
The Alien Plants That Threaten South Africa’s Mountain Ecosystems

 ,
, 
Abstract
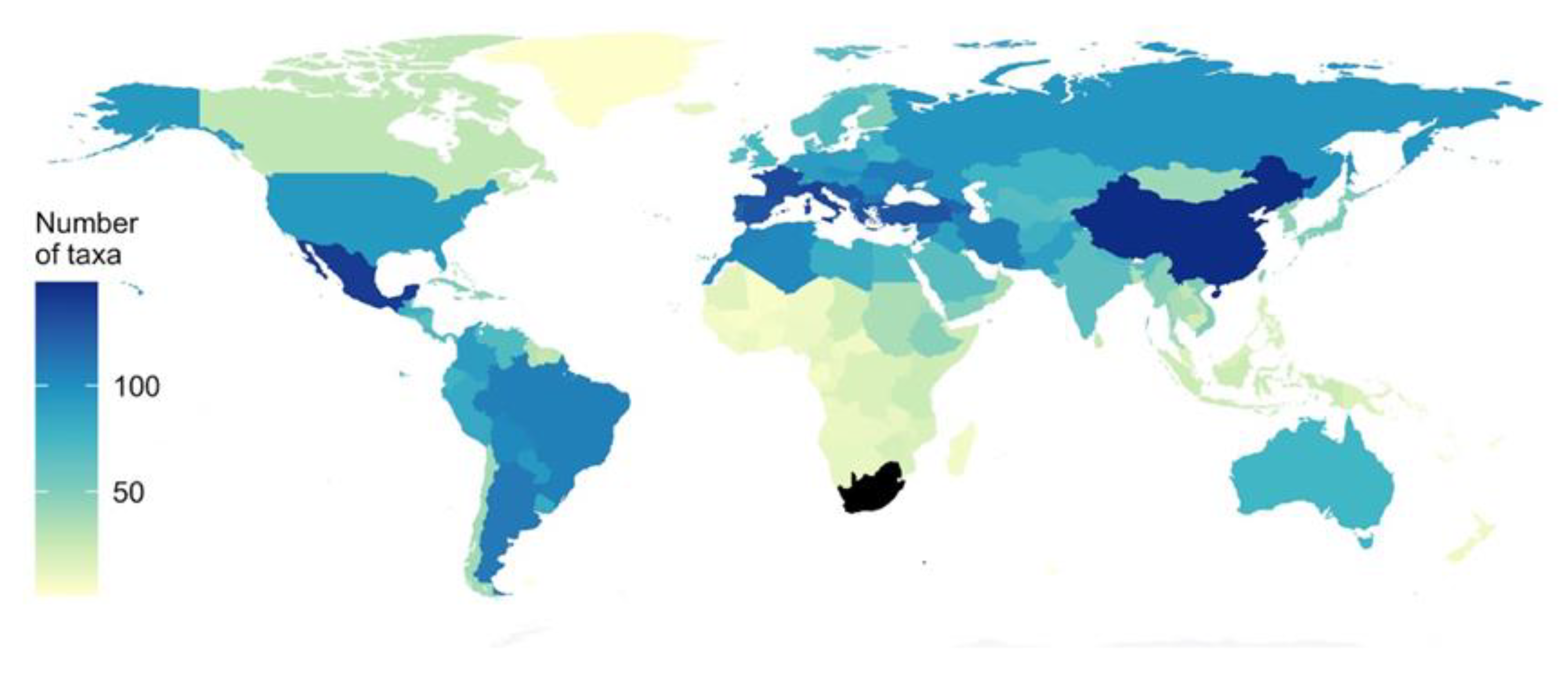
:1. Introduction
2. Data and Methods
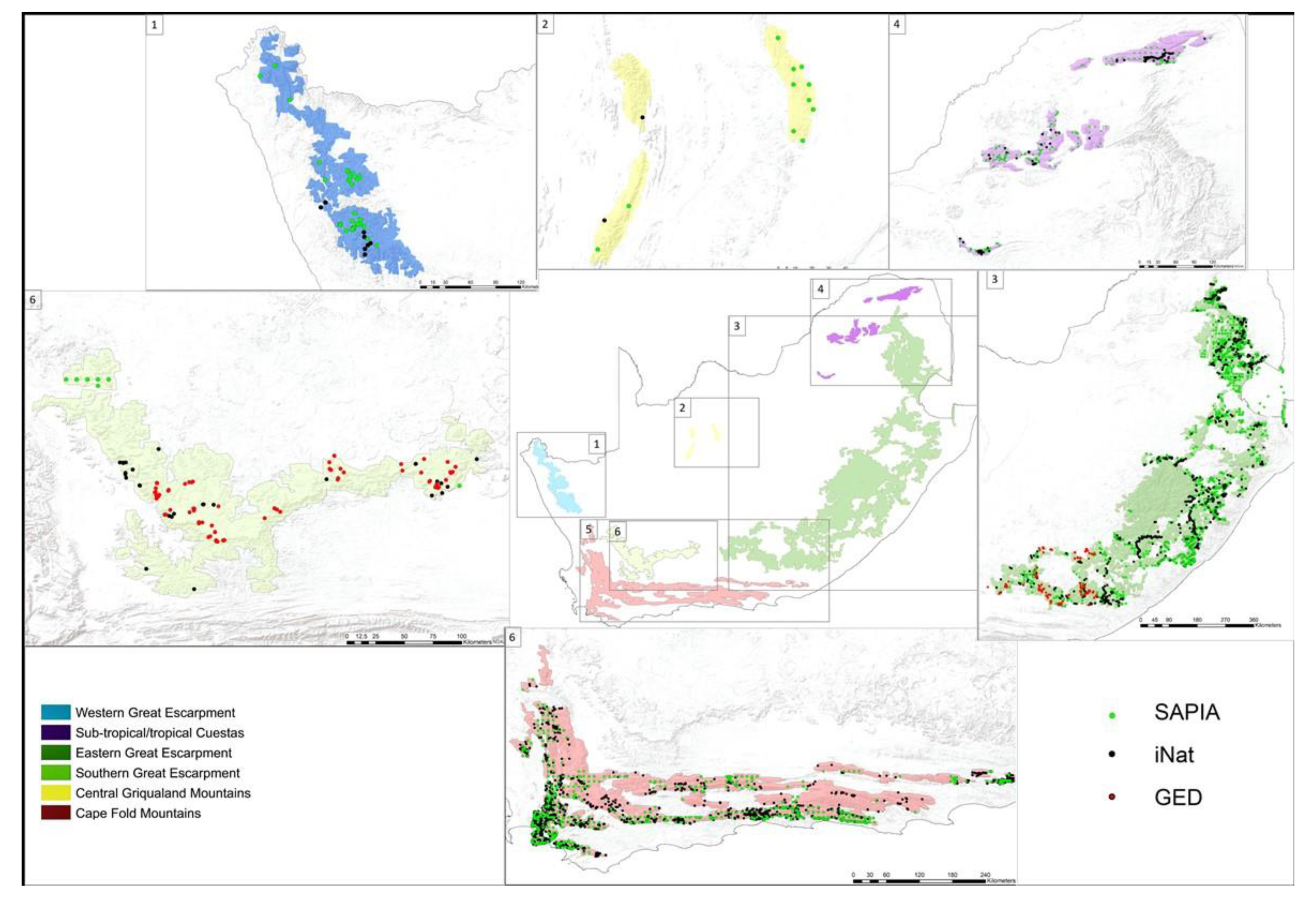
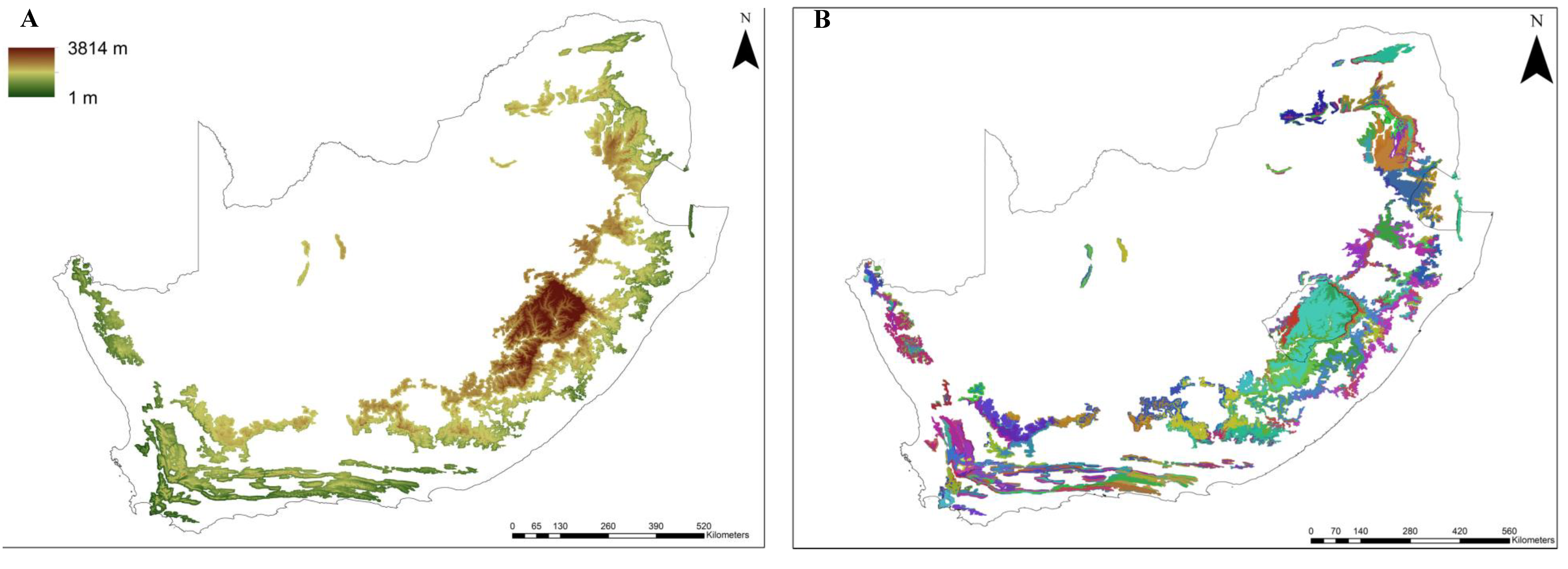
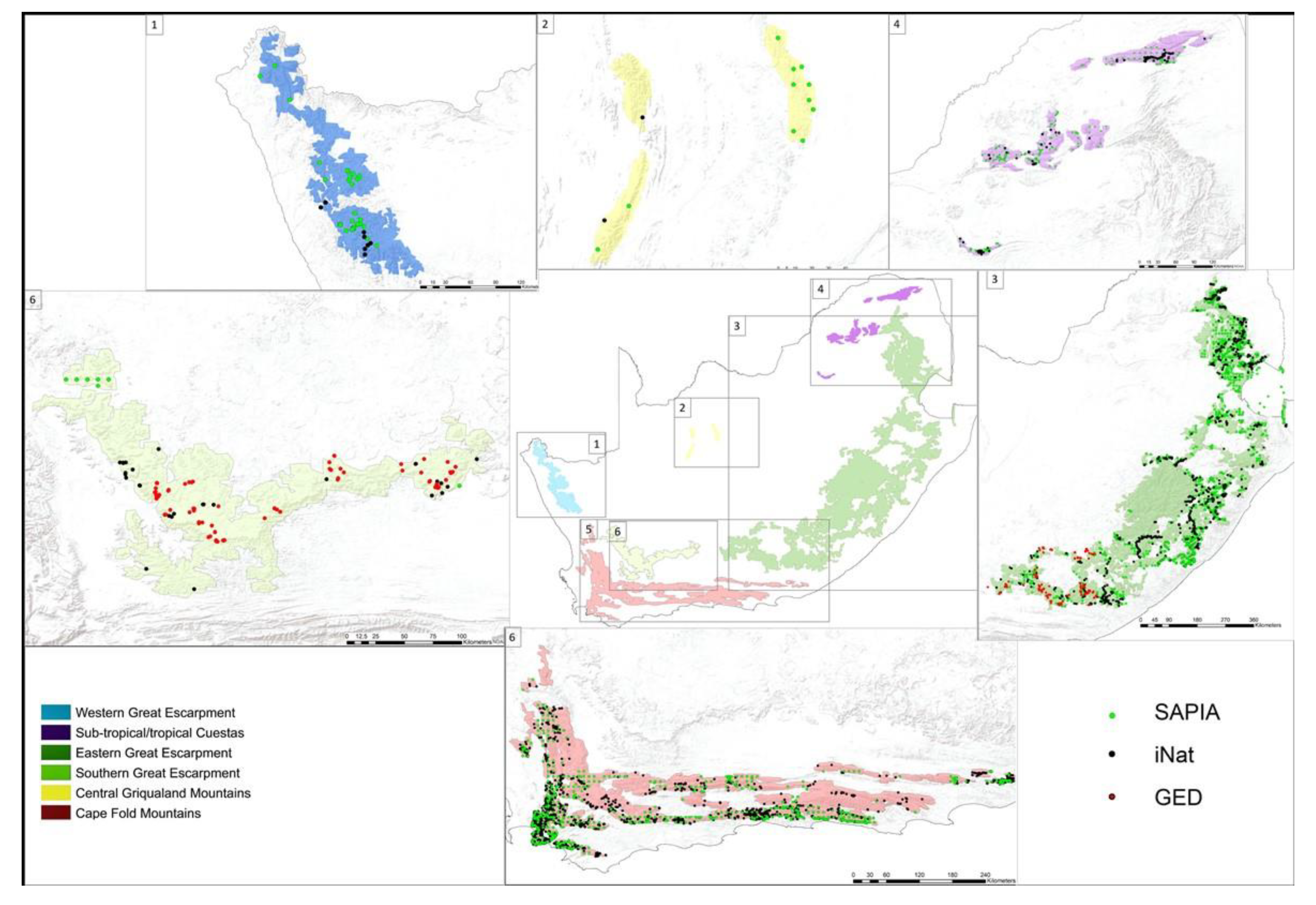
2.1. Delineating Mountain Ranges and Their Characteristics
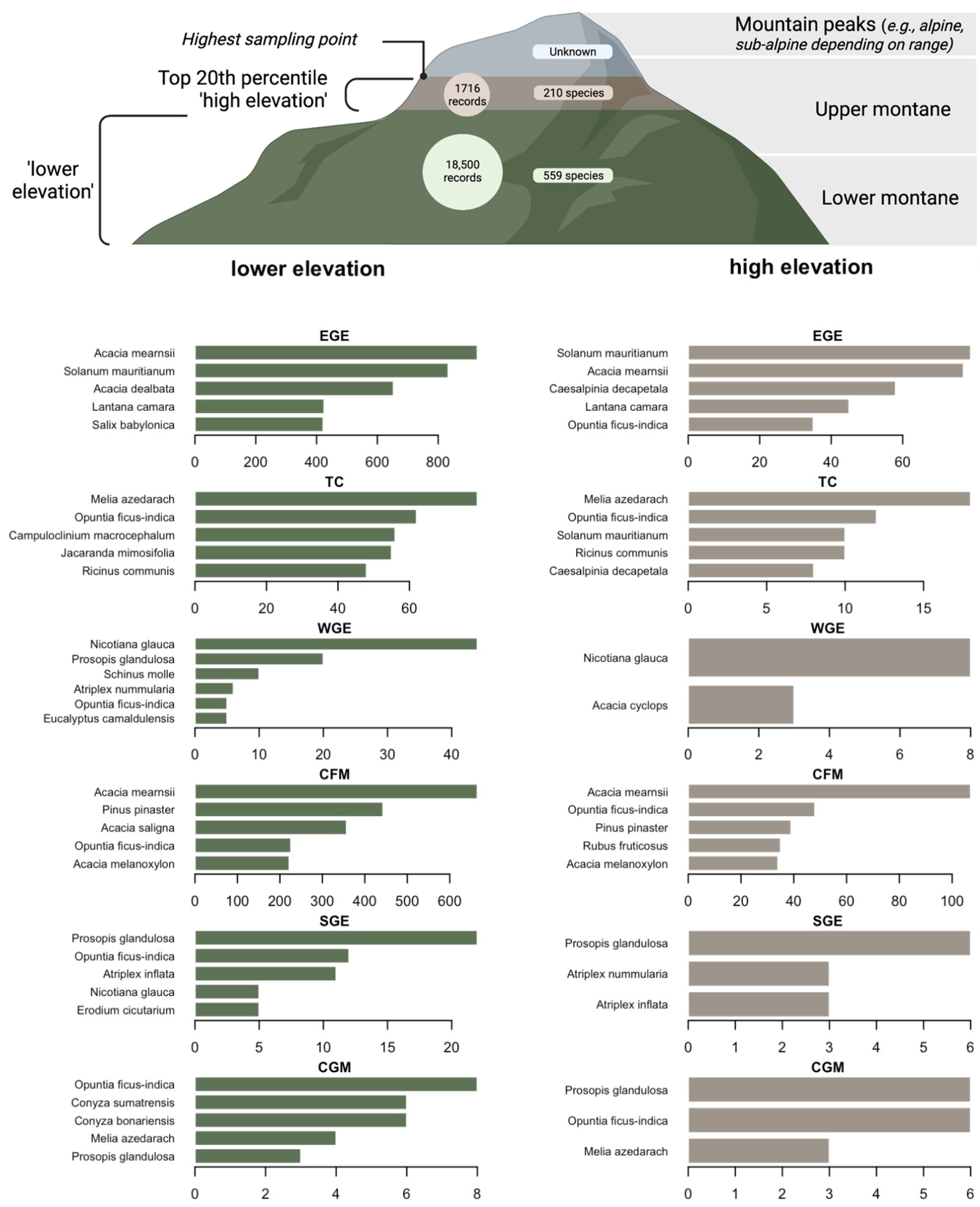
2.2. Sampling Effort
2.3. Species Composition
2.4. Species Abundance
3. Results and Analyses
3.1. Delineating Mountain Ranges And Their Characteristics
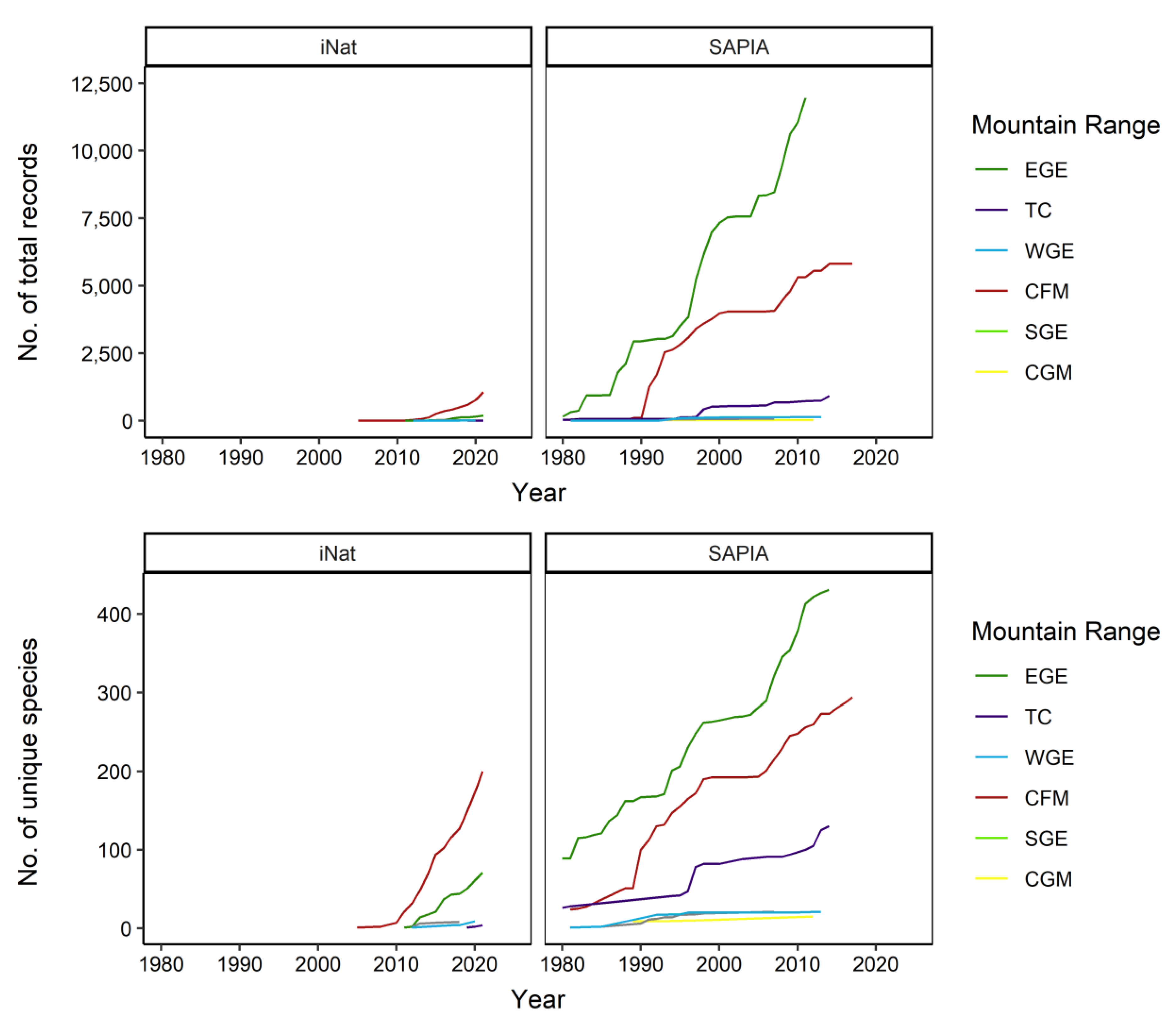
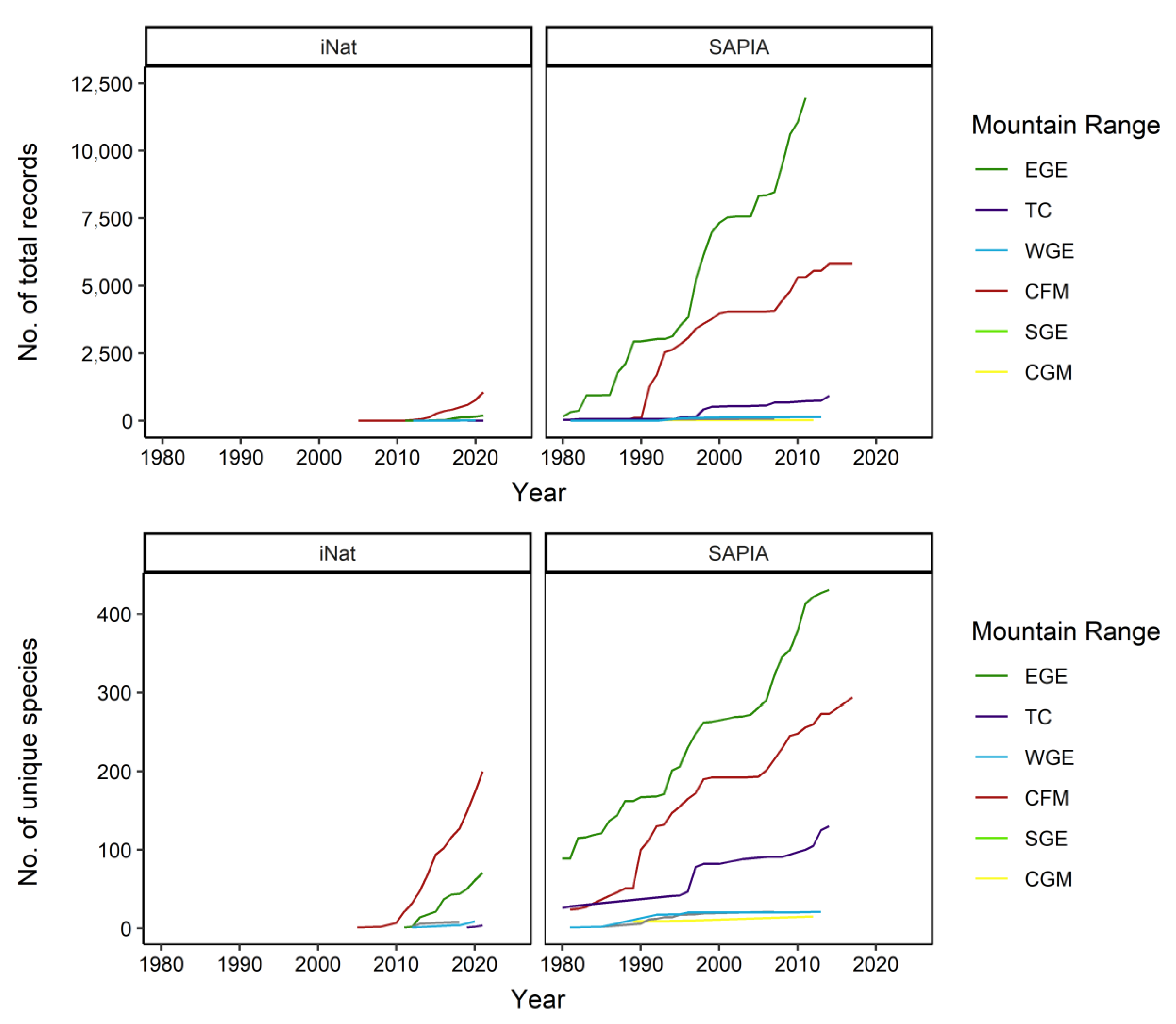
3.2. Sampling Effort
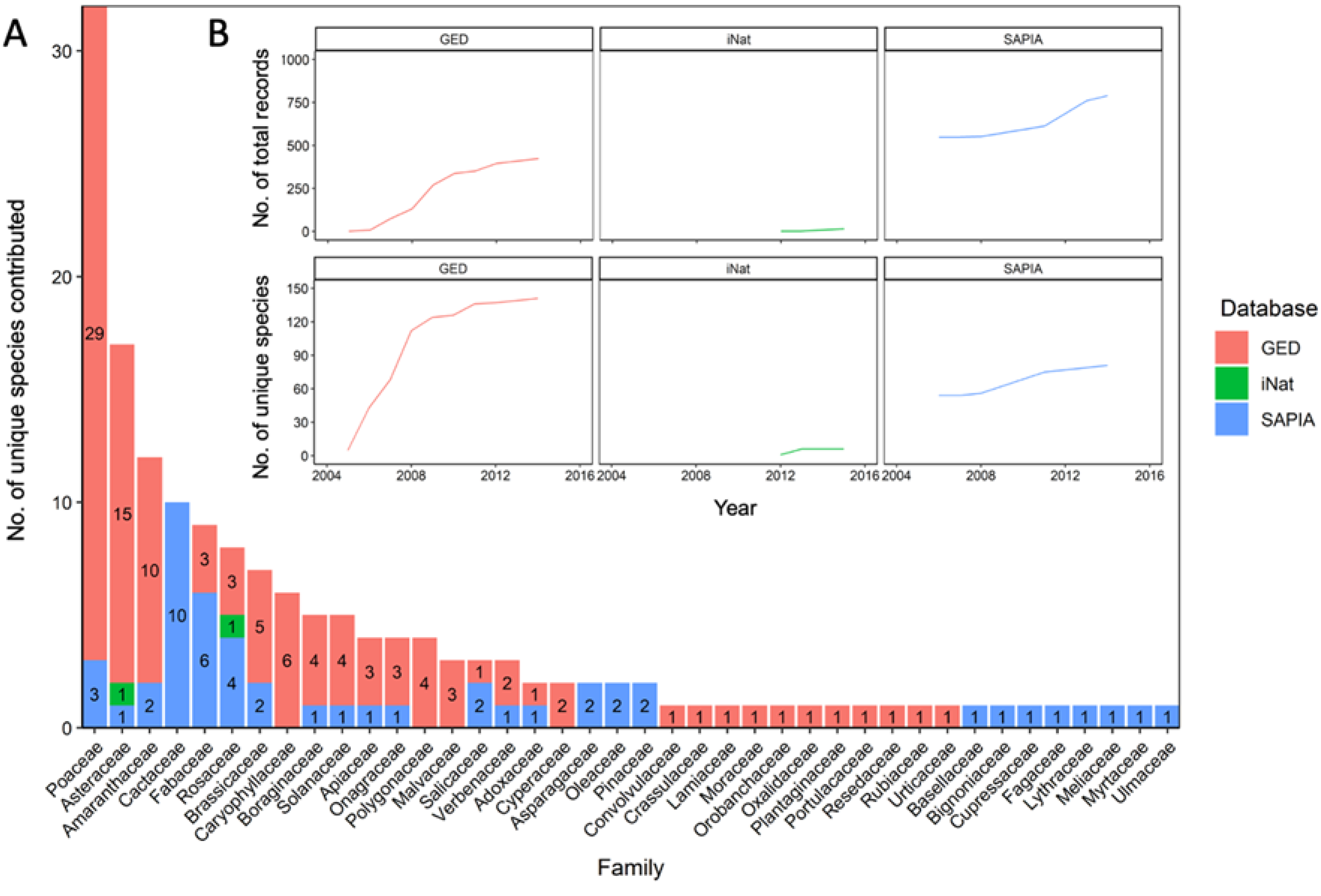
3.3. Species Composition
3.4. Species Abundance within Each Range
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Turpie, J.K.; Marais, C.; Blignaut, J.N. The Working for Water Programme: Evolution of a Payments for Ecosystem Services Mechanism That Addresses Both Poverty and Ecosystem Service Delivery in South Africa. Ecol. Econ. 2008, 65, 788–798. [Google Scholar] [CrossRef] [Green Version]
- Currie, B.; Milton, S.J.; Steenkamp, J. Cost–Benefit Analysis of Alien Vegetation Clearing for Water Yield and Tourism in a Mountain Catchment in the Western Cape of South Africa. Ecol. Econ. 2009, 68, 2574–2579. [Google Scholar] [CrossRef]
- Viviroli, D.; Dürr, H.H.; Messerli, B.; Meybeck, M.; Weingartner, R. Mountains of the World, Water Towers for Humanity: Typology, Mapping, and Global Significance. Water Resour. Res. 2007, 43, 7. [Google Scholar] [CrossRef] [Green Version]
- Matthews, W.; Van Wyk, A.; Bredenkamp, G. Endemic Flora of the North-Eastern Transvaal Escarpment, South Africa. Biol. Conserv. 1993, 63, 83–94. [Google Scholar] [CrossRef]
- Clark, V.R.; Barker, N.P.; Mucina, L. The Great Escarpment of Southern Africa: A New Frontier for Biodiversity Exploration. Biodivers. Conserv. 2011, 20, 2543–2561. [Google Scholar] [CrossRef]
- Moraswi, I.; Bamigboye, S.O.; Tshisikhawe, M.P. Conservation Status and Threats to Vascular Plant Species Endemic to Soutpansberg Mountain Range in Limpopo Province, South Africa. Int. J. Plant Biol. 2019, 10, 7978. [Google Scholar] [CrossRef] [Green Version]
- Godde, P.M.; Price, M.F.; Zimmermann, F.M. Tourism and Development in Mountain Regions: Moving Forward into the New Millennium. Tour. Dev. Mt. Reg. 2000, 1–25. [Google Scholar]
- Mathibela, K.M. An Investigation into Aspects of Medicinal Plant Use by Traditional Healers from Blouberg Mountain, Limpopo Province, South Africa. Ph.D. Thesis, University of Limpopo, Polokwane, Turfloop Campus, Mankweng, South Africa, 2013. [Google Scholar]
- Adelabu, D.B.; Clark, V.R.; Bredenhand, E. Potential for Sustainable Mountain Farming. Mt. Res. Dev. 2020, 40, 1–11. [Google Scholar]
- Everson, T.M.; Morris, C.D. Conservation of Biodiversity in the Maloti–Drakensberg Mountain Range. In Land Use Change and Mountain Biodiversity; CRC Press: Boca Raton, FL, USA, 2006; pp. 285–292. ISBN 0-429-12521-6. [Google Scholar]
- McDougall, K.L.; Alexander, J.M.; Haider, S.; Pauchard, A.; Walsh, N.G.; Kueffer, C. Alien Flora of Mountains: Global Comparisons for the Development of Local Preventive Measures against Plant Invasions. Divers. Distrib. 2011, 17, 103–111. [Google Scholar] [CrossRef]
- Mukwada, G.; Manatsa, D. Spatiotemporal Analysis of the Effect of Climate Change on Vegetation Health in the Drakensberg Mountain Region of South Africa. Environ. Monit. Assess. 2018, 190, 1–21. [Google Scholar] [CrossRef]
- Canavan, K.; Canavan, S.; Clark, V.R.; Gwate, O.; Mapaura, A.; Richardson, D.M.; Steenhuisen, S.; Martin, G.D. Invasive Alien Plants in the Montane Areas of South Africa: Impacts and Management Options. In Exploring the Multiple Values of Nature: Connecting Ecosystems and People Across Landscapes; Springer: Cham, Switzerland, 2022; in press. [Google Scholar]
- Tecco, P.A.; Pais-Bosch, A.I.; Funes, G.; Marcora, P.I.; Zeballos, S.R.; Cabido, M.; Urcelay, C. Mountain Invasions on the Way: Are There Climatic Constraints for the Expansion of Alien Woody Species along an Elevation Gradient in Argentina? J. Plant Ecol. 2016, 9, 380–392. [Google Scholar] [CrossRef] [Green Version]
- Turner, S.C. Examining the Spatiotemporal Dynamics of Exotic Plants in a Montane Grassland: Mechanisms and Management. Master’s Thesis, Stellenbosch University, Stellenbosch, South Africa, 2019. [Google Scholar]
- Pauchard, A.; Kueffer, C.; Dietz, H.; Daehler, C.C.; Alexander, J.; Edwards, P.J.; Arévalo, J.R.; Cavieres, L.A.; Guisan, A.; Haider, S.; et al. Ain’t No Mountain High Enough: Plant Invasions Reaching New Elevations. Front. Ecol. Environ. 2009, 7, 479–486. [Google Scholar] [CrossRef] [Green Version]
- Alexander, J.M.; Lembrechts, J.J.; Cavieres, L.A.; Daehler, C.; Haider, S.; Kueffer, C.; Liu, G.; McDougall, K.; Milbau, A.; Pauchard, A.; et al. Plant Invasions into Mountains and Alpine Ecosystems: Current Status and Future Challenges. Alp. Bot. 2016, 126, 89–103. [Google Scholar] [CrossRef]
- Arévalo, J.R.; Delgado, J.D.; Otto, R.; Naranjo, A.; Salas, M.; Fernández-Palacios, J.M. Distribution of Alien vs. Native Plant Species in Roadside Communities along an Altitudinal Gradient in Tenerife and Gran Canaria (Canary Islands). Perspect. Plant Ecol. Evol. Syst. 2005, 7, 185–202. [Google Scholar] [CrossRef]
- Lembrechts, J.J.; Alexander, J.M.; Cavieres, L.A.; Haider, S.; Lenoir, J.; Kueffer, C.; McDougall, K.; Naylor, B.J.; Nuñez, M.A.; Pauchard, A.; et al. Mountain Roads Shift Native and Non-native Plant Species’ Ranges. Ecography 2017, 40, 353–364. [Google Scholar]
- Moradi, H.; Fattorini, S.; Oldeland, J. Influence of Elevation on the Species–Area Relationship. J. Biogeogr. 2020, 47, 2029–2041. [Google Scholar] [CrossRef]
- Alexander, M.J.; Teitelbaum, H. Three dimensional Properties of Andes Mountain Waves Observed by Satellite: A Case Study. J. Geophys. Res. Atmos. 2011, 116. [Google Scholar] [CrossRef] [Green Version]
- Marini, L.; Battisti, A.; Bona, E.; Federici, G.; Martini, F.; Pautasso, M.; Hulme, P.E. Alien and Native Plant Life-forms Respond Differently to Human and Climate Pressures. Glob. Ecol. Biogeogr. 2012, 21, 534–544. [Google Scholar] [CrossRef]
- Haider, S.; Kueffer, C.; Bruelheide, H.; Seipel, T.; Alexander, J.M.; Rew, L.J.; Arévalo, J.R.; Cavieres, L.A.; McDougall, K.L.; Milbau, A.; et al. Mountain Roads and Non-native Species Modify Elevational Patterns of Plant Diversity. Glob. Ecol. Biogeogr. 2018, 27, 667–678. [Google Scholar] [CrossRef]
- Wilson, J.R.; Foxcroft, L.C.; Geerts, S.; Hoffman, M.T.; MacFadyen, S.; Measey, J.; Mills, A.; Richardson, D.M.; Robertson, M.P.; van Wilgen, B.W. The Role of Environmental Factors in Promoting and Limiting Biological Invasions in South Africa. In Biological Invasions in South Africa; Springer: Cham, Switzerland, 2020; pp. 355–385. [Google Scholar]
- Marini, L.; Bertolli, A.; Bona, E.; Federici, G.; Martini, F.; Prosser, F.; Bommarco, R. Beta-diversity Patterns Elucidate Mechanisms of Alien Plant Invasion in Mountains. Glob. Ecol. Biogeogr. 2013, 22, 450–460. [Google Scholar] [CrossRef]
- Carbutt, C.; Thompson, D.I. Mountain Watch: How LT (S) ER Is Safeguarding Southern Africa’s People and Biodiversity for a Sustainable Mountain Future. Land 2021, 10, 1024. [Google Scholar] [CrossRef]
- Carbutt, C.; Edwards, T. The Flora of the Drakensberg Alpine Centre. Edinb. J. Bot. 2004, 60, 581–607. [Google Scholar] [CrossRef]
- Carbutt, C. The Emerging Invasive Alien Plants of the Drakensberg Alpine Centre, Southern Africa. Bothalia 2012, 42, 71–85. [Google Scholar] [CrossRef] [Green Version]
- Kalwij, J.M.; Robertson, M.P.; van Rensburg, B.J. Human Activity Facilitates Altitudinal Expansion of Exotic Plants along a Road in Montane Grassland, South Africa. Appl. Veg. Sci. 2008, 11, 491–498. [Google Scholar] [CrossRef] [Green Version]
- Shepard, M.K.; Campbell, B.A.; Bulmer, M.H.; Farr, T.G.; Gaddis, L.R.; Plaut, J.J. The roughness of natural terrain: A planetary and remote sensing perspective. J. Geophys. Res. Planets 2001, 106, 32777–32795. [Google Scholar] [CrossRef]
- Schulze, R.E. South African Atlas of Agrohydrology and Climatology. In Contribution towards a Final Report to the Water Research Commission on Project 492: Modelling Impacts of the Agricultural Environment on Water Resources: TT82-96; Water Research Commission (WRF): Pretoria, South Africa, 1997. [Google Scholar]
- Rutherford, M.C.; Mucina, L.; Powrie, L.W. Biomes and Bioregions of Southern Africa. In The Vegetation of South Africa, Lesotho and Swaziland; Phillips, K., Powrie, L.W., Eds.; Strelitzia, South African National Biodiversity Institute: Pretoria, South Africa, 2006; Volume 19, pp. 30–51. [Google Scholar]
- Google Scholar. Available online: https://scholar.google.com/ (accessed on 29 September 2021).
- Scopus. Available online: https://www.scopus.com/ (accessed on 29 September 2021).
- ScienceDirect. Available online: https://www.sciencedirect.com/ (accessed on 24 September 2021).
- Web of Science. Available online: https://www.webofknowledge.com/ (accessed on 23 September 2021).
- Taylor and Francis. Available online: https://www.tandfonline.com/ (accessed on 21 September 2021).
- SAPIA. Atlas 40 Years of SAPIA. 2014. Available online: https://www.arc.agric.za/ARC%20Newsletters/SAPIA%20NEWS%20No.%2055,%20January%202020.pdf (accessed on 10 August 2021).
- iNaturalist. Available online: https://www.inaturalist.org/ (accessed on 24 August 2021).
- Barker, N.; Clark, V.; Dold, T. FBIP: Great Escarpment Biodiversity Research Programme Data Mobilisation; South African National Biodiversity Institute. 2020. Available online: https://www.gbif.org/dataset/f8cd9c4d-4693-40d7-9c0c-a3ce2ea2a1c3/project (accessed on 9 August 2021).
- Richardson, D.M.; Rouget, M.; Ralston, S.J.; Cowling, R.M.; Van Rensburg, B.J.; Thuiller, W. Species Richness of Alien Plants in South Africa: Environmental Correlates and the Relationship with Native Plant Species Richness. Ecoscience 2005, 12, 391–402. [Google Scholar] [CrossRef]
- Aristeidou, M.; Herodotou, C.; Ballard, H.L.; Young, A.N.; Miller, A.E.; Higgins, L.; Johnson, R.F. Exploring the Participation of Young Citizen Scientists in Scientific Research: The Case of iNaturalist. PLoS ONE 2021, 16, e0245682. [Google Scholar] [CrossRef] [PubMed]
- Nugent, J. iNaturalist. Sci. Scope 2018, 41, 12–13. [Google Scholar] [CrossRef]
- Laider, G. Using iNaturalist as a New Model for Recording Species for CREW Field Trips. CREW Newsl. 2019, 15, 46–47. [Google Scholar]
- POWO. Plants of the World Online. Available online: http://www.plantsoftheworldonline.org/ (accessed on 30 September 2021).
- The Plant List. Available online: http://www.theplantlist.org/ (accessed on 30 September 2021).
- Brownrigg, R. Package ‘maps’, Version 3.4.0. 2021. Available online: https://cran.r-project.org/web/packages/maps/maps.pdf (accessed on 1 October 2021).
- Rouget, M.; Hui, C.; Renteria, J.; Richardson, D.M.; Wilson, J.R.U. Plant Invasions as a Biogeographical Assay: Vegetation Biomes Constrain the Distribution of Invasive Alien Species Assemblages. S. Afr. J. Bot. 2015, 101, 24–31. [Google Scholar]
- Hui, F.K.C. Boral-Bayesian ordination and regression analysis of multivariate abundance data. R. Meth. Ecol. Evol. 2016, 7, 744–750. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Naumann, U.; Wright, S.T.; Warton, D.I. Mvabund—An R Package for Model-based Analysis of Multivariate Abundance Data. Meth. Ecol. Evol. 2012, 3, 471–474. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing (Version 4.0.3); R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Carbutt, C. The Imperiled Alpine Grasslands of the Afrotropic Realm. In Earth Systems and Environmental Sciences; Elsevier: Amsterdam, The Netherlands, 2020. [Google Scholar] [CrossRef]
- Weihrauch, D.M. Alpine Vegetation Communities. In Proceedings of the Wilderness Science in a Time of Change Conference, Missoula, Montana, 23–27 May 1999; US Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2000; Volume 3, p. 93. [Google Scholar]
- Jury, M.R. An inter-comparison of model-simulated east-west climate gradients over South Africa. Water SA 2012, 38, 467–478. [Google Scholar] [CrossRef] [Green Version]
- Henderson, L.; Wilson, J.R.U. Changes in the composition and distribution of alien plants in South Africa: An update from the South African Plant Invaders Atlas. Bothalia 2017, 47, a2172. [Google Scholar] [CrossRef]
- Ruwanza, S.; Mhlongo, E.S. Lantana Camara Invasion along Road-River Interchanges and Roadsides in Soutpansberg, Vhembe Biosphere Reserve in South Africa. S. Afr. J. Sci. 2020, 116, 1–5. [Google Scholar] [CrossRef]
- Clark, V.; Barker, N.; Mucina, L. The Sneeuberg: A New Centre of Floristic Endemism on the Great Escarpment, South Africa. S. Afr. J. Bot. 2009, 75, 196–238. [Google Scholar] [CrossRef]
- Clark, V.; Barker, N.; McMaster, C.; Mucina, L. The Boschberg (Somerset East, Eastern Cape)—A Floristic Cross-Roads of the Southern Great Escarpment. S. Afr. J. Bot. 2011, 77, 94–104. [Google Scholar] [CrossRef] [Green Version]
- Clark, V.R.; Perera, S.J.; Stiller, M.; Stirton, C.H.; Weston, P.H.; Stoev, P.; Coombs, G.; Morris, D.B.; Ratnayake-Perera, D.; Barker, N.P. A Rapid Multi-Disciplinary Biodiversity Assessment of the Kamdebooberge (Sneeuberg, Eastern Cape, South Africa): Implications for Conservation. SpringerPlus 2012, 1, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, L.; Bezuidenhout, H. The Vegetation of the Farms Ingleside and Welgedacht of the Mountain Zebra National Park, Eastern Cape. Koedoe 2005, 48, 23–42. [Google Scholar] [CrossRef] [Green Version]
- Bezuidenhout, H.; Brown, L.R. Vegetation Description of the Doornhoek Section of the Mountain Zebra National Park (MZNP), South Africa. Koedoe: Afr. Prot. Area Conserv. Sci. 2008, 50, 82–92. [Google Scholar] [CrossRef]
- Brown, L.; Bezuidenhout, H. Ecosystem Description and Diversity of the Jurisdam–Seekoegat Sections of the Mountain Zebra National Park, South Africa. S. Afr. J. Bot. 2018, 118, 166–178. [Google Scholar] [CrossRef]
- Steyn, C.; Greve, M.; Robertson, M.P.; Kalwij, J.M.; le Roux, P.C. Alien Plant Species That Invade High Elevations Are Generalists: Support for the Directional Ecological Filtering Hypothesis. J. Veg. Sci. 2017, 28, 337–346. [Google Scholar] [CrossRef]
- Popp, M.R.; Kalwij, J.M. Abiotic Conditions Shape the Relationship between Indigenous and Exotic Species Richness in a Montane Biodiversity Hotspot. Plant Ecol. 2021, 222, 421–432. [Google Scholar] [CrossRef]
- Brand, R.F.; Du Preez, P.J.; Brown, L.R. A Floristic Description of the Afromontane Fynbos Communities on Platberg, Eastern Free State, South Africa. Koedoe: Afr. Prot. Area Conserv. Sci. 2008, 50, 202–213. [Google Scholar] [CrossRef] [Green Version]
- Brand, R.F.; Brown, L.R.; Du Preez, P.J. The Grassland Vegetation of Platberg, Eastern Free State, South Africa. Koedoe: Afr. Prot. Area Conserv. Sci. 2011, 53, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Grieve, G.R.; Downs, C.T. A Checklist of the Plants of the Forests and Grasslands in the Weza District, Southern KwaZulu-Natal and a Review of Their Status in the Red Data List: Checklist. Koedoe: Afr. Prot. Area Conserv. Sci. 2015, 57, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Clark, V.; Dold, A.; McMaster, C.; McGregor, G.; Bredenkamp, C.; Barker, N. Rich Sister, Poor Cousin: Plant Diversity and Endemism in the Great Winterberg–Amatholes (Great Escarpment, Eastern Cape, South Africa). S. Afr. J. Bot. 2014, 92, 159–174. [Google Scholar] [CrossRef] [Green Version]
- Omar, M.Y.; Maroyi, A.; Van Tol, J.J. Floral Diversity, Composition and Distribution in a Montane Wetland in Hogsback, the Eastern Cape Province, South Africa. Pak. J. Bot. 2016, 48, 1861–1870. [Google Scholar]
- Hoffman, M.T.; Van der Merwe, H. Vegetation of Akkerendam Nature Reserve, Northern Cape: Delineation and Dynamics over 100 Years. Bothalia-Afr. Biodiv. Conserv. 2019, 49, 1–9. [Google Scholar]
- Clark, V.; Barker, N.; Mucina, L. The Roggeveldberge—Notes on a Botanically Hot Area on a Cold Corner of the Southern Great Escarpment, South Africa. S. Afr. J. Bot. 2011, 77, 112–126. [Google Scholar] [CrossRef] [Green Version]
- Clark, V.; Barker, N.; Mucina, L. A Phytogeographic Assessment of the Nuweveldberge, South Africa. S. Afr. J. Bot. 2011, 77, 147–159. [Google Scholar] [CrossRef] [Green Version]
- Moll, E.; Trinder-Smith, T. Invasion and Control of Alien Woody Plants on the Cape Peninsula Mountains, South Africa—30 Years On. Biol. Conserv. 1992, 60, 135–143. [Google Scholar] [CrossRef]
- Walton, B.A. Vegetation patterns and dynamics of Renosterveld at Ager-Groeneberg conservancy, Western Cape, South Africa. Master’s Thesis, Stellenbosch University, Stellenbosch, South Africa, 2006. [Google Scholar]
- Kraaij, T.; Baard, J.A. Use of a Rapid Roadside Survey to Detect Potentially Invasive Plant Species along the Garden Route, South Africa. Koedoe: Afr. Prot. Area Conserv. Sci. 2019, 61, 1–10. [Google Scholar]
- Invasive Species Compendium. Centre for Agriculture and Bioscience International datasheets. Available online: https://www.cabi.org/isc/ (accessed on 10 October 2021).
- Department of Environment Australia. Atriplex lindleyi subsp inflata. Available online: https://www.water.wa.gov.au/__data/assets/pdf_file/0014/5423/84797.pdf (accessed on 10 October 2021).
- Environmental Affairs. State of the Environment: Chapter 3. 2018. Available online: https://www.environment.gov.za/sites/default/files/docs/SKAIEMPChapter3.pdf (accessed on 20 October 2021).
- Abu-Zanat, M.; Samarah, N. Physical and Chemical Treatments for Enhancing Seed Germination of Oldman Saltbush (Atriplex nummularia). Afr. J. Range Forage Sci. 2005, 22, 141–145. [Google Scholar] [CrossRef]
- Gitonga, L.; Cron, G.; McConnachie, A.; Byrne, M. Genetic Variation of the Invasive Campuloclinium macrocephalum, Asteraceae in South Africa, Inferred from Molecular Markers. Weed Res. 2015, 55, 51–61. [Google Scholar] [CrossRef]
- Goodall, J.; Witkowski, E.; McConnachie, A.J.; Keen, C. Altered Growth, Population Structure and Realised Niche of the Weed Campuloclinium macrocephalum (Asteraceae) after Exposure to the Naturalised Rust Puccinia eupatorii (Pucciniaceae). Biol. Inv. 2012, 14, 1947–1962. [Google Scholar] [CrossRef]
- Latch, B. Weeping Willow Rust in New Zealand. J. Agric. Res. 1980, 23, 535–538. [Google Scholar] [CrossRef]
- Wangdi, K.; Roder, W. Willow (Salix babylonica) Fodder Tree for the Temperate: An Experience from Bhutan. 2005. Available online: http://citeseerx.ist.psu.edu/viewdoc/download?doi=10.1.1.485.1&rep=rep1&type=pdf (accessed on 20 October 2021).
- Encarnação, J.; Teodósio, M.A.; Morais, P. Citizen Science and Biological Invasions: A Review. Front. Environ. Sci. 2021, 8, 303. [Google Scholar] [CrossRef]
- Clark, V.R. First quantification of plant endemics in the Manica Highlands (Zimbabwe-Mozambique), and circumscription of the Manica Highlands Centre of Floristic Endemism. In Proceedings of the Conference 57th Annual Grassland Society of Southern Africa Congress, Virtual Event, 26–30 July 2021. [Google Scholar]
- Chari, L.D.; Martin, G.D.; Steenhuisen, S.; Adams, L.D.; Clark, V.R. Biology of Invasive Plants 1. Pyracantha angustifolia (Franch.) CK Schneid. Invasive Plant Sci. Manag. 2020, 13, 120–142. [Google Scholar] [CrossRef]
- Tsunamoto, Y.; Koike, S.; Tayasu, I.; Masaki, T.; Kato, S.; Kikuchi, S.; Nagamitsu, T.; Haraguchi, T.; Naganuma, T.; Naoe, S. Timing of fruiting rather than topography determines the direction of vertical seed dispersal by mammals and birds. bioRxiv 2020. [Google Scholar] [CrossRef]
- McDougall, K.L.; Khuroo, A.A.; Loope, L.L.; Parks, C.G.; Pauchard, A.; Reshi, Z.A.; Rushworth, I.; Kueffer, C. Plant Invasions in Mountains: Global Lessons for Better Management. Mt. Res. Dev. 2011, 31, 380–387. [Google Scholar] [CrossRef]
- Becker, T.; Dietz, H.; Billeter, R.; Buschmann, H.; Edwards, P.J. Altitudinal Distribution of Alien Plant Species in the Swiss Alps. Perspect. Plant Ecol. Evol. Syst. 2005, 7, 173–183. [Google Scholar] [CrossRef]
- Boshoff, W.H.; Fourie, J. The Significance of the Cape Trade Route to Economic Activity in the Cape Colony: A Medium-Term Business Cycle Analysis. Eur. Rev. Econ. Hist. 2010, 14, 469–503. [Google Scholar] [CrossRef] [Green Version]
- Ellis, B. The Impact of the White Settlers on the Natural Environment of Natal, 1845–1870. Ph.D. Thesis, University of KwaZulu-Natal, Pietermaritzburg, South Africa, 1998. [Google Scholar]
- Venter, Z.; Cramer, M.; Hawkins, H.-J. Drivers of Woody Plant Encroachment over Africa. Nature Commu. 2018, 9, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Sylvester, S.P.; Soreng, R.J.; Sylvester, M.D.; Mapaura, A.; Clark, V.R. New Records of Alien and Potentially Invasive Grass (Poaceae) Species for Southern Africa. Bothalia-Afr. Biodiv. Conserv. 2021, 51, 1–9. [Google Scholar] [CrossRef]
- Mapaura, A.; Canavan, K.; Richardson, D.M.; Clark, V.R.; Steenhuisen, S.-L. The Invasive Grass Genus Nassella in South Africa: A Synthesis. S. Afr. J. Bot. 2020, 135, 336–348. [Google Scholar] [CrossRef]
- O’Connor, T.G.; van Wilgen, B.W. The Impact of Invasive Alien Plants on Rangelands in South Africa. In Biological Invasions in South Africa; Springer: Cham, Switzerland, 2020; pp. 459–488. [Google Scholar]
- Grime, J. Benefits of Plant Diversity to Ecosystems: Immediate, Filter and Founder Effects. J. Ecol. 1998, 86, 902–910. [Google Scholar] [CrossRef]
- Le Maitre, D.C.; Blignaut, J.N.; Clulow, A.; Dzikiti, S.; Everson, C.S.; Görgens, A.H.; Gush, M.B. Impacts of Plant Invasions on Terrestrial Water Flows in South Africa. In Biological Invasions in South Africa; Springer: Cham, Switzerland, 2020; pp. 431–457. [Google Scholar]
- Roura-Pascual, N.; Richardson, D.M.; Krug, R.M.; Brown, A.; Chapman, R.A.; Forsyth, G.G.; Le Maitre, D.C.; Robertson, M.P.; Stafford, L.; Van Wilgen, B.W.; et al. Ecology and Management of Alien Plant Invasions in South African Fynbos: Accommodating Key Complexities in Objective Decision Making. Biol. Conserv. 2009, 142, 1595–1604. [Google Scholar] [CrossRef] [Green Version]
- Marais, C.; van Wilgen, B.W.; Stevens, D. The clearing of invasive alien plants in South Africa, a preliminary assessment of costs and progress, working for water. S. Afr. J. Sci. 2004, 100, 97–103. [Google Scholar]
- Martin, G. Prospects for the biological control of Northern Temperate Weeds in South Africa. Afr. Entomol. 2021, in press. [Google Scholar]
- Hobbs, R.J. The Working for Water Programme in South Africa: The Science behind the Success. Divers. Distrib. 2004, 10, 501–503. [Google Scholar] [CrossRef]
- Venter, C. High Altitude Teams Caring for the Environment. FarmBiz 2019, 5, 10–11. [Google Scholar]
- Downey, P.O.; Williams, M.C.; Whiffen, L.K.; Auld, B.A.; Hamilton, M.A.; Burley, A.L.; Turner, P.J. Managing Alien Plants for Biodiversity Outcomes—The Need for Triage. Invasive Plant Sci. Manag. 2010, 3, 1–11. [Google Scholar] [CrossRef]
- Clark, V.; Mukwada, G.; Hansen, M.; Adelabu, S.; Magaiza, G.; le Roux, A.; Bredenhand, E.; Voua Otoma, P.; Steenhuisen, S.-L.; Franke, A.; et al. The Afromontane Research Unit: Driving Connections and Capacity Building for the Sustainable Development of Southern African Mountains. Mt. Res. Dev. 2021, 41, 1–5. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
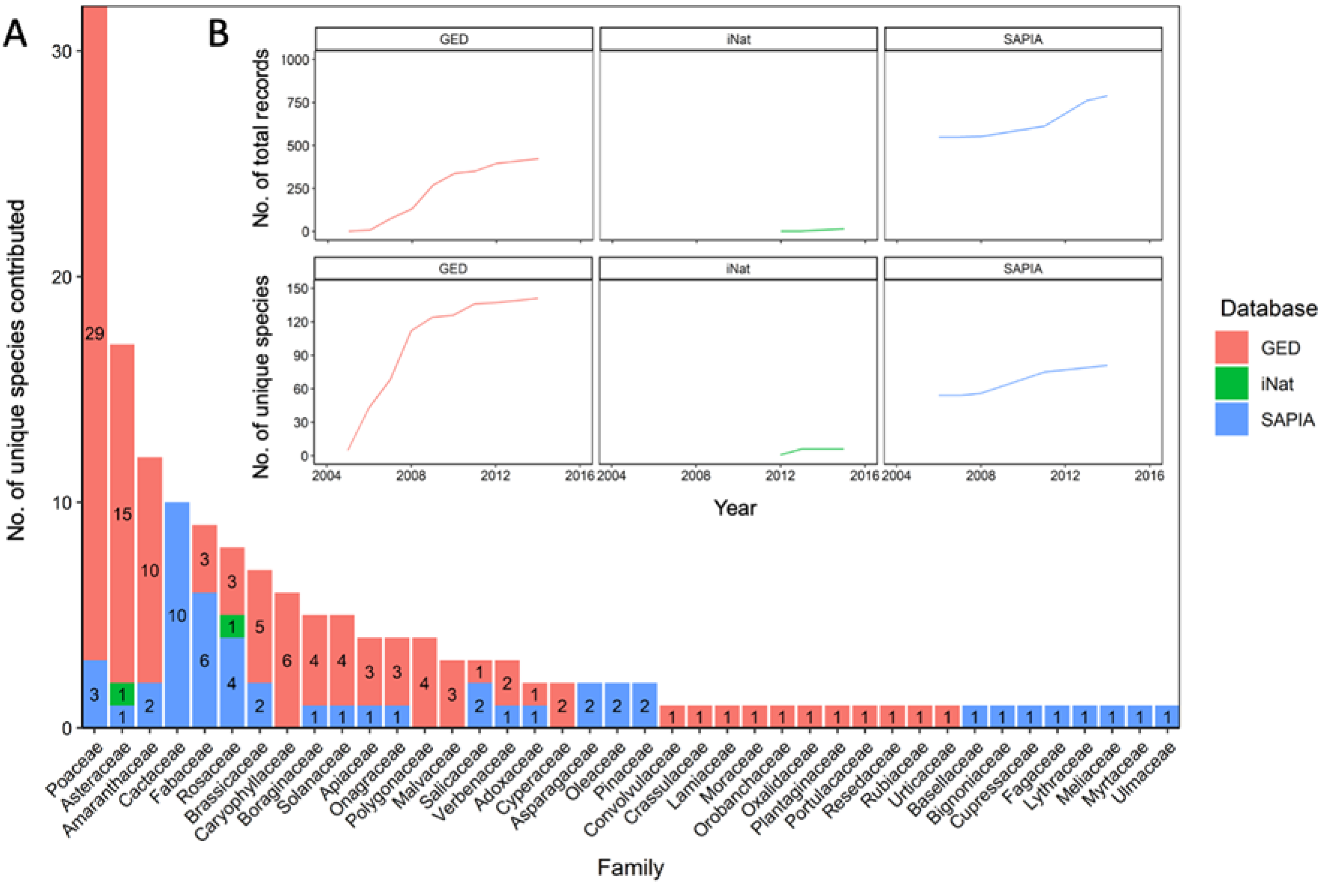
| Database | Total Records in SA | Total Species Richness in SA | Total Alien Plant Records in Mountain Areas | Percent of Records in Mountain Areas | Total Species Richness in Mountains | Observers | Methodology | Level of Botanical Verification | Spatial Scale | Temporal Scale | Funding Dependent | Alien Specific | Source |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SAPIA * | 80,226 | 969 | 18,278 | 26% | 588 | 710 | Nation-wide roadside survey, initially at the quarter degree scale, later using precise coordinates | High | National | 1979–2018 | Yes | Yes | Henderson and Wilson [55]; SAPIA Atlas [38] |
| iNaturalist * | 59,704 | 554 | 1472 | 3% | 239 | 9816 | Citizen science observations | Medium to high | Global | 2008–present (continuously updated) | No | No | Nugent [43] |
| GED | N/A | N/A | 466 | 100% | 160 | 3 | Botanist focused plant surveys | High | Regional | 2005−2014 | Yes | No | Barker [40] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Canavan, K.; Canavan, S.; Clark, V.R.; Gwate, O.; Richardson, D.M.; Sutton, G.F.; Martin, G.D. The Alien Plants That Threaten South Africa’s Mountain Ecosystems. Land 2021, 10, 1393. https://doi.org/10.3390/land10121393
Canavan K, Canavan S, Clark VR, Gwate O, Richardson DM, Sutton GF, Martin GD. The Alien Plants That Threaten South Africa’s Mountain Ecosystems. Land. 2021; 10(12):1393. https://doi.org/10.3390/land10121393
Chicago/Turabian StyleCanavan, Kim, Susan Canavan, Vincent Ralph Clark, Onalenna Gwate, David Mark Richardson, Guy Frederick Sutton, and Grant Douglas Martin. 2021. "The Alien Plants That Threaten South Africa’s Mountain Ecosystems" Land 10, no. 12: 1393. https://doi.org/10.3390/land10121393
APA StyleCanavan, K., Canavan, S., Clark, V. R., Gwate, O., Richardson, D. M., Sutton, G. F., & Martin, G. D. (2021). The Alien Plants That Threaten South Africa’s Mountain Ecosystems. Land, 10(12), 1393. https://doi.org/10.3390/land10121393