
Biomat Resilience to Desiccation and Flooding Within a Shallow, Unit Process Open Water Engineered Wetland

 , ,
, ,
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
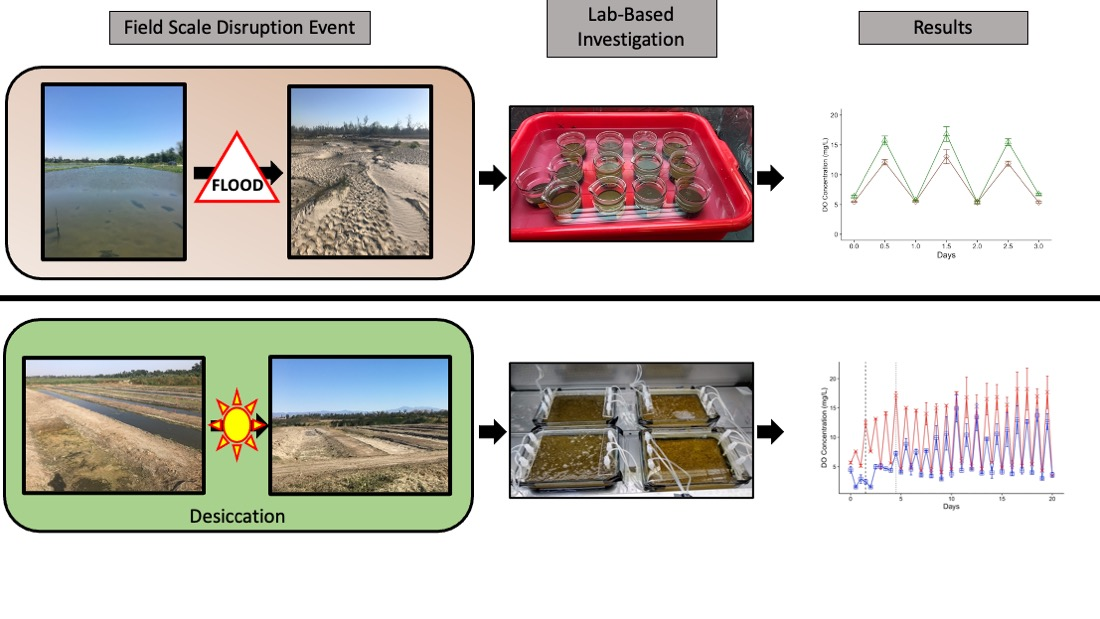
2.1. Field Systems
2.2. Flood-Impacted Batch Microcosms
2.3. Rehydration Flow-Through Microcosms
2.4. Aqueous Phase Sampling and Analysis
2.5. DNA Extraction and Sequence Analysis
2.6. Bioinformatic and Statistical Analysis
3. Results
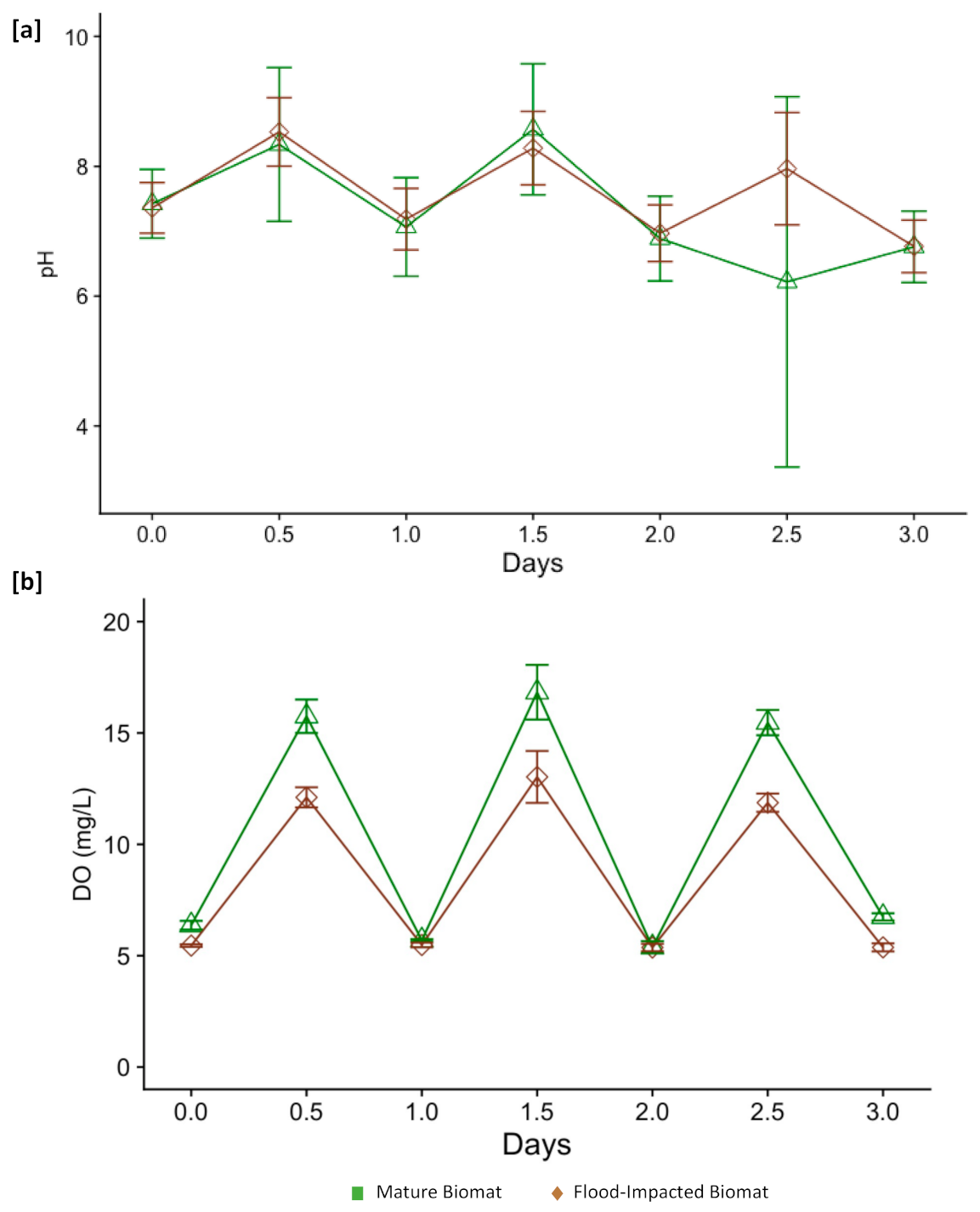
3.1. Environmental Disruption Extremes
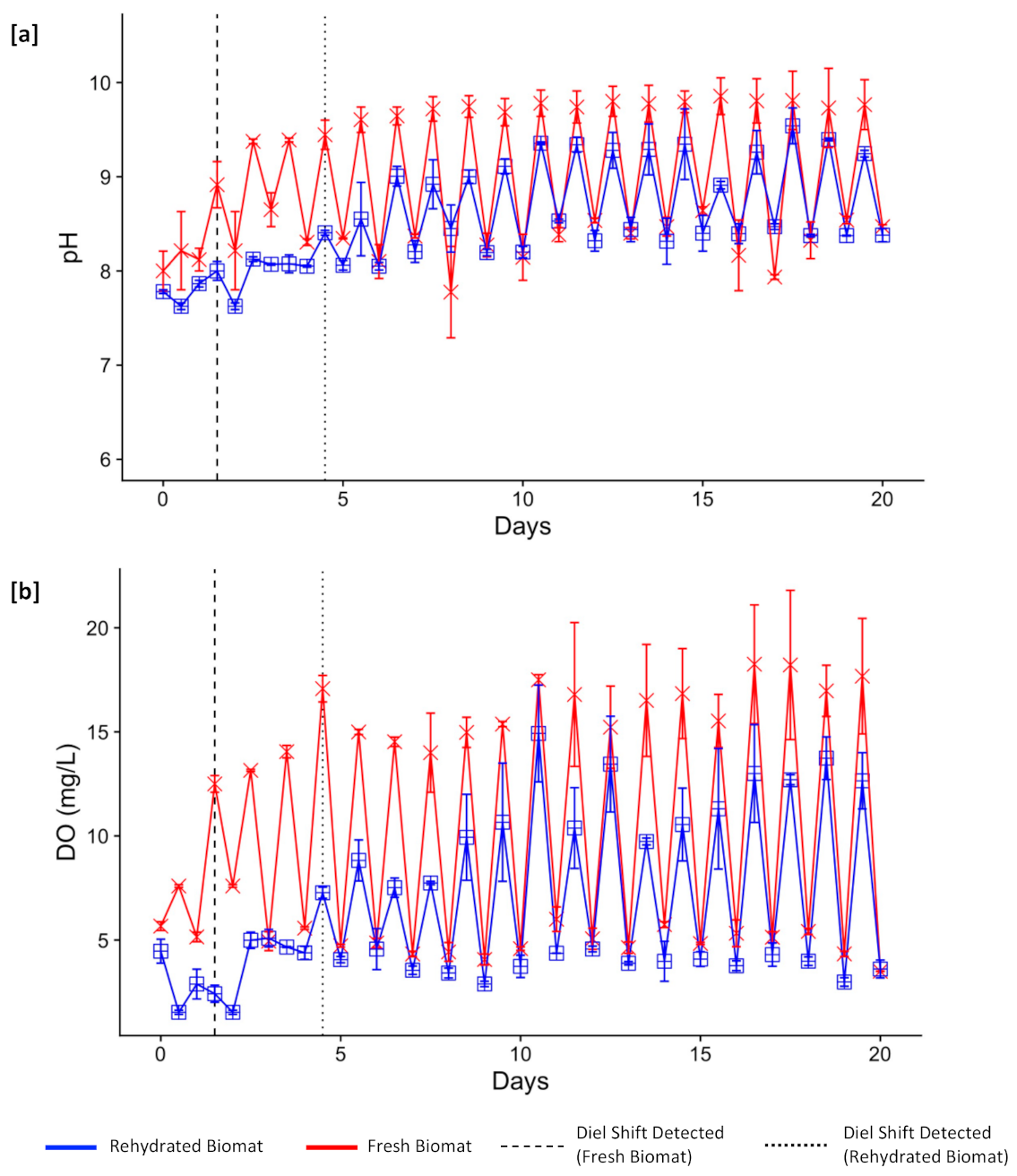
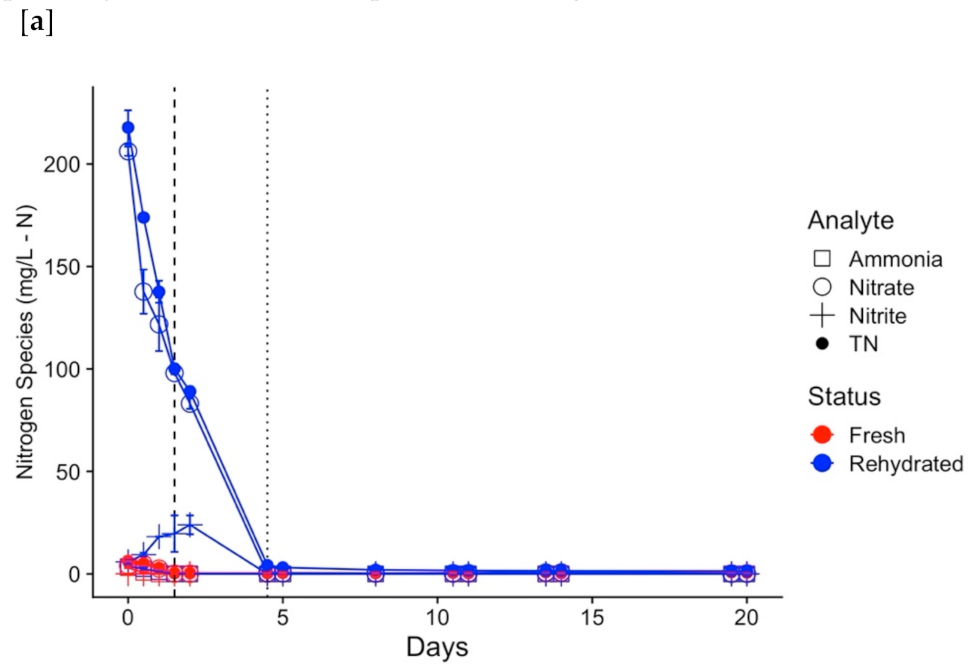
3.2. Physicochemical Changes Associated with Disruption Extremes
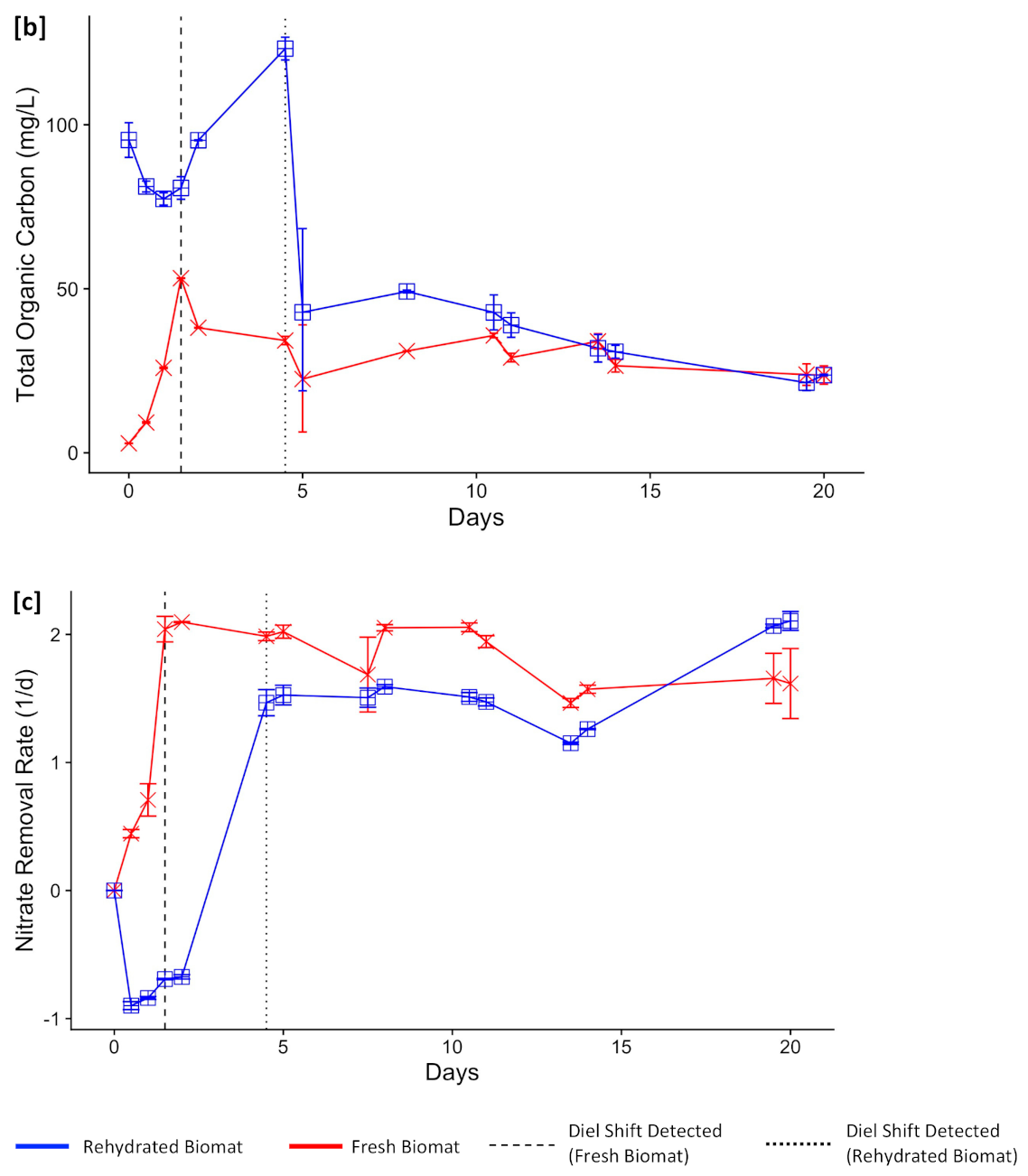
3.3. Functional Resilience
3.4. Operational Resilience
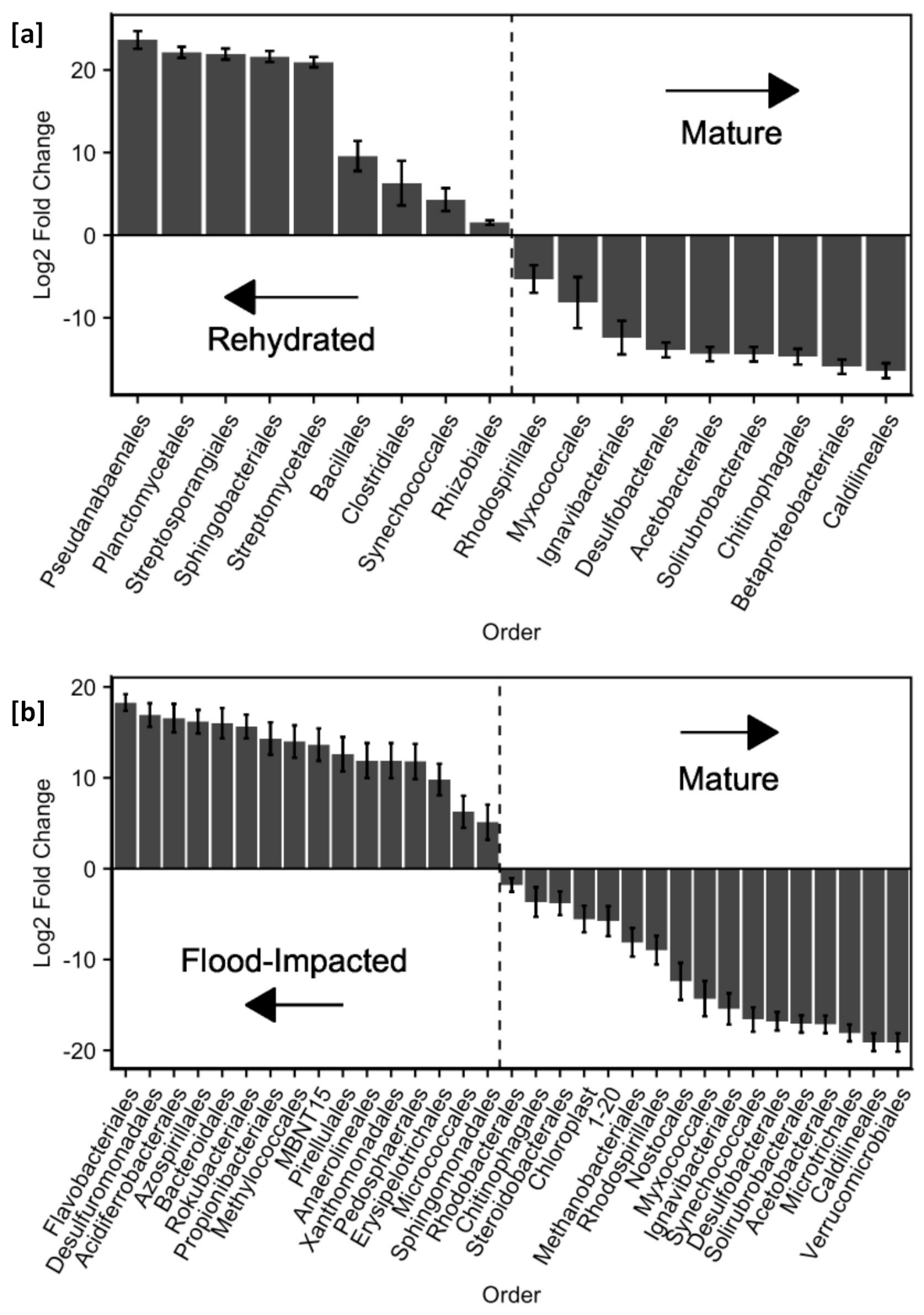
3.5. Ecological Resistance of Microbial Assemblage
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Seneviratne, S.; Nicholls, N.; Easterling, D.; Goodess, C.; Kanae, S.; Kossin, J.; Luo, Y.; Marengo, J.; McInnes, K.; Rahimi, M.; et al. Changes in climate extremes and their impacts on the natural physical environment. In Managing the Risks of Extreme Events and Disasters to Advance Climate Change Adaptation (SREX); Field, C.B., Barros, V., Stocker, T.F., Qin, D., Dokken, D.J., Ebi, K.L., Mastrandrea, M.D., Mach, K.J., Plattner, G.-K., Allen, S.K., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2012; pp. 109–230. Available online: https://www.cambridgorg/core/books/managing-the-risks-of-extreme-events-and-disasters-to-advance-climate-change-adaptation/changes-in-climate-extremes-and-their-impacts-on-the-natural-physical-environment/A4B05D458547ACA9017591A37860DDD3 (accessed on 22 February 2021).
- Milly, P.C.D.D.; Wetherald, R.T.; Dunne, K.A.; Delworth, T.L. Increasing risk of great floods in a changing climat. Nature 2002, 415, 514–517. [Google Scholar] [CrossRef]
- U.S. Bureau of Reclamatio Climate Change Analysis for the Santa Ana River Watershed—Santa Ana Watershed Basin Study 2013. Available online: https://www.usbgov/lc/socal/basinstudies/OWOWReferences/FinalReport/TM1ClimateChangpdf (accessed on 7 November 2019).
- Cook, B.; Mankin, J.S.; Anchukaitis, K.J. Climate Change and Drought: From Past to Futur. Curr. Clim. Chang. Rep. 2018, 4, 164–179. [Google Scholar] [CrossRef]
- Olsson, L.; Jerneck, A.; Thoren, H.; Persson, J.; O’Byrne, D. Why resilience is unappealing to social science: Theoretical and empirical investigations of the scientific use of resilienc. Sci. Adv. 2015, 1, e1400217. [Google Scholar] [CrossRef] [PubMed]
- Juan-Garcia, P.; Butler, D.; Comas, J.; Darch, G.; Sweetapple, C.; Thornton, A.; Corominas, L. Resilience theory incorporated into urban wastewater systems managemen State of the air. Water Res. 2017, 115, 149–161. [Google Scholar] [CrossRef] [PubMed]
- Shade, A.; Peter, H.; Allison, S.D.; Baho, D.L.; Berga, M.; Buergmann, H.; Huber, D.H.; Langenheder, S.; Lennon, J.T.; Martiny, J.B.H.; et al. Fundamentals of microbial community resistance and resilienc. Front. Microbiol. 2012, 3, 417. [Google Scholar] [CrossRef] [PubMed]
- Allison, S.D.; Martiny, J.B.H. Resistance, resilience, and redundancy in microbial communities. Proc. Natl. Acad. Sci. USA 2009, 105, 149–166. [Google Scholar] [CrossRef] [PubMed]
- Gersonius, B.; Ashley, R.; Pathirana, A.; Zevenbergen, C. Climate change uncertainty: Building flexibility into water and flood risk infrastructure. Clim. Chang. 2013, 116, 411–423. [Google Scholar] [CrossRef]
- Youn, B.D.; Hu, C.; Wang, P. Resilience-driven system design of complex engineered systems. J. Mech. Des. 2011, 133, 101011-1–101011–5. [Google Scholar] [CrossRef]
- Jackson, S.; Ferris, T. Resilience Principles for Engineered Systems. Syst. Eng. 2013, 16, 152–164. [Google Scholar] [CrossRef]
- Kiparsky, M.; Thompson, B.H.; Binz, C.; Sedlak, D.L.; Tummers, L.; Truffer, B. Barriers to Innovation in Urban Wastewater Utilities: Attitudes of Managers in California. Environ. Manag. 2016, 57, 1204–1216. [Google Scholar] [CrossRef]
- Acreman, M.; Holden, J. How wetlands affect floods. Wetlands 2013, 33, 773–786. [Google Scholar] [CrossRef]
- Brody, S.D.; Sebastian, A.; Blessing, R.; Bedient, P.B. Case study results from southeast Houston, Texas: Identifying the impacts of residential location on flood risk and loss. J. Flood Risk Manag. 2018, 11, S110–S120. [Google Scholar] [CrossRef]
- Shimabuku, M.; Diringer, S.; Cooley, H. Stormwater Capture in California: Innovative Policies and Funding Opportunities; Pacific Institute: Oakland, CA, USA, 2018; 33p, Available online: http://pacinsorg/wp-content/uploads/2018/06/Pacific-Institute-Stormwater-Capture-in-California.pdf (accessed on 4 February 2021).
- Regnery, J.; Lee, J.; Kitanidis, P.; Illangasekare, T.; Sharp, J.O.; Drewes, J.E. Integration of artificial recharge and recovery systems for impaired water sources in urban settings: Overcoming current limitations and engineering challenges. Environ. Eng. Sci. 2013, 30, 409–420. [Google Scholar] [CrossRef]
- Jasper, J.T.; Nguyen, M.T.; Jones, Z.L.; Ismail, N.S.; Sedlak, D.L.; Sharp, J.O.; Luthy, R.G.; Horne, A.J.; Nelson, K.L. Unit Process Wetlands for Removal of Trace Organic Contaminants and Pathogens from Municipal Wastewater Effluents. Environ. Eng. Sci. 2013, 30, 421–436. [Google Scholar] [CrossRef]
- Grebel, J.E.; Mohanty, S.K.; Torkelson, A.A.; Boehm, A.B.; Higgins, C.P.; Maxwell, R.M.; Nelson, K.L.; Sedlak, D.L. Engineered Infiltration Systems for Urban Stormwater Reclamatio. Environ. Eng. Sci. 2013, 30, 437–454. [Google Scholar] [CrossRef]
- Ashoori, N.; Teixido, M.; Spahr, S.; LeFevre, G.H.; Sedlak, D.L.; Luthy, R.G. Evaluation of pilot-scale biochar-amended woodchip bioreactors to remove nitrate, metals, and trace organic contaminants from urban stormwater runoff. Water Res. 2019, 154, 1–11. [Google Scholar] [CrossRef]
- Kadlec, R.H.; Wallace, S.D. Treatment Wetlands, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2009. [Google Scholar]
- Jasper, J.T.; Sedlak, D.L. Phototransformation of Wastewater-Derived Trace Organic Contaminants in Open-Water Unit Process Treatment Wetlands. Environ. Sci. Technol. 2013, 47, 10781–10790. [Google Scholar] [CrossRef]
- Bear, S.E.; Nguyen, M.T.; Jasper, J.T.; Nygren, S.; Nelson, K.L.; Sedlak, D.L. Removal of nutrients, trace organic contaminants, and bacterial indicator organisms in a demonstration-scale unit process open-water treatment wetland. Ecol. Eng. 2017, 109, 76–83. [Google Scholar] [CrossRef]
- Jasper, J.T.; Jones, Z.L.; Sharp, J.O.; Sedlak, D.L. Biotransformation of trace organic contaminants in open-water unit process treatment wetlands. Environ. Sci. Technol. 2014, 48, 5136–5144. [Google Scholar] [CrossRef] [PubMed]
- Jasper, J.T.; Jones, Z.L.; Sharp, J.O.; Sedlak, D.L. Nitrate removal in shallow, Open-water treatment wetlands. Environ. Sci. Technol. 2014, 48, 11512–11520. [Google Scholar] [CrossRef] [PubMed]
- Jones, Z.L.; Mikkelson, K.M.; Nygren, S.; Sedlak, D.L.; Sharp, J.O. Establishment and convergence of photosynthetic microbial biomats in shallow unit process open-water wetlands. Water Res. 2018, 133, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Jones, Z.L.; Jasper, J.T.; Sedlak, D.L.; Sharp, J.O. Sulfide-Induced Dissimilatory Nitrate Reduction to Ammonium Supports Anaerobic Ammonium Oxidation (Anammox) in an Open-Water Unit Process Wetland. Appl. Environ. Microbiol. 2017, 83, e00782-17. [Google Scholar] [CrossRef] [PubMed]
- Reed, A.P. Diel Redox Cycling and Its Impact on Inorganic Nitrogen in An Engineered Wetland Designed for Water Treatment. Master’s Thesis, Colorado School of Mines, Golden, Colorado, 2019. Available online: https://mountainscholaorg/handle/11124/173018 (accessed on 10 June 2019).
- Topp, G.C.; Parkin, G.W.; Ferre, T.P.A. Chapter 70: Soil Water Conten Second. In Soil Sampling and Methods of Analysis, 2nd ed.; Carter, M.R., Gregorich, E.G., Eds.; Taylor and Francis Group, LLC: Boca Raton, FL, USA, 2008; pp. 939–962. [Google Scholar]
- APHA. 4500-NO2 B. Colorimetric Method. In Standard Methods for the Examination of Water and Wastewate, 18th ed.; Greenberg, A., Clesceri, L., Eaton, A., Franson, M.A., Eds.; APHA, AWWA, WPFC: Washington, DC, USA, 1992; pp. 4–86. [Google Scholar]
- APHA. 4500-NH3 F. Phenate Method. In Standard Methods for the Examination of Water and Wastewate, 21st ed.; APHA, AWWA, WPFC: Washington, DC, USA, 1997; pp. 4108–4117. [Google Scholar]
- Cataldo, D.A.; Maroon, M.; Schrader, L.E.; Youngs, V.L. Rapid colorimetric determination of nitrate in plant tissue by nitration of salicylic acid. Commun. Soil Sci. Plant Anal. 1975, 6, 71–80. [Google Scholar] [CrossRef]
- DeBusk, T.A.; Grace, K.A.; Dierberg, F.E.; Jackson, S.D.; Chimney, M.J.; Gu, B. An investigation of the limits of phosphorus removal in wetlands: A mesocosm study of a shallow periphyton-dominated treatment system. Ecol. Eng. 2004, 23, 1–14. [Google Scholar] [CrossRef]
- Reed, A.P.; Sharp, J.O.; Tobias, C.R.; Repert, D.A.; Smith, R.L. Surface Water and Porewater Chemistry of Cell 3A; Prado Wetlands: Riverside, CA, USA, 2020; Available online: https://www.sciencebasgov/catalog/item/5cdaf823e4b0ab16db3a82f9 (accessed on 10 January 2021).
- El Haïkali, B.; Bensoussan, N.; Romano, J.C.; Bousquet, V. Estimation of photosynthesis and calcification rates of Corallina elongata Ellis and Solander, 1786, by measurements of dissolved oxygen, pH and total alkalinity. Sci. Mar. 2004, 68, 45–56. [Google Scholar] [CrossRef][Green Version]
- Revs Bech, N.P.; Ward, D.M. Microelectrode Studies of Interstitial Water Chemistry and Photosynthetic Activity in a Hot Spring Microbial Mat. Appl. Environ. Microbiol. 1984, 48, 270–275. [Google Scholar] [CrossRef]
- Neal, C.; Harrow, M.; Williams, R.J. Dissolved carbon dioxide and oxygen in the River Thames: Spring-summer 1997. Sci. Total Environ. 1998, 210/211, 205–217. [Google Scholar] [CrossRef]
- Kraus, E.A.; Beeler, S.R.; Mors, R.A.; Floyd, J.G.; Stamps, B.W.; Nunn, H.S.; Stevenson, B.S.; Johnson, H.A.; Shapiro, R.S.; Loyd, S.J.; et al. Microscale biosignatures and abiotic mineral authigenesis in Little Hot Creek, California. Front. Microbiol. 2018, 9, 1–13. [Google Scholar] [CrossRef]
- Parada, A.E.; Needham, D.M.; Fuhrman, J.A. Every base matters: Assessing small subunit rRNA primers for marine microbiomes with mock communities, time series and global field samples. Environ. Microbiol. 2016, 18, 1403–1414. [Google Scholar] [CrossRef] [PubMed]
- Stamps, B.W.; Lyles, C.N.; Suflita, J.M.; Masoner, J.R.; Cozzarelli, M.; Kolpin, D.W.; Stevenson, B.S. Municipal solid waste landfills harbor distinct microbiomes. Front. Microbiol. 2016, 7, 534. [Google Scholar] [CrossRef]
- Honeyman, A.S.; Day, M.L.; Spear, J.R. Regional fresh snowfall microbiology and chemistry are driven by geography in storm-tracked events, Colorado, USA. PeerJ 2018, 6, e5961. [Google Scholar] [CrossRef]
- Schubert, S.; Lindgreen, S.; Orlando, L. AdapterRemoval v2: Rapid adapter trimming, identification, and read merging. BMC Res. Notes 2016, 9, 88. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2—Approximately maximum-likelihood trees for large alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef] [PubMed]
- Paradis, E.; Claude, J.; Strimmer, K. APE: Analyses of phylogenetics and evolution in R language. Bioinformatics 2004, 20, 289–290. [Google Scholar] [CrossRef] [PubMed]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e62217. [Google Scholar] [CrossRef] [PubMed]
- Andersen, K.S.; Kirkegaard, R.H.; Karst, S.M.; Albertsen, M. ampvis2: An R package to analyse and visualise 16S rRNA amplicon data. bioRxiv 2018, 299537. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Wagner, H. vegan: Community Ecology Package 2019. Available online: https://crar-projecorg/package=vega (accessed on 24 May 2020).
- Love, M.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Lozupone, C.; Knight, R. UniFrac: A new phylogenetic method for comparing microbial communities. Appl. Environ. Microbiol. 2005, 71, 8228–8235. [Google Scholar] [CrossRef]
- Deshmukh, S.; Herndon, R.L.; Huang, W.B.; Markus, M.R.; Miller, C.D. Santa Ana River Watermaster Report for Water Year 1 October 2018–30 September 2019 San Bernandino, CA. 2020. Available online: https://www.ocwd.com/media/8837/2018-19-watermaster-reporpdf (accessed on 9 December 2020).
- McKinley, V.L.; Peacock, A.D.; White, D.C. Microbial community PLFA and PHB responses to ecosystem restoration in tallgrass prairie soils. Soil Biol. Biochem. 2005, 37, 1946–1958. [Google Scholar] [CrossRef]
- Inglett, K.S.; Inglett, P.W.; Reddy, K.R. Soil Microbial Community Composition in a Restored Calcareous Subtropical Wetland. Soil Sci. Soc. Am. J. 2011, 75, 1731–1740. [Google Scholar] [CrossRef]
- Garrido-Oter, R.; Nakano, R.T.; Dombrowski, N.; Ma, K.W.; McHardy, A.C.; Schulze-Lefert, P. Modular Traits of the Rhizobiales Root Microbiota and Their Evolutionary Relationship with Symbiotic Rhizobia. Cell Host Microbe 2018, 24, 155–167.e5. [Google Scholar] [CrossRef] [PubMed]
- Zumft, W.G. Cell biology and molecular basis of denitrification. Microbiol. Mol. Biol. Rev. 1997, 61, 533–616. [Google Scholar] [CrossRef]
- Dhillon, A.; Teske, A.; Dillon, J.; Stahl, D.A.; Sogin, M.L. Molecular characterization of sulfate-reducing bacteria in the Guaymas basi. Appl. Environ. Microbiol. 2003, 69, 2765–2772. [Google Scholar] [CrossRef]
- Han Congeevaram, S.; Ki, D.W.; Oh, B.T.; Park, J. Bacterial community analysis of swine manure treated with autothermal thermophilic aerobic digestion. Appl. Microbiol. Biotechnol. 2011, 89, 835–842. [Google Scholar]
- Dedysh, S.N.; Kulichevskaya, S.; Beletsky, A.V.; Ivanova, A.A.; Rijpstra, W.C.; Damsté, J.S.S.; Mardanov, A.V.; Ravin, N.V. Lacipirellula parvula ge nov., sp. nov., representing a lineage of planctomycetes widespread in low-oxygen habitats, description of the family Lacipirellulaceae fam. nov. and proposal of the orders Pirellulales ord. nov., Gemmatales ord. nov. and Isosph. Syst. Appl. Microbiol. 2020, 43, 126050. [Google Scholar] [CrossRef]
- Hazarika, S.N.; Thakur, D. Actinobacteria. In Beneficial Microbes in Agro-Ecology; Amaresan, N., Senthil Kumar, M., Annapurna Krishna Kumar, K., Sankaranarayanan, A., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 443–476. Available online: http://dx.doi.org/10.1016/B978-0-12-823414-3.00021-6 (accessed on 25 January 2021).
- Bonin, A.S.; Boone, D.R. The Order Methanobacteriales. In Martin Dworkin (Editor-in-Chief), Stanley Falkow, Eugene Rosenberg, Karl-Heinz; Schleifer, E.S., Ed.; The Prokaryotes Third; Springer: Singapore, 2006; pp. 231–243. Available online: http://www.springerlink.com/index/10.1007/0-387-30743-5 (accessed on 16 December 2020).
- Komárek, J.; Kaštovs.ký, J.; Mareš, J.; Johansen, J.R. Taxonomic classification of cyanoprokaryotes (cyanobacterial genera) 2014, using a polyphasic approach. Preslia 2014, 86, 295–335. [Google Scholar]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brady, A.R.; Vega, M.A.; Riddle, K.N.; Peel, H.F.; Lundeen, E.J.; Siegmund, J.G.; Sharp, J.O. Biomat Resilience to Desiccation and Flooding Within a Shallow, Unit Process Open Water Engineered Wetland. Water 2021, 13, 815. https://doi.org/10.3390/w13060815
Brady AR, Vega MA, Riddle KN, Peel HF, Lundeen EJ, Siegmund JG, Sharp JO. Biomat Resilience to Desiccation and Flooding Within a Shallow, Unit Process Open Water Engineered Wetland. Water. 2021; 13(6):815. https://doi.org/10.3390/w13060815
Chicago/Turabian StyleBrady, Adam R., Michael A. Vega, Kimberly N. Riddle, Henry F. Peel, Evelyn J. Lundeen, Julia G. Siegmund, and Jonathan O. Sharp. 2021. "Biomat Resilience to Desiccation and Flooding Within a Shallow, Unit Process Open Water Engineered Wetland" Water 13, no. 6: 815. https://doi.org/10.3390/w13060815
APA StyleBrady, A. R., Vega, M. A., Riddle, K. N., Peel, H. F., Lundeen, E. J., Siegmund, J. G., & Sharp, J. O. (2021). Biomat Resilience to Desiccation and Flooding Within a Shallow, Unit Process Open Water Engineered Wetland. Water, 13(6), 815. https://doi.org/10.3390/w13060815