
Otolith Fingerprints and Tissue Stable Isotope Information Enable Allocation of Juvenile Fishes to Different Nursery Areas


 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
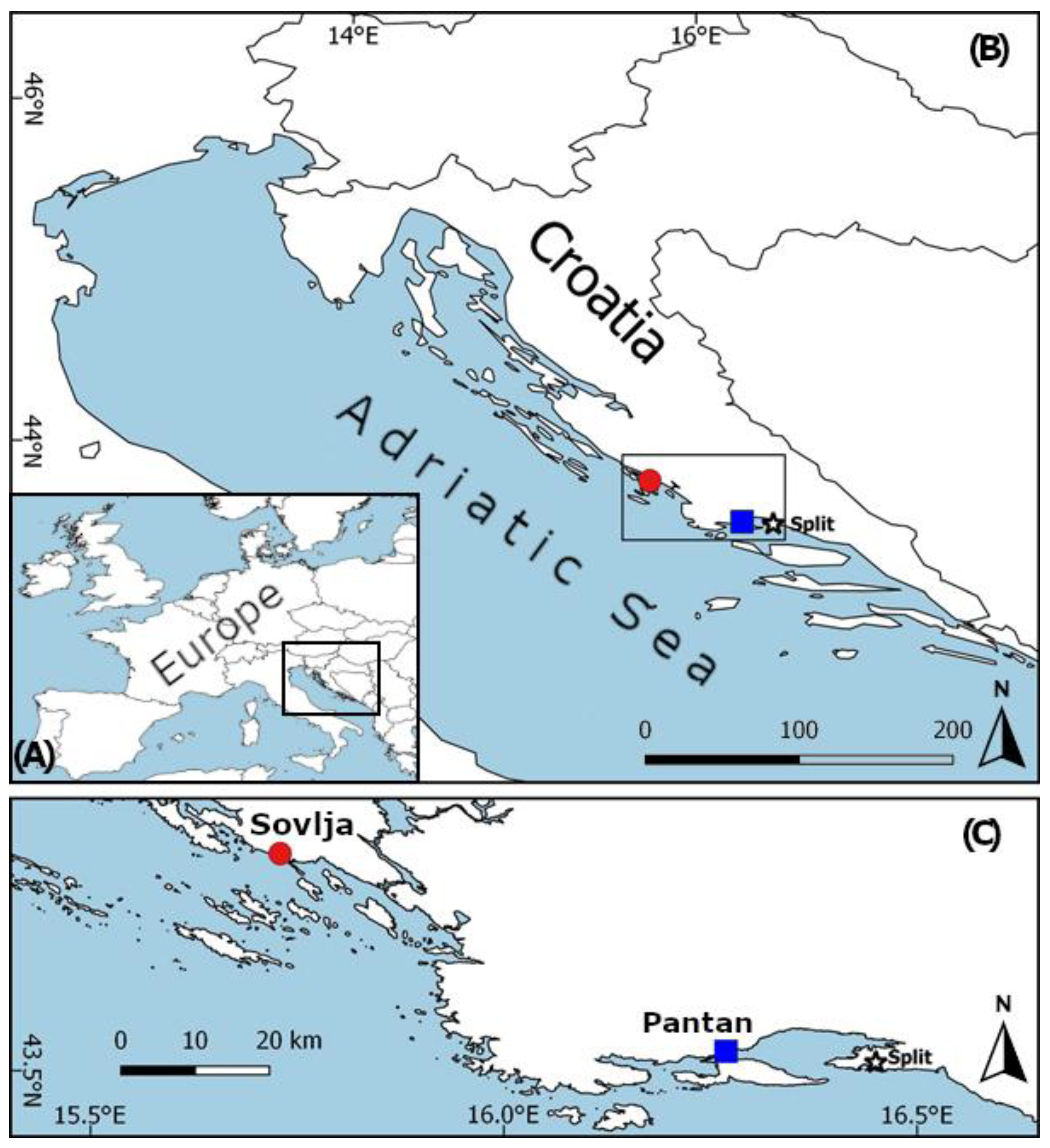
2.1. Study Locations and Fish Collection
2.2. Sample Preparation
2.3. Element and Stable Isotopes Analyses
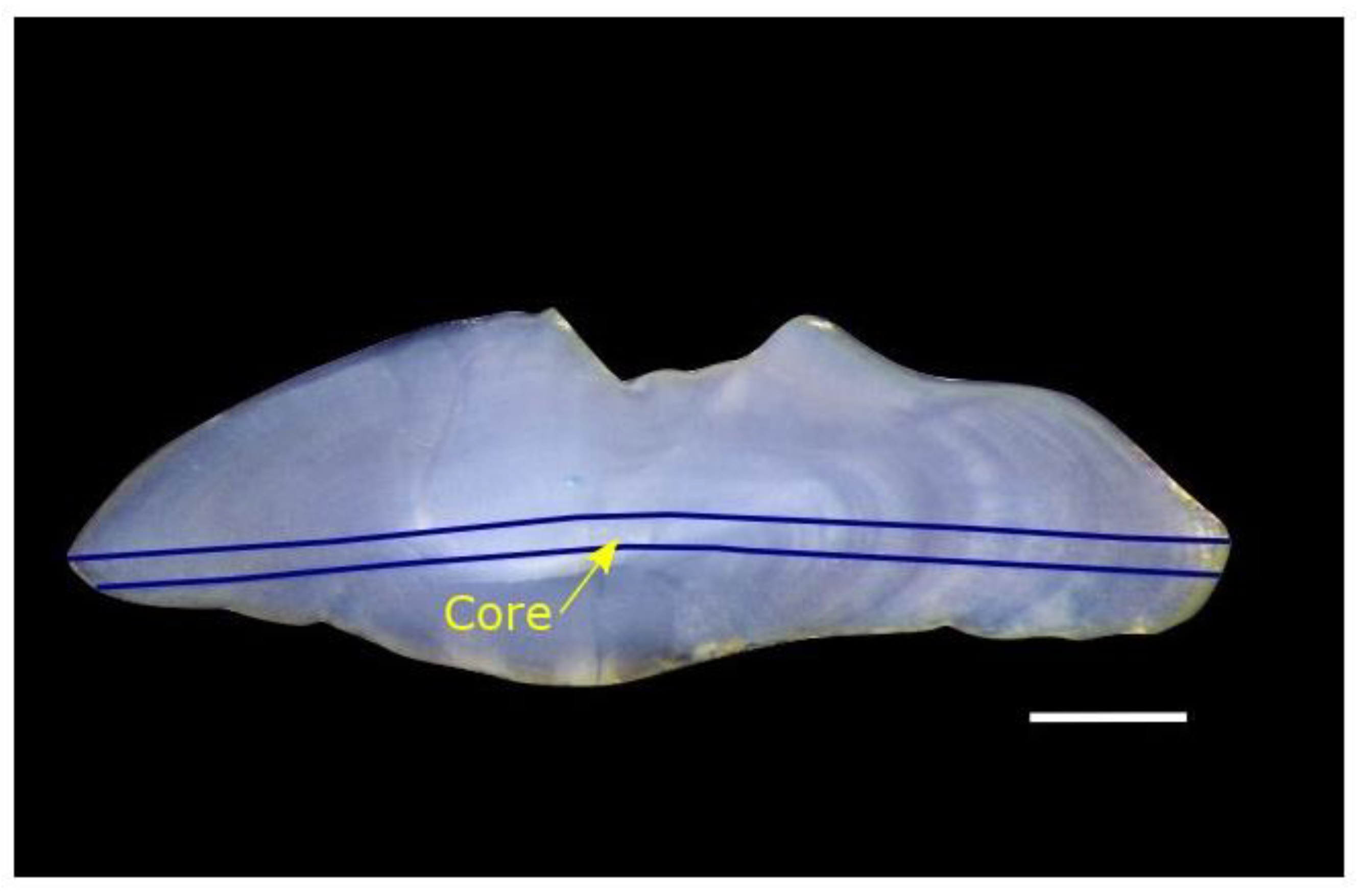
2.3.1. LA-ICP-MS Analysis of Otoliths
2.3.2. Stable Isotope Analyses of Muscle Tissue
2.4. Data Analysis
3. Results
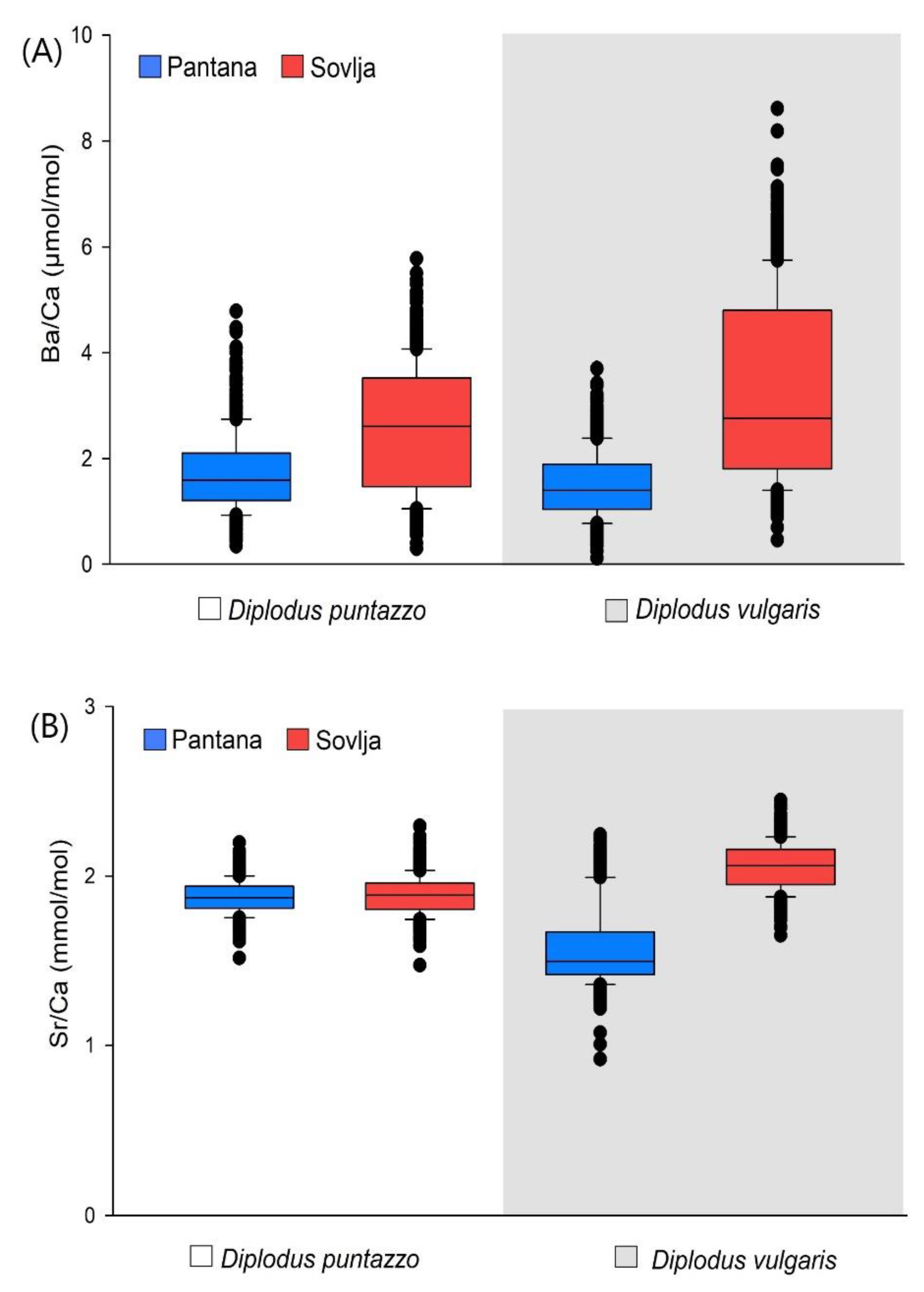
3.1. Otolith Trace Element Chemistry
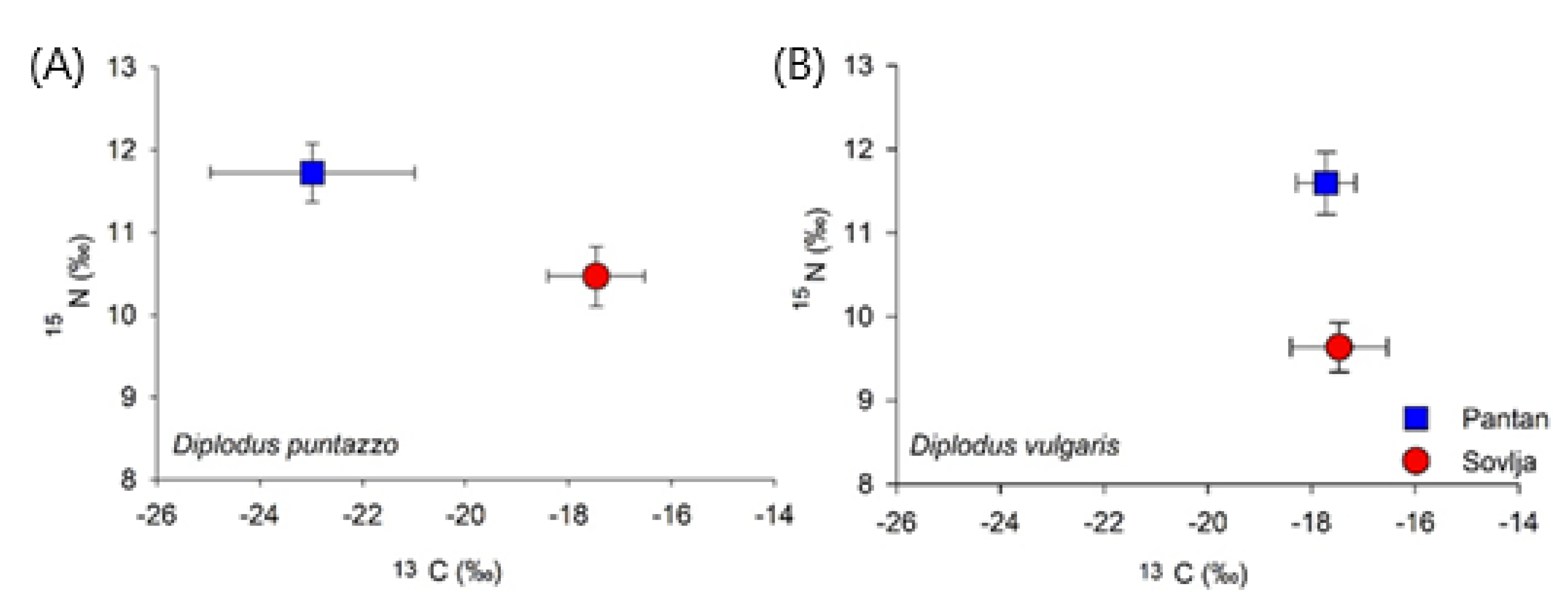
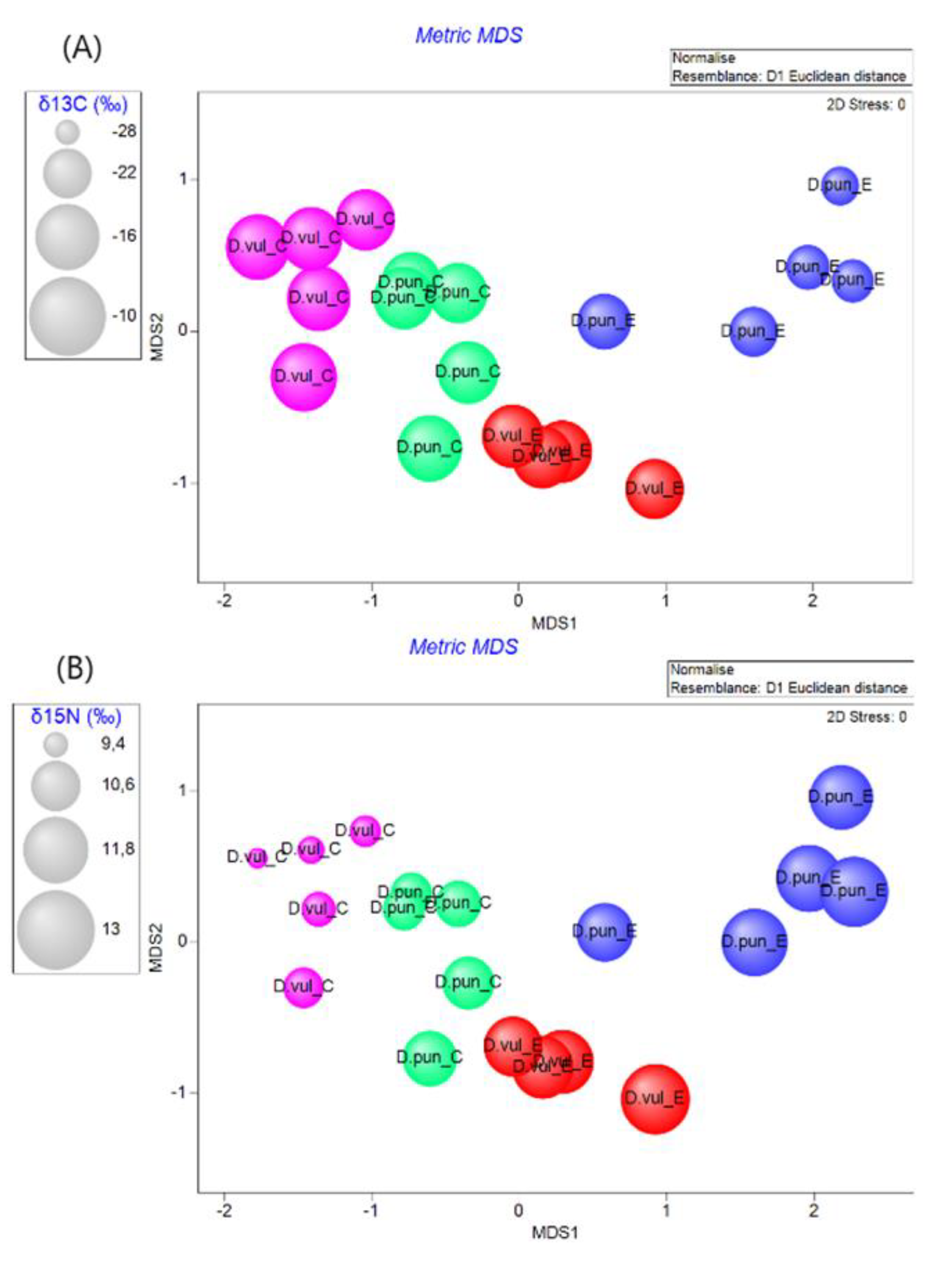
3.2. Stable Isotope Analyses
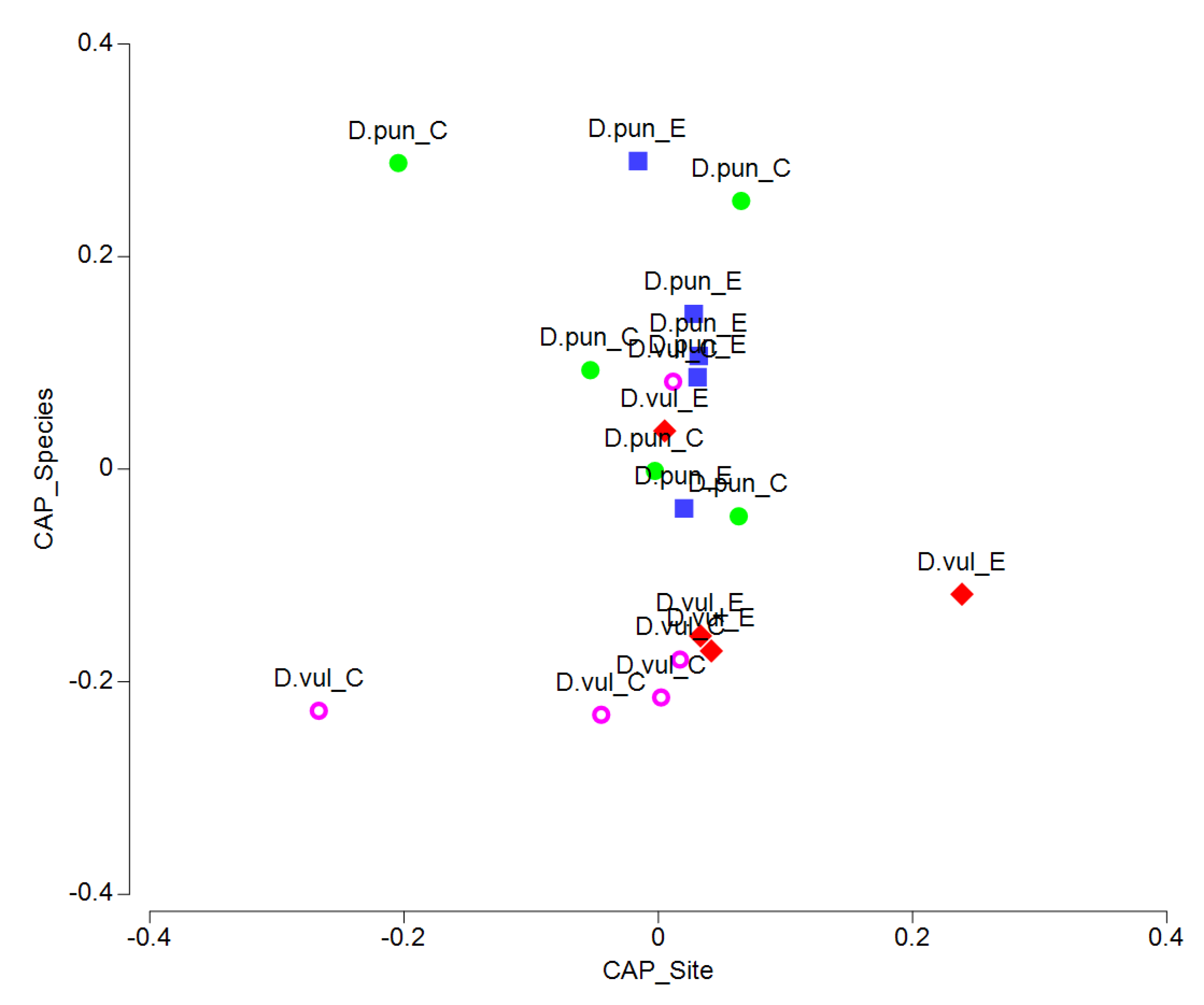
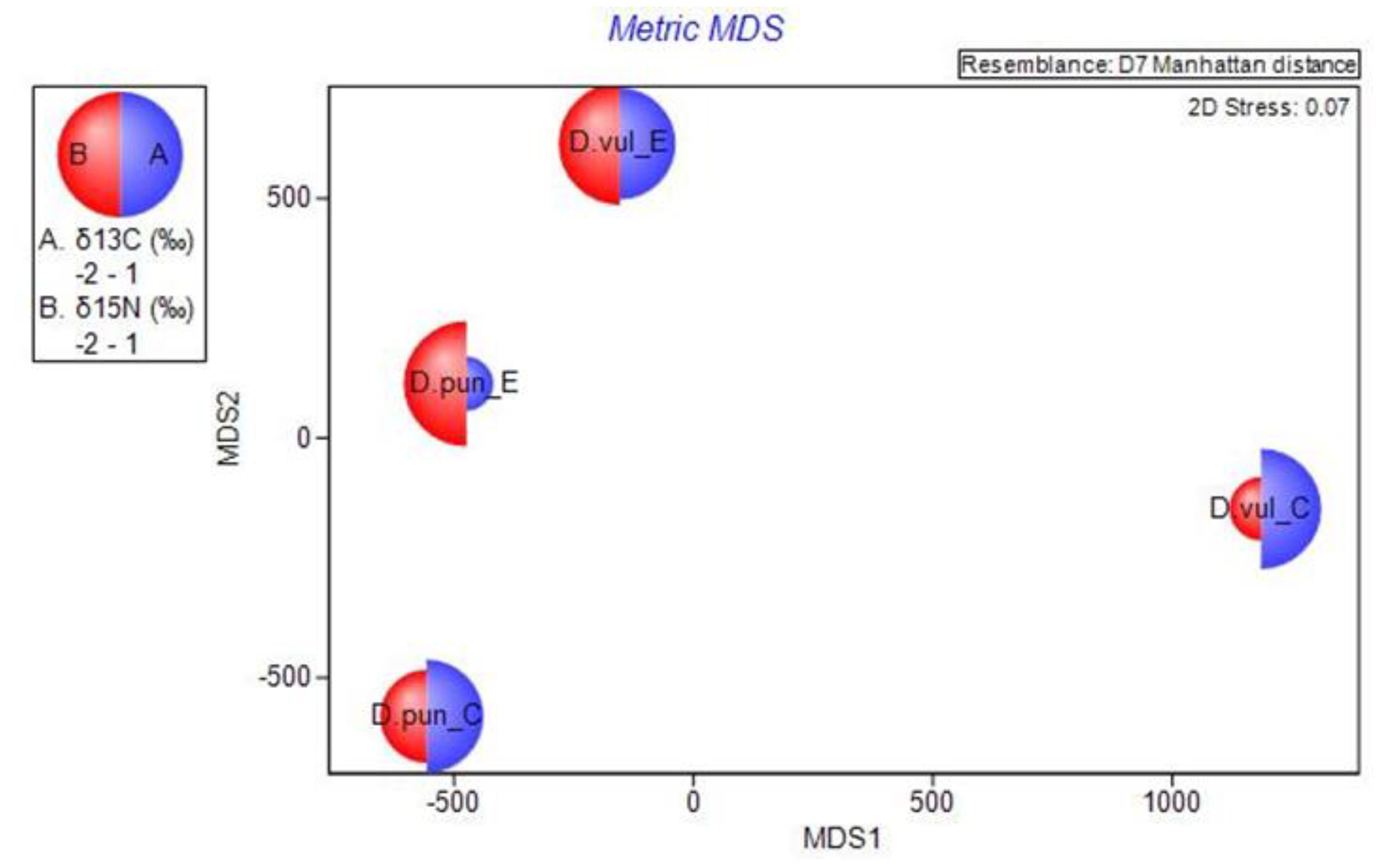
3.3. Multi-parameter Comparison
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Elsdon, T.S.; Wells, B.K.; Campana, S.E.; Gillanders, B.M.; Jones, C.M.; Limburg, K.E.; Secor, D.H.; Thorrold, S.R.; Walther, B.D. Otolith chemistry to describe movements and life-history parameters of fishes: Hypotheses, assumptions, limitations and inferences. Oceanogr. Mar. Biol. 2008, 46, 297–330. [Google Scholar]
- Catalán, I.A.; Alós, J.; Díaz-Gil, C.; Pérez-Mayol, S.; Basterretxea, G.; Morales-Nin, B.; Palmer, M. Potential fishing-related effects on fish life history revealed by otolith microchemistry. Fish. Res. 2018, 199, 186–195. [Google Scholar] [CrossRef]
- Darnaude, A.M.; Hunter, E. Validation of otolith δ18O values as effective natural tags for shelf-scale geolocation of migrating fish. Mar. Ecol. Prog. Ser. 2018, 598, 167–185. [Google Scholar] [CrossRef]
- Beck, M.W.; Heck, K.L.; Able, K.W.; Childers, D.L.; Eggleston, D.B.; Gillanders, B.M.; Halpern, B.; Hays, C.G.; Hoshino, K.; Minello, T.J.; et al. The identification, conservation, and management of estuarine and marine nurseries for fish and invertebrates. Bioscience 2001, 51, 633–641. [Google Scholar] [CrossRef]
- Dahlgren, C.P.; Todd Kellison, G.; Adams, A.J.; Gillanders, B.M.; Kendall, M.S.; Layman, C.A.; Ley, J.A.; Nagelkerken, I.; Serafy, J.E. Marine nurseries and effective juvenile habitats: Concepts and applications. Mar. Ecol. Prog. Ser. 2006, 312, 291–295. [Google Scholar] [CrossRef]
- Lotze, H.K.; Lenihan, H.S.; Bourque, B.J.; Bradbury, R.H.; Cooke, R.G.; Kay, M.C.; Kidwell, S.M.; Kirby, M.X.; Peterson, C.H.; Jackson, J.B.C. Depletion degradation, and recovery potential of estuaries and coastal seas. Science 2006, 312, 1806–1809. [Google Scholar] [CrossRef]
- Worm, B.; Barbier, E.B.; Beaumont, N.; Duffy, J.E.; Folke, C.; Halpern, B.S.; Jackson, J.B.C.; Lotze, H.K.; Micheli, F.; Palumbi, S.R.; et al. Impacts of biodiversity loss on ocean ecosystem services. Science 2006, 344, 787–790. [Google Scholar] [CrossRef]
- Waycott, M.; Duarte, C.M.; Carruthers, T.J.B.; Orth, R.J.; Dennison, W.C.; Olyarnik, S.; Calladine, A.; Fourqurean, J.W.; Heck, K.L., Jr.; Hughes, A.R.; et al. Accelerating loss of seagrasses across the globe threatens coastal ecosystems. Proc. Natl. Acad. Sci. USA 2009, 106, 12377–12381. [Google Scholar] [CrossRef]
- Claudet, J.; Fraschetti, S. Human-driven impacts on marine habitats: A regional meta-analysis in the Mediterranean Sea. Biol. Conserv. 2010, 143, 2195–2206. [Google Scholar] [CrossRef]
- Barausse, A.; Duci, A.; Mazzoldi, C.; Artioli, Y.; Palmeri, L. Trophic network model of the Northern Adriatic Sea: Analysis of an exploited and eutrophic ecosystem. Est. Coast. Shelf. Sci. 2009, 83, 577–590. [Google Scholar] [CrossRef]
- Teodósio, M.A.; Paris, C.B.; Wolanski, E.; Morais, P. Biophysical processes leading to the ingress of temperate fish larvae into estuarine nursery areas: A review. Estuar. Coast. Shelf Sci. 2016, 183, 187–202. [Google Scholar] [CrossRef]
- Rogers, T.A.; Fowler, A.J.; Steer, M.A.; Gillanders, B.M. Spatial connectivity during the early life history of a temperate marine fish inferred from otolith microstructure and geochemistry. Estuar. Coast. Shelf Sci. 2019, 227, 106342. [Google Scholar] [CrossRef]
- Cowen, R.K.; Lwiza, K.M.M.; Sponaugle, S.; Paris, C.B.; Olson, D.B. Connectivity of marine populations: Open or closed? Science 2000, 287, 857–859. [Google Scholar] [CrossRef]
- Cowen, R.K.; Sponaugle, S. Larval dispersal and marine population Connectivity. Ann. Rev. Mar. Sci. 2009, 1, 443–466. [Google Scholar] [CrossRef]
- Campana, S.E.; Thorrold, S.R. Otoliths, increments, and elements: Keys to a comprehensive understanding of fish populations? Can. J. Fish. Aquat. Sci. 2001, 58, 30–38. [Google Scholar] [CrossRef]
- Campana, S.E.; Chouinard, G.A.; Hanson, J.M.; Fréchet, A.; Brattey, J. Otolith elemental fingerprints as biological tracers of fish stocks. Fish. Res. 2000, 46, 343–357. [Google Scholar] [CrossRef]
- Tanner, S.E.; Vasconcelos, R.P.; Cabral, H.N.; Thorrold, S.R. Testing an otolith geochemistry approach to determine population structure and movements of European hake in the northeast Atlantic Ocean and Mediterranean Sea. Fish. Res. 2012, 125, 198–205. [Google Scholar] [CrossRef]
- Gillanders, B.M.; Kingsford, M.J. Elemental fingerprints of otoliths of fish may distinguish estuarine “nursery” habitats. Mar. Ecol. Prog. Ser. 2000, 201, 273–286. [Google Scholar] [CrossRef]
- Reis-Santos, P.; Tanner, S.E.; Vasconcelos, R.P.; Elsdon, T.S.; Cabral, H.N.; Gillanders, B.M. Connectivity between estuarine and coastal fish populations: Contributions of estuaries are not consistent over time. Mar. Ecol. Prog. Ser. 2013, 491, 177–186. [Google Scholar] [CrossRef]
- Reis-Santos, P.; Tanner, S.E.; França, S.; Vasconcelos, R.P.; Gillanders, B.M.; Cabral, H.N. Connectivity within estuaries: An otolith chemistry and muscle stable isotope approach. Ocean Coast. Manag. 2015, 118, 51–59. [Google Scholar] [CrossRef]
- Sadekov, A.; Eggins, S.M.; De Deckker, P. Characterization of Mg/Ca distributions in planktonic foraminifera species by electron microprobe mapping. Geochem. Geophys. Geosyst. 2005, 6. [Google Scholar] [CrossRef]
- Montagna, P.; McCulloch, M.; Mazzoli, C.; Silenzi, S.; Odorico, R. The non-tropical coral Cladocora caespitosa as the new climate archive for the Mediterranean: High-resolution (∼weekly) trace element systematics. Quat. Sci. Rev. 2007, 26, 441–462. [Google Scholar] [CrossRef]
- Sadekov, A.; Eggins, S.M.; De Deckker, P.; Ninnemann, U.; Kuhnt, W.; Bassinot, F. Surface and subsurface seawater temperature reconstruction using Mg/Ca microanalysis of planktonic foraminifera Globigerinoides ruber, Globigerinoides sacculifer, and Pulleniatina obliquiloculata. Paleoce. Paleoclim. 2009, 24, PA3201. [Google Scholar]
- Long, K.; Stern, N.; Williams, I.S.; Kinsley, L.; Wood, R.; Sporcic, K.; Fallon, S.; Kokkonen, H.; Moffat, I.; Grün, R. Fish otolith geochemistry, environmental conditions and human occupation at Lake Mungo, Australia. Quaternery Sci. Rev. 2014, 88, 82–95. [Google Scholar] [CrossRef]
- Fowler, A.M.; Smith, S.M.; Booth, D.J.; Stewart, J. Partial migration of grey mullet (Mugil cephalus) on Australia’s east coast revealed by otolith chemistry. Mar. Environ. Res. 2016, 119, 238–244. [Google Scholar] [CrossRef]
- Gillanders, B.M. Using elemental chemistry of fish otoliths to determine connectivity between estuarine and coastal habitats. Estuar. Coast. Shelf Sci. 2005, 64, 47–57. [Google Scholar] [CrossRef]
- Gillikin, D.P.; Wanamaker, A.D.; Andrus, C.F.T. Chemical sclerochronology. Chem. Geol. 2019, 526. [Google Scholar] [CrossRef]
- Secor, D.H.; Rooker, J.R. Is otolith strontium a useful scalar of life-cycles in estuarine fishes? Fish. Res. 2000, 46, 359–371. [Google Scholar] [CrossRef]
- Kraus, R.T.; Secor, D.H. Dynamics of white perch Morone americana population contingents in the Patuxent River estuary, Maryland, USA. Mar. Ecol. Prog. Ser. 2004, 279, 247–259. [Google Scholar] [CrossRef]
- Tabouret, H.; Lord, C.; Bareille, G.; Pécheyran, C.; Monti, D.; Keith, P. Otolith microchemistry in Sicydium punctatum: Indices of environmental condition changes after recruitment. Aqua. Liv. Res. 2011, 24, 369–378. [Google Scholar] [CrossRef]
- Izzo, C.; Reis-Santos, P.; Gillanders, B.M. Otolith chemistry does not just reflect environmental conditions: A meta-analytic evaluation. Fish Fish. 2018, 19, 441–454. [Google Scholar] [CrossRef]
- Green, B.C.; Smith, D.J.; Earley, S.E.; Hepburn, L.J.; Underwood, G.J.C. Seasonal changes in community composition and trophic structure of fish populations of five salt marshes along the Essex coastline, United Kingdom. Estuar. Coast. Shelf. Sci. 2009, 85, 1–10. [Google Scholar] [CrossRef]
- Correira, A.T.; Pipac, T.; Gonçalves, J.M.S.; Erzini, K.; Hamer, P.A. Insights into population structure of Diplodus vulgaris along the SW Portuguese coast from otolith elemental signatures. Fish. Res. 2011, 111, 82–91. [Google Scholar] [CrossRef]
- Bouchoucha, M.; Pécheyran, C.; Gonzalez, J.L.; Lenfant, P.; Darnaude, A.M. Otolith fingerprints as natural tags to identify juvenile fish life in ports. Estuar. Coast. Shelf Sci. 2018, 212, 210–218. [Google Scholar] [CrossRef]
- Hobson, K.A. Tracing origins and migration of wildlife using stable isotopes: A review. Oecologia. 1999, 120, 314–326. [Google Scholar] [CrossRef] [PubMed]
- Herzka, S.Z. Assessing connectivity of estuarine fishes based on stable isotope ratio analysis. Estuar. Coast. Shelf Sci. 2005, 64, 58–69. [Google Scholar] [CrossRef]
- Trueman, C.N.; Mackenzie, K.M.; Palmer, M.R. Identifying migrations in marine fishes through stable-isotope analysis. J. Fish Biol. 2012, 81, 826–847. [Google Scholar] [CrossRef]
- Suzuki, K.W.; Kasai, A.; Ohta, T.; Nakayama, K.; Tanaka, M. Migration of Japanese temperate bass Lateolabrax japonicus juveniles within the Chikugo River estuary revealed by δ13C analysis. Mar. Ecol. Prog. Ser. 2008, 358, 246–256. [Google Scholar] [CrossRef]
- Verweij, M.C.; Nagelkerken, I.; Hans, I.; Ruseler, S.M.; Mason, P.R.D. Seagrass nurseries contribute to coral reef fish populations. Limnol. Oceanogr. 2008, 53, 1540–1547. [Google Scholar] [CrossRef]
- Green, B.C.; Smith, D.J.; Grey, J.; Underwood, G.J.C. High site fidelity and low site connectivity in temperate salt marsh fish populations: A stable isotope approach. Oecologia 2012, 168, 245–255. [Google Scholar] [CrossRef]
- Vinagre, C.; Salgado, J.; Costa, M.J.; Cabral, H.N. Nursery fidelity, food web interactions and primary sources of nutrition of the juveniles of Solea solea and S. senegalensis in the Tagus estuary (Portugal): A stable isotope approach. Estuar. Coast. Shelf Sci. 2008, 76, 255–264. [Google Scholar] [CrossRef]
- Abecasis, D.; Bentes, D.; Erzini, K. Home range, residency and movements of Diplodus sargus and Diplodus vulgaris in a coastal lagoon: Connectivity between nursery and adult habitats. Estuar. Coast. Shelf Sci. 2009, 85, 55–529. [Google Scholar] [CrossRef]
- Fry, B. Using stable isotopes to monitor watershed influences on aquatic trophodynamics. Can. J. Fish. Aquat. Sci. 1999, 56, 2167–2171. [Google Scholar]
- Rubenstein, D.R.; Hobson, K.A. From birds to butterflies: Animal movement patterns and stable isotopes. Trends Ecol. Evol. 2004, 19, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Lawton, R.J.; Wing, S.R.; Lewis, A.M. Evidence for discrete subpopulations of sea perch (Helicolenus ercoides) across four fjords in Fiordland, New Zealand. New Zeal. J. Mar. Fresh. Res. 2010, 44, 309–322. [Google Scholar] [CrossRef]
- Dierking, J.; Morat, F.; Letourneur, Y.; Harmelin-Vivien, M. Fingerprints of lagoonal life: Migration of the marine flatfish Solea solea assessed by stable isotopes and otolith microchemistry. Estuar. Coast. Shelf Sci. 2012, 104, 23–32. [Google Scholar] [CrossRef]
- Fodrie, F.J.; Herzka, S.Z. A Comparison of Otolith Geochemistry and Stable Isotope Markers to Track Fish Movement: Describing Estuarine Ingress by Larval and Post-Larval Halibut. Estaur. Coast. 2013, 36, 906–917. [Google Scholar]
- Marengo, M.; Durieux, E.D.H.; Marchand, B.; Francour, P. A review of biology, fisheries and population structure of Dentex dentex (Sparidae). Rev. Fish. Biol. Fisheries 2014, 24, 1065–1088. [Google Scholar] [CrossRef]
- Divanach, P. Contribution de la Biologie et de l’élevage de 6 Sparidés Mediterranéens: Sparus aurata, Diplodus sargus, Diplodus vulgaris, Diplodus annularis, Lithognathus mormyrus, Puntazzo puntazzo (Poissons Téleosteens). Thèse d’Etat, Université des Sciences et Techniques de Languedoc, Montpellier, France, 1985; p. 479. [Google Scholar]
- MacPherson, E. Ontogenetic shifts in habitat use and aggregation in juvenile sparid fishes. J. Exp. Mar. Bio. Ecol. 1998, 220, 127–150. [Google Scholar]
- Vigliola, L.; Harmelin-Vivien, M.L.; Biagi, E.; Galzin, R.; Garcia-Rubies, A.; Harmelin, J.G.; Jouvenel, J.Y.; Le Direach-Boursier, L.; Macpherson, E.; Tunesi, L. Spatial and temporal patterns of settlement among sparid fishes of the genus Diplodus in the north-western Mediterranean. Mar. Ecol. Prog. Ser. 1998, 168, 45–56. [Google Scholar] [CrossRef]
- Mouine, N.; Francour, P.; Ktari, M.H.; Chakroun-Marzouk, N. Reproductive biology of four Diplodus species Diplodus vulgaris, D. annularis, D. sargus sargus and D. puntazzo (Sparidae) in the Gulf of Tunis (central Mediterranean). J. Mar. Biol. Ass. UK. 2012, 92, 623–631. [Google Scholar]
- Dulčić, J.; Kraljević, M.; Grbec, B.; Pallaoro, A. Composition and temporal fluctuations of inshore juvenile fish populations in the Kornati Archipelago, eastern middle Adriatic. Mar. Biol. 1997, 129, 267–277. [Google Scholar] [CrossRef]
- Di Franco, A.; Bulleri, F.; Pennetta, A.; De Benedetto, G.; Clarke, K.R.; Guidetti, P. Within-Otolith Variability in Chemical Fingerprints: Implications for Sampling Designs and Possible Environmental Interpretation. PLoS ONE 2014, 9, e101701. [Google Scholar] [CrossRef]
- Vasconcelos, R.P.; Reis-Santos, P.; Maia, A.; Fonseca, V.; França, S.; Wouters, N.; Costa, M.J.; Cabral, H.N. Nursery use patterns of commercially important marine fish species in estuarine systems along the Portuguese coast. Estuar. Coast. Shelf Sci. 2010, 86, 613–624. [Google Scholar] [CrossRef]
- Zeigler, J.M.; Whitledge, G.W. Otolith trace element and stable isotopic compositions differentiate fishes from the Middle Mississippi River, its tributaries, and floodplain lakes. Hydrobiologia 2011, 661, 289–302. [Google Scholar] [CrossRef]
- Gibb, F.M.; Régnier, T.; Donald, K.; Wright, P.J. Connectivity in the early life history of sandeel inferred from otolith microchemistry. J. Sea Res. 2017, 119, 8–16. [Google Scholar] [CrossRef]
- Avigliano, E.; Pisonero, J.; Dománico, A.; Silva, N.; Sánchez, S.; Vanina Volpedo, A. Spatial segregation and connectivity in young and adult stages of Megaleporinus obtusidens inferred from otolith elemental signatures: Implications for management. Fish. Res. 2018, 204, 239–244. [Google Scholar] [CrossRef]
- Ley, L.A.; Rolls, H.J. Using otolith microchemistry to assess nursery habitat contribution and function at a fine spatial scale. Mar. Ecol. Prog. Ser. 2018, 606, 151–173. [Google Scholar] [CrossRef]
- Dulčić, J.; Matić, S.; Kraljević, M. Shallow coves as nurseries for non-resident fish: A case study in the eastern middle Adriatic. J. Mar. Biol. Ass. U.K. 2002, 82, 991–993. [Google Scholar] [CrossRef]
- Dulčić, J.; Matić-Skoko, S.; Kraljević, M.; Fencil, M.; Glamuzina, B. Seasonality of a fish assemblage in shallow waters of Duće-Glava, eastern middle Adriatic. Cybium 2005, 29, 57–63. [Google Scholar]
- Matić-Skoko, S.; Kraljević, M.; Dulčić, J.; Pallaoro, A.; Lučić, D.; Glamuzina, B. Growth of juvenile sharpsnout seabream, Diplodus puntazzo (Teleostei: Sparidae) in the Kornati Archipelago, eastern Adriatic Sea. Vie Milieu. 2007, 57, 13–19. [Google Scholar]
- Fukumori, K.; Oi, M.; Doi, H.; Takahashi, D.; Okuda, N.; Miller, T.W. Bivalve tissue as a carbon and nitrogen isotope baseline indicator in coastal ecosystems. Estuar. Coast. Shelf Sci. 2008, 79, 45–50. [Google Scholar] [CrossRef]
- Post, D.M. Using stable isotopes to estimate trophic position: Models, methods, and assumptions. Ecology 2002, 83, 703–718. [Google Scholar] [CrossRef]
- Post, D.M.; Layman, C.A.; Arrington, D.A.; Takimoto, G.; Quattrochi, J.; Montaña, C.G. Getting to the fat of the matter: Models, methods and assumptions for dealing with lipids in stable isotope analyses. Oecologia 2007, 152, 179–189. [Google Scholar] [CrossRef]
- Available online: http://georem.mpch-mainz.gwdg.de/ (accessed on 15 May 2020).
- Jochum, K.P.; Nohl, U.; Herwig, K.; Lammel, E.; Stoll, B.; Hofmann, A.W. GeoReM: A new geochemical database for reference materials and isotopic standards. Geostand. Geoanalytical Res. 2005, 29, 333–338. [Google Scholar] [CrossRef]
- Jochum, K.P.; Weis, U.; Stoll, B.; Kuzmin, D.; Yang, Q.; Raczek, I.; Jacob, D.E.; Stracke, A.; Birbaum, K.; Frick, D.A.; et al. Determination of reference values for NIST SRM 610-617 glasses following ISO guidelines. Geostand. Geoanalytical Res. 2011, 36, 397–429. [Google Scholar] [CrossRef]
- Jochum, K.P.; Scholz, D.; Stoll, B.; Weis, U.; Wilson, S.A.; Yang, Q.; Schwalb, A.; Börner, N.; Jacob, D.E.; Andreae, M.O. Accurate trace element analysis of speleothems and biogenic calcium carbonates by LA-ICP-MS. Chem. Geol. 2012, 318–319, 31–44. [Google Scholar] [CrossRef]
- Jochum, K.P.; Stoll, B.; Herwig, K.; Willbold, M. Validation of LA-ICP-MS trace element analysis of geological glasses using a new solid-state 193 nm Nd:YAG laser and matrix-matched calibration. J. Anal. At. Spectrom. 2007, 22, 112–121. [Google Scholar] [CrossRef]
- Mischel, S.A.; Mertz-Kraus, R.; Jochum, K.P.; Scholz, D. TERMITE: An R script for fast reduction of laser ablation inductively coupled plasma mass spectrometry data and its application to trace element measurements. Rapid Commun. Mass Spectrom. 2017, 131, 1079–1087. [Google Scholar] [CrossRef]
- Yoshinaga, J.; Nakama, A.; Morita, M.; Edmonds, J.S. Fish otolith reference material for quality assurance of chemical analyses. Mar. Chem. 2000, 69, 91–97. [Google Scholar] [CrossRef]
- Sharp, Z. Principles of stable isotope geochemistry. Choice Rev. Online. 2007. [CrossRef]
- Vrdoljak, D.; Matić-Skoko, S.; Peharda, M.; Uvanović, H.; Markulin, K.; Mertz-Kraus, R. Otolith fingerprints reveals potential pollution exposure of newly settled juvenile Sparus aurata. Mar. Poll. Bull. 2020, 160, 111695. [Google Scholar] [CrossRef] [PubMed]
- Marali, S.; Schöne, B.R.; Mertz-Kraus, R.; Griffin, S.M.; Wanamaker, A.D., Jr.; Butler, P.G. Reproducibility of trace element time-series (Na/Ca, Mg/Ca, Mn/Ca, Sr/Ca, and Ba/Ca) within and between specimens of the bivalve Arctica islandica—a LA-ICP-MS line scan study. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2017, 484, 109–128. [Google Scholar] [CrossRef]
- Marali, S.; Schöne, B.R.; Mertz-Kraus, R.; Griffin, S.M.; Wanamaker, A.D., Jr.; Matras, U. Ba/Ca ratios in shells of Arctica islandica—potential environmental proxy and crossdating tool. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2017, 465, 347–361. [Google Scholar] [CrossRef]
- Anderson, M.J.; Willis, T.J. Canonical analysis of principal coordinates: A useful method of constrained ordination for ecology. Ecology 2003, 84, 511–525. [Google Scholar] [CrossRef]
- Dulčić, J.; Pallaoro, A.; Matić-Skoko, S.; Dragičević, B.; Tutman, P.; Grgičević, R.; Stagličić, N.; Bukvić, V.; Pavličević, J.; Glamuzina, B.; et al. Age, growth and mortality of common two-banded seabream, Diplodus vulgaris (Geoffroy Saint-Hilaire, 1817), in the eastern Adriatic Sea (Croatian coast). J. Appl. Icthyol. 2011, 27, 1254–1258. [Google Scholar] [CrossRef]
- Kraljević, M.; Matić-Skoko, S.; Dulčić, J.; Pallaoro, A.; Jardas, I.; Glamuzina, B. Age and growth of sharpsnout seabream Diplodus puntazzo (Cetti, 1777) in the eastern Adriatic Sea. Cah. Biol. Mar. 2007, 48, 145–154. [Google Scholar]
- Miller, J.A. Effects of water temperature and barium concentration on otolith composition along a salinity gradient: Implications for migratory reconstructions. J. Exp. Mar. Bio. Ecol. 2011, 405, 42–52. [Google Scholar] [CrossRef]
- Herrera-Reveles, A.T.; Lemus, M.; Marín, B.; Prin, J.L. Trace metal incorporation in otoliths of a territorial coral reef fish (Abudefduf saxatilis) as an environmental monitoring tool. E3S Web Conf. 2013, 1, 34007. [Google Scholar] [CrossRef]
- Sturrock, A.M.; Trueman, C.N.; Milton, J.A.; Waring, C.P.; Cooper, M.J.; Hunter, E. Physiological influences can outweigh environmental signals in otolith microchemistry research. Mar. Ecol. Prog. Ser. 2014, 500, 245–264. [Google Scholar] [CrossRef]
- Markulin, K.; Peharda, M.; Mertz-Kraus, R.; Schöne, B.R.; Uvanović, H.; Kovač, Ž.; Janeković, I. Trace and minor element records in aragonitic bivalve shells as environmental proxies. Chem. Geo. 2019, 507, 120–133. [Google Scholar] [CrossRef]
- Kalish, J.M. Otolith chemistry: Validation of the effects of physiology, age and environment on otolith composition. J. Exp. Mar. Biol. Ecol. 1989, 132, 151–178. [Google Scholar] [CrossRef]
- Kalish, J.M. Determinants of otolith chemistry: Seasonal variation in the composition of blood plasma, endolymph and otoliths of bearded rock cod Pseudophycis barbatus. Mar. Ecol. Prog. Ser. 1991, 74, 137–159. [Google Scholar] [CrossRef]
- Radtke, R.L.; Shafer, D.J. Environmental sensitivity of fish otolith microchemistry. Aust. J. Mar. Freshwater Res. 1992, 43, 935–951. [Google Scholar] [CrossRef]
- Sadovy, Y.; Severin, K. Elemental patterns in Red Hind (Epinephelus guttatus) otoliths from Bermuda and Puerto Rico reflect growth rate, not temperature. Can. J. Fish. Aquat. Sci. 1994, 51, 133–141. [Google Scholar] [CrossRef][Green Version]
- Tzeng, W.N. Temperature effects on the incorporation on strontium in otoliths of Japanese eel Anguilla japonica. J. Fish Biol. 1994, 45, 1055–1066. [Google Scholar] [CrossRef]
- Campana, S.E. Chemistry and composition of fish otoliths: Pathways, mechanisms and applications. Mar. Ecol. Prog. Ser. 1999, 188, 263–297. [Google Scholar] [CrossRef]
- Elsdon, T.S.; Gillanders, B.M. Reconstructing migratory patterns of fish based on environmental influences on otolith chemistry. Rev. Fish Biol. Fisher. 2003, 13, 219–235. [Google Scholar] [CrossRef]
- Hamer, P.A.; Jenkins, G.P.; Coutin, P. Barium variation in Pagrus auratus (Sparidae) otoliths: A potential indicator of migration between an embayment and ocean waters in south-eastern Australia. Estuar. Coast. Shelf Sci. 2006, 68, 686–702. [Google Scholar] [CrossRef]
- Walther, B.D.; Thorrold, S.R. Water, not food, contributes the majority of strontium and barium deposited in the otoliths of a marine fish. Mar. Ecol. Prog. Ser. 2006, 311, 125–130. [Google Scholar] [CrossRef]
- Davis, W.J. Contamination of coastal versus open ocean surface waters: A brief meta-analysis. Mar. Pollut. Bull. 1993, 26, 128–134. [Google Scholar] [CrossRef]
- Patterson, H.M.; Thorrold, S.R.; Shenker, J.M. Analysis of otolith chemistry in Nassau grouper (Epinephelus striatus) from the Bahamas and Belize using solution-based ICP-MS. Coral Reefs 1999, 18, 171–178. [Google Scholar] [CrossRef]
- Patterson, H.M.; Kingsford, M.J.; McCulloch, M.T. Elemental signatures of Pomacentrus coelestis otoliths at multiple spatial scales on the Great Barrier Reef, Australia. Mar. Ecol. Prog. Ser. 2004, 270, 229–239. [Google Scholar]
- Elsdon, T.S.; Gillanders, B.M. Temporal variability in strontium, calcium, barium, and manganese in estuaries: Implications for reconstructing environmental histories of fish from chemicals in calcified structures. Estuar. Coast. Shelf Sci. 2006, 66, 147–156. [Google Scholar]
- Bath, G.E.; Thorrold, S.R.; Jones, C.M.; Campana, S.E.; McLaren, J.W.; Lam, J.W.H. Strontium and barium uptake in aragonitic otoliths of marine fish. Geochim. Cosmochim. Acta. 2000, 64, 1705–1714. [Google Scholar] [CrossRef]
- Grønkjær, P. Otoliths as individual indicators: A reappraisal of the link between fish physiology and otolith characteristics. Mar. Fresh. Res. 2006, 67, 881–888. [Google Scholar] [CrossRef]
- Fry, B.; Baltz, D.M.; Benfield, M.C.; Fleeger, J.W.; Gace, A.; Haas, H.L.; Quiñones-Rivera, Z.J. Stable isotope indicators of movement and residency for brown shrimp (Farfantepenaeus aztecus) in coastal Louisiana marshscapes. Estuaries 2003, 26, 82–97. [Google Scholar] [CrossRef]
- Schlacher, T.A.; Liddell, B.; Gaston, T.F.; Schlacher-Hoenlinger, M. Fish track wastewater pollution to estuaries. Oecologia 2005, 144, 570–584. [Google Scholar] [PubMed]
- França, S.; Vasconcelos, R.P.; Tanner, S.; Maguas, C.; Costa, M.J.; Cabral, H.N. Assessing food web dynamics and relative importance of organic matter sources for fish species in two Portuguese estuaries: A stable isotope approach. Mar. Environ. Res. 2011, 72, 204–215. [Google Scholar] [CrossRef]
- Hesslein, R.H.; Hallard, K.A.; Ramlal, P. Replacement of sulfur, carbon, and nitrogen in tissue of growing broad whitefish (Coregonus nasus) in response to a change in diet traced by δ34S, δ13C, and δ15N. Can. J. Fish. Aqua. Sci. 1993, 50, 2071–2076. [Google Scholar] [CrossRef]
- MacNeil, M.A.; Drouillard, K.G.; Fisk, A.T. Variable uptake and elimination of stable nitrogen isotopes between tissues in fish. Can. J. Fish. Aquat. Sci. 2006, 63, 345–353. [Google Scholar]
- Yoshioka, T.; Wada, E. A stable isotope study on seasonal food web dynamics in a eutrophic lake. Ecology 1994, 75, 835–846. [Google Scholar]
- Elliot, M.; Quintino, V. The Estuarine Quality Paradox, Environmental Homeostasis and the difficulty of detecting anthropogenic stress in naturally stressed areas. Mar. Poll. Bull. 2007, 54, 640–645. [Google Scholar]
- Nunn, A.D.; Tewson, L.H.; Cowx, I.G. The foraging ecology of larval and juvenile fishes. Rev. Fish Biol. Fisheries 2012, 22, 377–408. [Google Scholar] [CrossRef]
- van Leeuwen, A.; Huss, M.; Gärdmark, A.; Casini, M.; Vitale, F.; Hjelm, J.; Persson, L.; de Roos, A.M. Predators with multiple ontogenetic niche shifts have limited potential for population growth and top-down control of their prey. American Naturalist. 2013, 182, 53–66. [Google Scholar] [CrossRef]
- Buljan, M.; Zore-Armanda, M. Oceanographic properties of the Adriatic Sea. Oceanogr. Mar. Biol. Ann. 1976, 14, 11–98. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Pantan | Sovlja | ||
|---|---|---|---|---|
| Bottom * | Surface | Bottom * | Surface | |
| Temperature (°C) | 27.8 | 26.5 | 24 | 26.4 |
| Salinity | 33.1 | 0.9 | 38.3 | 38 |
| Oxygen (mg/L) | 8.88 | 8.63 | 10.48 | 8.92 |
| Element | USG MACS-3 | USG BCR-2G | NIST SRM 610 | |||
|---|---|---|---|---|---|---|
| Measured values (µg/g) | Reference values (µg/g) | Measured values (µg/g) | Reference values (µg/g) | Measured values (µg/g) | Reference values (µg/g) | |
| Sr | 6181 ± 174 | 6760 ± 350 | 345.6 ± 1 | 342 ± 4 | 530.5 ± 7 | 515.5 ± 1 |
| Ba | 57.9 ± 2 | 58.7 ± 2 | 647.1 ± 5 | 683 ± 7 | 438.8 ± 9 | 452 ± 9 |
| Factors | (a) Sr and Ba | (b) δ13C and δ15N | |||||
|---|---|---|---|---|---|---|---|
| df | MS | Pseudo-F | P (perm) | MS | Pseudo-F | P (perm) | |
| Species | 1 | 9.588E + 05 | 2.239 | 0.049 | 6.517 | 19.325 | 0.0005 |
| Site | 1 | 6.134E + 05 | 1.432 | 0.227 | 19.936 | 59.111 | 0.0001 |
| Sp x Site | 1 | 6.495E + 05 | 1.157 | 0.199 | 3.394 | 10.064 | 0.0001 |
| Residauls | 15 | 4.283E + 05 | 0.337 | ||||
| Total | 18 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vrdoljak, D.; Matić-Skoko, S.; Peharda, M.; Uvanović, H.; Markulin, K.; Mertz-Kraus, R.; Grønkjær, P. Otolith Fingerprints and Tissue Stable Isotope Information Enable Allocation of Juvenile Fishes to Different Nursery Areas. Water 2021, 13, 1293. https://doi.org/10.3390/w13091293
Vrdoljak D, Matić-Skoko S, Peharda M, Uvanović H, Markulin K, Mertz-Kraus R, Grønkjær P. Otolith Fingerprints and Tissue Stable Isotope Information Enable Allocation of Juvenile Fishes to Different Nursery Areas. Water. 2021; 13(9):1293. https://doi.org/10.3390/w13091293
Chicago/Turabian StyleVrdoljak, Dario, Sanja Matić-Skoko, Melita Peharda, Hana Uvanović, Krešimir Markulin, Regina Mertz-Kraus, and Peter Grønkjær. 2021. "Otolith Fingerprints and Tissue Stable Isotope Information Enable Allocation of Juvenile Fishes to Different Nursery Areas" Water 13, no. 9: 1293. https://doi.org/10.3390/w13091293
APA StyleVrdoljak, D., Matić-Skoko, S., Peharda, M., Uvanović, H., Markulin, K., Mertz-Kraus, R., & Grønkjær, P. (2021). Otolith Fingerprints and Tissue Stable Isotope Information Enable Allocation of Juvenile Fishes to Different Nursery Areas. Water, 13(9), 1293. https://doi.org/10.3390/w13091293