.; Bentouati, A.; Bigler, C.; Burk, C.J.; Camarero, J.J. Forest and woodland replacement patterns following drought-related mortality. Ecology 2020, 117, 29720–29729. [Google Scholar] [CrossRef]
- Anderegg, W.R.; Kane, J.M.; Anderegg, L.D. Consequences of widespread tree mortality triggered by drought and temperature stress. Nat. Clim. Chang. 2013, 3, 30–36. [Google Scholar] [CrossRef]
- Jara-Guerrero, A.; Gonzalez-Sanchez, D.; Escudero, A.; Espinosa, C.I. Chronic disturbance in a tropical dry forest: Disentangling direct and indirect pathways behind the loss of plant richness. Front. For. Glob. Chang. 2021, 4, 723985. [Google Scholar] [CrossRef]
- Levick, S.; Setterfield, S.; Rossiter-Rachor, N.; Hutley, L.; McMaster, D.; Hacker, J. Monitoring the Distribution and Dynamics of an Invasive Grass in Tropical Savanna Using Airborne LiDAR. Remote Sens. 2015, 7, 5117–5132. [Google Scholar] [CrossRef]
- Rose, R.A.; Byler, D.; Eastman, J.R.; Fleishman, E.; Geller, G.; Goetz, S.; Guild, L.; Hamilton, H.; Hansen, M.; Headley, R. Ten ways remote sensing can contribute to conservation. Conserv. Biol. 2015, 29, 350–359. [Google Scholar] [CrossRef] [PubMed]
- Sisodia, P.S.; Tiwari, V.; Kumar, A. Analysis of supervised maximum likelihood classification for remote sensing image. In Proceedings of the International Conference on Recent Advances and Innovations in Engineering (ICRAIE-2014), Jaipur, India, 9–11 May 2014; pp. 1–4. [Google Scholar]
- Sykas, D. Spectral Indicies in Remote Sensing and How to Interpret Them. Available online: https://www.geo.university/pages/spectral-indices-in-remote-sensing-and-how-to-interpret-them (accessed on 10 October 2022).
- Pettorelli, N.; Vik, J.O.; Mysterud, A.; Gaillard, J.-M.; Tucker, C.J.; Stenseth, N.C. Using the satellite-derived NDVI to assess ecological responses to environmental change. Trends Ecol. Evol. 2005, 20, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Pettorelli, N. The Normalized Difference Vegetation Index; Oxford University Press: New York, NY, USA, 2013. [Google Scholar]
- Verma, N.K.; Lamb, D.W.; Sinha, P. Airborne LiDAR and high resolution multispectral data integration in Eucalyptus tree species mapping in an Australian farmscape. Geocarto Int. 2022, 37, 70–90. [Google Scholar] [CrossRef]
- Karna, Y.K.; Penman, T.D.; Aponte, C.; Bennett, L.T. Assessing Legacy Effects of Wildfires on the Crown Structure of Fire-Tolerant Eucalypt Trees Using Airborne LiDAR Data. Remote Sens. 2019, 11, 2433. [Google Scholar] [CrossRef]
- Cotrozzi, L. Spectroscopic detection of forest diseases: A review (1970–2020). J. For. Res. 2021, 33, 21–38. [Google Scholar] [CrossRef]
- Exelis VIS. ENVI 5.3; Exelis VIS: Boulder, CO, USA, 2015. [Google Scholar]
- Näsi, R.; Honkavaara, E.; Lyytikäinen-Saarenmaa, P.; Blomqvist, M.; Litkey, P.; Hakala, T.; Viljanen, N.; Kantola, T.; Tanhuanpää, T.; Holopainen, M. Using UAV-Based Photogrammetry and Hyperspectral Imaging for Mapping Bark Beetle Damage at Tree-Level. Remote Sens. 2015, 7, 15467–15493. [Google Scholar] [CrossRef]
- Mitchell, P.J.; Benyon, R.G.; Lane, P.N. Responses of evapotranspiration at different topographic positions and catchment water balance following a pronounced drought in a mixed species eucalypt forest, Australia. J. Hydrol. 2012, 440, 62–74. [Google Scholar] [CrossRef]
- Hawthorne, S.; Miniat, C.F. Topography may mitigate drought effects on vegetation along a hillslope gradient. Ecohydrology 2018, 11, e1825. [Google Scholar] [CrossRef]
- Tian, Y.; Davies-Colley, R.; Gong, P.; Thorrold, B. Estimating solar radiation on slopes of arbitrary aspect. Agric. For. Meteorol. 2001, 109, 67–74. [Google Scholar] [CrossRef]
- Auslander, M.; Nevo, E.; Inbar, M. The effects of slope orientation on plant growth, developmental instability and susceptibility to herbivores. J. Arid Environ. 2003, 55, 405–416. [Google Scholar] [CrossRef]
- ESRI. ArcGIS Pro. 2019. Available online: https://www.esri.com/en-us/arcgis/products/arcgis-pro/resources (accessed on 12 July 2022).
- Hollunder, R.K.; Mariotte, P.; Carrijo, T.T.; Holmgren, M.; Luber, J.; Stein-Soares, B.; Guidoni-Martins, K.G.; Ferreira-Santos, K.; Scarano, F.R.; Garbin, M.L. Topography and vegetation structure mediate drought impacts on the understory of the South American Atlantic Forest. Sci. Total Environ. 2021, 766, 144234. [Google Scholar] [CrossRef]
- Moeslund, J.E.; Arge, L.; Bøcher, P.K.; Dalgaard, T.; Odgaard, M.V.; Nygaard, B.; Svenning, J.-C. Topographically controlled soil moisture is the primary driver of local vegetation patterns across a lowland region. Ecosphere 2013, 4, 1–26. [Google Scholar] [CrossRef]
- BOM. Climate Statistics for Australian Locations; BOM: Melbourne, Australian, 2022. [Google Scholar]
- DENR. Spring Gully Conservation Park (Visitor’s Pamphlet); DENR: Quezon City, Philippines, 2010. [Google Scholar]
- SASCC. Eucalyptus macrorhyncha ssp. In Macrorhyncha (Myrtaceae): Red Stringybark. Seeds of South Australia.; 2021. Available online: environment.sa.gov.au (accessed on 10 November 2021).
- AgiSoft. AgiSoft Program. 2022. Available online: https://www.agisoft.com/ (accessed on 30 July 2022).
- Rwanga, S.S.; Ndambuki, J.M. Accuracy assessment of land use/land cover classification using remote sensing and GIS. Int. J. Geosci. 2017, 8, 611. [Google Scholar] [CrossRef]
- Stehman, S. Estimating the kappa coefficient and its variance under stratified random sampling. Photogramm. Eng. Remote Sens. 1996, 62, 401–407. [Google Scholar]
- LLC. Global Mapper Version 9.0 Software. 2009. Available online: https://www.bluemarblegeo.com/global-mapper/ (accessed on 30 July 2022).
- Sörensen, R.; Zinko, U.; Seibert, J. On the calculation of the topographic wetness index: Evaluation of different methods based on field observations. Hydrol. Earth Syst. Sci. 2006, 10, 101–112. [Google Scholar] [CrossRef]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Wu, X.; Xiao, Q.; Wen, J.; You, D.; Hueni, A. Advances in quantitative remote sensing product validation: Overview and current status. Earth-Sci. Rev. 2019, 196, 102875. [Google Scholar] [CrossRef]
- Huang, S.; Tang, L.; Hupy, J.P.; Wang, Y.; Shao, G. A commentary review on the use of normalized difference vegetation index (NDVI) in the era of popular remote sensing. J. For. Res. 2021, 32, 1–6. [Google Scholar] [CrossRef]
- Xue, J.; Su, B. Significant remote sensing vegetation indices: A review of developments and applications. J. Sens. 2017, 201, 13536917. [Google Scholar] [CrossRef]
- Hussain, M.; Chen, D.; Cheng, A.; Wei, H.; Stanley, D. Change detection from remotely sensed images: From pixel-based to object-based approaches. ISPRS J. Photogramm. Remote Sens. 2013, 80, 91–106. [Google Scholar] [CrossRef]
- Almalki, K.A.; Bantan, R.A.; Hashem, H.I.; Loni, O.A.; Ali, M.A. Improving geological mapping of the Farasan Islands using remote sensing and ground-truth data. J. Maps 2017, 13, 900–908. [Google Scholar] [CrossRef]
- Hegarty-Craver, M.; Polly, J.; O’Neil, M.; Ujeneza, N.; Rineer, J.; Beach, R.H.; Lapidus, D.; Temple, D.S. Remote crop mapping at scale: Using satellite imagery and UAV-acquired data as ground truth. Remote Sens. 2020, 12, 1984. [Google Scholar] [CrossRef]
- Samiappan, S.; Prasad, S.; Bruce, L.M.; Robles, W. NASA’s upcoming HyspIRI mission—Precision vegetation mapping with limited ground truth. In Proceedings of the 2010 IEEE International Geoscience and Remote Sensing Symposium, Honolulu, HI, USA, 25–30 July 2010; pp. 3744–3747. [Google Scholar]
- Wang, B.; Zhang, G.; Duan, J. Relationship between topography and the distribution of understory vegetation in a Pinus massoniana forest in Southern China. Int. Soil Water Conserv. Res. 2015, 3, 291–304. [Google Scholar] [CrossRef]
- Knudby, A. Change detection. In Remote Sensing; Electronic book. 2021. Available online: https://ecampusontario.pressbooks.pub/remotesensing/chapter/chapter-8-change-detection/ (accessed on 29 April 2022).
- Heisig, J. Detecting Drought Effects on Tree Mortality in Forests of Franconia with Remote Sensing. Master’s Thesis, University of Bayreuth, Bayreuth, Germany, 2020. [Google Scholar]
- Whyte, G.; Howard, K.; Hardy, G.S.J.; Burgess, T. The tree decline recovery seesaw; a conceptual model of the decline and recovery of drought stressed plantation trees. For. Ecol. Manag. 2016, 370, 102–113. [Google Scholar] [CrossRef]
- McDowell, N.; Pockman, W.T.; Allen, C.D.; Breshears, D.D.; Cobb, N.; Kolb, T.; Plaut, J.; Sperry, J.; West, A.; Williams, D.G. Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? New Phytol. 2008, 178, 719–739. [Google Scholar] [CrossRef]
- Bachelard, E. Studies on the formation of epicormic shoots on eucalypt stem segments. Aust. J. Biol. Sci. 1969, 22, 1291–1296. [Google Scholar] [CrossRef]
- Huang, C.-Y.; Asner, G.P.; Barger, N.N.; Neff, J.C.; Floyd, M.L. Regional aboveground live carbon losses due to drought-induced tree dieback in piñon–juniper ecosystems. Remote Sens. Environ. 2010, 114, 1471–1479. [Google Scholar] [CrossRef]
- Wang, W.; Peng, C.; Kneeshaw, D.D.; Larocque, G.R.; Luo, Z. Drought-induced tree mortality: Ecological consequences, causes, and modelling. Environ. Rev. 2012, 20, 109–121. [Google Scholar] [CrossRef]
- Guerin, G.R.; Keppel, G.; Peters, S.; Hurren, A. Dieback of stringybark eucalypt forests in the Mount Lofty Ranges. Trans. R. Soc. South Aust. 2023, 147, 17–38. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fitzgerald, D.L.; Peters, S.; Guerin, G.R.; McGrath, A.; Keppel, G. Quantifying Dieback in a Vulnerable Population of Eucalyptus macrorhyncha Using Remote Sensing. Land 2023, 12, 1271. https://doi.org/10.3390/land12071271
Fitzgerald DL, Peters S, Guerin GR, McGrath A, Keppel G. Quantifying Dieback in a Vulnerable Population of Eucalyptus macrorhyncha Using Remote Sensing. Land. 2023; 12(7):1271. https://doi.org/10.3390/land12071271
Chicago/Turabian StyleFitzgerald, Donna L., Stefan Peters, Gregory R. Guerin, Andrew McGrath, and Gunnar Keppel. 2023. "Quantifying Dieback in a Vulnerable Population of Eucalyptus macrorhyncha Using Remote Sensing" Land 12, no. 7: 1271. https://doi.org/10.3390/land12071271
APA StyleFitzgerald, D. L., Peters, S., Guerin, G. R., McGrath, A., & Keppel, G. (2023). Quantifying Dieback in a Vulnerable Population of Eucalyptus macrorhyncha Using Remote Sensing. Land, 12(7), 1271. https://doi.org/10.3390/land12071271