
Exploring the Drivers Influencing Multidimensional Alpha and Beta Diversity of Macroinvertebrates in Mountain Streams



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
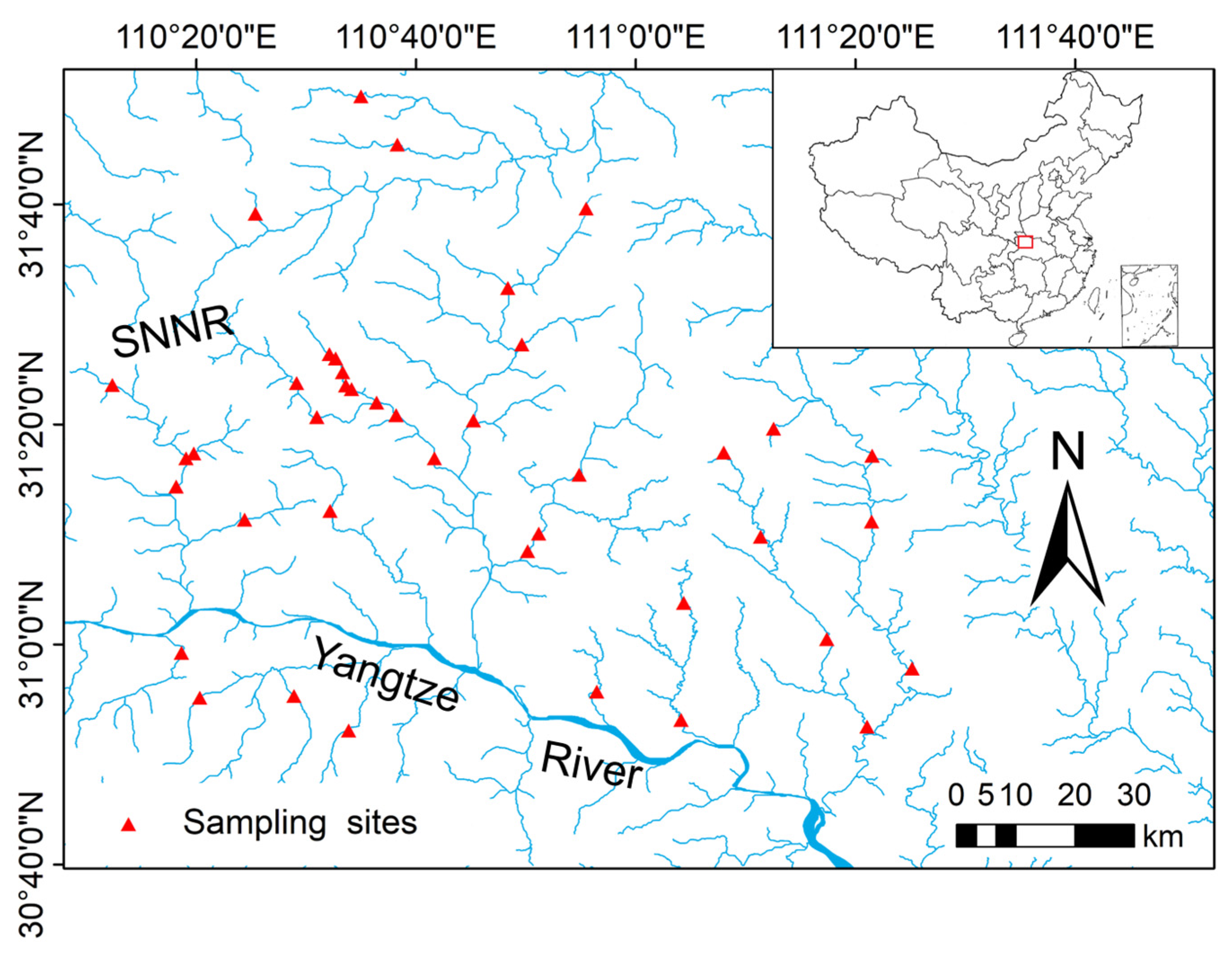
2.1. Study Area
2.2. Macroinvertebrates Sampling
2.3. Functional Traits and Phylogenetic Proxy
2.4. Environmental Factors
2.5. Spatial Factors
2.6. Data Analysis
2.6.1. Alpha Diversity
2.6.2. Beta Diversity
3. Results
3.1. Alpha Diversity
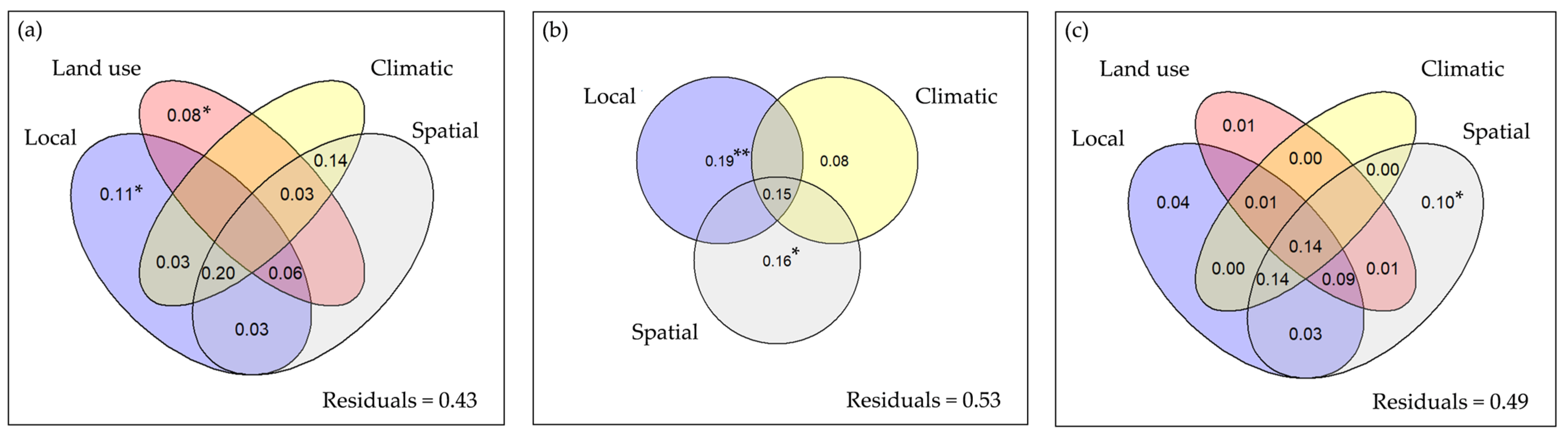
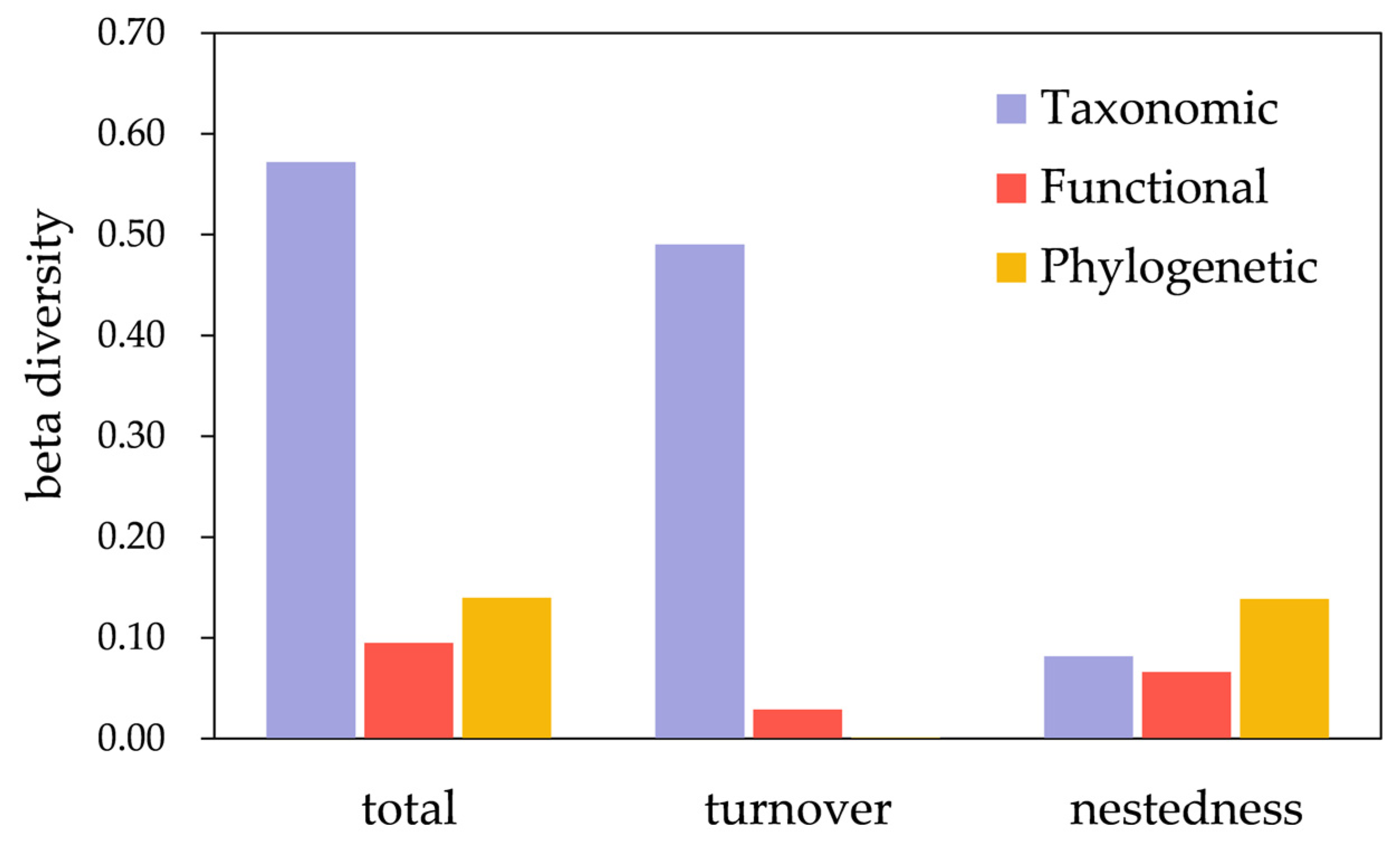
3.2. Beta Diversity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pascual, U.; Adams, W.M.; Díaz, S.; Lele, S.; Mace, G.M.; Turnhout, E. Biodiversity and the challenge of pluralism. Nat. Sustain. 2021, 4, 567–572. [Google Scholar] [CrossRef]
- Tilman, D.; Isbell, F.; Cowles, J.M. Biodiversity and ecosystem functioning. Annu. Rev. Ecol. Evol. Syst. 2014, 45, 471–493. [Google Scholar] [CrossRef]
- Saito, V.S.; Siqueira, T.; Fonseca-Gessner, A.A. Should phylogenetic and functional diversity metrics compose macroinvertebrate multimetric indices for stream biomonitoring? Hydrobiologia 2015, 745, 167–179. [Google Scholar] [CrossRef]
- Ackerly, D.D.; Dudley, S.A.; Sultan, S.E.; Schmitt, J.; Coleman, J.S.; Linder, C.R.; Sandquist, D.R.; Geber, M.A.; Evans, A.S.; Dawson, T.E. The evolution of plant ecophysiological traits: Recent advances and future directions: New research addresses natural selection, genetic constraints, and the adaptive evolution of plant ecophysiological traits. Bioscience 2000, 50, 979–995. [Google Scholar] [CrossRef]
- Colombo, G.T.; Di Ponzio, R.; Benchimol, M.; Peres, C.A.; Bobrowiec, P.E.D. Functional diversity and trait filtering of insectivorous bats on forest islands created by an amazonian mega dam. Funct. Ecol. 2023, 37, 99–111. [Google Scholar] [CrossRef]
- Schmera, D.; Heino, J.; Podani, J.; Erős, T.; Dolédec, S. Functional diversity: A review of methodology and current knowledge in freshwater macroinvertebrate research. Hydrobiologia 2017, 787, 27–44. [Google Scholar] [CrossRef]
- Rangel, T.F.; Edwards, N.R.; Holden, P.B.; Diniz-Filho, J.A.F.; Gosling, W.D.; Coelho, M.T.P.; Cassemiro, F.A.; Rahbek, C.; Colwell, R.K. Modeling the ecology and evolution of biodiversity: Biogeographical cradles, museums, and graves. Science 2018, 361, eaar5452. [Google Scholar] [CrossRef] [PubMed]
- Mascarenhas, R.; Miyaki, C.Y.; Dobrovolski, R.; Batalha-Filho, H. Late pleistocene climate change shapes population divergence of an atlantic forest passerine: A model-based phylogeographic hypothesis test. J. Ornithol. 2019, 160, 733–748. [Google Scholar] [CrossRef]
- Winter, M.; Devictor, V.; Schweiger, O. Phylogenetic diversity and nature conservation: Where are we? Trends Ecol. Evol. 2013, 28, 199–204. [Google Scholar] [CrossRef]
- Alahuhta, J.; Erős, T.; Kärnä, O.-M.; Soininen, J.; Wang, J.; Heino, J. Understanding environmental change through the lens of trait-based, functional, and phylogenetic biodiversity in freshwater ecosystems. Environ. Rev. 2019, 27, 263–273. [Google Scholar] [CrossRef]
- Craven, D.; Eisenhauer, N.; Pearse, W.D.; Hautier, Y.; Isbell, F.; Roscher, C.; Bahn, M.; Beierkuhnlein, C.; Bönisch, G.; Buchmann, N. Multiple facets of biodiversity drive the diversity–stability relationship. Nat. Ecol. Evol. 2018, 2, 1579–1587. [Google Scholar] [CrossRef] [PubMed]
- Southwood, T.R. Habitat, the templet for ecological strategies? J. Anim. Ecol. 1977, 46, 337–365. [Google Scholar] [CrossRef]
- Melles, S.; Jones, N.; Schmidt, B. Review of theoretical developments in stream ecology and their influence on stream classification and conservation planning. Freshw. Biol. 2012, 57, 415–434. [Google Scholar] [CrossRef]
- Leibold, M.A.; Holyoak, M.; Mouquet, N.; Amarasekare, P.; Chase, J.M.; Hoopes, M.F.; Holt, R.D.; Shurin, J.B.; Law, R.; Tilman, D. The metacommunity concept: A framework for multi-scale community ecology. Ecol. Lett. 2004, 7, 601–613. [Google Scholar] [CrossRef]
- Özkundakci, D.; Hamilton, D.P.; Kelly, D.; Schallenberg, M.; de Winton, M.; Verburg, P.; Trolle, D. Ecological integrity of deep lakes in new zealand across anthropogenic pressure gradients. Ecol. Indic. 2014, 37, 45–57. [Google Scholar] [CrossRef]
- Li, Z.; Liu, Z.; Heino, J.; Jiang, X.; Wang, J.; Tang, T.; Xie, Z. Discriminating the effects of local stressors from climatic factors and dispersal processes on multiple biodiversity dimensions of macroinvertebrate communities across subtropical drainage basins. Sci. Total Environ. 2020, 711, 134750. [Google Scholar] [CrossRef]
- Alahuhta, J.; Aroviita, J. Quantifying the relative importance of natural variables, human disturbance and spatial processes in ecological status indicators of boreal lakes. Ecol. Indic. 2016, 63, 240–248. [Google Scholar] [CrossRef]
- Whittaker, R.H. Evolution and measurement of species diversity. Taxon 1972, 21, 213–251. [Google Scholar] [CrossRef]
- Baselga, A. Partitioning the turnover and nestedness components of beta diversity. Glob. Ecol. Biogeogr. 2010, 19, 134–143. [Google Scholar] [CrossRef]
- Legendre, P.; Gauthier, O. Statistical methods for temporal and space–time analysis of community composition data. Proc. R. Soc. B Biol. Sci. 2014, 281, 20132728. [Google Scholar] [CrossRef]
- Stubbington, R.; Sarremejane, R.; Datry, T. Alpha and beta diversity of connected benthic–subsurface invertebrate communities respond to drying in dynamic river ecosystems. Ecography 2019, 42, 2060–2073. [Google Scholar] [CrossRef]
- González-Caro, S.; Umaña, M.N.; Álvarez, E.; Stevenson, P.R.; Swenson, N.G. Phylogenetic alpha and beta diversity in tropical tree assemblages along regional-scale environmental gradients in northwest south America. J. Plant Ecol. 2014, 7, 145–153. [Google Scholar] [CrossRef]
- Li, Z.; García-Girón, J.; Zhang, J.; Jia, Y.; Jiang, X.; Xie, Z. Anthropogenic impacts on multiple facets of macroinvertebrate α and β diversity in a large river-floodplain ecosystem. Sci. Total Environ. 2023, 874, 162387. [Google Scholar] [CrossRef]
- Pool, T.K.; Grenouillet, G.; Villéger, S. Species contribute differently to the taxonomic, functional, and phylogenetic alpha and beta diversity of freshwater fish communities. Divers. Distrib. 2014, 20, 1235–1244. [Google Scholar] [CrossRef]
- Lin, Z.; Liu, G.; Guo, K.; Wang, K.; Wijewardene, L.; Wu, N. Scales matter: Regional environment factors affect α diversity but local factors affect β diversity of macroinvertebrates in thousand islands lake catchment area. Ecol. Indic. 2024, 158, 111561. [Google Scholar] [CrossRef]
- Hill, M.J.; Heino, J.; White, J.C.; Ryves, D.B.; Wood, P.J. Environmental factors are primary determinants of different facets of pond macroinvertebrate alpha and beta diversity in a human-modified landscape. Biol. Conserv. 2019, 237, 348–357. [Google Scholar] [CrossRef]
- Clarke, A.; Mac Nally, R.; Bond, N.; Lake, P.S. Macroinvertebrate diversity in headwater streams: A review. Freshw. Biol. 2008, 53, 1707–1721. [Google Scholar] [CrossRef]
- MacArthur, R.H.; MacArthur, J.W. On bird species diversity. Ecology 1961, 42, 594–598. [Google Scholar] [CrossRef]
- Chen, Z.; Yang, J.; Xie, Z. Economic development of local communities and biodiversity conservation: A case study from shennongjia national nature reserve, China. Biodivers. Conserv. 2005, 14, 2095–2108. [Google Scholar] [CrossRef]
- Lopez-Pujol, J.; Ren, M.-X. Biodiversity and the three gorges reservoir: A troubled marriage. J. Nat. Hist. 2009, 43, 2765–2786. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, N.; Tang, T.; Zhou, S.; Cai, Q. Small run-of-river dams affect taxonomic and functional β-diversity, community assembly process of benthic diatoms. Front. Ecol. Evol. 2022, 10, 895328. [Google Scholar] [CrossRef]
- Dudgeon, D. Tropical Asian Streams: Zoobenthos, Ecology and Conservation; Hong Kong University Press: Hong Kong, China, 1999; Volume 1, ISBN 9622094694. [Google Scholar]
- Morse, J.C.; Yang, L.; Tian, L. Aquatic Insects of China Useful for Monitoring Water Quality; Hohai University Press: Nanjing, China, 1994; ISBN 7563002405. [Google Scholar]
- Zhou, C.; Gui, H.; Zhou, K. Larval key to families of ephemeroptera from china (insecta). J. Nanjing Norm. Univ. 2003, 26, 65–68. [Google Scholar] [CrossRef]
- Poff, N.L.; Olden, J.D.; Vieira, N.K.M.; Finn, D.S.; Simmons, M.P.; Kondratieff, B.C. Functional trait niches of north american lotic insects: Traits-based ecological applications in light of phylogenetic relationships. J. N. Am. Benthol. Soc. 2006, 25, 730–755. [Google Scholar] [CrossRef]
- Li, Z.; Heino, J.; Liu, Z.; Meng, X.; Chen, X.; Ge, Y.; Xie, Z. The drivers of multiple dimensions of stream macroinvertebrate beta diversity across a large montane landscape. Limnol. Oceanogr. 2020, 66, 226–236. [Google Scholar] [CrossRef]
- Tolonen, K.T.; Vilmi, A.; Karjalainen, S.; Hellsten, S.; Heino, J. Do different facets of littoral macroinvertebrate diversity show congruent patterns in a large lake system? Community Ecol. 2017, 18, 109–116. [Google Scholar] [CrossRef]
- Rocha, M.P.; Bini, L.M.; Domisch, S.; Tolonen, K.T.; Jyrkankallio-Mikkola, J.; Soininen, J.; Hjort, J.; Heino, J. Local environment and space drive multiple facets of stream macroinvertebrate beta diversity. J. Biogeogr. 2018, 45, 2744–2754. [Google Scholar] [CrossRef]
- Jiang, X.; Pan, B.; Jiang, W.; Hou, Y.; Yang, H.; Zhu, P.; Heino, J. The role of environmental conditions, climatic factors and spatial processes in driving multiple facets of stream macroinvertebrate beta diversity in a climatically heterogeneous mountain region. Ecol. Indic. 2021, 124, 107407. [Google Scholar] [CrossRef]
- Clarke, K.; Warwick, R. A further biodiversity index applicable to species lists: Variation in taxonomic distinctness. Mar. Ecol. Prog. Ser. 2001, 216, 265–278. [Google Scholar] [CrossRef]
- Heino, J. Functional biodiversity of macroinvertebrate assemblages along major ecological gradients of boreal headwater streams. Freshw. Biol. 2005, 50, 1578–1587. [Google Scholar] [CrossRef]
- Huang, X.; Chen, W.; Cai, Q. Standard Methods for Observation and Analysis in Chinese Ecosystem Research Network-Survey, Observation and Analysis of Lake Ecology; Standards Press of China: Beijing, China, 1999; ISBN 7506621398. [Google Scholar]
- Yang, J.; Huang, X. The 30 m annual land cover dataset and its dynamics in China from 1990 to 2019. Earth Syst. Sci. Data 2021, 13, 3907–3925. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Griffith, D.A.; Peres-Neto, P.R. Spatial modeling in ecology: The flexibility of eigenfunction spatial analyses. Ecology 2006, 87, 2603–2613. [Google Scholar] [CrossRef] [PubMed]
- Borcard, D.; Gillet, F.; Legendre, P. Spatial analysis of ecological data. In Numerical Ecology with R; Springer: New York, NY, USA, 2011; pp. 227–292. [Google Scholar] [CrossRef]
- Declerck, S.A.; Coronel, J.S.; Legendre, P.; Brendonck, L. Scale dependency of processes structuring metacommunities of cladocerans in temporary pools of high-andes wetlands. Ecography 2011, 34, 296–305. [Google Scholar] [CrossRef]
- Laliberté, E.; Legendre, P.; Shipley, B.; Laliberté, M.E. Package ‘fd’: Measuring Functional Diversity from Multiple Traits, and Other Tools for Functional Ecology. Available online: https://rpkg.net/packages/FD/reference/FD-package.ob (accessed on 2 December 2021).
- Oksanen, J.; Blanchet, F.G.; Michael, F.; Kindt, R.; Legendre, P.; Dan, M.; Minchin, P.R.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Package ‘Vegan’. Community Ecology Package, Version 2.5-7. 2020. Available online: https://github.com/vegandevs/vegan (accessed on 5 December 2021).
- Kembel, S.W.; Cowan, P.D.; Helmus, M.R.; Cornwell, W.K.; Morlon, H.; Ackerly, D.D.; Blomberg, S.P.; Webb, C.O. Picante: R tools for integrating phylogenies and ecology. Bioinformatics 2010, 26, 1463–1464. [Google Scholar] [CrossRef]
- Harrell, F.E., Jr. Package ‘Hmisc’. Harrell Miscellaneous, Version 4.6. 2021. Available online: https://hbiostat.org/R/Hmisc/ (accessed on 14 December 2021).
- Ripley, B.; Venables, B.; Bates, D.M.; Hornik, K.; Gebhardt, A.; Firth, D. Package ‘Mass’. Available online: http://www.stats.ox.ac.uk/pub/MASS4/ (accessed on 2 December 2021).
- Peres-Neto, P.R.; Legendre, P.; Dray, S.; Borcard, D. Variation partitioning of species data matrices: Estimation and comparison of fractions. Ecology 2006, 87, 2614–2625. [Google Scholar] [CrossRef]
- Andres, B.; David, O.; Sebastien, V.; Bortoli, D.J.; Fabien, L.; Maxime, L.; Renato, H.-S. Package ‘Betapart’: Partitioning Beta Diversity into Turnover and Nestedness Components. Version1.5.4. 2021. Available online: https://CRAN.R-project.org/package=betapart (accessed on 10 January 2022).
- Nekola, J.C.; White, P.S. The distance decay of similarity in biogeography and ecology. J. Biogeogr. 1999, 26, 867–878. [Google Scholar] [CrossRef]
- Legendre, P.; Anderson, M.J. Distance-based redundancy analysis: Testing multispecies responses in multifactorial ecological experiments. Ecol. Monogr. 1999, 69, 1–24. [Google Scholar] [CrossRef]
- Laliberté, E.; Legendre, P. A distance-based framework for measuring functional diversity from multiple traits. Ecology 2010, 91, 299–305. [Google Scholar] [CrossRef]
- Li, Z.; Jiang, X.; Wang, J.; Meng, X.; Heino, J.; Xie, Z. Multiple facets of stream macroinvertebrate alpha diversity are driven by different ecological factors across an extensive altitudinal gradient. Ecol. Evol. 2019, 9, 1306–1322. [Google Scholar] [CrossRef]
- Bonacina, L.; Fasano, F.; Mezzanotte, V.; Fornaroli, R. Effects of water temperature on freshwater macroinvertebrates: A systematic review. Biol. Rev. 2023, 98, 191–221. [Google Scholar] [CrossRef]
- Guo, F.; Kainz, M.J.; Sheldon, F.; Bunn, S.E. The importance of high-quality algal food sources in stream food webs–current status and future perspectives. Freshw. Biol. 2016, 61, 815–831. [Google Scholar] [CrossRef]
- Heino, J.; Tolonen, K.T. Ecological drivers of multiple facets of beta diversity in a lentic macroinvertebrate metacommunity. Limnol. Oceanogr. 2017, 62, 2431–2444. [Google Scholar] [CrossRef]
- Kefford, B.J.; Nugegoda, D.; Zalizniak, L.; Fields, E.J.; Hassell, K.L. The salinity tolerance of freshwater macroinvertebrate eggs and hatchlings in comparison to their older life-stages: A diversity of responses: The salinity tolerance of freshwater macroinvertebrate eggs and hatchlings. Aquat. Ecol. 2007, 41, 335–348. [Google Scholar] [CrossRef]
- Heino, J.; Melo, A.S.; Siqueira, T.; Soininen, J.; Valanko, S.; Bini, L.M. Metacommunity organisation, spatial extent and dispersal in aquatic systems: Patterns, processes and prospects. Freshw. Biol. 2015, 60, 845–869. [Google Scholar] [CrossRef]
- Shanks, A.L. Mechanisms of cross-shelf dispersal of larval invertebrates and fish. In Ecology of Marine Invertebrate Larvae, CRC Press: Boca Raton, FL, USA, 2020; pp. 323–367. [CrossRef]
- Burgmer, T.; Hillebrand, H.; Pfenninger, M. Effects of climate-driven temperature changes on the diversity of freshwater macroinvertebrates. Oecologia 2007, 151, 93–103. [Google Scholar] [CrossRef]
- Hering, D.; Schmidt-Kloiber, A.; Murphy, J.; Lücke, S.; Zamora-Munoz, C.; López-Rodríguez, M.J.; Huber, T.; Graf, W. Potential impact of climate change on aquatic insects: A sensitivity analysis for european caddisflies (trichoptera) based on distribution patterns and ecological preferences. Aquat. Sci. 2009, 71, 3–14. [Google Scholar] [CrossRef]
- Olson, A.R.; Stewart, T.W.; Thompson, J.R. Direct and indirect effects of human population density and land use on physical features and invertebrates of iowa (USA) streams. Urban Ecosyst. 2016, 19, 159–180. [Google Scholar] [CrossRef]
- Poff, N.L.; Pyne, M.I.; Bledsoe, B.P.; Cuhaciyan, C.C.; Carlisle, D.M. Developing linkages between species traits and multiscaled environmental variation to explore vulnerability of stream benthic communities to climate change. J. N. Am. Benthol. Soc. 2010, 29, 1441–1458. [Google Scholar] [CrossRef]
- Dray, S.; Pélissier, R.; Couteron, P.; Fortin, M.-J.; Legendre, P.; Peres-Neto, P.R.; Bellier, E.; Bivand, R.; Blanchet, F.G.; De Cáceres, M. Community ecology in the age of multivariate multiscale spatial analysis. Ecol. Monogr. 2012, 82, 257–275. [Google Scholar] [CrossRef]
- Krynak, E.M.; Lindo, Z.; Yates, A.G. Patterns and drivers of stream benthic macroinvertebrate beta diversity in an agricultural landscape. Hydrobiologia 2019, 837, 61–75. [Google Scholar] [CrossRef]
- Maasri, A.; Schechner, A.E.; Erdenee, B.; Dodds, W.K.; Chandra, S.; Gelhaus, J.K.; Thorp, J.H. Does diel variation in oxygen influence taxonomic and functional diversity of stream macroinvertebrates? Freshw. Sci. 2019, 38, 692–701. [Google Scholar] [CrossRef]
- Connolly, N.; Crossland, M.; Pearson, R. Effect of low dissolved oxygen on survival, emergence, and drift of tropical stream macroinvertebrates. J. N. Am. Benthol. Soc. 2004, 23, 251–270. [Google Scholar] [CrossRef]
- Schneider, S.C.; Petrin, Z. Effects of flow regime on benthic algae and macroinvertebrates-a comparison between regulated and unregulated rivers. Sci. Total Environ. 2017, 579, 1059–1072. [Google Scholar] [CrossRef] [PubMed]
- Wu, N.; Guo, K.; Zou, Y.; He, F.; Riis, T. Ser: An r package to characterize environmental regimes. Ecol. Evol. 2023, 13, e9882. [Google Scholar] [CrossRef]
- Oikonomou, A.; Stefanidis, K. A- and β-diversity patterns of macrophytes and freshwater fishes are driven by different factors and processes in lakes of the unexplored southern balkan biodiversity hotspot. Water 2020, 12, 1984. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.; Wang, X.; Tan, L.; Cai, Q. Exploring the Drivers Influencing Multidimensional Alpha and Beta Diversity of Macroinvertebrates in Mountain Streams. Water 2024, 16, 2915. https://doi.org/10.3390/w16202915
Li S, Wang X, Tan L, Cai Q. Exploring the Drivers Influencing Multidimensional Alpha and Beta Diversity of Macroinvertebrates in Mountain Streams. Water. 2024; 16(20):2915. https://doi.org/10.3390/w16202915
Chicago/Turabian StyleLi, Shudan, Xingzhong Wang, Lu Tan, and Qinghua Cai. 2024. "Exploring the Drivers Influencing Multidimensional Alpha and Beta Diversity of Macroinvertebrates in Mountain Streams" Water 16, no. 20: 2915. https://doi.org/10.3390/w16202915
APA StyleLi, S., Wang, X., Tan, L., & Cai, Q. (2024). Exploring the Drivers Influencing Multidimensional Alpha and Beta Diversity of Macroinvertebrates in Mountain Streams. Water, 16(20), 2915. https://doi.org/10.3390/w16202915