
Diclofenac Produces Diverse Toxic Effects on Aquatic Organisms of Different Trophic Levels, Including Microalgae, Cladocerans, and Fish

 ,
, 
Abstract
1. Introduction
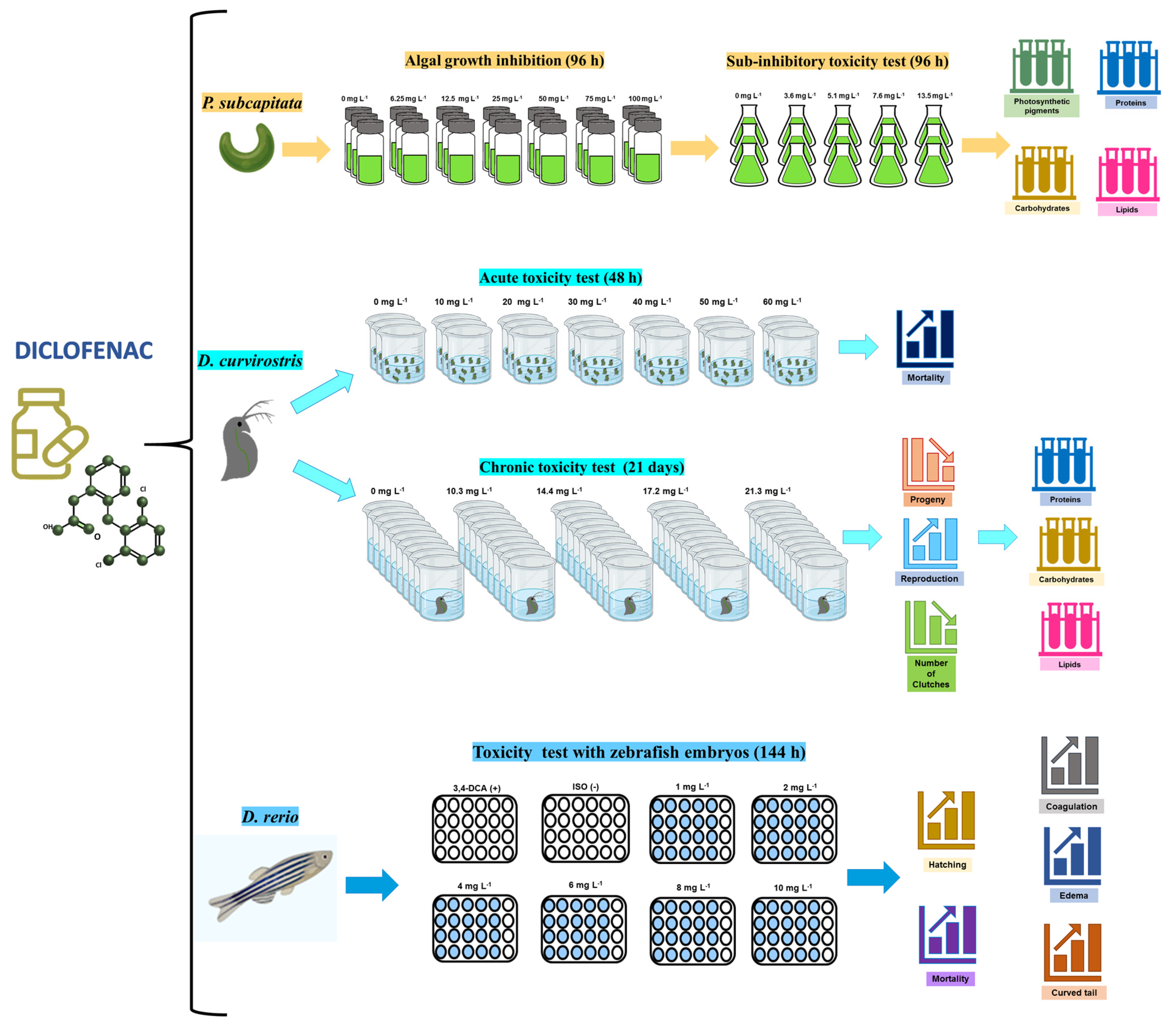
2. Materials and Methods
2.1. Test Organisms
2.2. Microalga
2.2.1. Toxicity of Diclofenac on the Growth of the Microalga P. subcapitata
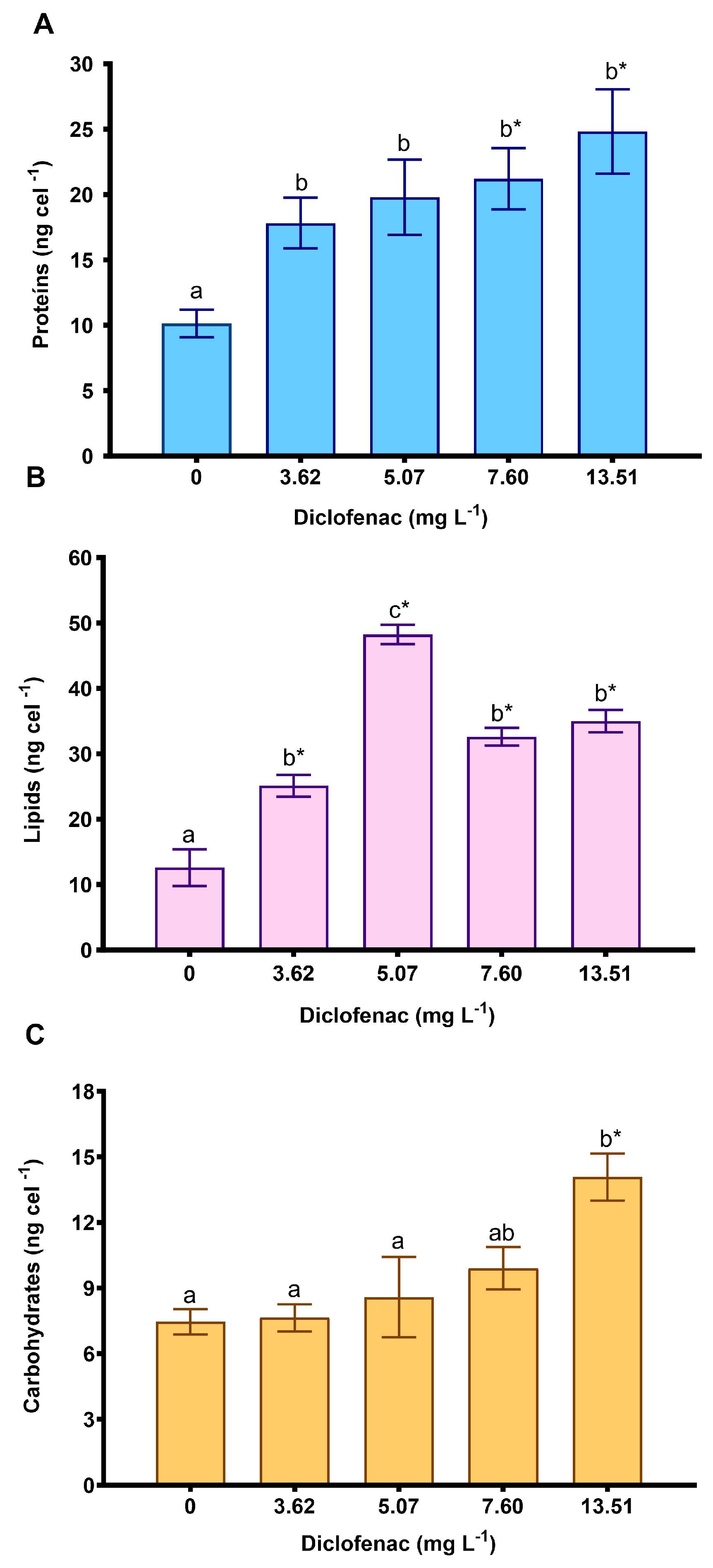
2.2.2. Effect of Diclofenac on Photosynthetic Pigments and Macromolecule Concentrations
- Chlorophyll-a = 12.19 A665 − 3.45 A649
- Chlorophyll-b = 21.99 A649 − 5.32 A665
- Carotenoids = (1000 A480 − 2.14 Chlorophyl-a − 70.16 Chlorophyl-b)/220
2.3. Toxicity of Diclofenac in Daphnia curvirostris
2.3.1. Acute and Chronic Toxicity
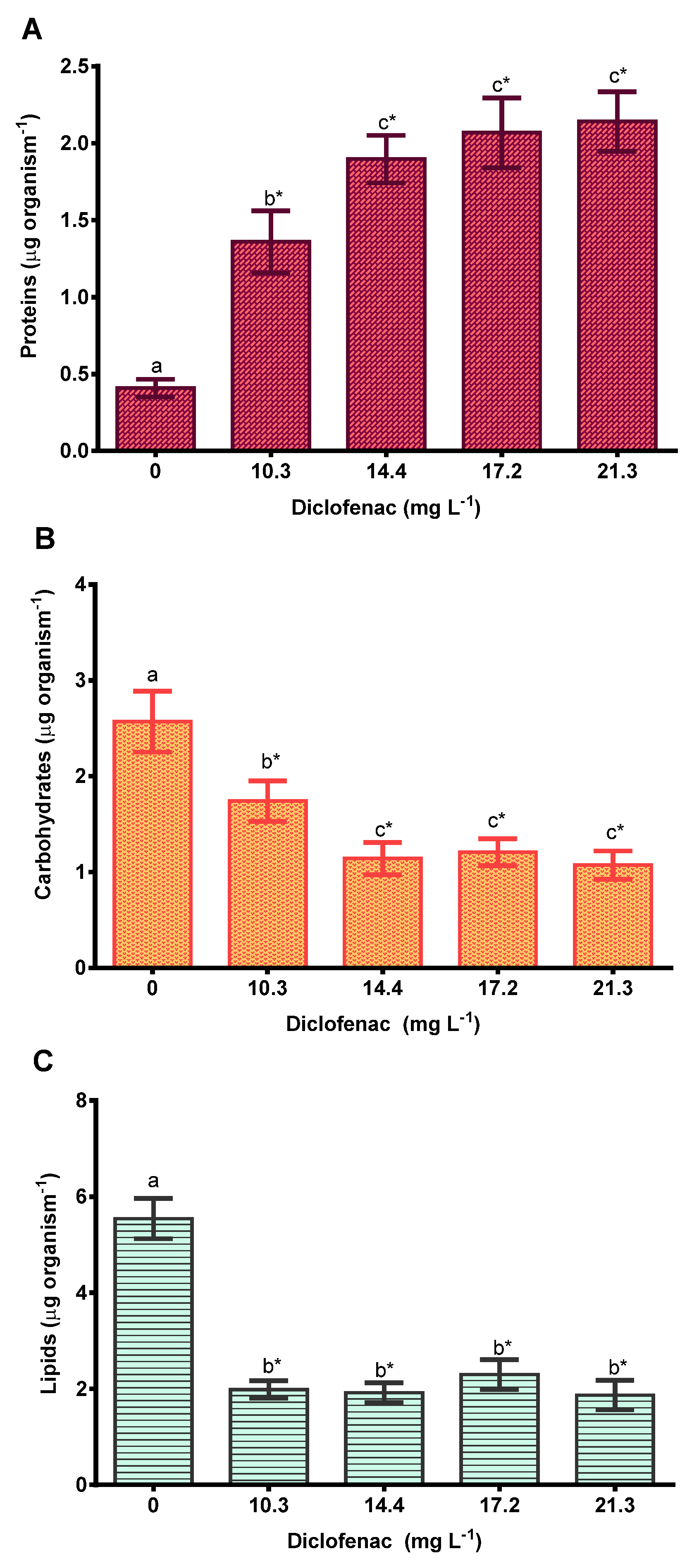
2.3.2. Toxic Effects of Diclofenac on the Biomolecule Concentrations of Progeny
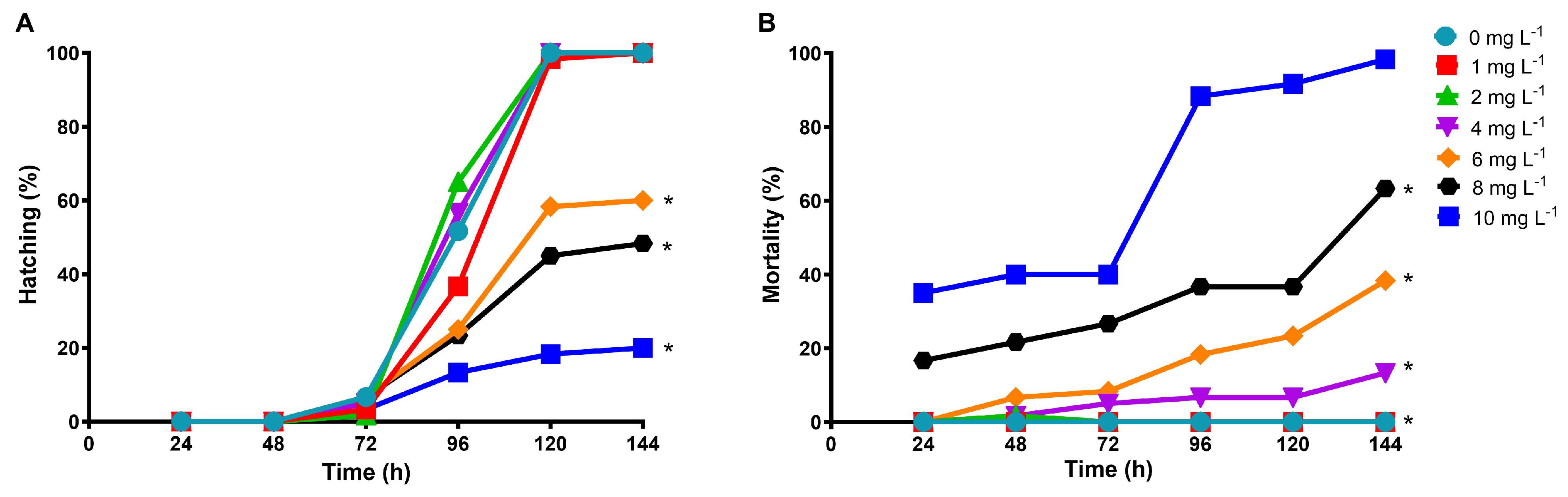
2.4. Lethal and Sublethal Effects of Diclofenac on Danio rerio Embryos
2.5. Statistical Analysis
3. Results
3.1. Effect of Diclofenac on P. subcapitata: Growth, Photosynthetic Pigments, and Macromolecules
3.2. Effect of Diclofenac on the Cladoceran D. curvirostris
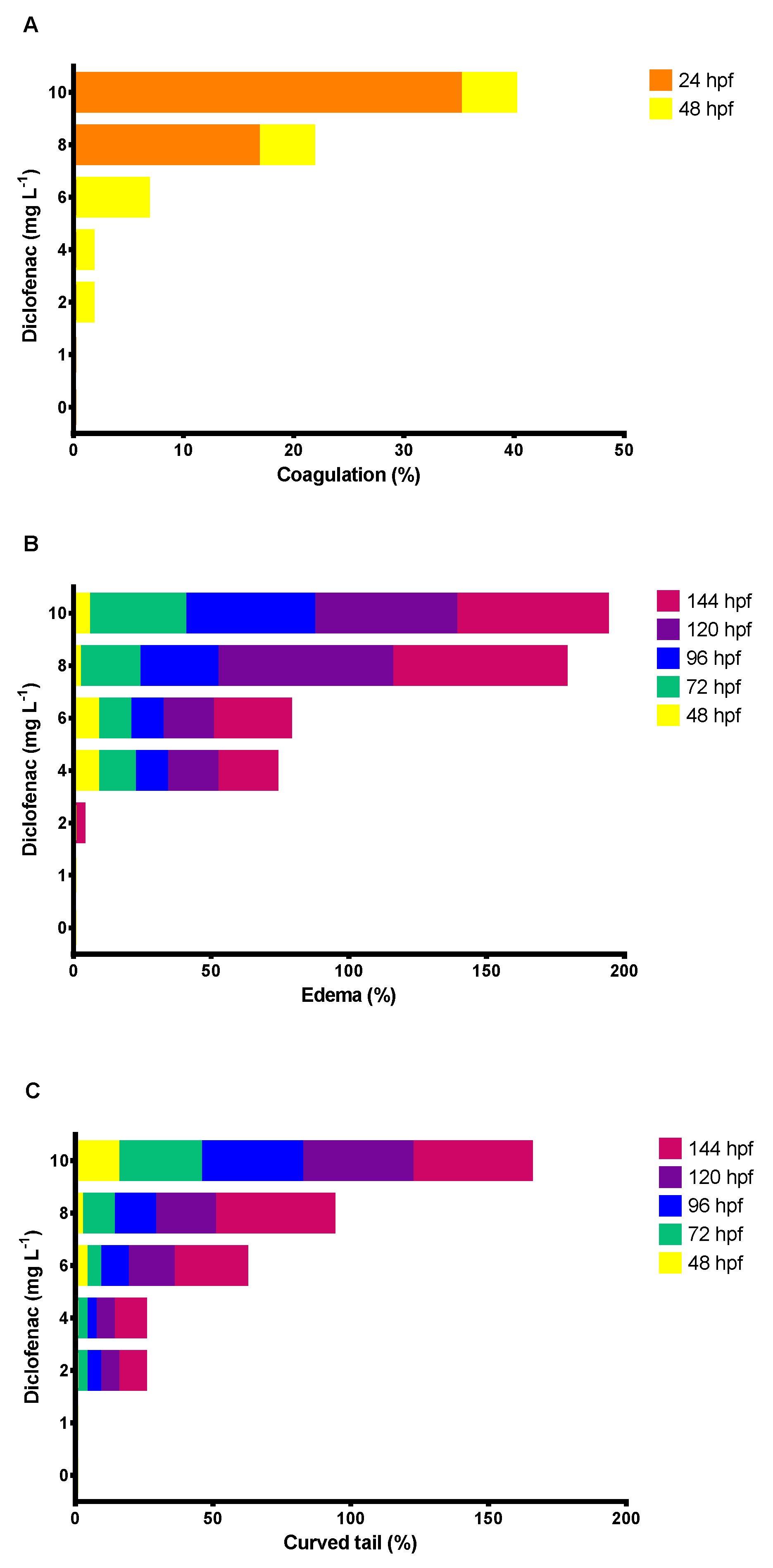
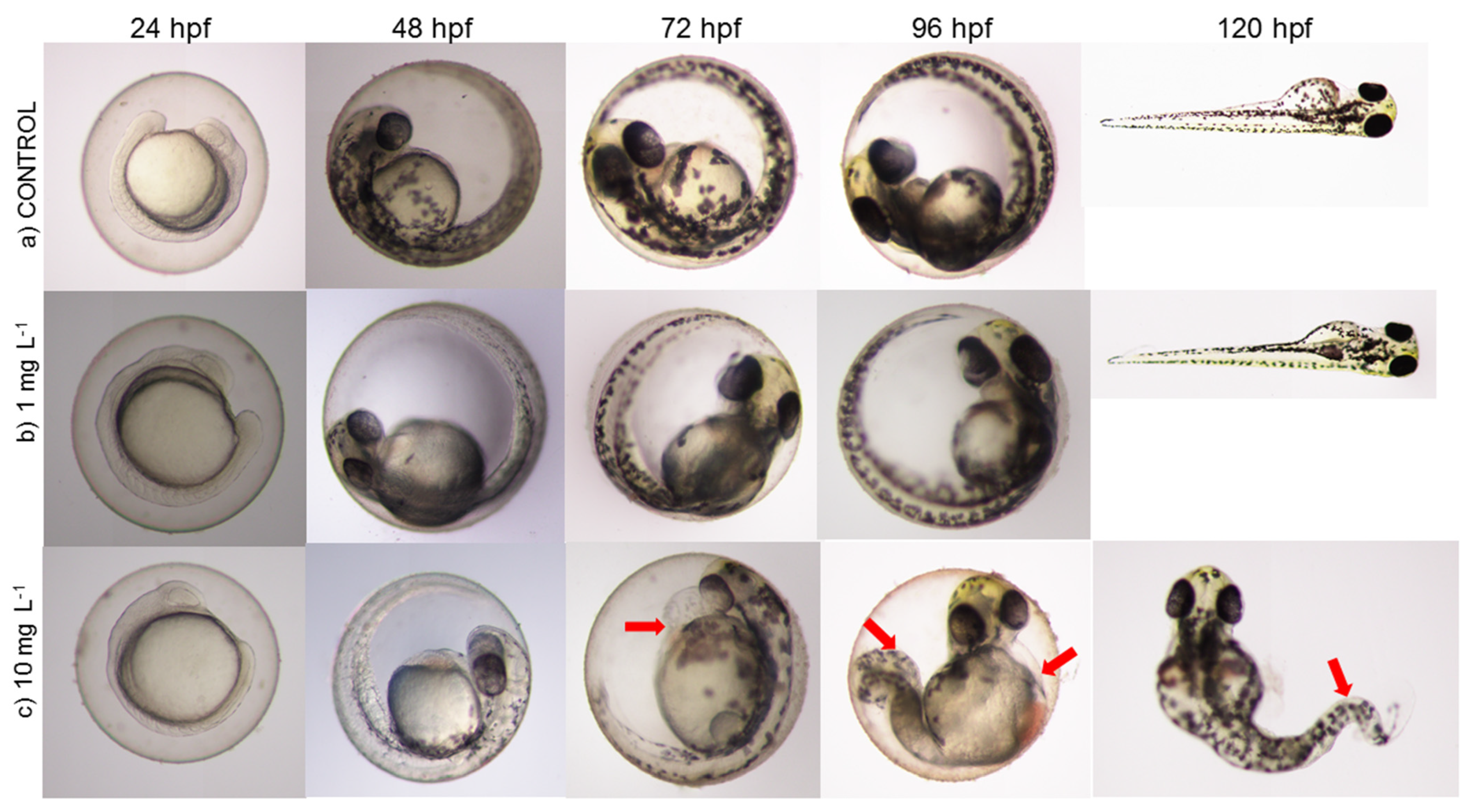
3.3. Effect of Diclofenac on D. rerio Embryos
4. Discussion

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Chemical Compound | Exposure Time | IC50/EC50/LC50 | Reference |
|---|---|---|---|---|
| Pseudokircheneriella subcapitata | Diclofenac | 96 h | IC50: 16.3 mg L−1 | Ferrari et al. [48] |
| P. subcapitata | Diclofenac | 72 h | IC50: 64.8 mg L−1 | Quinn et al. [49] |
| P. subcapitata | Diclofenac | 96 h | EC50: 6 mg L−1 | Zind et al. [50] |
| P. subcapitata | Hydroxy-diclofenac | 96 h | IC50: 3.42 mg L−1 | Zind et al. [50] |
| P. subcapitata | 2-[(2-chlorophenyl) amino] benzaldehyde | 96 h | IC50: 8.62mg L−1 | Zind et al. [50] |
| P. subcapitata | Diclofenac | 96 h | IC50: 16.57 mg L−1 | This study |
| Daphnia magna | Diclofenac | 48 h | EC50: 22.4 mg L−1 | Ferrari et al. [48] |
| D. magna | Diclofenac | 48 h | EC50: 39.9-44.7 mg L−1 | Haap et al. [61] |
| D. magna | Diclofenac | 48 h | LC50: 80.1 mg L−1 | Zind et al. [50] |
| D. magna | Diclofenac | 48 h | LC50: 24.6 g L−1 | Barbosa et al. [63] |
| D. similis | Diclofenac | 48 h | EC50: 46 mg L−1 | Junqueira de Castro et al. [62] |
| D. curvirostris | Diclofenac | 48 h | LC50: 32.29 mg L−1 | This study |
| Danio rerio | Diclofenac | 120 h | LC50: 37.7 mg L−1 | Chabchoubi et al. [80] |
| D. rerio | Diclofenac | 144 h | LC50: 6.11 mg L−1 | Praskova et al. [81] |
| D. rerio | Diclofenac | 96 h | LC50: 166.6 mg L−1 | Praskova et al. [81] |
| D. rerio | Diclofenac | 72 h | LC50: 7.8 mg L−1 | van den Brandhof and Montforts [82] |
| D. rerio | Diclofenac | 72 h | EC50: 5.3 mg L−1 | van den Brandhof and Montforts [82] |
| D. rerio | Diclofenac | 96 h | LC50: 480 μg L−1 | Dietrich and Prietz [83] |
| D. rerio | Diclofenac | 144 h | LC50: 6.27 mgL−1 | This study |
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Lonappan, L.; Brar, S.K.; Das, R.K.; Verma, M.; Surampalli, R.Y. Diclofenac and its transformation products: Environmental occurrence and toxicity-a review. Environ. Int. 2016, 96, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Ellis, J.B. Pharmaceutical and personal care products (PPCPs) in urban receiving waters. Environ. Pollut. 2006, 144, 184–189. [Google Scholar] [CrossRef] [PubMed]
- Abdulrazaq, Y.; Abdulsalam, A.; Rotimi, A.L.; Abdulbasit, A.A.; Clifford, O.; Abdulsalam, O.A.; Bilal, M. Classification, Potential Routes and Risk of Emerging Pollutants/Contaminant. In Emerging Contaminants; Nuro, L., Ed.; IntechOpen: London, UK, 2020; pp. 1–12. [Google Scholar]
- Mezzelani, M.; Gorbi, S.; Regoli, F. Pharmaceuticals in the aquatic environments: Evidence of emerged threat and future challenges for marine organisms. Mar. Environ. Res. 2018, 140, 41–60. [Google Scholar] [CrossRef] [PubMed]
- Wada, O.Z.; Olawade, D.B. Recent occurrence of pharmaceuticals in freshwater, emerging treatment technologies, and future considerations: A review. Chemosphere 2025, 374, 144153. [Google Scholar] [CrossRef]
- Parolini, M. Toxicity of the Non-Steroidal Anti-Inflammatory Drugs (NSAIDs) acetylsalicylic acid, paracetamol, diclofenac, ibuprofen and naproxen towards freshwater invertebrates: A review. Sci. Total Environ. 2020, 740, 140043. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information NCBI. 2023; PubChem Compound Summary for CID 3033, Diclofenac. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Diclofenac (accessed on 13 June 2024).
- Lara-Pérez, C.; Leyva, E.; Zermeño, B.; Osorio, I.; Montalvo, C.; Moctezuma, E. Photocatalytic degradation of diclofenac sodium salt: Adsorption and reaction kinetic studies. Environ. Earth Sci. 2020, 79, 277. [Google Scholar] [CrossRef]
- Gómez-Oliván, L.; Carmona-Zepeda, F.; Galar-Martínez, M.; Téllez-López, A.; Amaya Chávez, A. Estudio de automedicación en una farmacia comunitaria de la ciudad de Toluca. Rev. Mex. Cienc. Farma 2009, 40, 5–11. [Google Scholar]
- Luongo, G.; Guida, M.; Siciliano, A.; Libralato, G.; Saviano, L.; Amoresano, A.; Zarrelli, A. Oxidation of diclofenac in water by sodium hypochlorite: Identification of new degradation by-products and their ecotoxicological evaluation. J. Pharm. Biomed. Anal. 2021, 194, 113762. [Google Scholar] [CrossRef]
- Mohan, H.; Rajput, S.S.; Jadhav, E.B.; Sankhla, M.S.; Sonone, S.S.; Jadhav, S.; Kumar, R. Ecotoxicity, Occurrence, and Removal of Pharmaceuticals and Illicit Drugs from Aquatic Systems. Bio. Res. Appl. Chem 2021, 11, 12530–12546. [Google Scholar]
- Langford, K.H.; Reid, M.; Thomas, K.V. Multi-residue screening of prioritised human pharmaceuticals, illicit drugs and bactericides in sediments and sludge. J. Environ. Monit. 2011, 13, 2284–2291. [Google Scholar] [CrossRef]
- Kunkel, U.; Radke, M. Fate of pharmaceuticals in rivers: Deriving a benchmark dataset at favorable attenuation conditions. Water Res. 2012, 46, 5551–5565. [Google Scholar] [CrossRef] [PubMed]
- Carmona, E.; Andreu, V.; Picó, Y. Occurrence of acidic pharmaceuticals and personal care products in Turia River basin: From waste to drinking water. Sci. Total Environ. 2014, 484, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Simazaki, D.; Kubota, R.; Suzuki, T.; Akiba, M.; Nishimura, T.; Kunikane, S. Occurrence of selected pharmaceuticals at drinking water purification plants in Japan and implications for human health. Water Res. 2015, 76, 187–200. [Google Scholar] [CrossRef]
- Tröger, R.; Klöckner, P.; Ahrens, L.; Wiberg, K. Micropollutants in drinking water from source to tap—Method development and application of a multiresidue screening method. Sci. Total Environ. 2018, 627, 1404–1432. [Google Scholar] [CrossRef]
- Shamsudin, M.S.; Farhan, S.A.; Suzylawati, I. A review of diclofenac occurrences, toxicology, and potential adsorption of clay-based materials with surfactant modifier. J. Environ. Chem. Eng. 2022, 10, 107541. [Google Scholar] [CrossRef]
- Iglesias, A.; Nebot, C.; Vázquez, B.I.; Coronel-Olivares, C.; Abuín, C.M.F.; Cepeda, A. Monitoring the presence of 13 active compounds in surface water collected from rural areas in Northwestern Spain. Int. J. Environ. Res. Public Health 2014, 11, 5251–5272. [Google Scholar]
- Hashim, N.H.; Nasir, M.H.; Ramlee, M.S. Emerging pollutant of concern: Occurrence of pharmaceutical compounds in Asia with particular preference to Southeast Asia countries. MATEC Web Conf. 2016, 47, 05026. [Google Scholar] [CrossRef]
- Gibson, R.; Durán-Álvarez, J.C.; Estrada, K.L.; Chávez, A.; Jiménez Cisneros, B. Accumulation and leaching potential of some pharmaceuticals and potential endocrine disruptors in soils irrigated with wastewater in the Tula Valley, Mexico. Chemosphere 2010, 81, 1437–1445. [Google Scholar] [CrossRef]
- González-González, E.; Gómez-Oliván, L.; Galar-Martínez, M.; Vieyra-Reyes, P.; Islas-Flores, H.; García-Medina, S.; Jiménez-Vargas, J.; Razo-Estrada, C.; Pérez-Pastén, R. Metals and nonsteroidal anti-inflammatory pharmaceuticals drugs present in water from Madín Reservoir (Mexico) induce oxidative stress in gill, blood, and muscle of common carp (Cyprinus carpio). Arch. Environ. Contam. Toxicol. 2014, 67, 281–295. [Google Scholar] [CrossRef]
- Siemens, J.; Huschek, G.; Siebe, C.; Kaupenjohann, M. Concentrations and mobility of human pharmaceuticals in the world’s largest wastewater irrigation system, Mexico City-Mezquital Valley. Water Res. 2008, 42, 2124–2134. [Google Scholar] [CrossRef]
- Schwaiger, J.; Ferling, H.; Mallow, U.; Wintermayr, H.; Negele, R.D. Toxic effects of the non-steroidal anti-inflammatory drug diclofenac: Part I: Histopathological alterations and bioaccumulation in rainbow trout. Aquat. Toxicol. 2004, 68, 141–150. [Google Scholar] [CrossRef]
- Sathishkumar, P.; Meena, R.A.A.; Palanisami, T.; Ashokkumar, V.; Palvannan, T.; Gu, F.L. Occurrence, interactive effects, and ecological risk of diclofenac in environmental compartments and biota-a review. Sci. Total Environ. 2020, 698, 134057. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Hong, S.; An, S.A.; Khim, J.S. Methodological advances and future directions of microalgal bioassays for evaluation of potential toxicity in environmental samples: A review. Environ. Int. 2023, 173, 107869. [Google Scholar] [CrossRef] [PubMed]
- Bauer, B.; Mally, A.; Liedtke, D. Zebrafish embryos and larvae as alternative animal models for toxicity testing. Int. J. Mol. Sci. 2021, 22, 13417. [Google Scholar] [CrossRef]
- Nestler, H.; Groh, K.J.; Schönenberger, R.; Behra, R.; Schirmer, K.; Eggen, R.I.; Suter, M.J.F. Multiple-endpoint assay provides a detailed mechanistic view of responses to herbicide exposure in Chlamydomonas reinhardtii. Aquat. Toxicol. 2012, 110, 214–224. [Google Scholar] [CrossRef]
- Martínez-Jerónimo, F.F. Conceptos Generales. In Ensayos Toxicológicos y Métodos de Evaluación de Calidad de Aguas; En Castillo, G., Ed.; IMTA: Jiutepec, Mexico, 2004; pp. 89–98. [Google Scholar]
- Mofeed, J. Effects of three commonly used pharmaceutical products on biochemical parameters of the Micro-alga Pseudokirchneriella subcapitata (Under Laboratory Conditions). Catrina Inter. J. Environ. Sci. 2020, 22, 1–10. [Google Scholar] [CrossRef]
- Udebuani, A.C.; Pereao, O.; Akharame, M.O.; Fatoki, O.S.; Opeolu, B.O. The potential ecological risk of veterinary pharmaceuticals from swine wastewater on freshwater aquatic environment. Water Environ. Res. 2023, 95, e10833. [Google Scholar] [CrossRef]
- Conde-Porcuna, J.M.; Ramos-Rodríguez, E.; Morales-Baquero, R. El zooplancton como integrante de la estructura trófica de los ecosistemas lénticos. Ecosistemas 2004, 13, 23–29. [Google Scholar]
- Nandini, S.; Silva-Briano, M.; García, G.G.; Sarma, S.S.S.; Adabache-Ortiz, A.; de la Rosa, R.G. First record of the temperate species Daphnia curvirostris Eylmann, 1887 emend. Johnson, 1952 (Cladocera: Daphniidae) in Mexico and its demographic characteristics in relation to algal food density. Limnology 2009, 10, 87–94. [Google Scholar] [CrossRef]
- Dalla Bona, M.; Di Leva, V.; De Liguoro, M. The sensitivity of Daphnia magna and Daphnia curvirostris to 10 veterinary antibacterials and to some of their binary mixtures. Chemosphere 2014, 115, 67–74. [Google Scholar] [CrossRef]
- Zhang, C.; Willett, C.; Fremgen, T. Zebrafish: An animal model for toxicological studies. Curr. Protoc. Toxicol. 2003, 17, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Bauer, D.E.; Conforti, V.; Ruiz, L.; Gomez, N. An in situ test to explore the responses of Scenedesmus acutus and Lepocinclis acus as indicators of the changes in water quality in lowland streams. Ecotoxicol. Environ. Saf. 2012, 77, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, P.; Cunha, V.; Oliva-Teles, L.; Ferreira, M.; Guimarães, L. Norfluoxetine and venlafaxine in zebrafish larvae: Single and combined toxicity of two pharmaceutical products relevant for risk assessment. J. Hazard. Mater. 2020, 400, 123171. [Google Scholar] [CrossRef]
- Zhang, K.; Yuan, G.; Werdich, A.A.; Zhao, Y. Ibuprofen and diclofenac impair the cardiovascular development of zebrafish (Danio rerio) at low concentrations. Environ. Pollut. 2020, 258, 113613. [Google Scholar] [CrossRef]
- Porretti, M.; Arrigo, F.; Di Bella, G.; Faggio, C. Impact of pharmaceutical products on zebrafish: An effective tool to assess aquatic pollution. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2022, 261, 109439. [Google Scholar] [CrossRef]
- Stein, J.R. Handbook of Phycological Methods. Culture Methods and Growth Measurements; Cambridge University Press: London, UK, 1973; pp. 7–24. [Google Scholar]
- US Environmental Protection Agency USEPA. Methods for Measuring the Acute Toxicity of Effluents and Receiving Waters to Freshwater and Marine Organism, 5th ed.; EPA-821-R-02-012; Office of Research and Development: Cincinnati, OH, USA, 2002.
- Organisation for Economic Co-Operation and Development (OECD). Test No. 201: Freshwater Alga and Cyanobacteria, Growth Inhibition Test, OECD Guidelines for the Testing of Chemicals, Section 2; OECD Publishing: Paris, France, 2011. [Google Scholar]
- Hernández-Zamora, M.; Martínez-Jerónimo, F. Congo red diversely affects organisms of different trophic levels: A comparative study with microalgae, cladocerans and zebrafish embryos. Environ. Sci. Pollut. Res. Int. 2019, 26, 11743–11755. [Google Scholar] [CrossRef]
- Wellburn, A.R. The spectral determination of chlorophyll a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.T.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Zöllner, N.; Kirsch, K. Über die quantitative Bestimmung von Lipoiden (Mikromethode) mittels der vielen natürlichen Lipoiden (allen bekannten Plasmalipoiden) gemeinsamen Sulfophosphovanillin-Reaktion. Z. Für Die Gesamte Exp. Med. 1962, 135, 545–561. [Google Scholar] [CrossRef]
- Organisation for Economic Co-operation and Development (OECD). Test No. 236: Fish Embryo Acute Toxicity (FET) Test. OECD Guidelines for the Testing of Chemicals; OECD: Paris, France, 2013. [Google Scholar]
- Ferrari, B.; Mons, R.; Vollat, B.; Fraysse, B.; Paxéus, N.; Lo Giudice, R.; Pollio, A.; Garric, J. Environmental risk assessment of six human pharmaceuticals: Are the current environmental risk assessment procedures sufficient for the protection of the aquatic environment? Environ. Toxicol. Chem. 2004, 23, 1344–1354. [Google Scholar] [CrossRef] [PubMed]
- Quinn, B.; Schmidt, W.; O’Rourke, K.; Hernan, R. Effects of the pharmaceuticals gemfibrozil and diclofenac on biomarker expression in the zebra mussel (Dreissena polymorpha) and their comparison with standardised toxicity tests. Chemosphere 2011, 84, 657–663. [Google Scholar] [CrossRef] [PubMed]
- Zind, H.; Mondamert, L.; Remaury, Q.B.; Cleon, A.; Leitner, N.K.V.; Labanowski, J. Occurrence of carbamazepine, diclofenac, and their related metabolites and transformation products in a French aquatic environment and preliminary risk assessment. Water Res. 2021, 196, 117052. [Google Scholar] [CrossRef]
- Zhang, Y.; Guo, J.; Yao, T.; Zhang, Y.; Zhou, X.; Chu, H. The influence of four pharmaceuticals on Chlorella pyrenoidosa culture. Sci. Rep. 2019, 9, 1624. [Google Scholar] [CrossRef]
- Gomaa, M.; Zien-Elabdeen, A.; Hifney, A.F.; Adam, M.S. Phycotoxicity of antibiotics and non-steroidal anti-inflammatory drugs to green algae Chlorella sp. and Desmodesmus spinosus: Assessment of combined toxicity by Box–Behnken experimental design. Environ. Technol. Innov. 2021, 23, 101586. [Google Scholar] [CrossRef]
- Wang, H.; Jin, M.; Mao, W.; Chen, C.; Fu, L.; Li, Z.; Du, S.; Liu, H. Photosynthetic toxicity of non-steroidal anti-inflammatory drugs (NSAIDs) on green algae Scenedesmus obliquus. Sci. Total Environ. 2020, 707, 136176. [Google Scholar] [CrossRef]
- Tang, J.; Wu, Y.; Esquivel-Elizondo, S.; Sørensen, S.J.; Rittmann, B.E. How microbial aggregates protect against nanoparticle toxicity. Trends Biotechnol. 2018, 36, 1171–1182. [Google Scholar] [CrossRef]
- You, X.; Xu, N.; Yang, X.; Sun, W. Pollutants affect algae-bacteria interactions: A critical review. Environ. Pollut. 2021, 276, 116723. [Google Scholar] [CrossRef]
- Zhu, N.; Wang, S.; Tang, C.; Duan, P.; Yao, L.; Tang, J.; Wong, P.K.; Dionnysiou, D.D.; Wu, Y. Protection mechanisms of periphytic biofilm to photocatalytic nanoparticle exposure. Environ. Sci. Technol. 2019, 53, 1585–1594. [Google Scholar] [CrossRef]
- Mou, S.; Xu, D.; Ye, N.; Zhang, X.; Liang, C.; Liang, Q.; Zheng, Z.; Zhuang, Z.; Miao, J. Rapid estimation of lipid content in an Antarctic ice alga (Chlamydomonas sp.) using the lipophilic fluorescent dye BODIPY505/515. J. Appl. Phycol. 2012, 24, 1169–1176. [Google Scholar] [CrossRef]
- Escher, B.; Bramaz, N.; Eggen, R.; Richter, M. In vitro assessment of modes of toxic action of pharmaceuticals in aquatic life. Environ. Sci. Technol. 2005, 39, 3090–3100. [Google Scholar] [CrossRef] [PubMed]
- Corcoll, N.; Acuna, V.; Barcelo, D.; Casellas, M.; Guasch, H.; Huerta, B.; Petrovic, M.; Ponsati, L.; Rodriguez-Mozaz, S.; Sabater, S. Pollution-induced community tolerance to non-steroidal anti-inflammatory drugs (NSAIDs) in fluvial biofilm communities affected by WWTP effluents. Chemosphere 2014, 112, 185–193. [Google Scholar] [CrossRef]
- Liang, S.X.T.; Wong, L.S.; Dhanapal, A.C.T.A.; Djearamane, S. Toxicity of metals and metallic nanoparticles on nutritional properties of microalgae. Water Air Soil. Pollut. 2020, 231, 52. [Google Scholar] [CrossRef]
- Happ, T.; Triebskorn, R.; Köhler, H.R. Acute effects of diclofenac and DMSO to Daphnia magna: Immobilisation and hsp70-induction. Chemosphere 2008, 73, 353–359. [Google Scholar] [CrossRef] [PubMed]
- Junqueira de Castro, F.; Alves dos Santos, D.R.; Picolomini, B.C.R.; Sanzi, C.F.; Brasil, C.R.; Cesar, A. Ecotoxicological assessment of four pharmaceuticals compounds through acute toxicity test. O Mundo Saúde Sao Paulo 2014, 38, 51–55. [Google Scholar] [CrossRef]
- Barbosa, I.R.; Martins, R.M.; Sá e Melo, M.L.; Soares, A.M.V.M. Acute and chronic toxicity of dimethyl sulfoxide to Daphnia magna. Bull. Environ. Contam. Toxicol. 2003, 70, 1264–1268. [Google Scholar] [CrossRef]
- Smith-Jansen, M.; Bartels, P.; Adler, N.; Altenburger, R. Phytotoxicity assessment of diclofenac and its phototransformation products. Anal. Bioanal. Chem. 2007, 387, 1389–1396. [Google Scholar] [CrossRef]
- Lee, J.; Ji, K.; Kho, Y.L.; Kim, P.; Choi, K. Chronic exposure to diclofenac on two freshwater cladocerans and Japanese medaka. Ecotoxicol. Environ. Saf. 2011, 74, 1216–1225. [Google Scholar] [CrossRef]
- Dodson, S.; Hanazato, T. Commentary on effects of anthropogenic and natural organic chemicals on development, swimming behavior, and reproduction of Daphnia, a key member of aquatic ecosystems. Environ. Health. Perspect. 1995, 103, 7–11. [Google Scholar]
- Smirnov, N.N. Physiology of the Cladocera, 2nd ed.; Academic Press: Cambridge, MA, USA, 2017; p. 402. [Google Scholar]
- Edward, D.A.; Chapman, T. Mechanisms underlying costs of reproduction. In Mechanisms of Life History Evolution: The Genetics and Physiology of Life History Traits and Trade-Offs; En Flatt, T., Heyland, A., Eds.; Oxford University Press: Oxford, UK, 2011; pp. 137–152. [Google Scholar]
- Beyers, R.J.; Odum, H.T. Metabolism and Homeostasis. In Ecological Microcosms; Springer Advanced Texts in Life Sciences; Springer: New York, NY, USA, 1993; pp. 11–40. [Google Scholar]
- Liu, Y.; Wang, L.; Pan, B.; Wang, C.; Bao, S.; Nie, X. Toxic effects of diclofenac on life history parameters and the expression of detoxification-related genes in Daphnia magna. Aquat. Toxicol. 2017, 183, 104–113. [Google Scholar] [CrossRef]
- LfW (Landesamt für Wasserwirtschaft). Arzneimittel in der Umwelt. Abschlussbericht des Bayerischen Landesamtes für Wasserwirtschaft zum Forschungs-und Entwicklungsvorhaben 2000–2003; LfW: Augsburg, Germany, 2003. [Google Scholar]
- Le, T.-H.; Lim, E.-S.; Lee, S.-K.; Park, J.-S.; Kim, Y.-H.; Min, J. Toxicity evaluation of verapamil and tramadol based on toxicity assay and expression patterns of Dhb, Vtg, Arnt, CYP4, and CYP314 in Daphnia magna. Environ. Toxicol. 2011, 26, 515–523. [Google Scholar] [CrossRef] [PubMed]
- Arzate-Cárdenas, M.; Martínez-Jerónimo, F. Energy resource reallocation in Daphnia schodleri (Anomopoda: Daphniidae) reproduction induced by exposure to hexavalent chromium. Chemosphere 2012, 87, 326–332. [Google Scholar] [CrossRef] [PubMed]
- Papchenkova, G.A.; Golovanova, I.L.; Shakova, N.V.U. The parameters of reproduction, sizes, and activities of hydrolases in Daphnia magna Straus of successive generations affected by Roundup Herbicide. Inland. Water Biol. 2009, 2, 286–291. [Google Scholar] [CrossRef]
- Calow, P.; Silby, M.M. A physiological basis of population processes: Ecotoxicological implications. Funct. Ecol. 1990, 4, 283–288. [Google Scholar] [CrossRef]
- Sancho, E.; Ferrando, M.D.; Andreu, E. Physiological stress responses of Anguilla anguilla to fenitrothion. J. Environ. Sci. Health B 1996, 31, 87–98. [Google Scholar] [CrossRef]
- Ferain, A.; De Saeyer, N.; Larondelle, Y.; Rees, J.-F.; Debier, C.; De Schamphelaere, K.A.C. Body lipid composition modulates acute cadmium toxicity in Daphnia magna adults and juveniles. Chemosphere 2018, 205, 328–338. [Google Scholar] [CrossRef]
- Arts, M.T.; Wainmann, B.C. Lipids in Freshwater Ecosystems; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012; 319p. [Google Scholar]
- Tessier, A.J.; Goulden, C.E. Estimating food limitation in cladoceran populations. Limnol. Oceanogr. 1982, 27, 707–717. [Google Scholar] [CrossRef]
- Chabchoubi, I.B.; Bouchhima, R.A.; Louhichi, N.; Baanannou, A.; Masmoudi, S.; Hentati, O. Short-term effects of various non-steroidal anti-inflammatory drugs (NSAIDs) on Danio rerio embryos. MethodsX 2023, 11, 102215. [Google Scholar] [CrossRef]
- Praskova, E.; Voslarova, E.; Siroka, Z.; Plhalova, L.; Macova, S.; Marsalek, P.; Pistekova, V.; Svobodova, Z. Assessment of diclofenac LC50 reference values in juvenile and embryonic stages of the zebrafish (Danio rerio). Pol. J. Vet. Sci. 2011, 14, 545–549. [Google Scholar] [CrossRef]
- Van den Brandhof, E.J.; Montforts, M. Fish embryo toxicity of carbamazepine, diclofenac and metoprolol. Ecotoxicol. Environ. Saf. 2010, 73, 1862–1866. [Google Scholar] [CrossRef]
- Dietrich, D.R.; Prietz, A. Fish embryo toxicity and teratogenicity of pharmaceuticals, detergents and pesticides regularly detected in sewage treatment plant effluents and surface waters. Toxicologist 1999, 48, 151. [Google Scholar]
- Hallare, A.V.; Köhler, H.-R.; Triebskorn, R. Developmental toxicity and stress protein responses in zebrafish embryos after exposure to diclofenac and its solvent, DMSO. Chemosphere 2004, 56, 659–666. [Google Scholar] [CrossRef] [PubMed]
- Xia, L.; Zheng, L.; Zhou, J.L. Effects of ibuprofen, diclofenac and paracetamol on hatch and motor behavior in developing zebrafish (Danio rerio). Chemosphere 2017, 182, 416–425. [Google Scholar] [CrossRef]
- Ribeiro, S.; Torres, T.; Martins, R.; Santos, M.M. Toxicity screening of diclofenac, propranolol, sertraline and simvastatin using Danio rerio and Paracentrotus lividus embryo bioassays. Ecotoxicol. Environ. Saf. 2015, 114, 67–74. [Google Scholar] [CrossRef]
- Horie, Y.; Yamagishi, T.; Yagi, A.; Shintaku, Y.; Iguchi, T.; Tatarazako, N. The non-steroidal anti-inflammatory drug diclofenac sodium induces abnormal embryogenesis and delayed lethal effects in early life stage zebrafish (Danio rerio). J. Appl. Toxicol. 2018, 39, 622–629. [Google Scholar] [CrossRef]
- Escapa, C.; Torres, T.; Neuparth, T.; Coimbra, R.N.; García, A.I.; Santos, M.M.; Otero, M. Zebrafissh embryo bioassays for a comprehensive evaluation of microalgae efficiency in the removal of diclofenac from water. Sci. Total Environ. 2018, 640–641, 1024–1033. [Google Scholar] [CrossRef]
- Yadav, P.; Verma, M.; Ahmed, S.; Singh, A.; Yadav, S.; Zahra, K. Risk assessment of diclofenac sodium on Zebra Fish, Danio rerio: Protein estimation in tissues. Res. J. Pharm. Technol. 2019, 12, 4635–4638. [Google Scholar] [CrossRef]
- Zhou, S.; Chen, Q.; Di Paolo, C.; Shao, Y.; Hollert, H.; Seiler, T.B. Behavioral profile alterations in zebrafish larvae exposed to environmentally relevant concentrations of eight priority pharmaceuticals. Sci. Tot. Environ. 2019, 664, 89–98. [Google Scholar] [CrossRef]
- Glaberman, S.; Padilla, S.; Barron, M.G. Evaluating the zebrafish embryo toxicity test for pesticide hazard screening. Environ. Toxicol. Chem. 2017, 36, 1221–1226. [Google Scholar] [CrossRef]
- Bereketoglu, C.; Pradhan, A.; Olsson, P.E. Nonsteroidal anti-inflammatory drugs (NSAIDs) cause male-biased sex differentiation in zebrafish. Aquat. Toxicol. 2020, 223, 105476. [Google Scholar] [CrossRef]
- Tomisato, W.; Tsutsumi, S.; Hoshino, T.; Hwang, H.J.; Mio, M.; Tsuchiya, T.; Mizushima, T. Role of direct cytotoxic effects of NSAIDs in the induction of gastric lesions. Biochem. Pharmacol 2004, 67, 575–585. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.B.; Gao, H.W.; Zhang, Y.L.; Zhang, Y.; Zhou, X.F.; Li, C.Q.; Gao, H.P. Developmental toxicity of diclofenac and elucidation of gene regulation in zebrafish (Danio rerio). Sci. Rep. 2014, 4, 4841. [Google Scholar] [CrossRef] [PubMed]
- Buchan, J.G.; Gray, R.S.; Gansner, J.M.; Alvarado, D.M.; Burgert, L.; Gitlin, J.D.; Gurnett, C.A.; Goldsmith, M.I. Kinesin family member 6 (kif6) is necessary for spine development in zebrafish. Dev. Dyn. 2014, 243, 1646–1657. [Google Scholar] [CrossRef] [PubMed]
- Gray, R.S.; Wilm, T.P.; Smith, J.; Bagnat, M.; Dale, R.M.; Topczewski, J.; Johnson, S.L.; Solnica-Krezel, L. Loss of col8a1a function during zebrafish embryogenesis results in congenital vertebral malformations. Dev. Biol. 2014, 386, 72–85. [Google Scholar] [CrossRef]
- Commission of the European Communities. Technical Guidance Document in Support of Commission Directive 93/67/EEC on Risk Assessment for New Notified Substances and Commission Regulation (EC) No. 1488/94 on Risk Assessment for Existing Substances. Part II: Environmental Risk Assessment; Office for Official Publications of the European Communities: Luxembourg, 1996. [Google Scholar]
- Heberer, T.; Feldmann, D. Contribution of effluents from hospitals and private households to the total loads of diclofenac and carbamazepine in municipal sewage effluents--modeling versus measurements. J. Hazard. Mater. 2005, 122, 211. [Google Scholar] [CrossRef]
- Oviedo-Gómez, D.G.; Galar-Martínez, M.; García-Medina, S.; Razo-Estrada, C.; Gómez-Oliván, L.M. Diclofenac-enriched artificial sediment induces oxidative stress in Hyalella azteca. Environ. Toxicol. Pharmacol. 2010, 29, 39–43. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hernández-Zamora, M.; Cruz-Castillo, L.M.; Martínez-Jerónimo, L.; Martínez-Jerónimo, F. Diclofenac Produces Diverse Toxic Effects on Aquatic Organisms of Different Trophic Levels, Including Microalgae, Cladocerans, and Fish. Water 2025, 17, 1489. https://doi.org/10.3390/w17101489
Hernández-Zamora M, Cruz-Castillo LM, Martínez-Jerónimo L, Martínez-Jerónimo F. Diclofenac Produces Diverse Toxic Effects on Aquatic Organisms of Different Trophic Levels, Including Microalgae, Cladocerans, and Fish. Water. 2025; 17(10):1489. https://doi.org/10.3390/w17101489
Chicago/Turabian StyleHernández-Zamora, Miriam, Laura Mariana Cruz-Castillo, Laura Martínez-Jerónimo, and Fernando Martínez-Jerónimo. 2025. "Diclofenac Produces Diverse Toxic Effects on Aquatic Organisms of Different Trophic Levels, Including Microalgae, Cladocerans, and Fish" Water 17, no. 10: 1489. https://doi.org/10.3390/w17101489
APA StyleHernández-Zamora, M., Cruz-Castillo, L. M., Martínez-Jerónimo, L., & Martínez-Jerónimo, F. (2025). Diclofenac Produces Diverse Toxic Effects on Aquatic Organisms of Different Trophic Levels, Including Microalgae, Cladocerans, and Fish. Water, 17(10), 1489. https://doi.org/10.3390/w17101489