1. Introduction
Mining activities produce mining rock wastes (tailings) that can cover vast areas. In Québec (Canada), these tailing storage facilities cover over 13,000 ha [
1]. Tailing impoundments are among the most damaging and longest-lasting environmental liabilities of the mining industry [
2]. Their creation converts productive ecosystems into degraded landscapes, representing a loss of ecological services, such as wildlife habitat, nutrient cycling, and carbon sequestration [
3,
4,
5,
6].
Reclamation efforts by the mining industry aim to mitigate some of these environmental impacts by accelerating plant colonization on tailing storage facilities. Re-establishing ecological functions is challenging because multiple factors limit colonization, including soil compaction, low soil organic matter content, soil erosion as well as the poor nutrient availability and low water-holding capacity of the wastes [
7,
8,
9,
10]. Standard reclamation methods include the use of amendments and the seeding of herbaceous plants to favor colonization [
11,
12,
13,
14,
15,
16]. This method allows the establishment of a rapid protective vegetation cover that helps limit erosion, as requested by the guidelines for preparing mine closure plans in Québec [
17]. Although this business-as-usual (BAU) minimal requirement can initiate the reclamation process, the planting of trees combined with the application of organic amendments and the seeding of herbaceous plants could benefit even more the reclamation of tailings.
Plantations on mining sites may initiate natural succession, help integrate former mining sites into natural forested landscapes, and increase carbon sequestration [
18,
19]. Plantations also modify the microclimate in the understorey, which influences the colonising plant community [
20,
21,
22,
23]. For instance, a tree canopy decreases light availability. This shading reduces the growth of grasses, which compete with tree seedlings [
8,
24]. Few studies have examined the role of plantations in enhancing understorey biodiversity on mine sites; however, the limited available data suggest that planting native trees on degraded landscapes fosters more diverse plant communities [
23,
25].
Organic amendments can enhance soil fertility and increase plant success on reclaimed sites [
5,
11,
26]. Their application on tailing storage facilities can increase tree survival [
2], biomass, and coverage of understorey vegetation [
26,
27,
28,
29,
30]. When applied at high rates, however, amendments can negatively influence plant communities by, for example, decreasing richness and diversity [
31,
32,
33]. In Québec, Canada, various organic amendments are used to reclaim mining sites. Topsoil, as an amendment, is often available on site, as it is collected when the tailing storage facility is created or enlarged. Topsoil can enhance soil conditions on tailings in the short term by increasing C and N concentrations and enhancing microbial activity [
34]. However, this amendment is not always available in sufficient quantities and is often stockpiled before being applied, which can lead to its compaction and decreased seed viability [
29,
35,
36,
37]. As an alternative to topsoil, paper mill sludge biosolid (PMS), a by-product of the pulp and paper industry, represents a potential source of organic amendments for tailing reclamation, in particular because PMSs are presently landfilled and, therefore, lost to other uses [
38,
39]. Animal manure is another amendment used to increase crop productivity in agriculture; it could also benefit plant growth on reclaimed tailing storage facilities [
27,
29].
The major goal of mine site reclamation is to enhance site conditions and establish an ecosystem to a near-predisturbance state [
37]. Evaluating progress towards this goal requires comparing the reclaimed plant community to a “natural” state at a similar successional stage. In the boreal forest, the greatest plant diversity is found in the forest understorey, a layer dominated by vascular plants, mosses, and lichens [
40,
41,
42]. Nonetheless, cyclical and frequent natural disturbances, such as wildfires and insect outbreaks, continually modify the composition of these communities [
43]. These frequent disturbances allow reclaimed mine site communities to be compared with naturally disturbed understorey communities; for example, Errington and Pinno [
44] used post-fire forests as natural references to capture the first years of plant community succession following the removal of the forest canopy and understorey vegetation. Post-fire reference plots located close to tailing storage facilities can serve as valid points of comparison for recovering plant communities.
Although organic amendments and plantations are expected to benefit multiple aspects of tailing storage facility reclamation in the boreal region, little is known about their effect on plant community diversity in these settings. We hypothesised that (1) reclamation of mine tailing storage facilities through the planting of woody species, the seeding of herbaceous plants, and the use of soil organic amendments produces a greater plant diversity relative to the business-as-usual minimal requirements, which rely on seeding herbaceous plants and applying amendments; (2) reclamation methods using tree planting, herbaceous plants, and amendments establish plant communities more similar to those found on recently naturally disturbed reference sites (i.e., 11 years post-fire) than the business-as-usual method. We compared the effect of various organic amendment applications (topsoil, PMS, and chicken manure) on the response of the understorey plant community (total percent cover, richness, evenness, diversity, and functional-group abundance) at two mine tailing storage facilities in Québec, Canada, and compare these reclaimed plant communities with natural reference sites.
2. Materials and Methods
2.1. Site Description
We collected field data in 2018 at two tailings storage facilities in Québec, Canada. The first site is located at a niobium mine (Niobec, Inc., Saint-Honoré, QC, Canada) in St-Honoré, Saguenay (48°32′ N, 71°08′ W). This site lies within the balsam fir–yellow birch bioclimatic domain of the boreal zone [
45]. The region receives 934.5 mm of precipitation annually (of which 223 mm is snow) and has a mean annual temperature of 2.8 °C [
46]. The second site is situated at an iron mine tailings facility operated by ArcelorMittal Mining Canada at Mont-Wright, Fermont (52°46′ N, 67°20′ W). This mine lies within the spruce–lichen bioclimatic domain [
45]. The region receives 839.5 mm of precipitation annually (of which 428.7 mm falls as snow) and has a mean daily temperature of −3.1 °C [
47]. At both sites, the tailings are non-acidic (pH 7–8) with a relatively coarse texture (150–180 μm). Our experimental plots were established on slopes of 15% at the Niobec mine and 10% at Mont-Wright.
We compared our reclaimed sites with recent post-fire natural reference sites, which were previously forested stands. We selected the reference sites, one per mining site, according to their proximity to the respective mine sites and the time since the last fire disturbance. Given the very limited number of reference sites fitting our criteria, our selected natural reference sites did not burn in the same years as the reclamation (2012 at Niobec and 2015 at Mont-Wright) within our experimental sites. Near the Niobec mine, our reference site was a mixed forest stand that burned in 2007 (48°29′30.4″ N, 71°00′53.3″ W), and the Mont-Wright reference stand was a young black spruce forest that burned in 2007 (52°15′11.0″ N, 67°41′27.5″ W).
2.2. Experimental Design
At the Niobec tailing storage facilities site, we established, in 2012, a complete randomized block and factorial design (split-split-plot) with four replicates. We tested ten reclamation treatments involving combinations of organic amendments (topsoil or topsoil paper mill sludges (PMS) mixture) and revegetation with trees (larch (Larix laricina (Du Roi) K. Koch), paper birch (Betula papyrifera Marshall), red pine (Pinus resinosa Aiton), a treatment with the three mixed tree species (larch, paper birch, and red pine), and a control without trees).
At the Niobec site, a 10 cm layer of topsoil was spread onto the tailings from an all-terrain vehicle (Caterpillar D10) and evened out over the ground. The vehicle operator also removed any large rocks found within the topsoil. The topsoil had been excavated from another area of the tailing facility and stockpiled for two years prior to its use. We split each block (25 m × 600 m) into two main plots. Each plot was randomly attributed to one of two organic amendment treatments: (1) no application of amendment or (2) the application (and rototilling) of PMS (obtained from Resolute Forest Products, Jonquière, Québec; see
Table 1 for its chemical characteristics) at a rate of 35 Mg(dry)·ha
−1. The PMS used contained a very low concentration of heavy metals (analyses not shown), according to Canadian standards [
48]. We, then, divided each main plot into five equal subplots (25 m × 60 m) in which we planted 40 to 60 cm-tall trees at a density of 2250 trees·ha
−1 (2 × 2 m spacing). We selected the tree species based on 11 criteria, including tolerance to drought, nutrient requirements, and cost (unpublished data), which had been developed by Niobec, Inc. We randomly attributed one of five selected tree covers to each subplot. The tree cover was either (1) no trees planted; (2)
Larix laricina; (3)
Pinus resinosa (specimens of both
Larix laricina and
Pinus resinosa were obtained from the Normandin nursery of the Québec Ministry of Forests, Wildlife and Parks); (4)
Betula papyrifera (obtained from the Boucher nursery, St-Ambroise, Québec); (5) a mixture of all three tree species. We, then, seeded all plantation plots with clovers (
Trifolium spp.) at a rate of 50 kg·ha
−1. A clover treatment (without tree) represents the business-as-usual reclamation scenario at the Niobec site.
For the Mont-Wright site, reclamation began on the tailing storage facility in 2013. About half of the reclaimed block received a “Norco” treatment representing a mixture of chicken manure (5 Mg·ha
−1), hay, and herbaceous seeds (grass and forb seeds spread at a rate of 220 kg·ha
−1; see Juge and Cossette [
49] for the composition of the seed mix). We established our experimental plots on the Mont-Wright tailing facilities in 2015 and applied a randomized block design with six amendment treatments, repeated in three blocks (156 m × 25 m; see
Table 1 for the chemical characteristics of the applied amendments). Each plot measured 26 m × 25 m. The amendments included topsoil, PMS, and the Norco treatment (chicken manure). The topsoil was collected in 2015 from another area of the tailing facility, and we applied the topsoil to the appropriate plots to a depth of approximately 10 cm using an all-terrain vehicle, as done for the Niobec site. PMS had been landfilled with ash before its recovery and application (the PMS was obtained from Resolute Forest Products, Baie-Comeau, Québec; see
Table 1 for PMS chemical characteristics). We removed the woody debris (stumps and branches) from all plots.
The six treatments were randomly assigned to six main plots. These treatments consisted of (1) a five-year Norco mixture (N5; plots reclaimed with Norco in 2013), (2) PMS applied at a rate of 50 Mg(dry)·ha−1, (3) PMS applied at a rate of 50 Mg(dry)·ha−1 on top of the N5 treatment (PMS50+N5), (4) topsoil, (5) topsoil on top of the N5 treatment (topsoil+N5), and (6) a three-year Norco mixture (N3; plots amended with Norco in 2015). The application of the Norco treatment alone is the business-as-usual approach used at the Mont-Wright site.
We selected a combination of local tree and shrubs species (jack pine (Pinus banksiana Lambert), green alder (Alnus alnobetula subsp. crispa (Aiton) Raus), and a hybrid poplar (Populus sp. Clone 915318)) on the basis of a preliminary greenhouse-based study investigating tree survival and growth (results not shown)49. We planted this mixture of species on all experimental units. Jack pine and hybrid poplar were obtained from the MFFP (Normandin nursery). We obtained green alder from the Girardville nursery (Girardville, Québec), and these alders were inoculated with Frankia (an N-fixing bacteria) at the Université du Sherbrooke, Québec.
2.3. Vegetation Survey
We conducted test replicates to determine the appropriate quadrat size for vegetation samplings. We confirmed that a 1 m2 quadrat makes accurate estimates for our sites. A quadrat was set in the center of each experimental unit on the tailing sites. At the reference sites, we established three quadrats aligned along cardinal directions (north, south, and west) at 3 m from the site center. Within each quadrat, we assessed plant communities between June and August 2018 by visually determining the cover (%) of these species six years (Niobec) and three years (Mont-Wright) post-reclamation. We assessed the combined cover; therefore, the percent cover for a quadrat can exceed 100%. Species were identified to the lowest possible taxonomic level. The inventory included vascular plants, mosses, and lichens. For both sites, the same person conducted all surveys. We collected a sample of each species to confirm its identification in the laboratory (at the vegetation and animal ecology lab at the Université du Québec à Chicoutimi, the bryology lab at Université du Québec en Abitibi-Témiscamingue, and the Louis-Marie Herbarium at Université Laval).
2.4. Statistical Analysis
Statistical analyses were conducted using R software, version 3.6.1 [
50]. Total percent cover, species richness (S), Pielou’s evenness (J′), and Simpson’s diversity (1−D) were calculated using the “vegan” package [
51]. We assessed the data for homogeneity of variance and transformed data when necessary (only richness at Mont-Wright was log-transformed). Sources of variation for the Niobec site were: (1) number of blocks (
n = 4; random), (2) amendment application (PMS35+topsoil and topsoil;
n = 2; fixed), and (3) the type of woody species used in plantations (
L. laricina; P. resinosa;
B. papyrifera; a mixture of these tree species; the no plantation control;
n = 5; fixed). Sources of variation at the Mont-Wright site were: (1) number of blocks (
n = 3; random) and (2) treatments applied (PMS50, PMS+N5, topsoil, topsoil+N5, N3, N5;
n = 6; fixed).
We used ANOVA to test for differences between plant community responses in terms of total percent cover, S, J′, 1−D, and functional groups (grasses, forbs, and mosses). We ran post hoc tests (estimated marginal means (least-squares means)) when the effects were significant (
p < 0.05). We analyzed community structures via multivariate analyses based on Bray–Curtis dissimilarity distances and matrices. Species assemblage data were transformed through square-root transformation, as suggested by Clarke and Warwick [
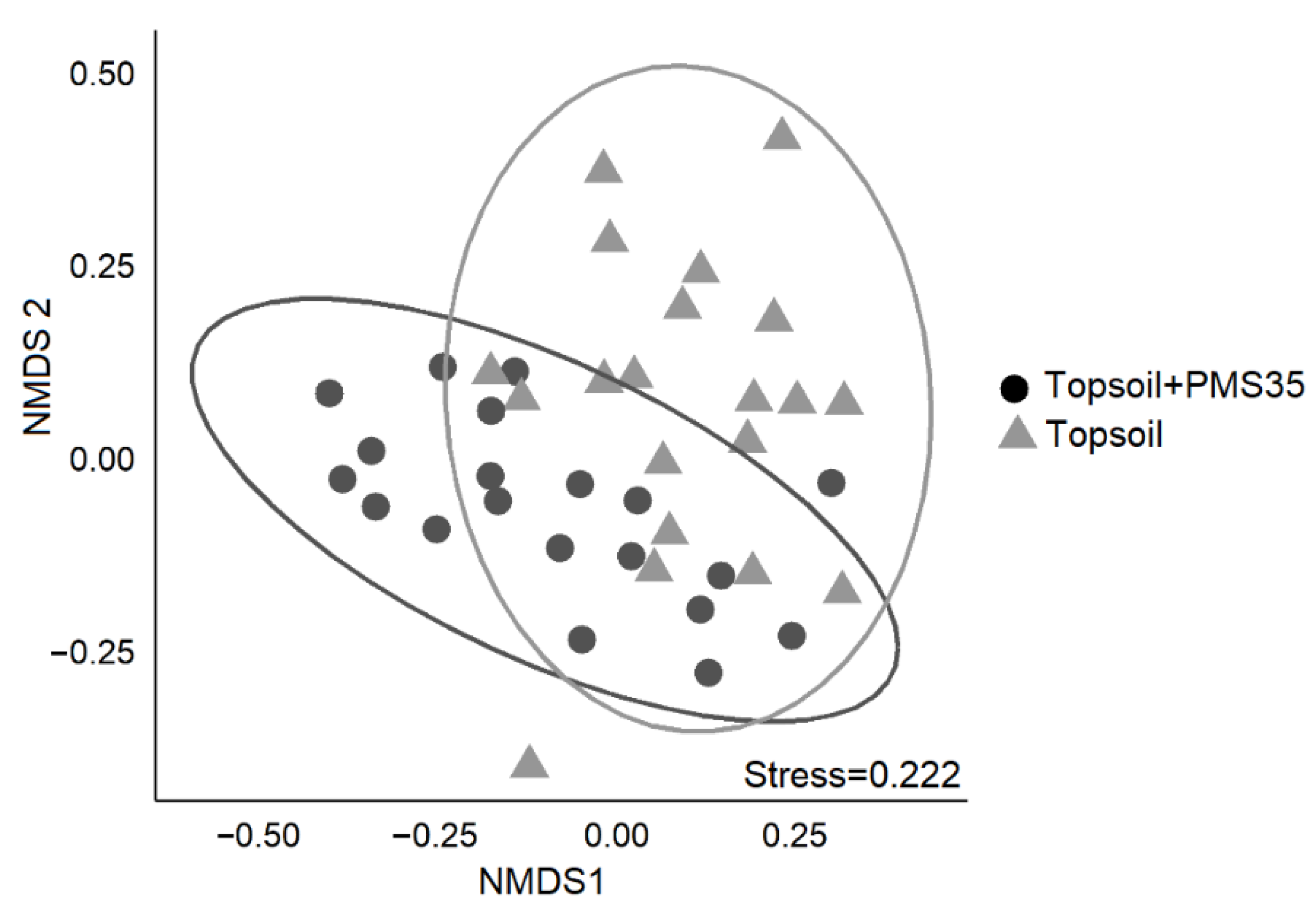
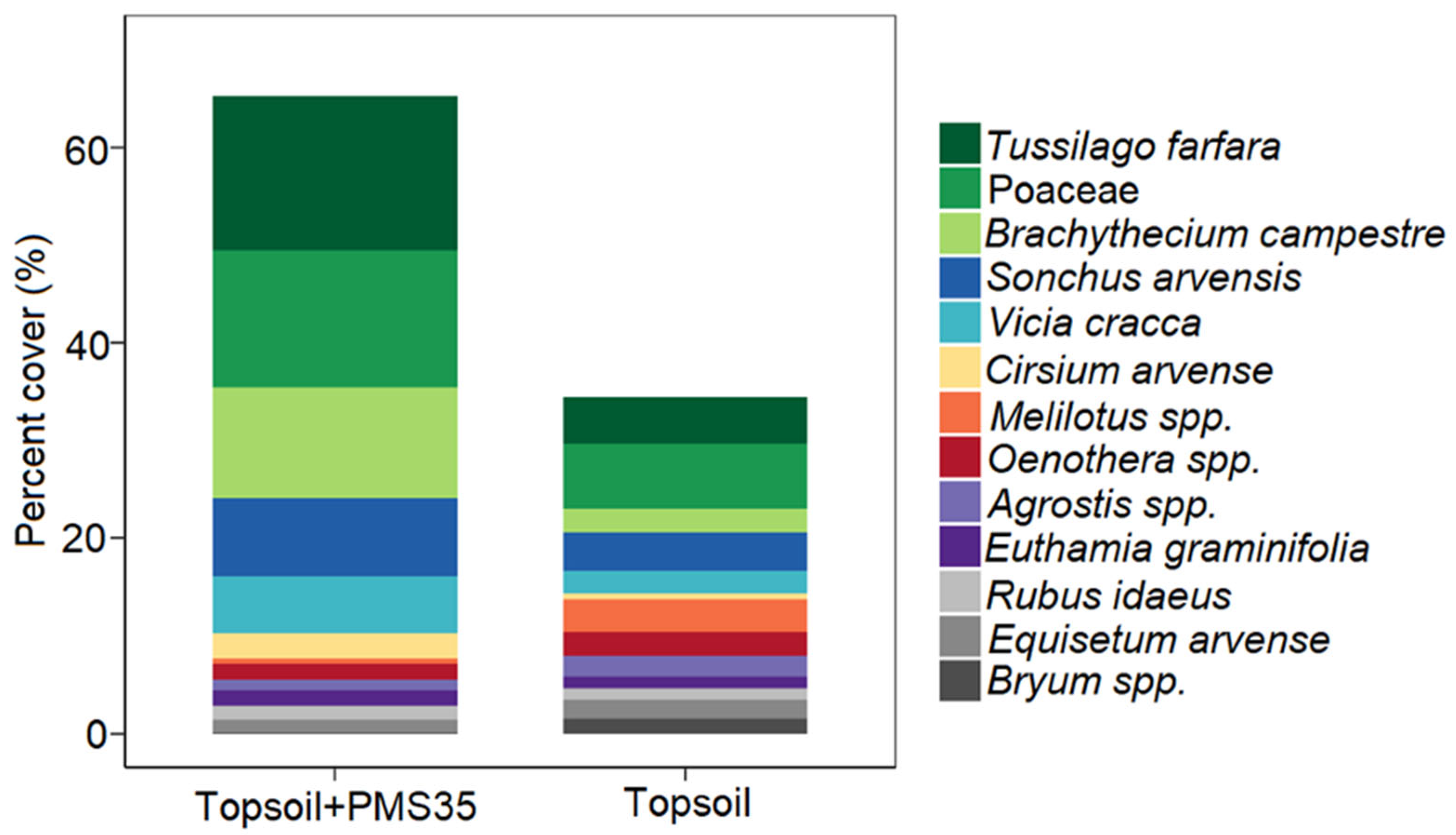
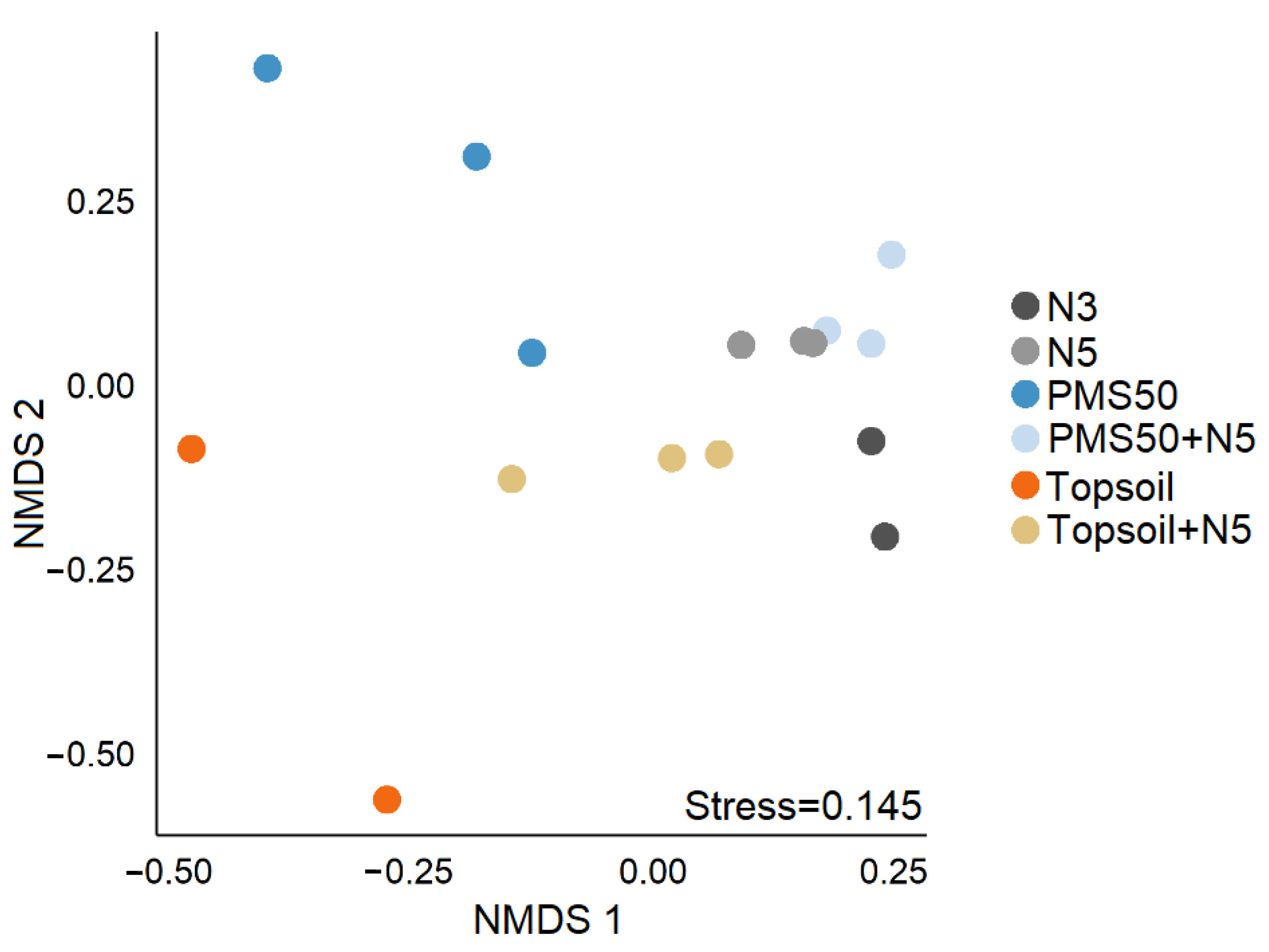
52]. To reduce noise in the dataset, we removed single-occurrence taxa before performing our analyses. We tested for differences in community structure among treatments using permutational multivariate analysis of variance (PERMANOVA). We then used non-metric multidimensional scaling (NMDS) to display dissimilarities between samples on a two-dimensional ordination. Finally, we ran SIMPER to identify the discriminant species that could explain differences in diversity between treatments at the Niobec site. We did not perform any statistical analyses on the reference sites because these sites were not part of the experimental design. The results measured at the references sites were used as indicators to make comparisons between natural disturbed sites and reclaimed mine sites.
5. Conclusions
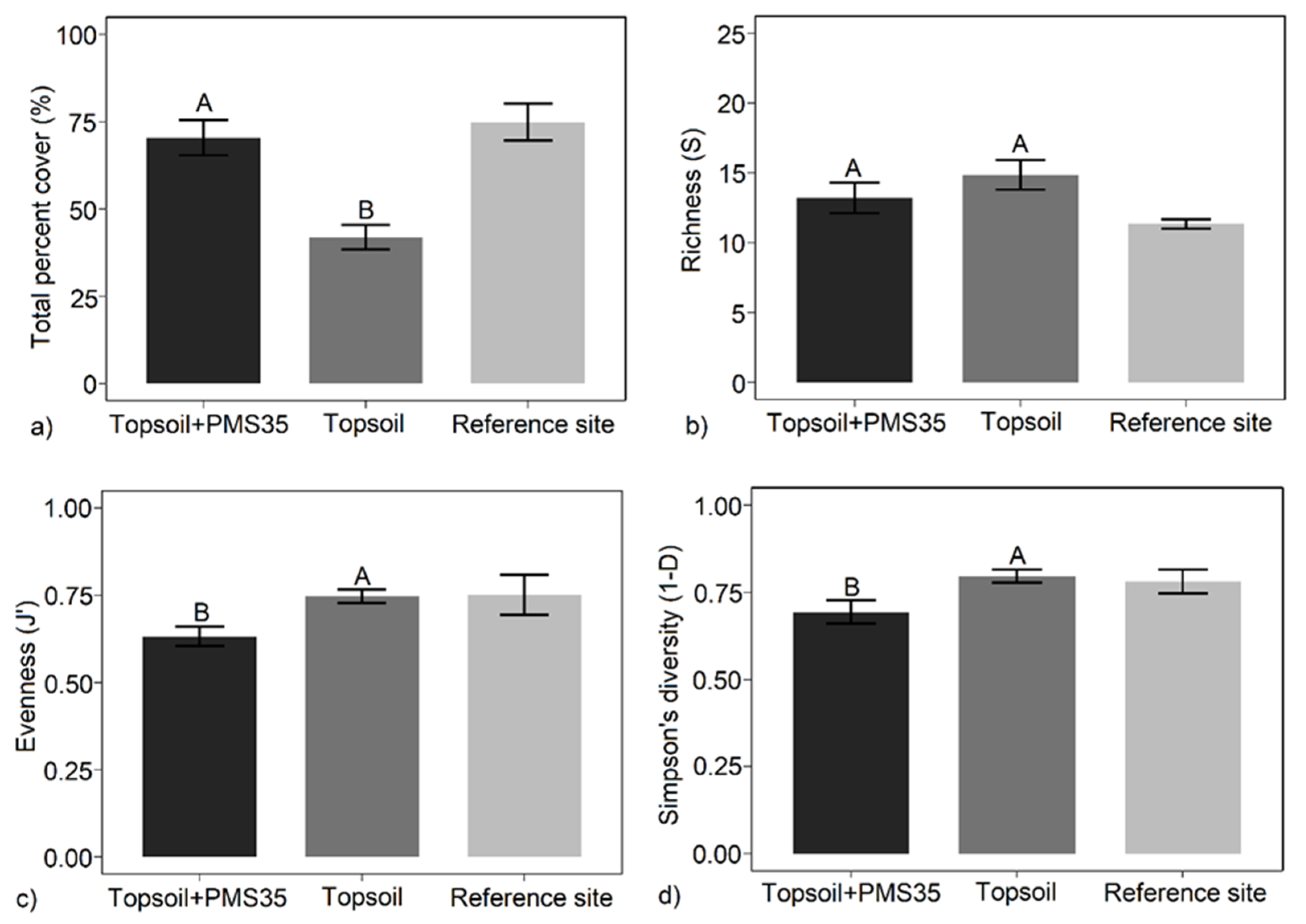
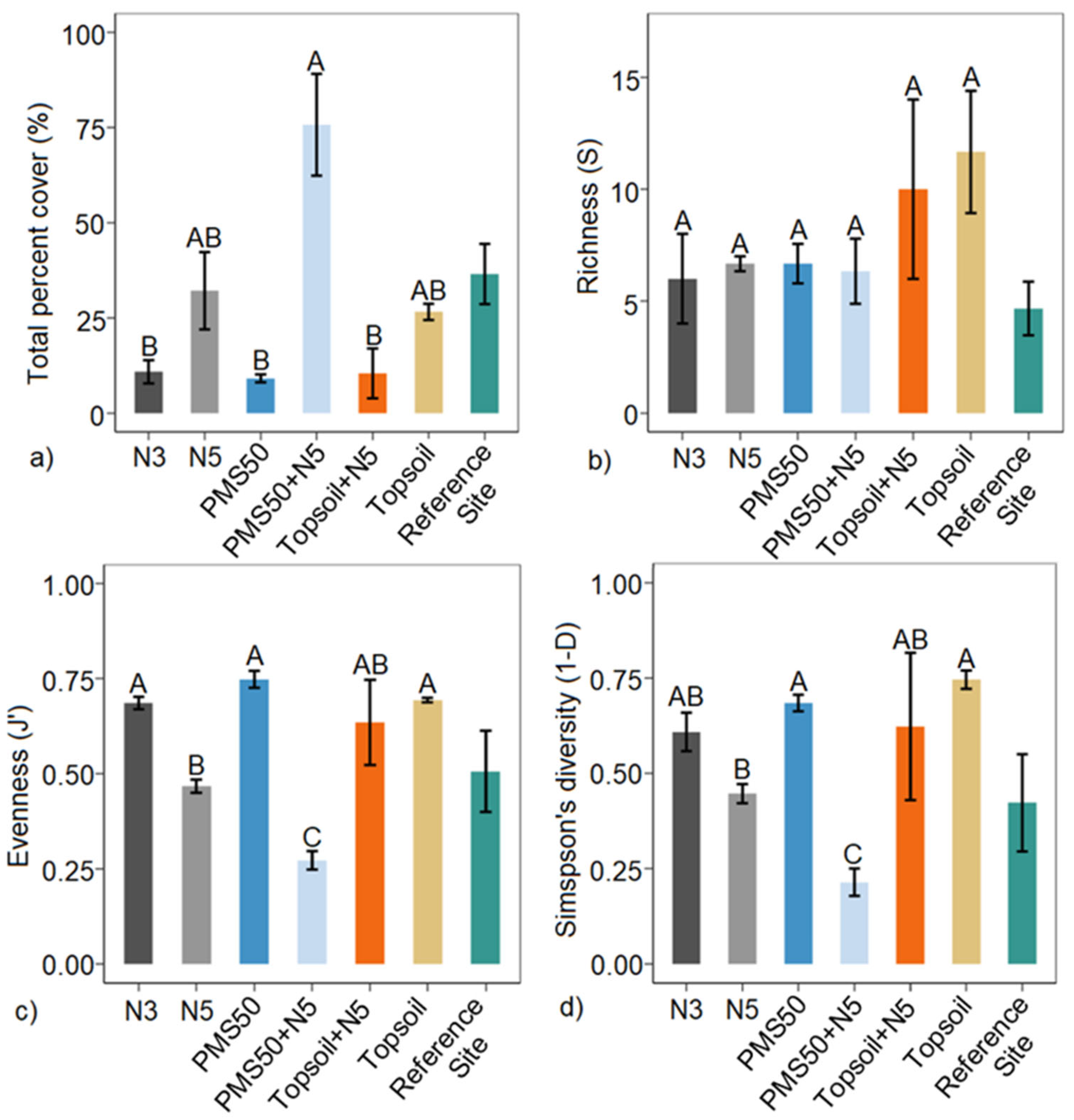
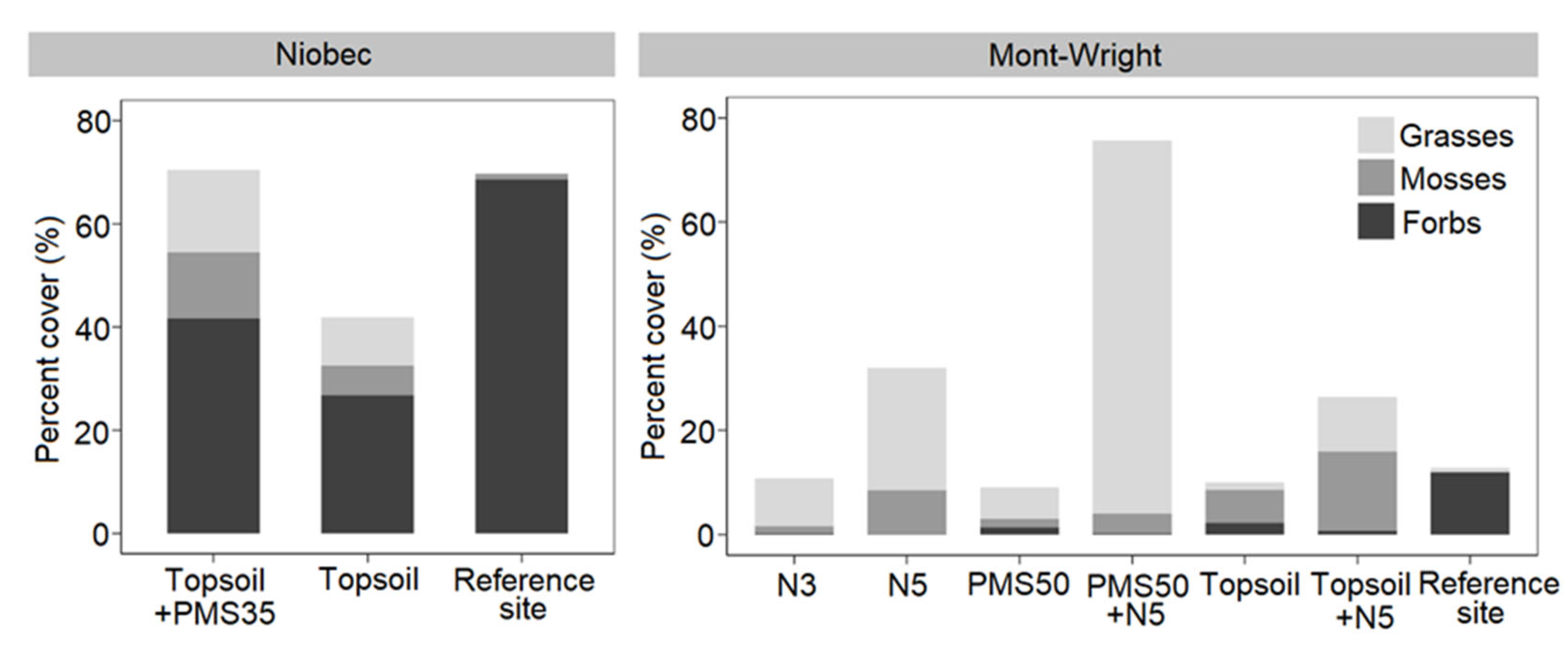
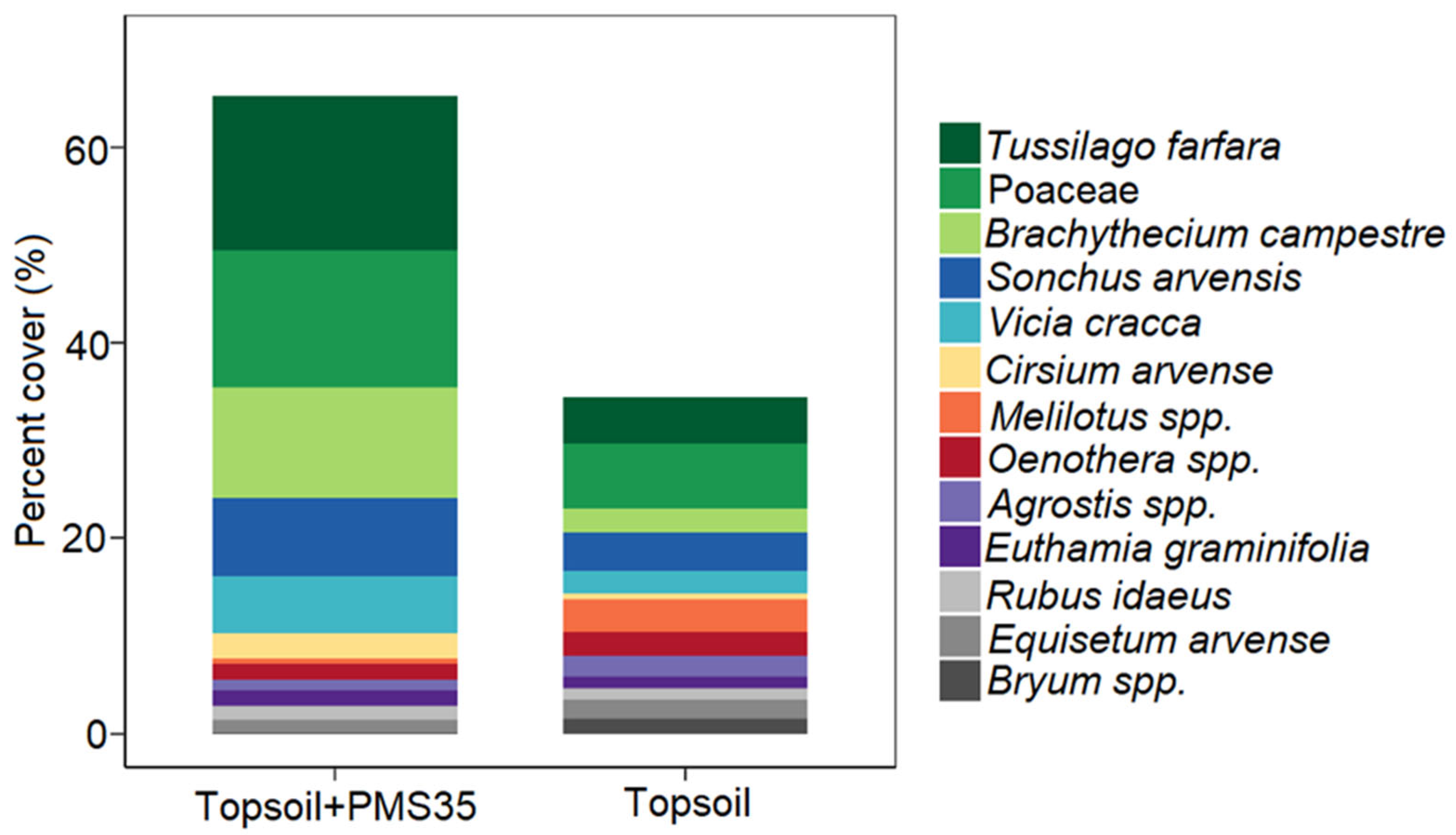
Our study showed that the application of an organic amendment had a significant influence on plant community response (total percent cover, J′, and 1−D) at both mine tailing sites (Niobec and Mont-Wright mines, Québec), whereas plantations did not produce any community response at the Niobec site. The application of PMS on topsoil produced less diverse communities than the use of the business-as-usual topsoil-only minimal method, six years after the reclamation. At the Mont-Wright site, the early response of vegetation (three years post-reclamation) showed that the combination of the Norco treatment (chicken manure, hay, and herbaceous seeds) with topsoil and the application of topsoil only or biosolids only resulted in the highest values of evenness (J′) and diversity (1−D).
Plant communities on tailings were distinct from those found on post-fire forest reference stands. We measured a higher proportion of mosses and grasses at the Niobec site than its reference site. At Mont-Wright, topsoil enhanced the abundance of mosses, and plots that received the Norco treatment mixed with PMS contained a high abundance of grasses. Natural sites contained mostly forbs. Our results provide valuable insight regarding the influence of different reclamation methods on the plant communities of reclaimed mine sites. Further research should explore the longer-term influences of these amendments in other climate zones and applied at different rates, including cost–benefit analyses on the selection of different organic amendments for tailing reclamation.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}