
Responses of Bacterial Taxonomical Diversity Indicators to Pollutant Loadings in Experimental Wetland Microcosms


Abstract
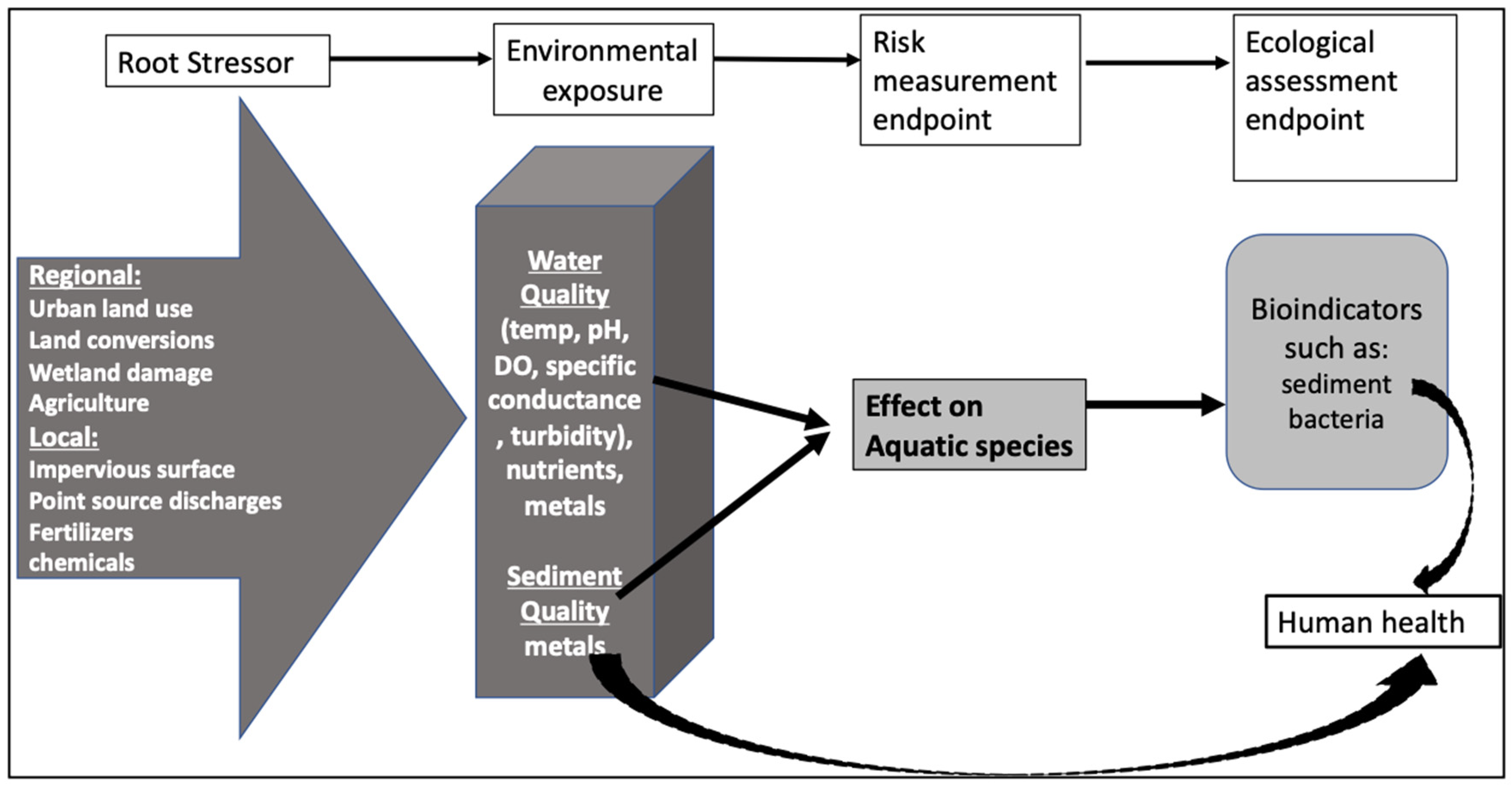
1. Introduction
2. Materials and Methods
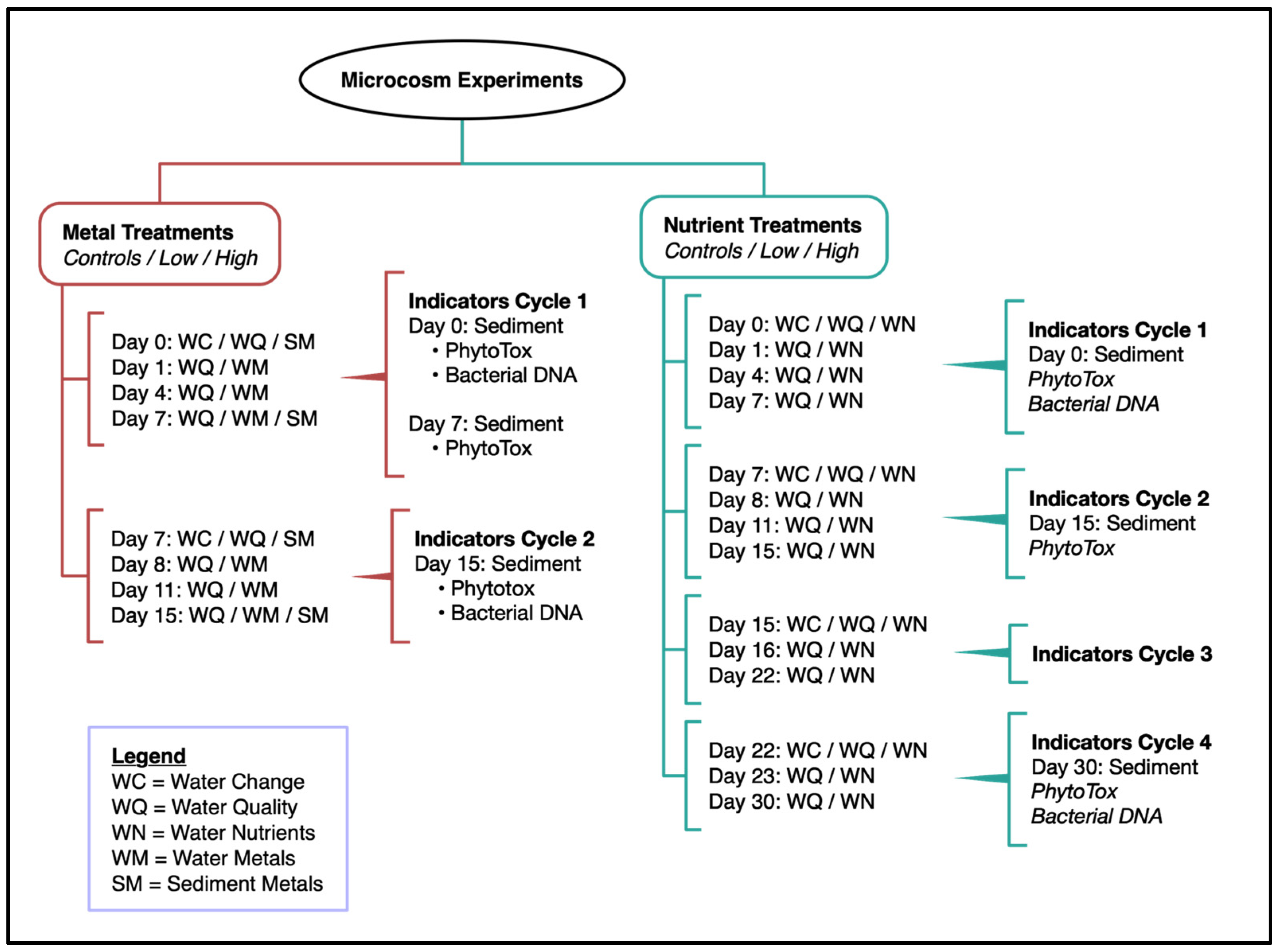
2.1. Sample Collection and Construction of Microcosms
2.2. Types of Microcosms
2.3. Pollutants
2.4. Measurements in Microcosm
2.5. Data Analysis
3. Results
3.1. Time and Treatment Effects on Metals and Nutrients in Microcosms
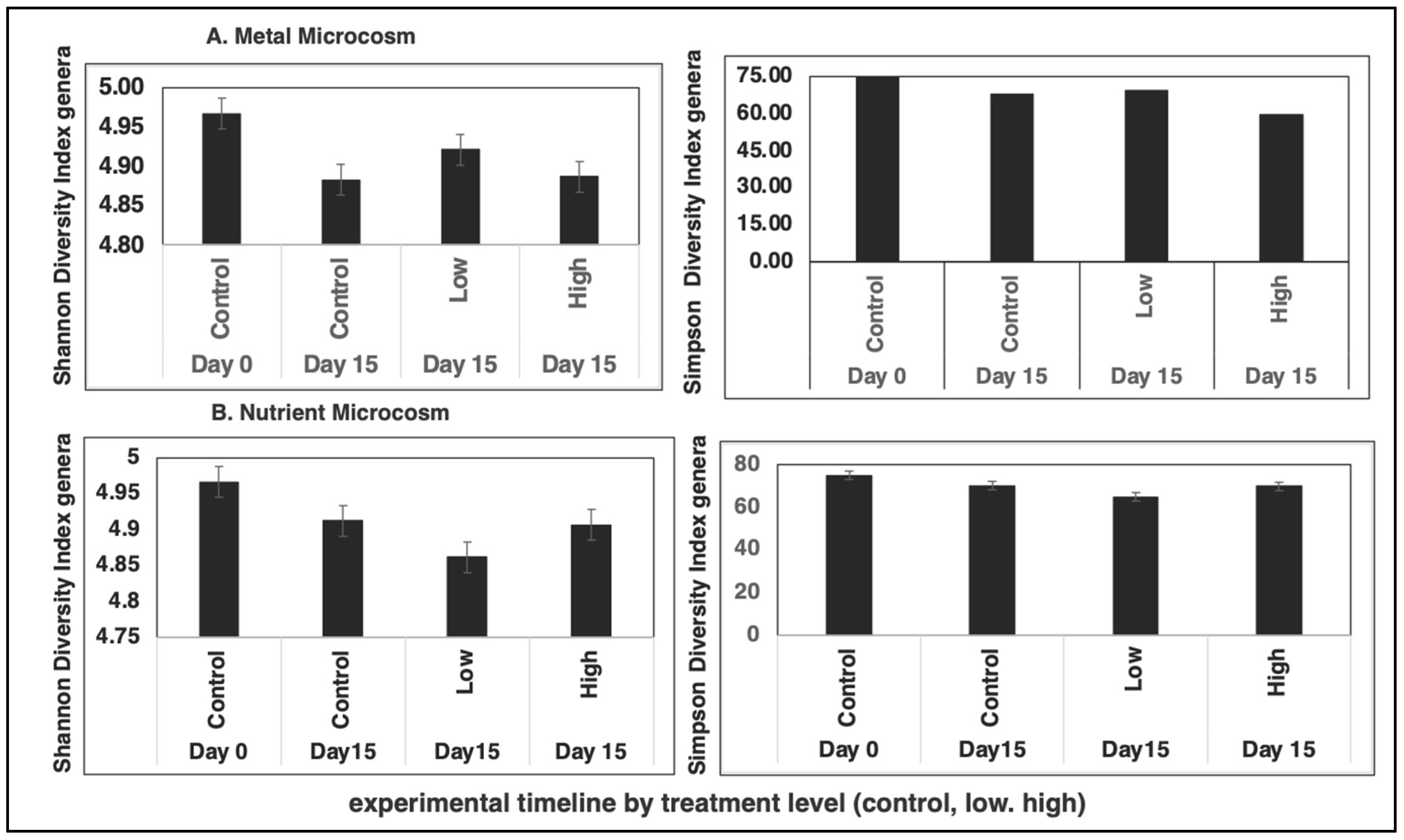
3.2. Time and Treatment Effects on Bacterial Bioindicators
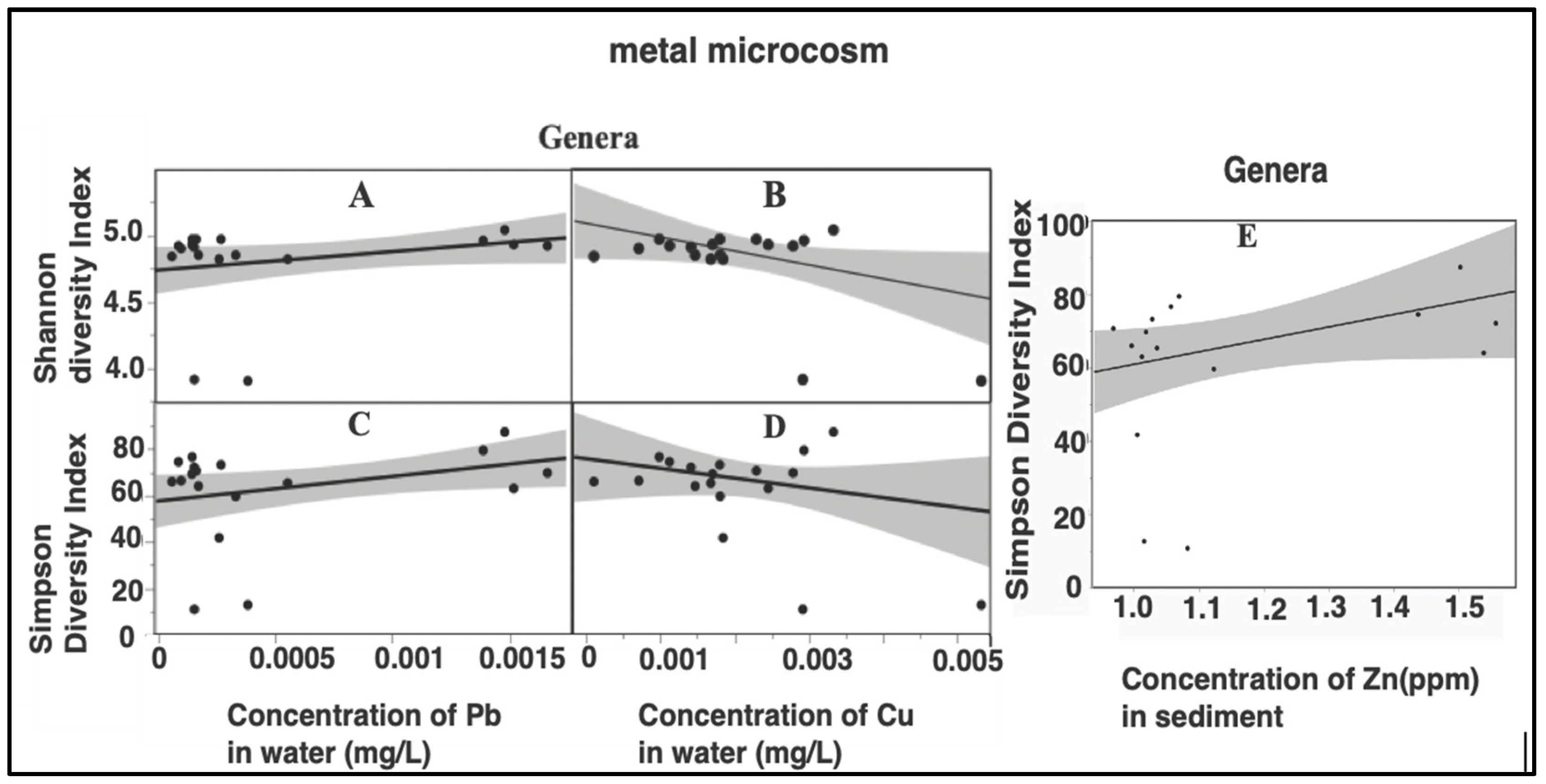
3.3. Effects of Pollutants on Bacterial Bioindicators
3.4. Predictive Indicator Categories Based on Key Pollutants in Field and Microcosm Study
4. Discussion
4.1. Time and Treatment Effects on Pollutants in Microcosms
4.2. Bacterial Bioindicators and Effect of Pollutants
4.3. Finding of Bacterial Bioindicators
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metal Microcosm—Metals Added in Water | ||||||||||||||||||
| Pb (mg/L) | Cu (mg/L) | |||||||||||||||||
| Effect Source | SS | F Ratio | p-Value | SS | F Ratio | p-Value | ||||||||||||
| Experimental duration | 0.000001 | 24.763 | <0.0001 * | 0.0003 | 1.1749 | 0.319 | ||||||||||||
| Treatment level | 0.000067 | 0.5031 | 0.61 | 0.0005 | 0.6749 | 0.515 | ||||||||||||
| Nutrient Microcosm—Nutrients Added in Water | ||||||||||||||||||
| NO3− (mg/L) | PO43− (mg/L) | |||||||||||||||||
| Effect Source | SS | F Ratio | p-Value | SS | F Ratio | p-Value | ||||||||||||
| Experimental duration | 9067 | 6.4182 | 0.0036 * | 26 | 11.9591 | <0.0001 * | ||||||||||||
| Treatment level | 79 | 0.0561 | NS | 6 | 2.5600 | NS | ||||||||||||
| Metal Microcosm—Metal Detected in Sediments | ||||||||||||||||||
| Ag (ppm) | As (ppm) | Cd (ppm) | ||||||||||||||||
| Effect Source | SS | F Ratio | p-Value | SS | F Ratio | p-Value | SS | F Ratio | p-Value | |||||||||
| Experimental duration | 0.0012 | 0.3312 | NS | 0.0767 | 4.2484 | 0.0223 * | 0.0054 | 0.4785 | NS | |||||||||
| Treatment level | 0.0017 | 0.4522 | NS | 0.0307 | 1.7017 | NS | 0.0194 | 1.7162 | NS | |||||||||
| Fe (ppm) | Hg (ppm) | Ni (ppm) | ||||||||||||||||
| Effect Source | SS | F Ratio | p-Value | SS | F Ratio | p-Value | SS | F Ratio | p-Value | |||||||||
| Experimental duration | 0.0002 | 0.0096 | NS | 0.0540 | 2.7600 | NS | 0.000131 | 0.252300 | NS | |||||||||
| Treatment level | 0.0001 | 0.0032 | NS | 0.0025 | 0.1261 | NS | 0.000078 | 0.149100 | NS | |||||||||
| Pb (ppm) | Rb (ppm) | |||||||||||||||||
| Effect Source | SS | F Ratio | p-Value | SS | F Ratio | p-Value | SS | F Ratio | p-Value | |||||||||
| Experimental duration | 0.000011 | 1.9716 | NS | 0.0057 | 0.2966 | NS | 0.0473 | 0.3754 | NS | |||||||||
| Treatment level | 0.000011 | 2.1328 | NS | 0.0083 | 0.4319 | NS | 0.0008 | 0.0067 | NS | |||||||||
| (A) Phyla Detected | Number of Sequences for Each Phylum | Percent of Total |
|---|---|---|
| Proteobacteria | 51,598 | 29.45% |
| Bacteroidetes | 23,252 | 13.27% |
| Chloroflexi | 16,236 | 9.27% |
| Planctomycetes | 10,013 | 5.71% |
| OD1 | 9477 | 5.41% |
| Firmicutes | 7513 | 4.29% |
| Acidobacteria | 7323 | 4.18% |
| Verrucomicrobia | 6934 | 3.96% |
| Actinobacteria | 5417 | 3.09% |
| Chlorobi | 3561 | 2.03% |
| Spirochaetes | 3428 | 1.96% |
| Armatimonadetes | 2702 | 1.54% |
| Elusimicrobia | 2148 | 1.23% |
| Cyanobacteria | 2099 | 1.20% |
| GN02 | 2090 | 1.19% |
| Shannon Diversity Index of Genera | Simpson Diversity Index of Genera | |||||
| Effect Source | SS | F Ratio | p-Value | SS | F Ratio | p-Value |
| Experimental duration | 0.2765 | 3.3789 | NS | 1688.4 | 5.3548 | 0.0314 * |
| Treatment level | 0.0553 | 0.3376 | NS | 410.21 | 0.6505 | NS |
| Shannon Diversity Index of Genera | Simpson Diversity Index of Genera | |||||
| Effect Source | SS | F Ratio | p-Value | SS | F Ratio | p-Value |
| Experimental duration | 0.0804 | 14.0738 | 0.0013 * | 512 | 4.731 | 0.0418 * |
| Treatment level | 0.0143 | 1.2515 | NS | 93 | 0.4282 | NS |
| Genera Shannon Diversity Index | Genera Simpson Diversity Index | |||||
| Parameter | Estimate | p-Value | R2 | Estimate | p-Value | R2 |
| Ag(ppm) | 0 | NS | 0.00 | 0 | NS | 0.27 |
| As(ppm) | 0 | NS | 0 | NS | ||
| Cd(ppm) | 0 | NS | 0 | NS | ||
| Fe(ppm) | 0 | NS | 0 | NS | ||
| Hg(ppm) | 0 | NS | 0 | NS | ||
| Ni(ppm) | 0 | NS | 0 | NS | ||
| Pb(ppm) | 0 | NS | −5606 | 0.0616 | ||
| Rb(ppm) | 0 | NS | 0 | NS | ||
| Zn(ppm) | 0 | NS | 57.85 | 0.0118 | ||
References
- Ding, G.-C.; Piceno, Y.M.; Heuer, H.; Weinert, N.; Dohrmann, A.B.; Carrillo, A.; Andersen, G.L.; Castellanos, T.; Tebbe, C.C.; Smalla, K. Changes of Soil Bacterial Diversity as a Consequence of Agricultural Land Use in a Semi-Arid Ecosystem. PLoS ONE 2013, 8, e59497. [Google Scholar] [CrossRef]
- Roy, S.G.; Wimpee, C.F.; McGuire, S.A.; Ehlinger, T.J. Effects of Land Use and Pollution Loadings on Ecotoxicological Assays and Bacterial Taxonomical Diversity in Constructed Wetlands. Diversity 2021, 13, 149. [Google Scholar] [CrossRef]
- Szoboszlay, M.; Dohrmann, A.B.; Poeplau, C.; Don, A.; Tebbe, C.C. Impact of land-use change and soil organic carbon quality on microbial diversity in soils across Europe. FEMS Microbiol. Ecol. 2017, 93, fix146. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Wang, H.; Liu, J.; Wang, Q.; Shen, T.; Guo, W.; Wang, R. Shifts in microbial community function and structure along the successional gradient of coastal wetlands in Yellow River Estuary. Eur. J. Soil Biol. 2012, 49, 12–21. [Google Scholar] [CrossRef]
- Kovacs, M. Biological Indicators of Environmental Pollution; Kovacs, M., Ed.; Ellis Horwood: New York, NY, USA, 1992. [Google Scholar]
- Karr, J.R.; Chu, E.W. Restoring Life in Running Waters; Island Press: Washington, DC, USA; Covelo, CA, USA, 1999. [Google Scholar]
- Dellinger, M.M.; Carvan, R.; Klingler, J.M.; Ehlinger, T.J. An Exploratory Analysis of Stream Teratogenicity and Human Health Using Zebrafish Whole-Sediment Toxicity Test. Challenges 2014, 5, 75–97. [Google Scholar] [CrossRef]
- Roy, S.G.; Ehlinger, T.J. Relationships between land use, predicted pollution loadings, and ecotoxicological assays in constructed wetlands. Rom. J. Ecol. Environ. Chem. 2020, 2, 118–129. [Google Scholar] [CrossRef]
- Zhao, X.; Huang, J.; Zhu, X.; Chai, J.; Ji, X. Ecological Effects of Heavy Metal Pollution on Soil Microbial Community Structure and Diversity on Both Sides of a River around a Mining Area. Int. J. Environ. Res. Public Health 2020, 17, 5680. [Google Scholar] [CrossRef] [PubMed]
- Stephanou, C.; Omirou, M.; Philippot, L.; Zissimos, A.M.; Christoforou, I.C.; Trajanoski, S.; Oulas, A.; Ioannides, I.M. Land use in urban areas impacts the composition of soil bacterial communities involved in nitrogen cycling. A case study from Lefkosia (Nicosia) Cyprus. Sci. Rep. 2021, 11, 8198. [Google Scholar] [CrossRef]
- Wang, H.; Marshall, C.W.; Cheng, M.; Xu, H.; Li, H.; Yang, X.; Zheng, T. Changes in land use driven by urbanization impact nitrogen cycling and the microbial community composition in soils. Sci. Rep. 2017, 7, 44049. [Google Scholar] [CrossRef]
- Torsvik, V.; Øvreås, L. Microbial diversity and function in soil: From genes to ecosystems. Curr. Opin. Microbiol. 2002, 5, 240–245. [Google Scholar] [CrossRef]
- Azarbad, H.; Van Gestel, C.A.; Niklińska, M.; Laskowski, R.; Röling, W.F.; Van Straalen, N.M. Resilience of soil microbial communities to metals and additional stressors: DNA-based approaches for assessing “stress-on-stress” responses. Int. J. Mol. Sci. 2016, 17, 933. [Google Scholar] [CrossRef]
- Vymazal, J. Removal of nutrients in various types of constructed wetlands. Sci. Total Environ. 2007, 380, 48–65. [Google Scholar] [CrossRef] [PubMed]
- Webb, J.S.; McGinness, S.; Lappin-Scott, H.M. Metal removal by sulphate-reducing bacteria from natural and constructed wetlands. J. Appl. Microbiol. 1998, 84, 240–248. [Google Scholar] [CrossRef]
- Kumar, S.; Pratap, B.; Dubey, D.; Dutta, V. Microbial Communities in Constructed Wetland Microcosms and Their Role in Treatment of Domestic Wastewater. Emerg. Eco-Friendly Green Technol. Wastewater Treat. 2020, 18, 311–327. [Google Scholar]
- Rajan, R.J.; Sudarsan, J.S.; Nithiyanantham, S. Microbial population dynamics in constructed wetlands: Review of recent advancements for wastewater treatment. Environ. Eng. Res. 2019, 24, 181–190. [Google Scholar] [CrossRef]
- Ke, X.; Wang, C.; Jing, D.; Zhang, Y.; Zhang, H. Assessing water quality by ratio of the number of dominant bacterium species between surface/subsurface sediments in Haihe River Basin. Mar. Pollut. Bull. 2015, 98, 267–273. [Google Scholar] [CrossRef]
- Aylagas, E.; Borja, Á.; Tangherlini, M.; Dell’Anno, A.; Corinaldesi, C.; Michell, C.T.; Irigoien, X.; Danovaro, R.; Rodríguez-Ezpeleta, N. A bacterial community-based index to assess the ecological status of estuarine and coastal environments. Mar. Pollut. Bull. 2017, 114, 679–688. [Google Scholar] [CrossRef] [PubMed]
- Chapman, D.P.M. Do sediment toxicity tests require field validation? Environ. Toxicol. Chem. 1995, 14, 1451–1453. [Google Scholar] [CrossRef]
- Jensen, J. Sediment Toxicity and the Recovery of Biological Integrity in a Restored Stream Channel; University of Wisconsin Milwaukee: Milwaukee, WI, USA, 2011. [Google Scholar]
- Crispell-Synder, I. Executive Summary of WDNR Facilitation Presentation for Pike River Improvements by Mount Pleasant Storm Water Drainage District No. 1. 1997. Available online: https://www.mtpleasantwi.gov/2325/Pike-River-Improvements-Project (accessed on 19 March 2016).
- Ehlinger, T.; DeThorne, L.; Berner, B. Monitoring of Stream Habitat and Aquatic Biotic Integrity—Pike River North and South Branches, Racine and Kenosha Counties, Wisconsin; Department of Biological Sciences, University of Wisconsin-Milwaukee: Milwaukee, WI, USA, 2002. [Google Scholar]
- Ehlinger, T.; DeThorne, L. Monitoring of Stream Habitat and Aquatic Biotic Integrity—Pike River North and South Branches, Racine and Kenosha Counties, Wisconsin; Interim Report; Department of Biological Sciences, University of Wisconsin-Milwaukee: Milwaukee, WI, USA, 2004. [Google Scholar]
- SEWRPC, Racine County Mapbook. 2010. Available online: https://www.sewrpc.org/SEWRPC.htm (accessed on 20 October 2020).
- Zhang, D.; Luo, J.; Lee, Z.; Maspolim, Y.; Gersberg, R.; Liu, Y. Characterization of bacterial communities in wetland mesocosms receiving pharmaceutical-enriched wastewater. Ecol. Eng. 2016, 90, 215–224. [Google Scholar] [CrossRef]
- Messer, T.L.; Burchell, M.; Birgand, F.; Broome, S.W.; Chescheir, G. Nitrate removal potential of restored wetlands loaded with agricultural drainage water: A mesocosm scale experimental approach. Ecol. Eng. 2017, 106, 541–554. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, M.; Xue, J.; Lv, Q.; Yang, J.; Han, X. Performance and microbial response in a multi-stage constructed wetland microcosm co-treating acid mine drainage and domestic wastewater. J. Environ. Chem. Eng. 2021, 9, 106786. [Google Scholar] [CrossRef]
- Ramond, J.B.; Welz, P.J.; Cowan, D.A.; Burton, S.G. Microbial community structure stability, a key parameter in monitoring the development of constructed wetland mesocosms during start-up. Res. Microbiol. 2012, 163, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Shange, R.; Haugabrooks, E.; Ankumah, R.; Ibekwe, A.M.; Smith, R.C.; Dowd, S. Assessing the Diversity and Composition of Bacterial Communities across a Wetland, Transition, Upland Gradient in Macon County Alabama. Diversity 2013, 5, 461–478. [Google Scholar] [CrossRef]
- Hermans, S.M.; Buckley, H.L.; Case, B.S.; Curran-Cournane, F.; Taylor, M.; Lear, G. Bacteria as Emerging Indicators of Soil Condition. Appl. Environ. Microbiol. 2017, 83, 02826-16. [Google Scholar] [CrossRef]
- Rüegg, S.R.; Nielsen, L.R.; Buttigieg, S.C.; Santa, M.; Aragrande, M.; Canali, M.; Ehlinger, T.; Chantziaras, I.; Boriani, E.; Radeski, M.; et al. A Systems Approach to Evaluate One Health Initiatives. Front. Vet. Sci. 2018, 5, 23. [Google Scholar] [CrossRef] [PubMed]
- Novotny, V.; Ehlinger, T.; Bartošová, A.; Manolakos, E.; O’Reilly, N. Developing Risk Propagation Model for Estimating Ecological Responses of Streams to Anthropogenic Watershed Stresses and Stream Modification. Cent. Urban Environ. Stud. Publ. 2007. paper 15. Available online: https://www.researchgate.net/publication/278678617_Developing_Risk_Propagation_Model_for_Estimating_Ecological_Responses_of_Streams_to_Anthropogenic_Watershed_Stresses_and_Stream_Modifications (accessed on 14 February 2020).
- Busnardo, M.J.; Gersberg, R.M.; Langis, R.; Sinicrope, T.L.; Zedler, J.B. Nitrogen and phosphorus removal by wetland mesocosms subjected to different hydroperiods. Ecol. Eng. 1992, 1, 287–307. [Google Scholar] [CrossRef]
- Ahn, C.; Gillevet, P.; Sikaroodi, M. Molecular characterization of microbial communities in treatment microcosm wetlands as influenced by macrophytes and phosphorus loading. Ecol. Indic. 2007, 7, 852–863. [Google Scholar] [CrossRef]
- Gikas, P.; Ranieri, E.; Tchobanoglous, G. Removal of iron, chromium and lead from waste water by horizontal subsurface flow constructed wetlands. J. Chem. Technol. Biotechnol. 2013, 88, 1906–1912. [Google Scholar] [CrossRef]
- Owens, P.L.; HInkle, R.C.; Best, R.G. Low-Energy Wastewater Recycling Through Wetland Ecosystem: Copper and Zinc in Wetland Microcosm. In Freshwater Wetlands and Wildlife, Issue DOE Symposium Series No. 61, 24–27 March; Sharitz, R.R., Gibbons, J.W., Eds.; USDOE Office of Scientific and Technical Information: Oak Ridge, TN, USA, 1989. [Google Scholar]
- Sinicrope, T.L.; Langis, R.; Gersberg, R.M.; Busnardo, M.J.; Zedler, J.B. Metal removal by wetland mesocosms subjected to different hydroperiods. Ecol. Eng. 1992, 1, 309–322. [Google Scholar] [CrossRef]
- Behrends, L.L. Use of Aquatic Plants for Removal of Nitrate-Nitrogen in Subsurface Flow Constructed Wetlands; ReCiprocating Water Technologies LLC: Florence, AL, USA, 2010. [Google Scholar]
- EPA. U.S. Environmental Protection Agency. Freshwater Wetlands for Wastewater Management Handbook. EPA Reg. IV Atlanta GA EPA 1985, 904, 9,85,135. [Google Scholar]
- Krachler, M.; Alvarez-Sarandes, R.; Rasmussen, G. High-Resolution Inductively Coupled Plasma Optical Emission Spectrometry for 234U/238Pu Age Dating of Plutonium Materials and Comparison to Sector Field Inductively Coupled Plasma Mass Spectrometry. Anal. Chem. 2016, 88, 8862–8869. [Google Scholar] [CrossRef][Green Version]
- YSI. YSI a Xylem Brand. 2020. Available online: https://www.ysi.com (accessed on 19 January 2020).
- Burbach, K.; Seifert, J.; Pieper, D.H.; Camarinha-Silva, A. Evaluation of DNA extraction kits and phylogenetic diversity of the porcine gastrointestinal tract based on Illumina sequencing of two hypervariable regions. MicrobiologyOpen 2016, 5, 70–82. [Google Scholar] [CrossRef]
- Li, J.; Li, B.; Zhou, Y.; Xu, J.; Zhao, J. A rapid DNA extraction method for PCR amplification from wetland soils. Lett. Appl. Microbiol. 2011, 52, 626–633. [Google Scholar] [CrossRef] [PubMed]
- MPBiomedicals. Fast DNATM Spinkit. 2017. Available online: http://www.mpbio.com/product.php?pid=116540600%0D%0D (accessed on 19 May 2019).
- SAS. JMP, Version 14; SAS Institute Inc.: Cary, NC, USA, 2019; Available online: https://www.jmp.com/en_us/home.html (accessed on 16 April 2020).
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef]
- Yi, J.; Lo, L.S.H.; Liu, H.; Qian, P.-Y.; Cheng, J. Study of Heavy Metals and Microbial Communities in Contaminated Sediments Along an Urban Estuary. Front. Mar. Sci. 2021, 8, 1–17. [Google Scholar] [CrossRef]
- Ranjan, R.K.; Sinha, A.K.; Gupta, D.; Sappal, S.M.; Kumar, A. Sedimentary Geochemistry of Kabar Tal wetland, Begusarai, Bihar, India. J. Appl. Geochem. 2016, 18, 414–429. [Google Scholar]
- Yu, L.; Kaiyi, S.; Jie, Y.; Qiyu, K. Evaluation of Heavy Metal Pollutants from Plateau Mines in Wetland Surface Deposits. Front. Environ. Sci. 2021, 8, 557302. [Google Scholar] [CrossRef]
- Pradit, S.; Wattayakorn, G.; Angsupanich, S.; Baeyens, W.; Leermakers, M. Distribution of trace elements in sediments and biota of Songkhla Lake, Southern Thailand. Water. Air. Soil Pollut. 2010, 206, 155–174. [Google Scholar] [CrossRef]
- Knox, A.S.; Dunn, D.L.; Paller, M.H.; Nelson, E.A.; Specht, W.L.; Seaman, J.C. Assessment of contaminant retention in constructed wetland sediments. Eng. Life Sci. 2006, 6, 31–36. [Google Scholar] [CrossRef]
- Jiménez-Ballesta, R.; García-Navarro, F.J.; Bravo, S.; Amorós, J.A.; Pérez-de-los-Reyes, C.; Mejías, M. Environmental assessment of potential toxic trace element contents in the inundated floodplain area of Tablas de Daimiel wetland (Spain). Environ. Geochem. Health 2017, 39, 1159–1177. [Google Scholar] [CrossRef]
- Bachand, P.; Horne, A. Denitrification in constructed free-water surface wetlands: II. Effects of vegetation and temperature. Ecol. Eng. 2000, 14, 17–32. [Google Scholar] [CrossRef]
- Reilly, J.F.; Horne, A.J.; Miller, C.D. Nitrate removal from a drinking water supply with large free-surface constructed wetlands prior to groundwater recharge. Ecol. Eng. 2000, 14, 33–47. [Google Scholar] [CrossRef]
- Shannon, R.D.; Flite, O.P.; Hunter, M.S. Subsurface flow constructed wetland performance at a Pennsylvania campground and conference center. J. Environ. Qual. 2000, 29, 2029–2036. [Google Scholar] [CrossRef]
- Walker, C.W.; Shannon, R.D. Nitrate and phosphate removal effects of compost amendments in wetland mesocosms. Am. Soc. Agric. Biol. Eng. 2006, 49, 1773–1778. [Google Scholar] [CrossRef]
- Chen, J.; He, F.; Zhang, X.; Sun, X.; Zheng, J.; Zheng, J. Heavy metal pollution decreases microbial abundance, diversity and activity within particle-size fractions of a paddy soil. FEMS Microbiol. Ecol. 2014, 87, 164–181. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.M.; Lyautey, E.; Bonnineau, C.; Dabrin, A.; Pesce, S. Environmental concentrations of copper, alone or in mixture with arsenic, can impact river sediment microbial community structure and functions. Front. Microbiol. 2018, 9, 1852. [Google Scholar] [CrossRef]
- Hemme, C.L.; Deng, Y.; Gentry, T.J.; Fields, M.W.; Wu, L.; Barua, S.; Barry, K.; Tringe, S.G.; Watson, D.B.; He, Z.; et al. Metagenomic insights into evolution of a heavy metal-contaminated groundwater microbial community. ISME J. 2010, 4, 660–672. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.; Van Nostrand, J.D.; Gough, H.L.; He, Z.; Hazen, T.C.; Stahl, D.A.; Zhou, J. Functional gene array-based analysis of microbial communities in heavy metals-contaminated lake sediments. FEMS Microbiol. Ecol. 2013, 86, 200–214. [Google Scholar] [CrossRef] [PubMed]
- Olson, B.H.; Thornton, I. The resistance patterns to metals of bacterial population in contaminated land. J. Soil Sci. 1982, 33, 271–277. [Google Scholar] [CrossRef]
- Junca, H.; Pieper, D.H. Functional gene diversity analysis in BTEX contaminated soils by means of PCR-SSCP DNA fingerprinting: Comparative diversity assessment against bacterial isolates and PCR-DNA clone libraries. Environ. Microbiol. 2004, 6, 95–110. [Google Scholar] [CrossRef]
- Anderson, J.A.H.; Hooper, M.J.; Zak, J.C.; Cox, S.B. Molecular and functional assessment of bacterial community convergence in metal-amended soils. Microb. Ecol. 2009, 58, 10–22. [Google Scholar] [CrossRef] [PubMed]
- Konopka, A.; Zakharova, T.; Bischoff, M.; Oliver, L.; Nakatsu, C.; Turco, R.F. Microbial biomass and activity in lead-contaminated soil. Appl. Environ. Microbiol. 1999, 65, 2256–2259. [Google Scholar] [CrossRef]
- Margesin, R.; Schinner, F. Bacterial heavy metal-tolerance—Extreme resistance to nickel in Arthrobacter spp. strains. J. Basic Microbiol. 1996, 36, 269–282. [Google Scholar] [CrossRef]
- Rensing, C.; Newby, D.T.; Pepper, I.L. The role of selective pressure and selfish DNA in horizontal gene transfer and soil microbial community adaptation. Soil Biol. Biochem. 2002, 34, 285–296. [Google Scholar] [CrossRef]
- Kulakovskaya, T. Phosphorus storage in Microorganisms: Diversity and Evolutionary Insight. Biochem. Physiol. Open Access 2014, 4, e130. [Google Scholar] [CrossRef]
- Ayangbenro, A.S.; Bababola, O.O.; Aremu, O.S. Bioflocculant production and heavy metal sorption by metal resistant bacterial isolates from gold mining soil. Chemosphere 2019, 231, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Liu, J.; Wang, Z.; Dong, L.; Fan, J.; Qu, J. Remediation of Pb-Resistant Bacteria to Pb Polluted Soil. J. Environ. Prot. 2011, 2, 130–141. [Google Scholar] [CrossRef]
- Figueiredo, N.L.L.; Canário, J.; Duarte, A.; Serralheiro, M.L.; Carvalho, C. Isolation and characterization of mercury-resistant bacteria from sediments of tagus estuary (Portugal): Implications for environmental and human health risk assessment. J. Toxicol. Environ. Health Part A Curr. Issues 2014, 77, 155–168. [Google Scholar] [CrossRef] [PubMed]
- Gummersheimer, B.S.; Giblin, T. Identification of Lead Resistant Bacteria from a Heavily Contaminated Site. Jstor 2003, 74, 48–54. Available online: https://www.jstor.org/stable/4608669 (accessed on 2 December 2020).
- Kafilzadeh, F.; Afrough, R.; Johari, H.; Tahery, Y. Range determination for resistance / tolerance and growth kinetic of indigenous bacteria isolated from lead contaminated soils near gas stations (Iran). Eur. J. Exp. Biol. 2012, 2, 62–69. [Google Scholar]
- Tipayno, S.C.; Truu, J.; Samaddar, S.; Truu, M.; Preem, J.K.; Oopkaup, K.; Espenberg, M.; Chatterjee, P.; Kang, Y.; Kim, K.; et al. The bacterial community structure and functional profile in the heavy metal contaminated paddy soils, surrounding a nonferrous smelter in South Korea. Ecol. Evol. 2018, 8, 6157–6168. [Google Scholar] [CrossRef]
- Zhang, M.J.; Huang, F.K.; Wang, G.Y.; Liu, X.Y.; Wen, J.K.; Zhang, X.S.; Huang, Y.S.; Xia, Y. Geographic distribution of cadmium and its interaction with the microbial community in the Longjiang River: Risk evaluation after a shocking pollution accident. Sci. Rep. 2017, 7, 227. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Huang, Z.; He, L.; Sheng, X. Assessment of bacterial communities and characterization of lead-resistant bacteria in the rhizosphere soils of metal-tolerant Chenopodium ambrosioides grown on lead-zinc mine tailings. Chemosphere 2012, 87, 1171–1178. [Google Scholar] [CrossRef] [PubMed]
- Altimira, F.; Yáñez, C.; Bravo, G.; González, M.; Rojas, L.A.; Seeger, M. Characterization of copper-resistant bacteria and bacterial communities from copper-polluted agricultural soils of central Chile. BMC Microbiol. 2012, 12, 193. [Google Scholar] [CrossRef]
- Andreazza, R.; Pieniz, S.; Okeke, B.C.; Camargo, F.A.O. Evaluation of copper resistant bacteria from vineyard soils and mining waste for copper biosorption. Braz. J. Microbiol. 2011, 42, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Berg, J.; Brandt, K.K.; Al-Soud, W.A.; Holm, P.E.; Hansen, L.H.; Sørensen, S.J.; Nybroe, O. Selection for Cu-tolerant bacterial communities with altered composition, but unaltered richness, via long-term cu exposure. Appl. Environ. Microbiol. 2012, 78, 7438–7446. [Google Scholar] [CrossRef] [PubMed]
- He, L.Y.; Zhang, Y.F.; Ma, H.Y.; Su, L.N.; Chen, Z.J.; Wang, Q.Y.; Qian, M.; Sheng, X.F. Characterization of copper-resistant bacteria and assessment of bacterial communities in rhizosphere soils of copper-tolerant plants. Appl. Soil Ecol. 2010, 44, 49–55. [Google Scholar] [CrossRef]
- Kunito, T.; Nagaoka, K.; Tada, N.; Senoo, K.; Oyaizu, H.; Matsumoto, S.; Saeki, K. Characterization of cu-resistant bacterial communities in cu-contaminated soils. Soil Sci. Plant Nutr. 1997, 43, 709–717. [Google Scholar] [CrossRef]
- Santo, C.E.; Morais, P.V.; Grass, G. Isolation and characterization of bacteria resistant to metallic copper surfaces. Appl. Environ. Microbiol. 2010, 76, 1341–1348. [Google Scholar] [CrossRef]
- Li, L.G.; Xia, Y.; Zhang, T. Co-occurrence of antibiotic and metal resistance genes revealed in complete genome collection. ISME J. 2017, 11, 651–662. [Google Scholar] [CrossRef] [PubMed]
- Boyd, E.S.; Barkay, T. The mercury resistance operon: From an origin in a geothermal environment to an efficient detoxification machine. Front. Microbiol. 2012, 3, 349. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Li, J.; Zhang, H.; Shi, W.; Liu, Y. Bacterial heavy-metal and antibiotic resistance genes in a copper tailing dam area in northern China. Front. Microbiol. 2019, 10, 1916. [Google Scholar] [CrossRef] [PubMed]










| Land Use Percent in Pike River Watershed | ||||||
|---|---|---|---|---|---|---|
| Wetland Site | Watershed Area (ha) | Percent | Percent | Percent | Percent | Percent |
| Residential | Commercial | Industrial | Agricultural | Underdeveloped | ||
| 1 | 104.45 | 11 | 15.1 | 12.1 | 61.6 | 0 |
| 2 | 334.18 | 42.3 | 0 | 0 | 57.5 | 0 |
| 3 | 493.72 | 15.7 | 14.2 | 20.08 | 0 | 49.3 |
| 4 | 720 | 0 | 72.2 | 20.2 | 0 | 7.2 |
| (A) Metal Microcosm | |||
| Test Sediment | |||
| Metal Added to Water | Control | Low Concentration | High Concentration |
| Cu as CuSO4 | 0 mg/L | 0.05 mg/L | 0.15 mg/L |
| Pb as PbNO3 | 0 mg/L | 0.1 mg/L | 0.3 mg/L |
| (B) Nutrient Microcosm | |||
| Test Sediment | |||
| Nutrient Added to Water | Control | Low Concentration | High Concentration |
| NO3− as KNO3 | 0 mg/L | 5.0 mg/L | 15.0 mg/L |
| PO43− as Na2HPO4 | 0 mg/L | 1.0 mg/L | 3.0 mg/L |
| Genera Shannon | Genera Simpson | |||||
|---|---|---|---|---|---|---|
| Diversity Index | Diversity Index | |||||
| Parameter | Estimate | p-Value | R2 | Estimate | p-Value | R2 |
| Concentration of Pb (mg/L) in water | 254 | <0.0001 * | 0.64 | 10187 | <0.0001 * | 0.64 |
| Concentration of Cu (mg/L) in water | −116 | <0.0001 * | 1797 | <0.0001 * | ||
| Genera Detected in Metal Microcosm | |||
|---|---|---|---|
| Intolerant | Sensitive | Tolerant | |
| Less tolerant | Highly tolerant | ||
| Methylobacterium, Microbacterium, Shewanella, Cupriavidus, Enterococcus, Rhizobium, Enterobacter, Corynebacterium, Chlorobium, Exiguobacterium, Psychrobacter | Aquicella, Agrobacterium, Gemmata, Sphingomonas, Nitrospira, Balneimonas, Chlorobaculum, Aeromonas, Pedosphaera, Arthrobacter | Aquicella, Candidatus Solibacter, Burkholderia, GOUTA19, Holophaga, Sphingomonas, Syntrophus, Anaerolinea, Geobacter, Cystobacter, Desulfococcus, Syntrophobacter, Paludibacter, Nitrospira, Methylocaldum, WCHB1-05, C1_B004, Candidatus_Xiphinematobacter, Synechococcus, Balneimonas, Trichococcus, Mycobacterium, Sulfurimonas, Epulopiscium, Ralstonia, Pedosphaera, Bdellovibrio, Flavobacterium, Clostridium, SJA-88, Luteolibacter, Treponema, Caldilinea, LCP-6, Thiobacillus, Rhodoplanes, Sulfuritalea, Rhodoferax, Methylotenera, Planctomyces, Rhodobacter, Spirochaeta, Halomonas, Gallionella, Roseomonas, PSB-M-3, Dechloromonas, Bacillus, Devosia, Hydrogenophaga, Sediminibacterium, Phenylobacterium, Sulfuricurvum, Pedobacter, Hyphomicrobium, Pseudomonas | Nocardioides, Holophaga, Gemmata, Syntrophus, Bacteroides, Anaerolinea, Geobacter, Candidatus_Solibacter, Cystobacter, Desulfococcu, Syntrophobacter, GOUTA9, Paludibacter, Nitrospira, Clostridium, Blvii28, Methylocaldum, Synechococcus, Candidatus_Xiphinematobacter, WCHB1-05, Aeromonas, Luteolibacter, C1_B004, Methylibium, Anaerovorax, Treponema, Sulfurimonas, Chlorobaculum, Epulopiscium, Caldilinea, Pedosphaera, Bdellovibrio, Flavobacterium, SJA-88, Blvii28, LCP-6, Thiobacillus, Rhodoplanes, Sulfuritalea, Rhodoferax, Methylotenera, Rhodobacter, Gemmatimonas, Spirochaeta, Halomonas, Gallionella, Roseomonas, PSB-M-3, Fusibacter, Dechloromonas, Bacillus, Devosia, Sediminibacterium, Hydrogenophaga, Prosthecobacter, Phenylobacterium, Sulfuricurvum, Arthrobacter, Pedobacter, Hyphomicrobium, Pseudomonas, Ralstonia |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghosh Roy, S.; Wimpee, C.F.; McGuire, S.A.; Ehlinger, T.J. Responses of Bacterial Taxonomical Diversity Indicators to Pollutant Loadings in Experimental Wetland Microcosms. Water 2022, 14, 251. https://doi.org/10.3390/w14020251
Ghosh Roy S, Wimpee CF, McGuire SA, Ehlinger TJ. Responses of Bacterial Taxonomical Diversity Indicators to Pollutant Loadings in Experimental Wetland Microcosms. Water. 2022; 14(2):251. https://doi.org/10.3390/w14020251
Chicago/Turabian StyleGhosh Roy, Subhomita, Charles F. Wimpee, S. Andrew McGuire, and Timothy J. Ehlinger. 2022. "Responses of Bacterial Taxonomical Diversity Indicators to Pollutant Loadings in Experimental Wetland Microcosms" Water 14, no. 2: 251. https://doi.org/10.3390/w14020251
APA StyleGhosh Roy, S., Wimpee, C. F., McGuire, S. A., & Ehlinger, T. J. (2022). Responses of Bacterial Taxonomical Diversity Indicators to Pollutant Loadings in Experimental Wetland Microcosms. Water, 14(2), 251. https://doi.org/10.3390/w14020251