
Marine Copepods as a Microbiome Hotspot: Revealing Their Interactions and Biotechnological Applications

 ,
,  , , , and
, , , and
Abstract
1. Introduction
2. Characterization of the Copepod-Associated Microbiome
3. Copepod-Microbiome Association
3.1. Copepod-Bacteria Interactions
3.1.1. Endobiotic and Exobiotic
3.1.2. Pathogen Bacteria Associated with Copepods
3.2. Copepod-Fungi Interactions
3.3. Copepod-Virus Interactions
3.4. Copepod-Protist Interactions

{kind=link}
| Copepod Species | Bacteria/Fungi | Habitat | Citation |
|---|---|---|---|
| Centropagidae and Clausocalanidae | Bacilli and Actinobacteria | Marine | [38] |
| Pleuromamma spp. | Lactobacillales, Bacillales | Marine | [44] |
| Acartia sp., Temora longicornis | Actinobacteria, Firmicutes | Marine | [46] |
| Pleuromamma, Undinula | Vibrionaceae, Oceanospirillales, and Rhodobacteraceae | Marine | [49] |
| Acartia tonsa | Unknown | Marine | [64] |
| Calanus hyperboreus and C. glacialis | Unknown | Marine | [65] |
| Undinula vulgaris, Pleuromamma spp. | Vibrio spp. | Marine | [66] |
| Calanus sp. | Bacteroidetes, Proteobacteria, Actinobacteria | Marine | [67] |
| Temora stylifera | Pseudomonas | Marine | [68] |
| Acartia tonsa | Bacillus spp. | Marine | [69] |
| Acartia bifilosa, Eurytemora affinis | Deltaproteobacteria, Firmicutes | Marine | [74] |
| Acartia tonsa | Vibrio cholerae and Pseudomonas sp. | Marine | [77] |
| Lepeophtheirus salmonis | Aeromonas salmonicida | Marine | [111] |
| Acartia omorii | Unknown | Marine | [112] |
| Pleuromamma, Undinula, and Sapphirina, among others | Gammaproteobacterial | Marine | [113] |
| Acartia tonsa | Vibrio cholerae | Marine | [114] |
| Acartia spp. | Unknown | Marine/freshwater | [71] |
| Mesocyclops aspericornis | Wolbachia | Freshwater | [60] |
| Diaptomus spp. | Pseudomonas, Bacillus, Acinetobacter, and Flavobacterium | Freshwater | [72] |
| Eurytemora affinis | Vibrio sp. F10 9ZB36 | Freshwater | [83] |
| Calanus | Dikarya | Marine | [115] |
| Diaptomus novamexicanus | Lagenidium (Phycomycetes) | Freshwater | [92] |
| Boeckella dilatate, Diaptomus gracilis | Aphanomyces ovidestruens | Freshwater | [93] |
| Boeckella dilatata | Aphanomyces sp. | Freshwater | [94] |
| Eudiaptomus gracilis | Aphanomyces sp. | Freshwater | [95] |
| Eudiaptomus intermedius | Aphanomyces sp. | Freshwater | [96] |
| Unknown | Chytridiomycetes, Oomycetes, and Peronosporales | Freshwater | [97] |
| Cyclops fuscus, Cyclops vicinus | Chytridiomycetes, Hyphochytriomycetes, and Oomycetes | Freshwater | [98] |
4. Factors Influencing Copepod-Associated Microbiome
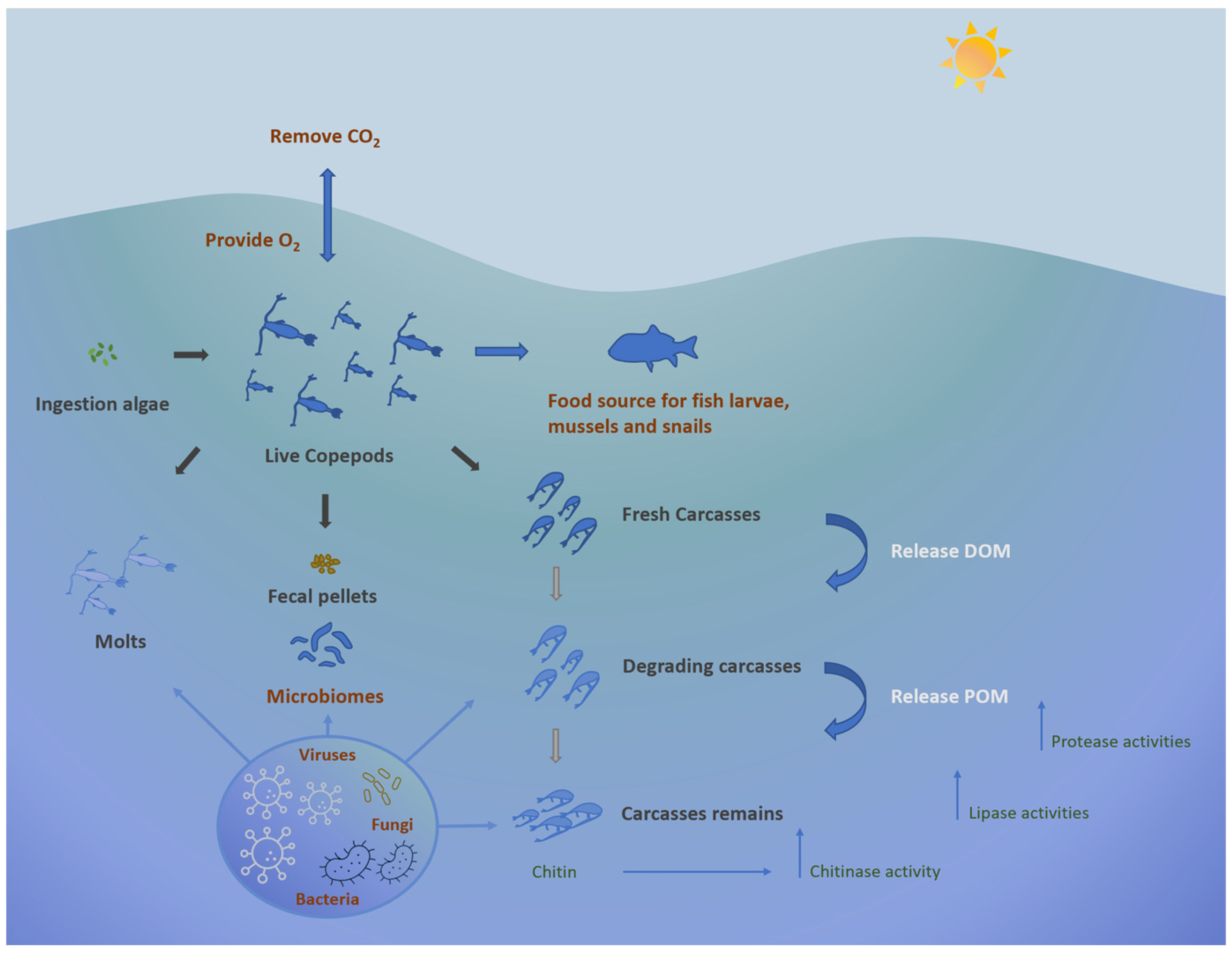
5. Biodegradation Role of the Copepod-Associated Microbiome
6. Conclusions and Future Perspectives
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Mauchline, J.; Blaxter, J.H.S.; Douglas, B.; Tyler, P.A. The Biology of Calanoid Copepods, Introduction; Academic Press: Cambridge, MA, USA, 1998; pp. 1–13. [Google Scholar]
- Ho, J.S. Phylogenetic analysis of copepod orders. J. Crustac. Biol. 1990, 10, 528–536. [Google Scholar] [CrossRef]
- Barbier, E.B. Marine ecosystem services. Curr. Biol. 2017, 27, R507–R510. [Google Scholar] [CrossRef] [PubMed]
- Boxshall, G.A.; Halsey, S.H. An Introduction to Copepod Diversity; Ray Society: Andover, UK, 2004. [Google Scholar]
- Hammervold, S.H.; Glud, R.N.; Evjemo, J.O.; Hagemann, A.; Hansen, B.W. A new large egg type from the marine live feed calanoid copepod Acartia tonsa (Dana) perspectives for selective breeding of designer feed for hatcheries. Aquaculture 2015, 436, 114–120. [Google Scholar] [CrossRef]
- Verity, P.; Smetacek, V. Organism life cycles, predation, and the structure of marine pelagic ecosystems. Mar. Ecol. Prog. Ser. 1996, 130, 277–293. [Google Scholar] [CrossRef]
- Walter, T.C.; Boxshall, G. World of Copepods Database. 2021. Available online: http://www.marinespecies.org/copepoda (accessed on 2 January 2023).
- Reid, J.W.; Williamson, C.E. Copepoda. In Ecology and Classification of North American Freshwater Invertebrates; Academic Press: Cambridge, MA, USA, 2010; pp. 829–899. [Google Scholar] [CrossRef]
- Bass, D.; Rueckert, S.; Stern, R.; Cleary, A.C.; Taylor, J.D.; Ward, G.M.; Huys, R. Parasites, pathogens, and other symbionts of copepods. Trends Parasitol. 2021, 37, 875–889. [Google Scholar] [CrossRef] [PubMed]
- Huys, R.; Boxshall, G.A. (Eds.) Copepod Evolution; Ray Society: London, UK, 1991. [Google Scholar]
- González, M.T.; Castro, R.; Muñoz, G.; López, Z. Sea lice (Siphonostomatoida: Caligidae) diversity on littoral fishes from the southeastern Pacific coast determined from morphology and molecular analysis, with description of a new species (Lepeophtheirus Confusum). Parasitol. Int. 2016, 65, 685–695. [Google Scholar] [CrossRef]
- Azam, F.; Fenchel, T.; Field, J.G.; Gray, J.S.; Meyer-Reil, L.; Thingstad, F. The ecological role of water-column microbes in the sea. Mar. Ecol. Prog. Ser. 1983, 10, 257–263. [Google Scholar] [CrossRef]
- Turner, J.T. The importance of small planktonic copepods and their roles in pelagic marine food webs. Zool. Stud. 2004, 43, 255–266. [Google Scholar]
- Thor, P.; Dam, H.G.; Rogers, D.R. Fate of organic carbon released from decomposing copepod fecal pellets in relation to bacterial production and ectoenzymatic activity. Aquat. Microb. Ecol. 2003, 33, 279–288. [Google Scholar] [CrossRef]
- Møller, E.F.; Thor, P.; Nielsen, T.G. Production of DOC by Calanus finmarchicus, C. glacialis, and C. hyperboreus through sloppy feeding and leakage from fecal pellets. Mar. Ecol. Prog. Ser. 2003, 262, 185–191. [Google Scholar] [CrossRef]
- Melianawati, R.; Rarastoeti, P.; Nyoman, P.; Pudji, A. The effect of various kinds of live feeds on the digestive enzyme activity of coral trout Plectropomus leopardus (Lacepède, 1802) larvae. Int. J. Fish. Aquat. Stud. 2015, 3, 83–88. [Google Scholar]
- Liu, G.; Xu, D. Effects of calanoid copepod Schmackeria poplesia as a live food on the growth, survival and fatty acid composition of larvae and juveniles of Japanese flounder, Paralichthys olivaceus. J. Ocean Univ. China 2009, 8, 359–365. [Google Scholar] [CrossRef]
- Karlsen, Ø.; van der Meeren, T.; Rønnestad, I.; Mangor-Jensen, A.; Galloway, T.F.; Kjørsvik, E.; Hamre, K. Copepods enhance nutritional status, growth and development in Atlantic cod (Gadus morhua L.) larvae—Can we identify the underlying factors? PeerJ 2015, 3, e902. [Google Scholar] [CrossRef] [PubMed]
- Buttino, I.; Ianora, A.; Buono, S.; Vitiello, V.; Malzone, M.G.; Rico, C.; Langellotti, A.L.; Sansone, G.; Gennari, L.; Miralto, A. Experimental cultivation of the Mediterranean calanoid copepods Temora stylifera and Centropages typicus in a pilot re-circulating system. Aquacult. Res. 2012, 43, 247–259. [Google Scholar] [CrossRef]
- Olivotto, I.; Avella, A.M.; Buttino, I.; Borroni, M.; Cutignano, A.; Carnevali, O. Calanoid copepod administration improves yellowtail clownfish (Amphiprion clarkii) larviculture: Biochemical and molecular implications. Aquac. Aquar. Conserv. Legis. Int. J. Bioflux Soc. 2009, 2, 355–367. [Google Scholar]
- Drillet, G.; Frouël, S.; Sichlau, M.H.; Jepsen, P.M.; Højgaard, J.K.; Joarder, A.K.; Hansen, B.W. Status and recommendations on marine copepod cultivation for use as live feed. Aquaculture 2011, 315, 155–166. [Google Scholar] [CrossRef]
- Hansen, B.W. Advances using copepods in aquaculture. J. Plankton Res. 2017, 39, 972–974. [Google Scholar] [CrossRef]
- Rong, R.L.; Gagnat, M.R.; Attramadal, Y.; Vadstein, O. Microbial water quality of the copepod Acartia tonsa in cultures for use as live feed. Aquaculture 2022, 560, 378439. [Google Scholar] [CrossRef]
- Krueger, D.M.; Gustafson, R.G.; Cavanaugh, C.M. Vertical transmission of chemoautotrophic symbionts in the bivalve Solemya velum (Bivalvia: Protobranchia). Biol. Bull. 1996, 190, 195–202. [Google Scholar] [CrossRef]
- Nuester, J.; Shema, S.; Vermont, A.; Fields, D.M.; Twining, B.S. The regeneration of highly bioavailable iron by meso- and microzooplankton. Limnol. Oceanogr. 2014, 59, 1399–1409. [Google Scholar] [CrossRef]
- Schmidt, K.; Schlosser, C.; Atkinson, A.; Fielding, S.; Venables, H.J.; Waluda, C.M.; Achterberg, E.P. Zooplankton gut passage mobilizes lithogenic iron for ocean productivity. Curr. Biol. 2016, 26, 2667–2673. [Google Scholar] [CrossRef] [PubMed]
- Gorokhova, E.; El-Shehawy, R.; Lehtiniemi, M.; Garbaras, A. How Copepods Can Eat Toxins Without Getting Sick: Gut Bacteria Help Zooplankton to Feed in Cyanobacteria Blooms. Front. Microbiol. 2021, 11, 589816. [Google Scholar] [CrossRef] [PubMed]
- Glud, R.N.; Grossart, H.P.; Larsen, M.; Tang, K.W.; Arendt, K.E.; Rysgaard, S. Copepod carcasses as microbial hot spots for pelagic denitrification. Limnol. Oceanogr. 2015, 60, 2026–2036. [Google Scholar] [CrossRef]
- Stief, P.; Lundgaard, A.B.; Morales-Ramirez, A.; Thamdrup, B.; Glud, R.N. Fixed-nitrogen loss associated with sinking zooplankton carcasses in a coastal oxygen minimum zone (Golfo Dulce, Costa Rica). Front. Mar. Sci. 2017, 4, 152. [Google Scholar] [CrossRef]
- Orphan, V.J.; Taylor, L.T.; Hafenbradl, D.; Delong, E. Culture-dependent and culture-independent characterization of microbial assemblages associated with high-temperature petroleum reservoirs. Appl. Environ. Microbiol. 2000, 66, 700–711. [Google Scholar] [CrossRef] [PubMed]
- Joint, I.; Mühling, M.; Querellou, J. Culturing marine bacteria—An essential prerequisite for biodiscovery. Microb. Biotechnol. 2010, 3, 564–575. [Google Scholar] [CrossRef] [PubMed]
- Doytchinov, V.V.; Dimov, S.G. Microbial community composition of the Antarctic ecosystems: Review of the bacteria, fungi, and archaea identified through an NGS-based metagenomics approach. Life 2022, 12, 916. [Google Scholar] [CrossRef] [PubMed]
- Su, C.; Lei, L.; Duan, Y.; Zhang, K.Q.; Yang, J. Culture-independent methods for studying environmental microorganisms: Methods, application, and perspective. Appl. Microbiol. Biotechnol. 2012, 93, 993–1003. [Google Scholar] [CrossRef]
- Liu, S.; Moon, C.D.; Zheng, N.; Huws, S.; Zhao, S.; Wang, J. Opportunities and challenges of using metagenomic data to bring uncultured microbes into cultivation. Microbiome 2022, 10, 76. [Google Scholar] [CrossRef]
- Akaçin, İ.; Ersoy, Ş.; Doluca, O.; Güngörmüşler, M. Comparing the significance of the utilization of next generation and third generation sequencing technologies in microbial metagenomics. Microbiol. Res. 2022, 264, 127154. [Google Scholar] [CrossRef]
- Ciuffreda, L.; Rodríguez-Pérez, H.; Flores, C. Nanopore sequencing and its application to the study of microbial communities. Comput. Struct. Biotechnol. J. 2021, 19, 1497–1511. [Google Scholar] [CrossRef] [PubMed]
- Ansorge, W.J. Next-generation DNA sequencing techniques. New Biotechnol. 2009, 25, 195–203. [Google Scholar] [CrossRef]
- De Corte, D.; Lekunberri, I.; Sintes, E.; Garcia, J.A.L.; Gonzales, S.; Herndl, G.J. Linkage between copepods and bacteria in the North Atlantic Ocean. Aquat. Microb. Ecol. 2014, 72, 215–225. [Google Scholar] [CrossRef]
- Moisander, P.H.; Sexton, A.D.; Daley, M.C. Stable associations masked by temporal variability in the marine copepod microbiome. PLoS ONE 2015, 10, e0138967. [Google Scholar] [CrossRef] [PubMed]
- Dorosz, J.N.A.; Castro-Mejia, J.L.; Hansen, L.H.; Nielsen, D.S.; Skovgaard, A. Different microbiomes associated with the copepods Acartia tonsa and Temora longicornis from the same marine environment. Aquat. Microb. Ecol. 2020, 78, 1–9. [Google Scholar] [CrossRef]
- Ortiz-Estrada, Á.M.; Gollas-Galván, T.; Martínez-Córdova, L.R.; Martínez-Porchas, M. Predictive functional profiles using metagenomic 16S rRNA data: A novel approach to understanding the microbial ecology of aquaculture systems. Rev. Aquac. 2019, 11, 234–245. [Google Scholar] [CrossRef]
- Meyer, M.; Kircher, M. Illumina sequencing library preparation for highly multiplexed target capture and sequencing. Cold Spring Harb. Protoc. 2010, 2010, t5448. [Google Scholar] [CrossRef] [PubMed]
- Mardis, E.R. The impact of next-generation sequencing technology on genetics. Trends Genet. 2008, 24, 133–141. [Google Scholar] [CrossRef]
- Shoemaker, K.M.; Moisander, P.H. Seasonal variation in the copepod gut microbiome in the subtropical North Atlantic Ocean. Environ. Microbiol. 2017, 19, 3087–3097. [Google Scholar] [CrossRef]
- Yeh, H.D.; Questel, J.M.; Maas, K.R.; Bucklin, A. Metabarcoding analysis of regional variation in gut contents of the copepod Calanus finmarchicus in the North Atlantic Ocean. Deep Sea Res. Part II Top. Stud. Oceanogr. 2020, 180, 104738. [Google Scholar] [CrossRef]
- Gerdts, G.; Brandt, P.; Kreisel, K.; Boersma, M.; Schoo, K.L.; Wichels, A. The microbiome of North Sea copepods. Helgol. Mar. Res. 2013, 67, 757–773. [Google Scholar] [CrossRef][Green Version]
- Bickel, S.L.; Tang, K.W.; Grossart, H.P. Structure and function of zooplankton-associated bacterial communities in a temperate estuary change more with time than with zooplankton species. Aquat. Microb. Ecol. 2014, 72, 1–15. [Google Scholar] [CrossRef]
- Zidour, M.; Chevalier, M.; Belguesmia, Y.; Cudennec, B.; Grard, T.; Drider, D.; Souissi, S.; Flahaut, C. Isolation and Characterization of Bacteria Colonizing Acartia tonsa Copepod Eggs and Displaying Antagonist Effects against Vibrio anguillarum, Vibrio alginolyticus, and Other Pathogenic Strains. Front. Microbiol. 2017, 8, 1919. [Google Scholar] [CrossRef] [PubMed]
- Shoemaker, K.M.; Duhamel, S.; Moisander, P.H. Copepods promote bacterial community changes in surrounding seawater through farming and nutrient enrichment. Environ. Microbiol. 2019, 21, 3737–3750. [Google Scholar] [CrossRef]
- Chae, Y.J.; Oh, H.J.; Chang, K.H.; Kwak, I.S.; Jo, H. Application of Next-Generation Sequencing for the Determination of the Bacterial Community in the Gut Contents of Brackish Copepod Species (Acartia hudsonica, Sinocalanus tenellus, and Pseudodiaptomus inopinus). Animals 2021, 11, 542. [Google Scholar] [CrossRef]
- Tang, K.W.; Turk, V.; Grossart, H.P. Linkage between crustacean zooplankton and aquatic bacteria. Aquat. Microb. Ecol. 2010, 61, 261–277. [Google Scholar] [CrossRef]
- Peduzzi, P.; Herndl, G.J. Zooplankton activity fueling the microbial loop: Differential growth response of bacteria from oligotrophic and eutrophic waters. Limnol. Oceanogr. 1992, 37, 1087–1092. [Google Scholar] [CrossRef]
- Datta, M.S.; Almada, A.A.; Baumgartner, M.F.; Mincer, T.J.; Tarrant, A.M.; Polz, M.F. Inter-individual variability in copepod microbiomes reveals bacterial networks linked to host physiology. ISME J. 2018, 12, 2103–2113. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Matson, P.A. Nutrient cycling and biogeochemistry. Princet. Guide Ecol. 2012, 1, 330–339. [Google Scholar]
- Steinberg, D.K.; Landry, M.R. Zooplankton and the ocean carbon cycle. Annu. Rev. Mar. Sci. 2017, 9, 413–444. [Google Scholar] [CrossRef]
- Gadd, G.M. Microbial roles in mineral transformations and metal cycling in the Earth’s critical zone. In Molecular Environmental Soil Science; Springer: Dordrecht, The Netherlands, 2013; pp. 115–165. [Google Scholar]
- Kappler, A.; Bryce, C.; Mansor, M.; Lueder, U.; Byrne, J.M.; Swanner, E.D. An evolving view on biogeochemical cycling of iron. Nat. Rev. Microbiol. 2021, 19, 360–374. [Google Scholar] [CrossRef] [PubMed]
- Gaur, N.; Narasimhulu, K.; PydiSetty, Y. Recent advances in the bio-remediation of persistent organic pollutants and its effect on the environment. J. Clean. Prod. 2018, 198, 1602–1631. [Google Scholar] [CrossRef]
- Stock, W.; Callens, M.; Houwenhuyse, S.; Schols, R.; Goel, N.; Coone, M.; Decaestecker, E. Human impact on symbioses between aquatic organisms and microbes. Aquat. Microb. Ecol. 2021, 87, 113–138. [Google Scholar] [CrossRef]
- Wiwatanaratanabutr, I. Distribution, diversity, and density of Wolbachial infections in cladocerans and copepods from Thailand. J. Invertebr. Pathol. 2013, 114, 341–345. [Google Scholar] [CrossRef] [PubMed]
- Zoccarato, L.; Grossart, H.P. Relationship between lifestyle and structure of bacterial communities and their functionality in aquatic systems. In The Structure and Function of Aquatic Microbial Communities; Springer: Cham, Switzerland, 2019; pp. 13–52. [Google Scholar]
- Carman, K.R.; Dobbs, F.C. Epibiotic microorganisms on copepods and other marine crustaceans. Microsc. Res. Tech. 1997, 37, 116–135. [Google Scholar] [CrossRef]
- Pérez-Brocal, V.; Latorre, A.; Moya, A. Symbionts and pathogens: What is the difference? In Between Pathogenicity and Commensalism, Current Topics in Microbiology and Immunology; Springer: Berlin/Heidelberg, Germany, 2011; pp. 215–243. [Google Scholar]
- Tang, K.W. Copepods as microbial hotspots in the ocean: Effects of host feeding activities on attached bacteria. Aquat. Microb. Ecol. 2005, 38, 31–40. [Google Scholar] [CrossRef]
- Tang, K.W.; Glud, R.N.; Glud, A.; Rysgaard, S.; Nielsen, T.G. Copepod guts as biogeochemical hotspots in the sea: Evidence from microelectrode profiling of Calanus spp. Limnol. Oceanogr. 2011, 56, 666–672. [Google Scholar] [CrossRef]
- Shoemaker, K.M.; Moisander, P.H. Microbial diversity associated with copepods in the North Atlantic subtropical gyre. FEMS Microbiol. Ecol. 2015, 91, fiv064. [Google Scholar] [CrossRef]
- De Corte, D.; Srivastava, A.; Koski, M.; Garcia, J.A.L.; Takaki, Y.; Yokokawa, T.; Nunoura, T.; Elisabeth, N.H.; Sintes, E.; Herndl, G.J. Metagenomic insights into zooplankton-associated bacterial communities. Environ. Microbiol. 2018, 20, 492–505. [Google Scholar] [CrossRef]
- Delille, D.; Razouls, S. Community structures of heterotrophic bacteria of copepod fecal pellets. J. Plankton Res. 1994, 16, 603–615. [Google Scholar] [CrossRef]
- Hansen, B.; Bech, G. Bacteria associated with a marine planktonic copepod in culture. I. Bacterial genera in seawater, body surface, intestines and fecal pellets and succession during fecal pellet degradation. J. Plankton Res. 1996, 18, 257–273. [Google Scholar] [CrossRef]
- Midya, S.; Ganguly, R.K.; Islam, S.S.; Chakraborty, S.K. Attachment and antimicrobial susceptibility of bacterial associates of zooplanktonic copepod: Lesson for environmental safety. Ecotoxicol. Environ. Saf. 2019, 180, 749–755. [Google Scholar] [CrossRef] [PubMed]
- Nagasawa, S. Bacterial epibionts of copepods. Sci. Prog. 1989, 73, 169–176. Available online: http://www.jstor.org/stable/43421029 (accessed on 29 November 2022).
- Holland, R.; Hergenrader, R. Bacterial epibionts of diaptomid copepods. Trans. Am. Microsc. Soc. 1981, 100, 56–65. [Google Scholar] [CrossRef]
- De Angelis, M.A.; Lee, C. Methane production during zooplankton grazing on marine-phytoplankton. Limnol. Oceanogr. 1994, 39, 1298–1308. [Google Scholar] [CrossRef]
- Gorokhova, E.; Soerensen, A.L.; Motwani, N.H. Mercury-methylating bacteria are associated with copepods: A proof-of-principle survey in the Baltic Sea. PLoS ONE 2020, 15, e0230310. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, S.G.; Ahmad, A.; Azam, F. Fate of particle-bound bacteria ingested by Calanus pacificus. Mar. Ecol. Prog. Ser. 1993, 97, 299–307. [Google Scholar] [CrossRef]
- Kaneko, T.; Colwell, R.R. Adsorption of Vibrio parahaemolyticus onto chitin and copepods. Appl. Environ. Microbiol. 1975, 29, 269–274. [Google Scholar] [CrossRef]
- Huq, A.; Small, E.B.; West, P.A.; Huq, M.I.; Rahman, R.; Colwell, R.R. Ecological relationships between Vibrio cholerae and planktonic crustacean copepods. Appl. Environ. Microbiol. 1983, 45, 275–283. [Google Scholar] [CrossRef]
- Huq, A.; West, P.A.; Small, E.B.; Huq, M.I.; Colwell, R.R. Influence of water temperature, salinity, and pH on survival and growth of toxigenic Vibrio cholerae serovar O1 associated with live copepods in laboratory microcosms. Appl. Environ. Microbiol. 1984, 48, 420–424. [Google Scholar] [CrossRef]
- Araújo, D.B.; Martins, S.C.S.; Albuquerque, L.M.B.D.; Hofer, E. Influence of the copepod Mesocyclops longisetus (Crustacea: Cyclopidae) on the survival of Vibrio cholerae O1 in fresh water. Cad. Saúde Pública 1996, 12, 551–554. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Thomas, K.U.; Joseph, N.; Raveendran, O.; Nair, S. Salinity-induced survival strategy of Vibrio cholerae associated with copepods in Cochin backwaters. Mar. Pollut. Bull. 2006, 52, 1425–1430. [Google Scholar] [CrossRef] [PubMed]
- Rawlings, T.K.; Ruiz, G.M.; Colwell, R.R. Association of Vibrio cholerae O1 El Tor and O139 Bengal with the Copepods Acartia tonsa and Eurytemora affinis. Appl. Environ. Microbiol. 2007, 73, 7926–7933. [Google Scholar] [CrossRef] [PubMed]
- De Magny, G.C.; Mozumder, P.K.; Grim, C.J.; Hasan, N.A.; Naser, M.N.; Alam, M.; Sack, R.B.; Huq, A.; Colwell, R.R. Role of zooplankton diversity in Vibrio cholerae population dynamics and in the incidence of cholera in the Bangladesh Sundarbans. Appl. Environ. Microbiol. 2011, 77, 6125–6132. [Google Scholar] [CrossRef] [PubMed]
- Almada, A.A.; Tarrant, A.M. Vibrio elicits targeted transcriptional responses from copepod hosts. FEMS Microbiol. Ecol. 2016, 92, fiw072. [Google Scholar] [CrossRef] [PubMed]
- Steinberg, D.K. Diet of copepods (Scopalatum vorax) associated with mesopelagic detritus (giant larvacean houses) in Monterey Bay, California. Mar. Biol. 1995, 122, 571–584. [Google Scholar] [CrossRef]
- Islam, R.; Hossain, M.B.; Das, N.G.; Rafi, R.U.N. Food and feeding behavior of grey mullet Mugil cephalus of Bangladesh coastal water. Bangladesh J. Progress. Sci. Technol. 2009, 7, 273–276. [Google Scholar]
- Ettinger, C.L.; Ostovar, T.; Yacoub, M.; Ahrendt, S.; Hice, R.H.; Federici, B.A.; Stajich, J.E. Genomes and transcriptomes help unravel the complex life cycle of the blastoclad fungus, Coelomomyces lativittatus, an obligate parasite of mosquitoes and microcrustaceans. Mycologia 2023, 115, 630–647. [Google Scholar] [CrossRef]
- Haag, C.R.; Ebert, D. Parasite-mediated selection in experimental metapopulations of Daphnia magna. Proc. R. Soc. B 2004, 271, 2149–2155. [Google Scholar] [CrossRef]
- Lynch, M. Complex interactions between natural coexploiters—Daphnia and Ceriodaphnia. Ecology 1978, 59, 552–564. [Google Scholar] [CrossRef]
- Zaret, T.M. Predation and Freshwater Communities; Yale University Press: New Haven, UK; London, UK, 1980. [Google Scholar]
- Minchella, D.J.; Scott, M.E. Parasitism: A cryptic determinant of animal community structure. Trends Ecol. Evol. 1991, 6, 250–254. [Google Scholar] [CrossRef] [PubMed]
- Green, A.J. Implications of pathogenic fungi for lifehistory evolution in amphibians. Funct. Ecol. 1999, 13, 573–575. [Google Scholar]
- Redfield, G.W.; Vincent, W.F. Stages of infection and ecological effects of a fungal epidemic on the eggs of a limnetic copepod. Freshw. Biol. 1979, 9, 503–510. [Google Scholar] [CrossRef]
- Burns, C.W. Occurrence of Aphanomyces ovidestruens, a fungus parasitic on copepods, in two eutrophic lakes. N. Z. J. Mar. Freshw. Res. 1980, 14, 23–29. [Google Scholar] [CrossRef]
- Burns, C.W. Fungal parasitism in a copepod population: The effects of Aphanomyces on the population dynamics of Boeckella dilatata Sars. J. Plankton Res. 1985, 7, 201–205. [Google Scholar] [CrossRef]
- Miao, S.; Nauwerck, A. Fungal infection of Eudiaptomus gracilis (Copepoda, Crustacea) in Lake Mondsee. Limnologica 1999, 29, 168–173. [Google Scholar] [CrossRef]
- Rossetti, G. Fungal parasitism in freshwater calanoid population: Ecological consequences and possible mechanisms involved in the infection process. Hydrobiologia 2005, 548, 167–176. [Google Scholar] [CrossRef]
- Czeczuga, B.; Godlewska, A.; Kozłowska, M. Zoosporic fungi growing on the carapaces of dead zooplankton organisms. Limnol. Ecol. Manag. Inland Waters 2000, 30, 37–43. [Google Scholar] [CrossRef]
- Czeczuga, B.; Kozłowska, M.; Godlewska, A. Zoosporic aquatic fungi growing on dead specimens of 29 freshwater crustacean species. Limnologica 2002, 32, 180–193. [Google Scholar] [CrossRef]
- Tang, K.W.; Hutalle, K.M.L.; Grossart, H.P. Microbial abundance, composition, and enzymatic activity during decomposition of copepod carcasses. Aquat. Microb. Ecol. 2006, 45, 219–227. [Google Scholar] [CrossRef]
- Wurzbacher, C.; Bärlocher, F.; Grossart, H.P. Fungi in lake ecosystems. Aquat. Microb. Ecol. 2010, 59, 125–149. [Google Scholar] [CrossRef]
- Vermont, A.I.; Martínez Martínez, J.; Waller, J.D.; Gilg, I.C.; Leavitt, A.H.; Floge, S.A.; Archer, S.D.; Wilson, W.H.; Fields, D.M. Virus infection of Emiliania huxleyi deters grazing by the copepod Acartia tonsa. J. Plankton Res. 2016, 38, 1194–1205. [Google Scholar] [CrossRef]
- Thingstad, T.F.; Våge, S.; Bratbak, G.; Egge, J.; Larsen, A.; Nejstgaard, J.C.; Sandaa, R.-A. Reproducing the virus-to-copepod link in Arctic mesocosms using host fitness optimization. Limnol. Oceanogr. 2021, 66, S303–S313. [Google Scholar] [CrossRef]
- Drake, L.A.; Dobbs, F.C. Do viruses affect fecundity and survival of the copepod Acartia tonsa Dana? J. Plankton Res. 2005, 27, 167–174. [Google Scholar] [CrossRef]
- Dunlap, D.S.; Ng, T.F.F.; Rosario, K.; Barbosa, J.G.; Greco, A.M.; Breitbart, M.; Hewson, I. Molecular and microscopic evidence of viruses in marine copepods. Proc. Natl. Acad. Sci. USA 2013, 110, 1375–1380. [Google Scholar] [CrossRef] [PubMed]
- Dunlap, D.S. Evidence of Viruses in the Copepods Labidocera aestiva and Acartia tonsa in Tampa Bay, Florida. Master’s Thesis, University of South Florida, Tampa, FL, USA, 2012. [Google Scholar]
- Traboni, C. Trophic Interactions between Mixoplankton and Copepods. Ph.D. Thesis, Free University of Brussels, Brussels, Belgium, 2022. [Google Scholar]
- Dutz, J.; Peters, J. Importance and nutritional value of large ciliates for the reproduction of Acartia clausi during the post-spring-bloom period in the North Sea. Aquat. Microb. Ecol. 2008, 50, 261–277. [Google Scholar] [CrossRef][Green Version]
- Burns, C.W.; Schallenberg, M. Calanoid copepods versus cladocerans: Consumer effects on protozoa in lakes of different trophic status. Limnol. Oceanogr. 2001, 46, 1558–1565. [Google Scholar] [CrossRef]
- Wickham, S.A. Trophic relations between cyclopoid copepods and ciliated protists: Complex interactions link the microbial and classic food webs. Limnol. Oceanogr. 1995, 40, 1173–1181. [Google Scholar] [CrossRef]
- Levinsen, H.; Turner, J.T.; Nielsen, T.G.; Hansen, B.W. On the trophic coupling between protists and copepods in Arctic marine ecosystems. Mar. Ecol. Prog. Ser. 2000, 204, 65–77. [Google Scholar] [CrossRef]
- Nese, L.; Enger, Ø. Isolation of Aeromonas salmonicida from salmon lice Lepeophtheirus salmonis and marine plankton. Dis. Aquat. Org. 1993, 16, 79–81. [Google Scholar] [CrossRef]
- Nagasawa, S.; Terazaki, M.; Nemoto, T. Bacterial attachment to the epipelagic copepod Acartia and the bathypelagic copepod Calanus. Proc. Jpn. Acad. Ser. B 1987, 63, 33–35. [Google Scholar] [CrossRef][Green Version]
- Shoemaker, K.M.; McCliment, E.A.; Moisander, P.H. Copepod-associated gammaproteobacterial alkaline phosphatases in the North Atlantic Subtropical Gyre. Front. Microbiol. 2020, 11, 1033. [Google Scholar] [CrossRef] [PubMed]
- Tang, K.; Dziallas, C.; Hutalle-Schmelzer, K.; Grossart, H.P. Effects of food on bacterial community composition associated with the copepod Acartia tonsa Dana. Biol. Lett. 2009, 5, 549–553. [Google Scholar] [CrossRef] [PubMed]
- Ray, J.L.; Althammer, J.; Skaar, K.S.; Simonelli, P.; Larsen, A.; Stoecker, D.; Sazhin, A.; Ijaz, U.Z.; Quince, C.; Nejstgaard, J.C.; et al. Metabarcoding and metabolome analyses of copepod grazing reveal feeding preference and linkage to metabolite classes in dynamic microbial plankton communities. Mol. Ecol. 2016, 25, 5585–5602. [Google Scholar] [CrossRef] [PubMed]
- Adair, K.L.; Douglas, A.E. Making a microbiome: The many determinants of host-associated microbial community composition. Curr. Opin. Microbiol. 2017, 35, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Autio, R. Response of seasonally cold-water bacterioplankton to temperature and substrate treatments. Estuar. Coast. Shelf Sci. 1998, 46, 465–474. [Google Scholar] [CrossRef]
- Hansen, P.J. Effect of high pH on the growth and survival of marine phytoplankton: Implications for species succession. Aquat. Microb. Ecol. 2002, 28, 279–288. [Google Scholar] [CrossRef]
- Cripps, G.; Lindeque, P.; Flynn, K. Parental exposure to elevated pCO2 influences the reproductive success of copepods. J. Plankton Res. 2014, 36, 1165–1174. [Google Scholar] [CrossRef]
- Skovgaard, A.; Castro-Mejia, J.L.; Hansen, L.H.; Nielsen, D.S. Host-specific and pH-dependent microbiomes of copepods in an extensive rearing system. PLoS ONE 2015, 10, e0132516. [Google Scholar] [CrossRef]
- Grossart, H.P.; Dziallas, C.; Leunert, F.; Tang, K.W. Bacteria dispersal by hitchhiking on zooplankton. Proc. Natl. Acad. Sci. USA 2010, 107, 11959–11964. [Google Scholar] [CrossRef] [PubMed]
- Do Sul, J.A.I.; Costa, M.F. The present and future of microplastic pollution in the marine environment. Environ. Pollut. 2014, 185, 352–364. [Google Scholar] [CrossRef] [PubMed]
- Gregory, M.R. Environmental implications of plastic debris in marine settings—Entanglement, ingestion, and alien invasions. Philos. Trans. R. Soc. 2009, 364, 2013–2025. [Google Scholar] [CrossRef] [PubMed]
- Wäge, J.; Strassert, J.F.; Landsberger, A.; Loick-Wilde, N.; Schmale, O.; Stawiarski, B.; Labrenz, M. Microcapillary sampling of Baltic Sea copepod gut microbiomes indicates high variability among individuals and the potential for methane production. FEMS Microbiol. Ecol. 2019, 95, fiz024. [Google Scholar] [CrossRef] [PubMed]
- Sunderland, E.M.; Krabbenhoft, D.P.; Moreau, J.W.; Strode, S.A.; Landing, W.M. Mercury sources, distribution, and bioavailability in the North Pacific Ocean: Insights from data and models. Glob. Biogeochem. Cycles 2009, 23, GB2010. [Google Scholar] [CrossRef]
- Sadaiappan, B.; PrasannaKumar, C.; Nambiar, V.U.; Subramanian, M.; Gauns, M.U. Meta-analysis cum machine learning approaches address the structure and biogeochemical potential of marine copepod associated bacteriobiomes. Sci. Rep. 2021, 11, 3312. [Google Scholar] [CrossRef] [PubMed]
- Glöckner, F.O.; Fuchs, B.M.; Amann, R. Bacterioplankton compositions of lakes and oceans: A first comparison based on fluorescence in situ hybridization. Appl. Environ. Microbiol. 1999, 65, 3721–3726. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, J.; Mazzei, M.; Di Gregorio, S.; Niccolini, L.; Vitiello, V.; Ye, Y.; Guo, B.; Yan, X.; Buttino, I. Marine Copepods as a Microbiome Hotspot: Revealing Their Interactions and Biotechnological Applications. Water 2023, 15, 4203. https://doi.org/10.3390/w15244203
Feng J, Mazzei M, Di Gregorio S, Niccolini L, Vitiello V, Ye Y, Guo B, Yan X, Buttino I. Marine Copepods as a Microbiome Hotspot: Revealing Their Interactions and Biotechnological Applications. Water. 2023; 15(24):4203. https://doi.org/10.3390/w15244203
Chicago/Turabian StyleFeng, Jiantong, Maurizio Mazzei, Simona Di Gregorio, Luca Niccolini, Valentina Vitiello, Yingying Ye, Baoying Guo, Xiaojun Yan, and Isabella Buttino. 2023. "Marine Copepods as a Microbiome Hotspot: Revealing Their Interactions and Biotechnological Applications" Water 15, no. 24: 4203. https://doi.org/10.3390/w15244203
APA StyleFeng, J., Mazzei, M., Di Gregorio, S., Niccolini, L., Vitiello, V., Ye, Y., Guo, B., Yan, X., & Buttino, I. (2023). Marine Copepods as a Microbiome Hotspot: Revealing Their Interactions and Biotechnological Applications. Water, 15(24), 4203. https://doi.org/10.3390/w15244203