
Orographic Factors as a Predictor of the Spread of the Siberian Silk Moth Outbreak in the Mountainous Southern Taiga Forests of Siberia

 , and
, and
Abstract
1. Introduction
- -
- to reveal the association between an insect outbreak occurrence and certain elements of the relief and analyze the spatiotemporal dynamics of the Siberian silk moth outbreak pattern based on the orographic features of the territory; and
- -
- to assess the prospects for the early detection of Siberian silk moth outbreaks using the known patterns in species ecology and remote sensing methods in mountainous southern taiga forests conditions.
2. Materials and Methods
2.1. Study Area
2.2. Satellite Data Collection and Analysis
2.3. On-Ground Studies
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Global Invasive Species Database. 2020. Available online: http://www.iucngisd.org/gisd/search.php (accessed on 10 December 2020).
- Isaev, A.S.; Palnikova, E.N.; Sukhovolsky, V.G.; Tarasova, O.V. Population Dynamics of Defoliating Forest Insects: Models and Forecasts; KMK Scientific Press Ltd.: Moscow, Russia, 2015; p. 262. [Google Scholar]
- Rozhkov, A.S. Outbreaks of Siberian Silk Moth and Measures of Their Control; Nauka: Moscow, Russia, 1965; p. 179. [Google Scholar]
- Kondakov, Y.P. Mass reproduction of Siberian silk moth. In Ekologiya Populyatsii Lesnykh Zhivotnykh Sibiri (Population Ecology of Forest Animals in Siberia); Nauka: Novosibirsk, Russia, 1974; pp. 206–264. [Google Scholar]
- Florov, D.N. Pests of Siberian Forests; OGIX: Irkutsk, Russia, 1948; p. 143. [Google Scholar]
- Foster, J.R.; Townsend, P.A.; Mladenoff, D.J. Spatial dynamics of a gypsy moth defoliation outbreak and dependence on habitat characteristics. Landsc. Ecol. 2013, 28, 1307–1320. [Google Scholar] [CrossRef]
- Nifontov, S.V.; Savchenko, A.A.; Gridnev, A.N. Monitoring the development and forecasting of the mass reproduction (gradation) outbreaks of the Dendrolimus sibiricus Tschetv. in the Khabarovsk region. In IOP Conference Series: Earth and Environmental Science; IOP Publishing: Bristol, UK, 2019; Volume 316. [Google Scholar] [CrossRef]
- Zhirin, V.M.; Knyazeva, S.V.; Eydlina, S.P. Long-term dynamics of vegetation indices in dark coniferous forest after Siberian moth disturbance. Contemp. Probl. Ecol. 2016, 9, 834–843. [Google Scholar] [CrossRef]
- Kirichenko, N.I.; Baranchikov, Y.N.; Vidal, S. Performance of the potentially invasive Siberian moth Dendrolimus superans sibiricus on coniferous species in Europe. Agric. For. Entomol. 2009, 11, 247–254. [Google Scholar] [CrossRef]
- Kharuk, V.I.; Demidko, D.A.; Fedotova, E.V.; Dvinskaya, M.L.; Budnik, U.A. Spatial and temporal dynamics of Siberian silk moth large-scale outbreak in dark-needle coniferous tree stands in Altai. Contemp. Probl. Ecol. 2016, 9, 711–720. [Google Scholar] [CrossRef]
- Flø, D.; Rafoss, T.; Wendell, M.; Sundheim, L. The Siberian moth (Dendrolimus sibiricus), a pest risk assessment for Norway. For. Ecosyst. 2020, 7, 48. [Google Scholar] [CrossRef]
- Zhuravlev, P.G. Recommendations for Siberian Silkworm Control in the Forests of Russian Far East; FEFRI: Khabarovsk, Russia, 1960; p. 33. [Google Scholar]
- Kharuk, V.I.; Im, S.T.; Soldatov, V.V. Siberian silkmoth outbreaks surpassed geoclimatic barrier in Siberian Mountains. J. Mt. Sci. 2020, 17, 1891–1900. [Google Scholar] [CrossRef]
- Okunev, P.P. Geographical distribution and zones of injuriousness of Siberian silk moth. Geogr. Probl. For. 1955, 5, 13–24. [Google Scholar]
- Kharuk, V.I.; Ranson, K.J.; Kuzmichev, V.V.; Im, S.T. Landsat-based analysis of insect outbreaks in southern Siberia. Can. J. Remote Sens. 2003, 29, 286–297. [Google Scholar] [CrossRef]
- Kharuk, V.I.; Im, S.T.; Dvinskaya, M.L.; Golukov, A.S.; Ranson, K.J. Climate-induced mortality of spruce stands in Belarus. Environ. Res. Lett. 2015, 10, 125006. [Google Scholar] [CrossRef]
- Thompson, L.M.; Faske, T.M.; Banahene, N.; Grim, D.; Agosta, S.J.; Parry, D.; Tobin, P.C.; Johnson, D.M.; Grayson, K.L. Variation in growth and developmental responses to supraoptimal temperatures near latitudinal range limits of gypsy moth Lymantria dispar (L.), an expanding invasive species. Physiol. Entomol. 2017, 42, 181–190. [Google Scholar] [CrossRef]
- Isaev, A.S.; Ryapolov, B.Y. Analysis of the landscape-ecological confinement of the Siberian silkworm outbreaks using aerial survey. In Investigation of Taiga Landscapes Using Methods of Remote Sensing; Nauka: Novosibirsk, Russia, 1979; pp. 152–166. [Google Scholar]
- Ryapolov, V.Y. Methods of creating maps of forest damage caused by insect pests. Geogr. Nat. Resour. 1985, 2, 97–106. [Google Scholar]
- Egan, J.M.; Kaiden, J.; Lestina, J.; Stasey, A.; Jenne, J.L. Techniques to Enhance Assessment and Reporting of Pest. Damage Estimated with Aerial Detection Surveys; R1–19-09; U.S. Department of Agriculture, Forest Service, Northern Region, Forest Health Protection: Missoula, MT, USA, 2019; p. 33.
- Meddens, A.J.H.; Hicke, J.A.; Vierling, L.A. Evaluating the potential of multispectral imagery to map multiple stages of tree mortality. Remote Sens. Environ. 2011, 115, 1632–1642. [Google Scholar] [CrossRef]
- Lyamtsev, N.I. Forecasting Gypsy Moth Outbreaks, a Threat to Oak Forests and the Necessity of Preventing Measure; VNIILM: Pushkino, Russia, 2018; p. 84. [Google Scholar]
- Townsend, P.A.; Singh, A.; Foster, J.R.; Rehberg, N.J.; Kingdon, C.C.; Eshleman, K.N.; Seagle, S.W. A general Landsat model to predict canopy defoliation in broadleaf deciduous forests. Remote Sens. Environ. 2012, 119, 255–265. [Google Scholar] [CrossRef]
- Buotte, P.C.; Hicke, J.A.; Preisler, H.K.; Abatzoglou, J.T.; Raffa, K.F.; Logan, J.A. Climate influences on whitebark pine mortality from mountain pine beetle in the Greater Yellowstone Ecosystem. Ecol. Appl. 2016, 26, 2507–2524. [Google Scholar] [CrossRef]
- Creeden, E.P.; Hicke, J.A.; Buotte, P.C. Climate, weather, and recent mountain pine beetle outbreaks in the western United States. For. Ecol. Manag. 2014, 312, 239–251. [Google Scholar] [CrossRef]
- Polikarpov, N.P.; Chebakova, N.M.; Nazimova, D.I. Climate and Mountain Forests in Southern Siberia; Nauka: Novosibirsk, Russia, 1986; p. 225. [Google Scholar]
- Smagin, V.N.; Ilinskaya, S.A.; Nazimova, D.I.; Novoseltseva, I.F.; Cherednikova, J.S. Forest Types of Mountains of Southern Siberia; Nauka: Novosibirsk, Russia, 1980; p. 336. [Google Scholar]
- Bright, B.C.; Hicke, J.A.; Hudak, A.T. Estimating aboveground carbon stocks of a forest affected by mountain pine beetle in Idaho using lidar and multispectral imagery. Remote Sens. Environ. 2012, 124, 270–281. [Google Scholar] [CrossRef]
- Bright, B.C.; Hudak, A.T.; Egan, J.M.; Jorgensen, C.L.; Rex, F.E.; Hicke, J.A.; Meddens, A.J.H. Using satellite imagery to evaluate bark beetle-caused tree mortality reported in aerial surveys in a mixed conifer forest in Northern Idaho, USA. Forests 2020, 11, 529. [Google Scholar] [CrossRef]
- Wulder, M.A.; Dymond, C.C.; White, J.C.; Leckie, D.G.; Carroll, A.L. Surveying mountain pine beetle damage of forests: A review of remote sensing opportunities. For. Ecol. Manag. 2006, 221, 27–41. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 20 July 2020).
- Liaw, A.; Wiener, M. Classification and regression by random forest. R News 2002, 2, 18–22. [Google Scholar]
- Chen, T.; He, T.; Benesty, M.; Khotilovich, V.; Tang, Y.; Cho, H.; Chen, K.; Mitchell, R.; Cano, I.; Zhou, T.; et al. Xgboost: Extreme Gradient Boosting. R Package Version 1.2.0.1. 2020. Available online: https://CRAN.R-project.org/package=xgboost (accessed on 5 August 2020).
- Kuhn, M. Caret: Classification and Regression Training. R Package Version 6.0-86. 2020. Available online: https://CRAN.R-project.org/package=caret (accessed on 20 July 2020).
- Hijmans, R.J. raster: Geographic Data Analysis and Modeling. R Package Version 3.3-13. 2020. Available online: https://CRAN.R-project.org/package=raster (accessed on 20 July 2020).
- De la Giroday, H.-M.C.; Carroll, A.L.; Aukema, B.H. Breach of the northern Rocky Mountain geoclimatic barrier: Initiation of range expansion by the mountain pine beetle. J. Biogeogr. 2012, 39, 1112–1123. [Google Scholar] [CrossRef]
- Haynes, K.J.; Allstadt, A.; Klimetzek, D. Forest defoliator outbreaks under climate change: Effects on the frequency and severity of outbreaks of five pine insect pests. Glob. Change Biol. 2014, 20, 2004–2018. [Google Scholar] [CrossRef] [PubMed]
- Eklundh, L.; Johansson, T.; Solberg, S. Mapping insect defoliation in Scots pine with MODIS time-series data. Remote Sens. Environ. 2009, 113, 1566–1573. [Google Scholar] [CrossRef]
- MODIS/Terra + Aqua Land Cover Type Yearly L3 Global 0.05Deg CMG. NASA LP DAAC. Available online: https://lpdaac.usgs.gov/products/mcd12c1v006/ (accessed on 20 July 2020).
- Mezentsev, V.S.; Karnatsevich, I.V. Humidification of the West-Siberian Plain; Gidrometeoizdat: Saint Petersburg, Russia, 1969; p. 168. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| File Name | Date |
|---|---|
| LC08_L1TP_141022_20180812_20180815_01_T1 | 25 June 2018 |
| LC08_L1TP_141022_20180929_20181009_01_T1 | 29 September 2018 |
| LC08_L1TP_140022_20190504_20190520_01_T1 | 4 May 2019 |
| LC08_L1TP_141022_20190730_20190801_01_T1 | 30 July 2019 |
| LC08_L1TP_140022_20190909_20190917_01_T1 | 9 September 2019 |
| LC08_L1TP_141022_20200513_20200526_01_T1 | 13 May 2020 |
| Date | Damaged Area, Ha | Classification Algorithm | Accuracy | Sensitivity | Specificity | Kappa |
|---|---|---|---|---|---|---|
| 29 September 2018 | 109.7 | xgboost | 1.000 | 1.000 | 0.886 | 0.918 |
| 30 July 2019 | 3459.6 | xgboost | 0.998 | 0.999 | 0.988 | 0.981 |
| 9 September 2019 | 17603.3 | random forest | 1.000 | 1.000 | 1.000 | 0.999 |
| Date | Area Increase, Ha | Altitude, m | Percentiles | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Average | Max. | Min. | 0.1 | 0.5 | 2.5 | 25 | 50 | 75 | 97.5 | 99.5 | 99.9 | ||
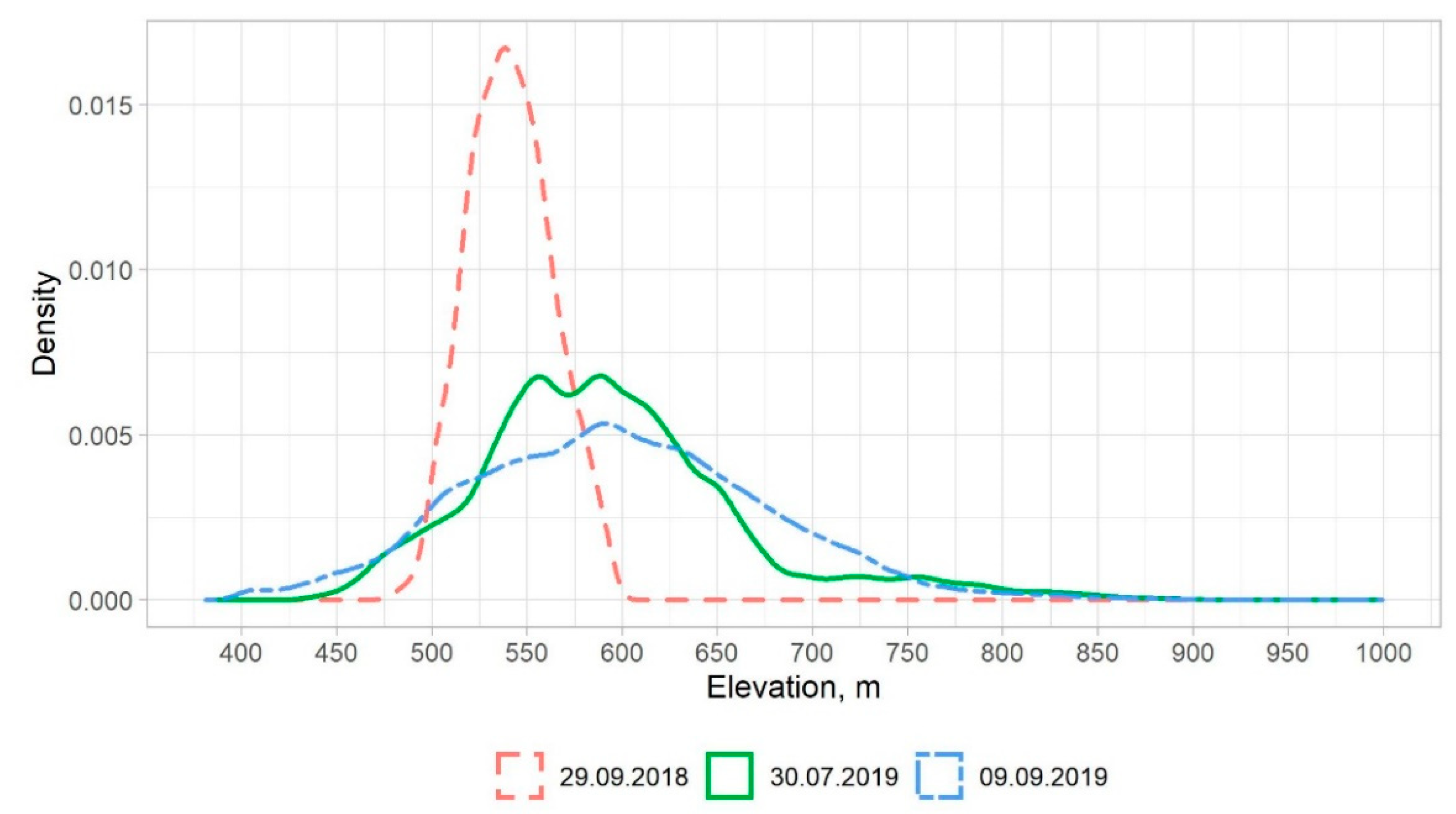
| 29 September 2018 | 109.7 | 541.1 | 596.2 | 478.3 | 481 | 491 | 501 | 525 | 540 | 557 | 586 | 592 | 596 |
| 30 July 2019 | 3349.9 | 593.2 | 999.3 | 429.2 | 441 | 457 | 477 | 547 | 586 | 627 | 779 | 843 | 883 |
| 9 September 2019 | 14253.4 | 597.0 | 997.9 | 381.1 | 400 | 412 | 452 | 541 | 594 | 648 | 757 | 833 | 884 |
| Date | Terrain Slope, Degrees | Percentiles | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Average | Median | Max. | Min. | 0.5 | 2.5 | 25 | 50 | 75 | 97.5 | 99.5 | |
| 29 September 2018 | 10.6 | 10.6 | 27.8 | 0.1 | 1.5 | 2.7 | 7.8 | 10.6 | 13.3 | 19.7 | 24.2 |
| 30 July 2019 | 10.2 | 9.2 | 45.1 | 0.1 | 0.8 | 1.8 | 5.9 | 9.2 | 13.3 | 24.0 | 30.5 |
| 9 September 2019 | 9.9 | 9.0 | 48.8 | 0.0 | 0.8 | 1.7 | 5.8 | 9.0 | 13.0 | 23.0 | 28.7 |
| Date | NW | W | SW | S | SE | E | NE | N |
|---|---|---|---|---|---|---|---|---|
| 29 September 2018 | 0.18 | 0.09 | 0.05 | 0.00 | 0.01 | 0.04 | 0.13 | 0.12 |
| 30 July 2019 | 2.10 | 2.26 | 2.34 | 1.69 | 1.94 | 2.41 | 2.47 | 2.75 |
| 9 September 2019 | 9.18 | 9.42 | 8.42 | 7.42 | 8.66 | 10.58 | 11.22 | 10.28 |
| Forest Type 1 | Aspect and Degree of Slopes | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N | NE | NW | S | SE | SW | E | W | |||||||||
| Up to 10° | ˃10° | Up to 10° | ˃10° | Up to 10° | ˃10° | Up to 10° | ˃10° | Up to 10° | ˃10° | Up to 10° | ˃10° | Up to 10° | ˃10° | Up to 10° | ˃10° | |
| Bil.ced | + 2 | - | + | - | + | - | - | - | - | - | - | - | + | - | + | + |
| Ber.ced | + | +++ | - | + | - | + | ++ | + | + | - | + | + | + | + | + | +++ |
| Bil.fir | ++ | - | - | - | - | - | + | + | ++ | + | + | - | - | - | + | - |
| Mos.fir | - | + | ++ | + | + | + | + | + | - | + | - | + | + | + | + | - |
| Ber.fir | - | + | - | - | - | - | - | + | ++ | + | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sultson, S.M.; Goroshko, A.A.; Verkhovets, S.V.; Mikhaylov, P.V.; Ivanov, V.A.; Demidko, D.A.; Kulakov, S.S. Orographic Factors as a Predictor of the Spread of the Siberian Silk Moth Outbreak in the Mountainous Southern Taiga Forests of Siberia. Land 2021, 10, 115. https://doi.org/10.3390/land10020115
Sultson SM, Goroshko AA, Verkhovets SV, Mikhaylov PV, Ivanov VA, Demidko DA, Kulakov SS. Orographic Factors as a Predictor of the Spread of the Siberian Silk Moth Outbreak in the Mountainous Southern Taiga Forests of Siberia. Land. 2021; 10(2):115. https://doi.org/10.3390/land10020115
Chicago/Turabian StyleSultson, Svetlana M., Andrey A. Goroshko, Sergey V. Verkhovets, Pavel V. Mikhaylov, Valery A. Ivanov, Denis A. Demidko, and Sergey S. Kulakov. 2021. "Orographic Factors as a Predictor of the Spread of the Siberian Silk Moth Outbreak in the Mountainous Southern Taiga Forests of Siberia" Land 10, no. 2: 115. https://doi.org/10.3390/land10020115
APA StyleSultson, S. M., Goroshko, A. A., Verkhovets, S. V., Mikhaylov, P. V., Ivanov, V. A., Demidko, D. A., & Kulakov, S. S. (2021). Orographic Factors as a Predictor of the Spread of the Siberian Silk Moth Outbreak in the Mountainous Southern Taiga Forests of Siberia. Land, 10(2), 115. https://doi.org/10.3390/land10020115