

Flooding Depth and Flooding Duration with the Zonation of Riparian Plant Communities in the Three Gorges Reservoir of China
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Vegetation Investigation
2.3. Inundation Characteristics
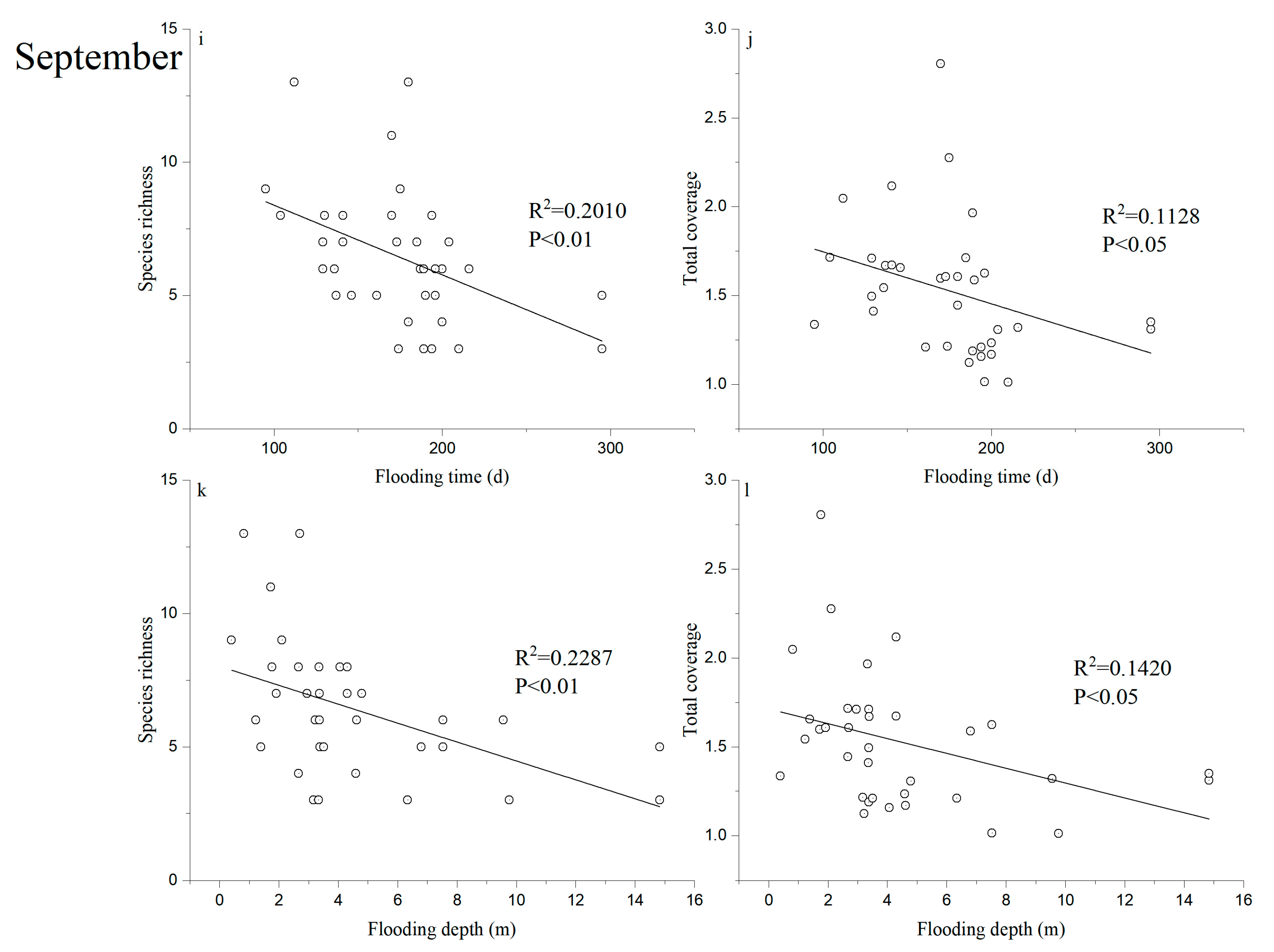
2.4. Data Analysis
3. Results
4. Discussion
4.1. Plant Community Varied throughout the Seasons in the Riparian Zone
4.2. Different Plant Community Constructions in Different Seasons of the Riparian Zone
4.3. Flooding Depth: A Better Indicator for the Zonation of the Riparian Vegetation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gregory, S.V.; Swanson, F.J.; McKee, W.A.; Cummins, K.W. An Ecosystem Perspective of Riparian Zones: Focus on links between land and water. BioScience 1991, 41, 540–551. [Google Scholar] [CrossRef]
- Naiman, R.J.; Décamps, H. The Ecology of Interfaces: Riparian Zones. Annu. Rev. Ecol. Syst. 1997, 28, 621–658. [Google Scholar] [CrossRef]
- Palmer, M.A.; Ruhi, A. Linkages between flow regime, biota, and ecosystem processes: Implications for river restoration. Science 2019, 365, eaaw2087. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, C.; Berggren, K. Alterations of riparian ecosystems caused by river regulation. BioScience 2000, 50, 783–792. [Google Scholar] [CrossRef]
- Tonkin, J.D.; Merritt, D.M.; Olden, J.D.; Reynolds, L.V.; Lytle, D.A. Flow regime alteration degrades ecological networks in riparian ecosystems. Nat. Ecol. Evol. 2018, 2, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Lou, Y.; Mei, X.; Dai, Z.; Wang, J.; Wei, W. Evolution of the mid-channel bars in the middle and lower reaches of the Changjiang (Yangtze) River from 1989 to 2014 based on the Landsat satellite images: Impact of the Three Gorges Dam. Environ. Earth Sci. 2018, 77, 394. [Google Scholar] [CrossRef]
- Kiss, T.; Balogh, M. Characteristics of point-bar development under the influence of a dam: Case study on the Dráva River at Sigetec, Croatia. J. Environ. Geogr. 2015, 8, 23–30. [Google Scholar] [CrossRef]
- Skalak, K.J.; Benthem, A.J.; Schenk, E.R.; Hupp, C.R.; Galloway, J.M.; Nustad, R.A.; Wiche, G.J. Large dams and alluvial rivers in the Anthropocene: The impacts of the Garrison and Oahe Dams on the Upper Missouri River. Anthropocene 2013, 2, 51–64. [Google Scholar] [CrossRef]
- Gu, Z.K.; Fan, H.; Wang, Y.H. Dynamic characteristics of sandbar evolution in the lower Lancang-Mekong River between 1993 and 2012 in the context of hydropower development. Estuar. Coast. Shelf Sci. 2020, 237, 11. [Google Scholar] [CrossRef]
- Fan, H.; He, D.M.; Wang, H.L. Environmental consequences of damming the mainstream Lancang-Mekong River: A review. Earth-Sci. Rev. 2015, 146, 77–91. [Google Scholar] [CrossRef]
- Yang, X.; Lu, X.X. Ten years of the Three Gorges Dam: A call for policy overhaul. Environ. Res. Lett. 2013, 8, 041006. [Google Scholar] [CrossRef]
- Li, D.; Lu, X.X.; Chen, L.; Wasson, R. Downstream geomorphic impact of the Three Gorges Dam: With special reference to the channel bars in the Middle Yangtze River. Earth Surf. Process. Landf. 2019, 44, 2660–2670. [Google Scholar] [CrossRef]
- Willison, J.; Li, R.; Yuan, X. Conservation and ecofriendly utilization of wetlands associated with the Three Gorges Reservoir. Environ. Sci. Pollut. Res. 2013, 20, 6907–6916. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Yuan, X.; Roß-Nickoll, M.; Hollert, H.; Schäffer, A. Moderate inundation stimulates plant community assembly in the drawdown zone of China’s Three Gorges Reservoir. Environ. Sci. Eur. 2020, 32, 79. [Google Scholar] [CrossRef]
- Strayer, D.L.; Findlay, S. Ecology of freshwater shore zones. Aquat. Sci. 2010, 72, 127–163. [Google Scholar] [CrossRef]
- Ye, C.; Butler, O.M.; Chen, C.; Liu, W.; Du, M.; Zhang, Q. Shifts in characteristics of the plant-soil system associated with flooding and revegetation in the riparian zone of Three Gorges Reservoir, China. Geoderma 2020, 361, 114015. [Google Scholar] [CrossRef]
- Lin, J.; Zhou, S.; Liu, D.; Zhang, S.; Yu, Z.; Yang, X. Relative contribution of environmental and nutritional variables to net primary production of Cynodon dactylon (Linn.) Pers in the riparian zone of a Three Gorges tributary. Ecol. Evol. 2020, 10, 7073–7081. [Google Scholar] [CrossRef]
- Hupp, C.R.; Rinaldi, M. Riparian Vegetation Patterns in Relation to Fluvial Landforms and Channel Evolution Along Selected Rivers of Tuscany (Central Italy). Ann. Assoc. Am. Geogr. 2007, 97, 12–30. [Google Scholar] [CrossRef]
- Sandercock, P.J.; Hooke, J.M.; Sandercock, P.J.; Hooke, J.M. Assessment of vegetation effects on hydraulics and of feedbacks on plant survival and zonation in ephemeral channels. Hydrol. Process. 2010, 24, 695–713. [Google Scholar] [CrossRef]
- Bejarano, M.D.; Jansson, R.; Nilsson, C. The effects of hydropeaking on riverine plants: A review. Biol. Rev. 2018, 93, 658–673. [Google Scholar] [CrossRef]
- Toner, M.; Keddy, P. River hydrology and riparian werlands: A predictive model for ecological assembly. Ecol. Appl. 1997, 7, 236–246. [Google Scholar] [CrossRef]
- Aguiar, F.C.; Segurado, P.; Martins, M.J.; Bejarano, M.D.; Nilsson, C.; Portela, M.M.; Merritt, D.M. The abundance and distribution of guilds of riparian woody plants change in response to land use and flow regulation. J. Appl. Ecol. 2018, 55, 2227–2240. [Google Scholar] [CrossRef]
- Butterfield, B.J.; Palmquist, E.; Ralston, B.E. Hydrological regime and climate interactively shape riparian vegetation composition along the Colorado River, Grand Canyon. Appl. Veg. Sci. 2018, 21, 572–583. [Google Scholar] [CrossRef]
- Jansson, R.; Strom, L.; Nilsson, C. Smaller future floods imply less habitat for riparian plants along a boreal river. Ecol. Appl. 2019, 29, e01977. [Google Scholar] [CrossRef]
- Su, X.; Bejarano, M.D.; Yi, X.; Lin, F.; Ayi, Q.; Zeng, B. Unnatural flooding alters the functional diversity of riparian vegetation of the Three Gorges Reservoir. Freshw. Biol. 2020, 20, 1585–1595. [Google Scholar] [CrossRef]
- Lytle, D.A.; Merritt, D.M.; Tonkin, J.D.; Olden, J.D.; Reynolds, L.V. Linking river flow regimes to riparian plant guilds: A community-wide modeling approach. Ecol. Appl. 2017, 27, 1338–1350. [Google Scholar] [CrossRef]
- Vreugdenhil, S.J.; Kramer, K.; Pelsma, T. Effects of flooding duration, -frequency and -depth on the presence of saplings of six woody species in north-west Europe. For. Ecol. Manag. 2006, 236, 47–55. [Google Scholar] [CrossRef]
- Engloner, A.I.; Németh, K.; Gere, D.; Stefán, D.; Óvári, M. Effects of water depth and water level fluctuation on the total and bio-available element concentrations in riverine reed stands. Ecol. Indic. 2020, 114, 106328. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, Z.; Ge, W.; Chen, X.; Wang, T. Impact of extreme floods on plants considering various influencing factors downstream of Luhun Reservoir, China. Sci. Total Environ. 2021, 768, 145312. [Google Scholar] [CrossRef]
- Kramer, K.; Vreugdenhil, S.J.; Werf, D. Effects of flooding on the recruitment, damage and mortality of riparian tree species: A field and simulation study on the Rhine floodplain. For. Ecol. Manag. 2008, 255, 3893–3903. [Google Scholar] [CrossRef]
- Garssen, A.G.; Baattrup-Pedersen, A.; Voesenek, L.A.C.J.; Verhoeven, J.T.A.; Soons, M.B. Riparian plant community responses to increased flooding: A meta-analysis. Glob. Chang. Biol. 2015, 21, 2881–2890. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Zamorano, M.F.; Ivanoff, D. Effect of Flooding Depth on Growth, Biomass, Photosynthesis, and Chlorophyll Fluorescence of Typha domingensis. Wetlands 2010, 30, 957–965. [Google Scholar] [CrossRef]
- Sorrell, B.K.; Tanner, C.C.; Brix, H. Regression analysis of growth responses to water depth in three wetland plant species. AoB Plants 2012, 2012, pls043. [Google Scholar] [CrossRef] [PubMed]
- New, T.; Xie, Z. Impacts of large dams on riparian vegetation: Applying global experience to the case of China’s Three Gorges Dam. Biodivers. Conserv. 2008, 17, 3149–3163. [Google Scholar] [CrossRef]
- Chen, J.; Li, G.; Xiao, B.; Wen, Z.; Lv, M.; Chen, C.; Jiang, Y.; Wang, X.; Wu, S. Assessing the transferability of support vector machine model for estimation of global solar radiation from air temperature. Energy Convers. Manag. 2015, 89, 318–329. [Google Scholar] [CrossRef]
- Lv, M.; Chen, J.; Mirza, Z.A.; Chen, C.; Wen, Z.; Jiang, Y.; Ma, M.; Wu, S. Spatial distribution and temporal variation of reference evapotranspiration in the Three Gorges Reservoir area during 1960–2013. Int. J. Climatol. 2016, 36, 4497–4511. [Google Scholar] [CrossRef]
- Chen, J.; He, L.; Yang, H.; Ma, M.; Chen, Q.; Wu, S.; Xiao, Z. Empirical models for estimating monthly global solar radiation: A most comprehensive review and comparative case study in China. Renew. Sustain. Energy Rev. 2019, 108, 91–111. [Google Scholar] [CrossRef]
- Yi, X.; Huang, Y.; Ma, M.; Wen, Z.; Wu, S. Plant trait-based analysis reveals greater focus needed for mid-channel bar downstream from the Three Gorges Dam of the Yangtze River. Ecol. Indic. 2020, 111, 105950. [Google Scholar] [CrossRef]
- DÍAz, S.; Lavorel, S.; McIntyre, S.U.E.; Falczuk, V.; Casanoves, F.; Milchunas, D.G.; Skarpe, C.; Rusch, G.; Sternberg, M.; Noy-Meir, I.; et al. Plant trait responses to grazing—A global synthesis. Glob. Chang. Biol. 2006, 13, 313–341. [Google Scholar] [CrossRef]
- Rao, C.R. Diversity and dissimilarity coefficients: A unified approach. Theor. Popul. Biol. 1982, 21, 24–43. [Google Scholar] [CrossRef]
- Botta-Dukát, Z. Rao’s quadratic entropy as a measure of functional diversity based on multiple traits. J. Veg. Sci. 2005, 16, 533–540. [Google Scholar] [CrossRef]
- Sasaki, T.; Okubo, S.; Okayasu, T.; Jamsran, U.; Ohkuro, T.; Takeuchi, K. Two-phase functional redundancy in plant communities along a grazing gradient in Mongolian rangelands. Ecology 2009, 90, 2598–2608. [Google Scholar] [CrossRef] [PubMed]
- Moor, H.; Rydin, H.; Hylander, K.; Nilsson, M.B.; Lindborg, R.; Norberg, J. Towards a trait-based ecology of wetland vegetation. J. Ecol. 2017, 105, 1623–1635. [Google Scholar] [CrossRef]
- Grygoruk, M.; Kochanek, K.; Mirosław-Świątek, D. Analysis of long-term changes in inundation characteristics of near-natural temperate riparian habitats in the Lower Basin of the Biebrza Valley, Poland. J. Hydrol. Reg. Stud. 2021, 36, 100844. [Google Scholar] [CrossRef]
- Jian, Z.; Ma, F.; Guo, Q.; Qin, A.; Xiao, W. Long-term responses of riparian plants’ composition to water level fluctuation in China’s Three Gorges Reservoir. PLoS ONE 2018, 13, e0207689. [Google Scholar] [CrossRef]
- Hoppenreijs, J.H.T.; Eckstein, R.L.; Lind, L. Pressures on Boreal Riparian Vegetation: A Literature Review. Front. Ecol. Evol. 2022, 9, 806130. [Google Scholar] [CrossRef]
- Eck, W.; Lenssen, J.; Steeg, H.; Blom, C.; Kroon, H.D.J.H. Seasonal Dependent Effects of Flooding on Plant Species Survival and Zonation: A Comparative Study of 10 Terrestrial Grassland Species. Hydrobiologia 2006, 565, 59–69. [Google Scholar]
- Mccoy-Sulentic, M.E.; Kolb, T.E.; Merritt, D.M.; Palmquist, E.; Ralston, B.E.; Sarr, D.A.; Shafroth, P.B. Changes in Community-Level Riparian Plant Traits over Inundation Gradients, Colorado River, Grand Canyon. Wetlands 2017, 37, 635–646. [Google Scholar] [CrossRef]
- Mayfield, M.M.; Boni, M.F.; Daily, G.C.; Ackerly, D. Species and Functional Diversity of Native and Human-Dominated Plant Communities. Ecology 2005, 86, 2365–2372. [Google Scholar] [CrossRef]
- Rood, S.B.; Pan, J.; Gill, K.M.; Franks, C.G.; Samuelson, G.M.; Shepherd, A. Declining summer flows of Rocky Mountain rivers: Changing seasonal hydrology and probable impacts on floodplain forests. J. Hydrol. 2008, 349, 397–410. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, Y.; Wang, J.; Ge, W.; Jiao, Y. Impact evaluation of geomorphic changes caused by extreme floods on inundation area considering geomorphic variations and land use types. Sci. Total Environ. 2021, 754, 142424. [Google Scholar] [CrossRef] [PubMed]
- Van der Pijl, L. Ecological dispersal classes, established on the basis of the dispersing agents. In Principles of Dispersal in Higher Plants; Springer: Berlin/Heidelberg, Germany, 1982; pp. 22–90. [Google Scholar]
- Hodgson, J.; Grime, J.; Hunt, R.; Thompson, K. The Electronic Comparative Plant Ecology; Springer: Berlin/Heidelberg, Germany, 1995. [Google Scholar]
- Mabry, C.M. Floristic Analysis of Central Iowa Woodlands, and Comparison of Reproduction and Regeneration in Common and Restricted Herbaceous Species; Iowa State University: Ames, IA, USA, 2000. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Estimate | Std. Error | p-Value | ||
|---|---|---|---|---|
| April | Intercept | 15.587 | 2.202 | p < 0.001 |
| flooding time | −0.068 | 0.016 | 0.000206 *** | |
| flooding depth | 0.201 | 0.206 | 0.336 | |
| regression equation | Y = 15.587 − 0.068X1 + 0.201X2, p < 0.001 | |||
| July | Intercept | 11.064 | 2.222 | p < 0.001 |
| flooding time | −0.010 | 0.017 | 0.560 | |
| flooding depth | −0.447 | 0.213 | 0.042 * | |
| regression equation | Y = 11.064 − 0. 010X1 − 0.447X2, p < 0.001 | |||
| September | Intercept | 11.080 | 2.246 | p < 0.001 |
| flooding time | −0.011 | 0.017 | 0.531 | |
| flooding depth | −0.286 | 0.213 | 0.189 | |
| regression equation | Y = 11.080 − 0.011X1 − 0.286X2, p < 0.001 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yi, X.; Huang, Y.; Jiang, Y.; Ma, M.; Chen, Q.; Wu, S. Flooding Depth and Flooding Duration with the Zonation of Riparian Plant Communities in the Three Gorges Reservoir of China. Water 2023, 15, 3228. https://doi.org/10.3390/w15183228
Yi X, Huang Y, Jiang Y, Ma M, Chen Q, Wu S. Flooding Depth and Flooding Duration with the Zonation of Riparian Plant Communities in the Three Gorges Reservoir of China. Water. 2023; 15(18):3228. https://doi.org/10.3390/w15183228
Chicago/Turabian StyleYi, Xuemei, Yuanyang Huang, Yi Jiang, Maohua Ma, Qiao Chen, and Shengjun Wu. 2023. "Flooding Depth and Flooding Duration with the Zonation of Riparian Plant Communities in the Three Gorges Reservoir of China" Water 15, no. 18: 3228. https://doi.org/10.3390/w15183228