
Water Lice and Other Macroinvertebrates in Drinking Water Pipes: Diversity, Abundance and Health Risk



Abstract
:1. Introduction
2. Methods
2.1. Laboratory Experiments
2.2. Sampling of Invertebrates
2.3. Invertebrates Analyses
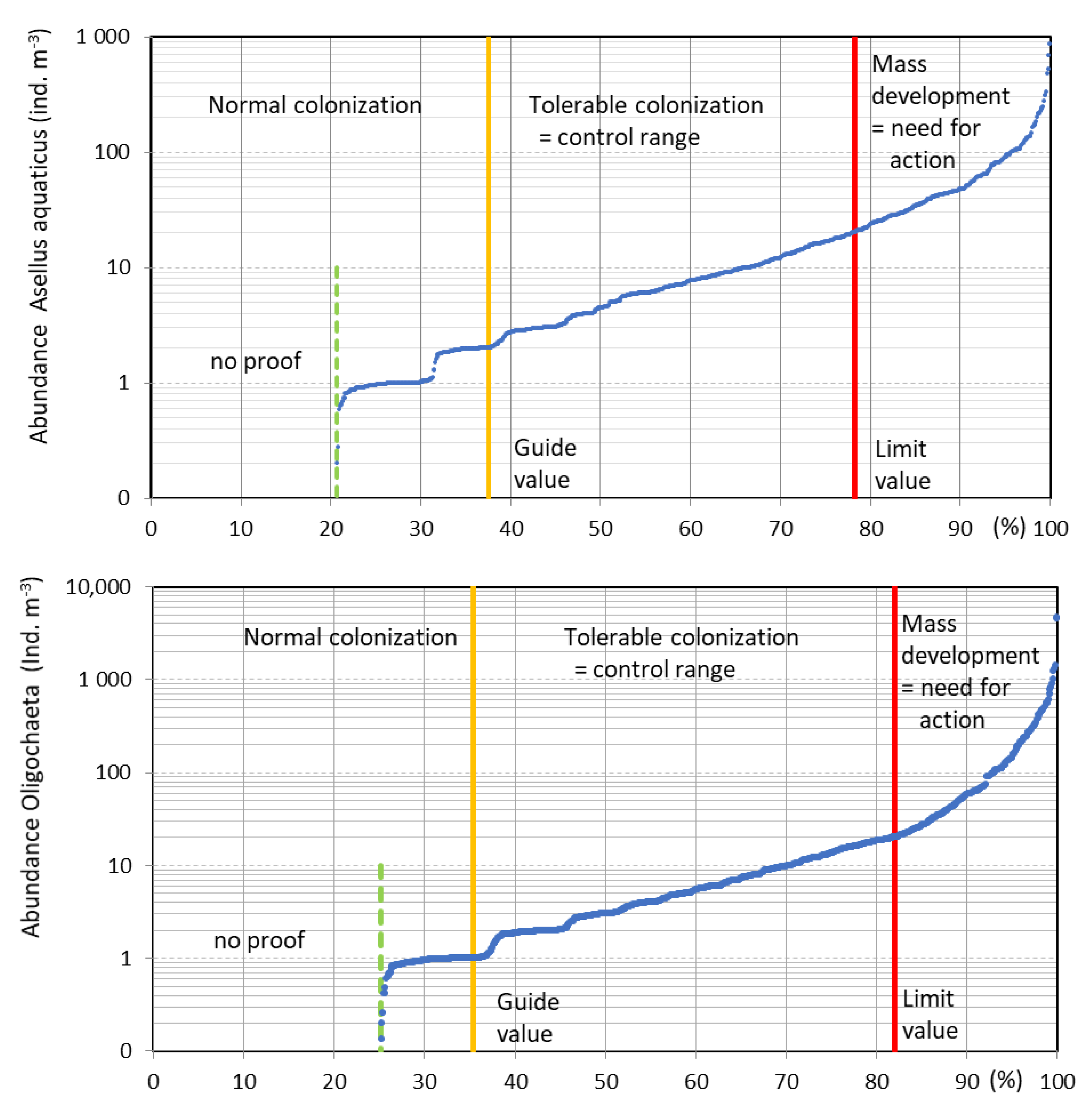
2.4. Calculation of Load Classes
3. Results
3.1. Diversity and Abundance of Macroinvertebrates
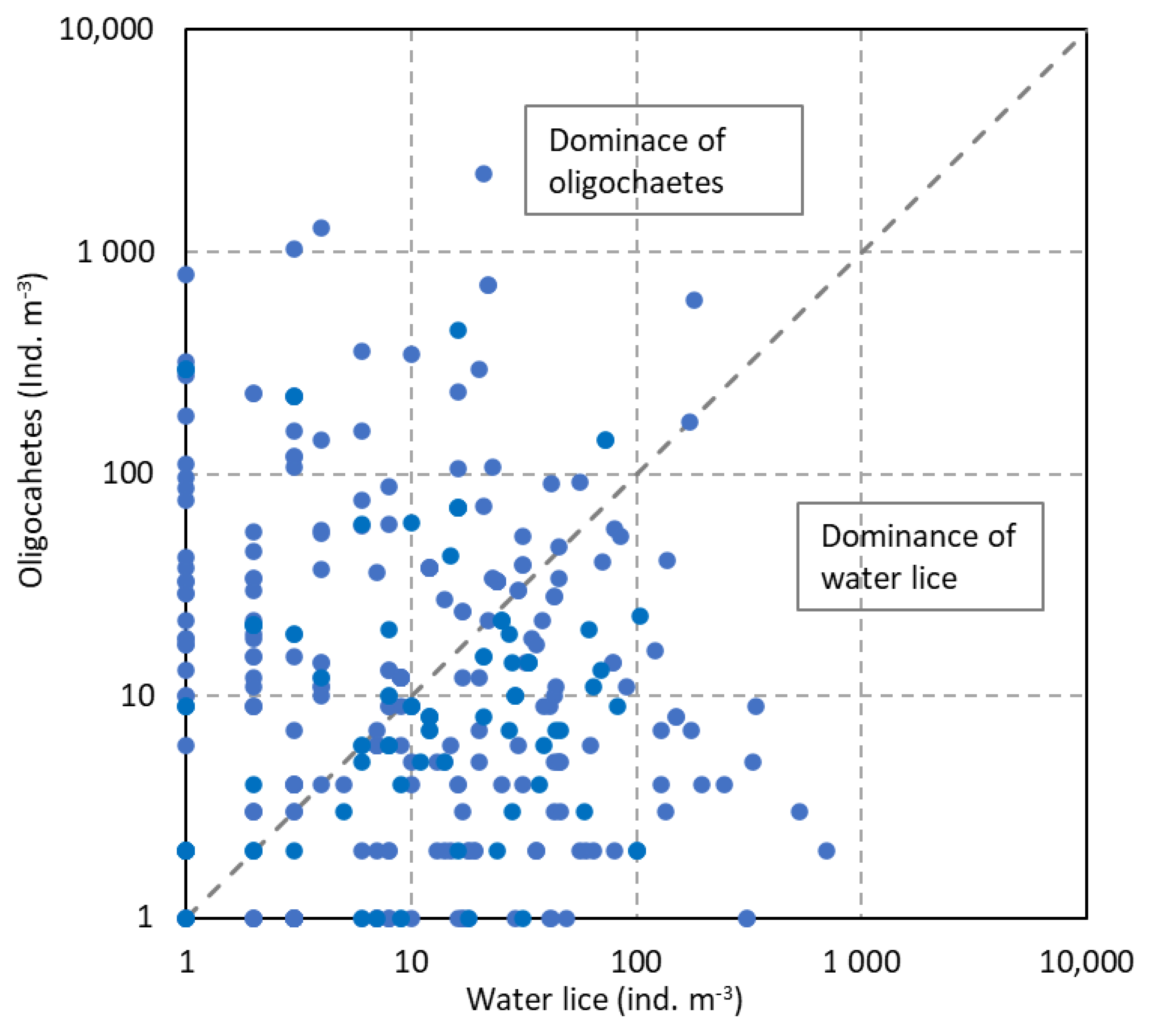
3.2. Co-Relationships of Macroinvertebrates
3.3. Load Classes of Macroinvertebrates
3.4. Reproduction of Asellus aquaticus
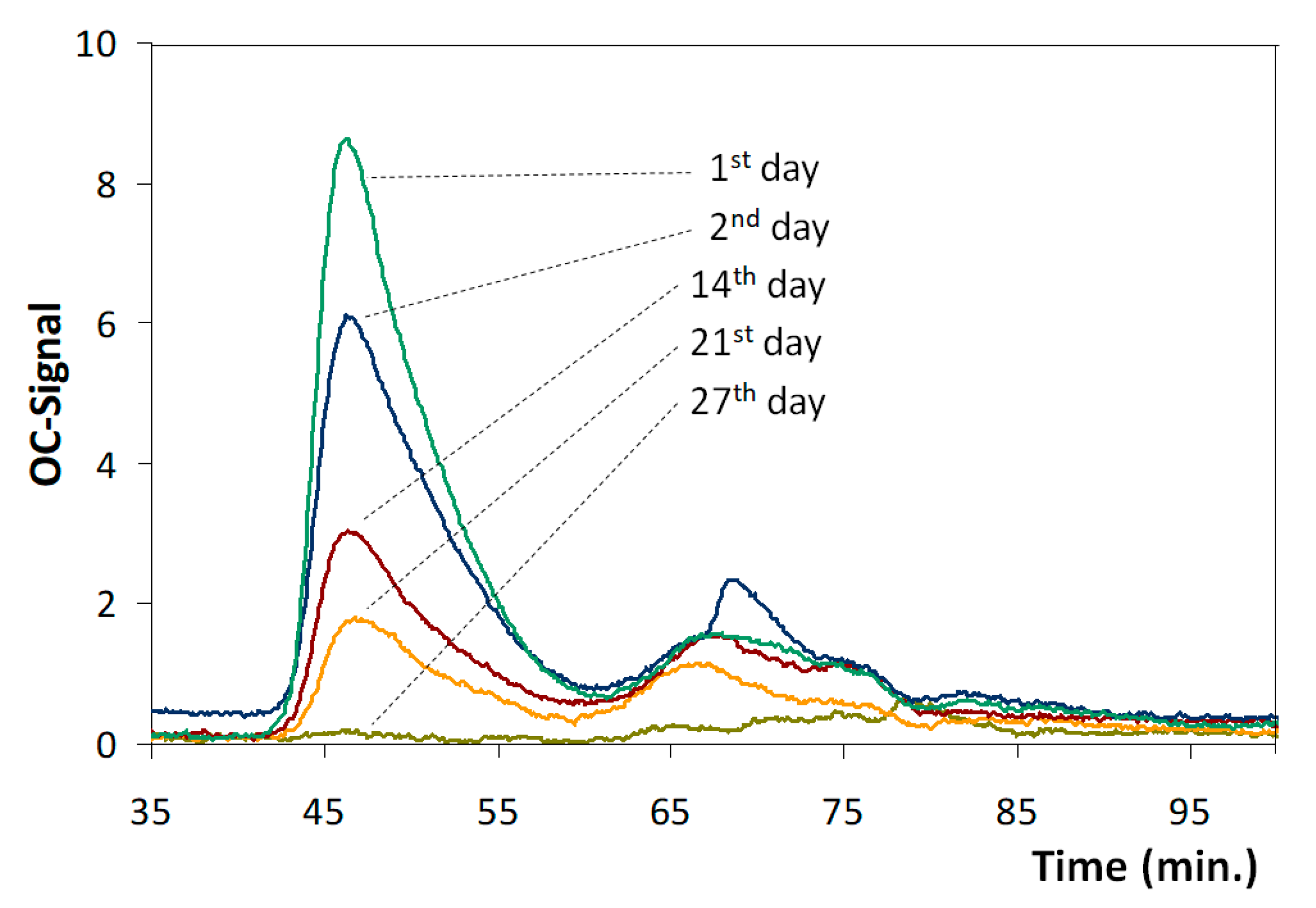
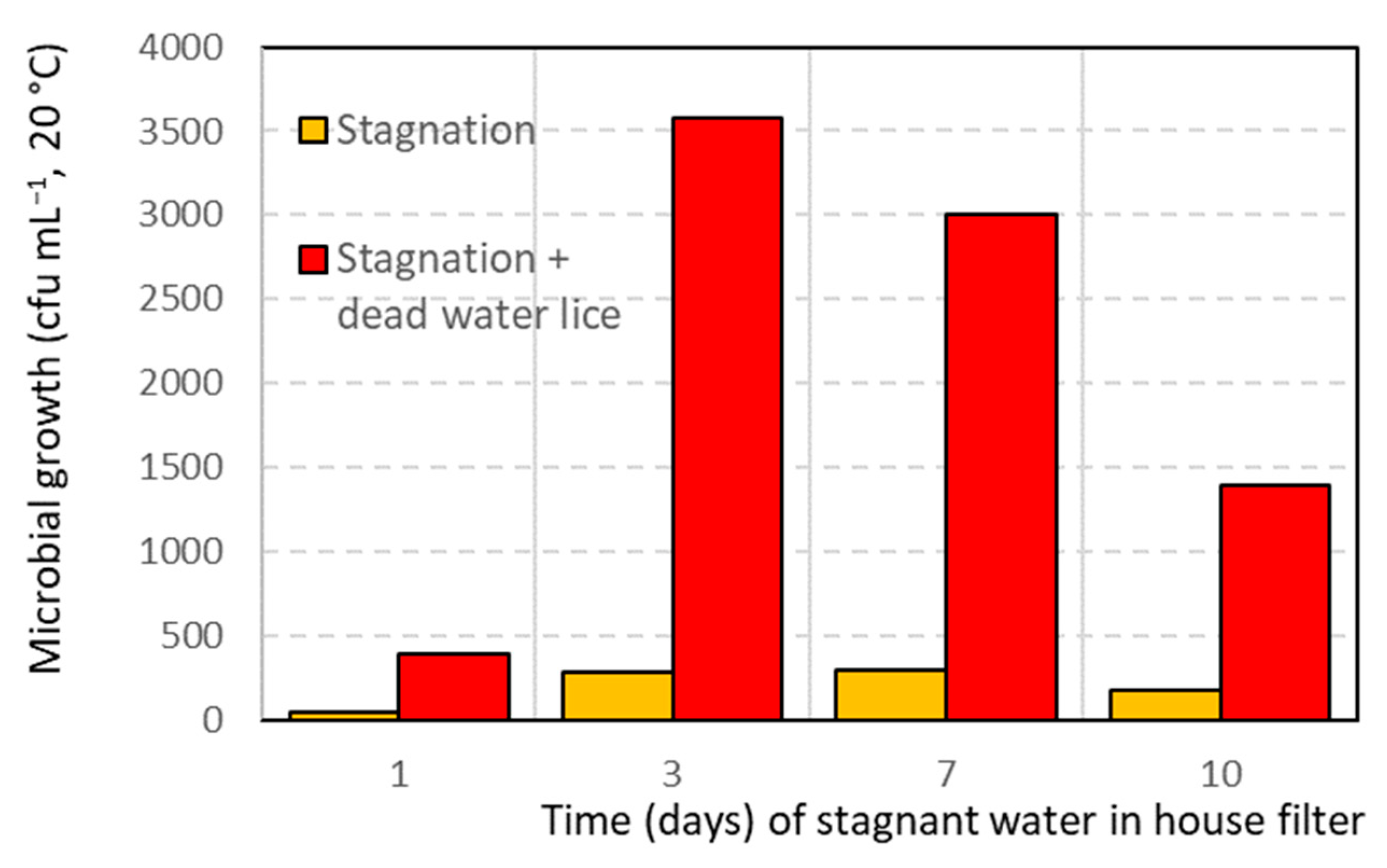
3.5. Impact of Asellus aquaticus on Drinking Water Quality
4. Discussion
4.1. Macroinvertebrates as Inhabitants of Drinking Water Pipes
4.2. Macroinvertebrates as Hosts for Harmful Bacteria
4.3. Proposed Activities for Biological Water Quality Control and Maintenance
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Gunkel, G.; Michels, U.; Scheideler, M.; Ripl, K. Vorkommen und Bedeutung von Kleintieren in Trinkwasser-Verteilungssystemen—Massnahmen zu deren Regulierung. Invertebr. Water Supply Facil. 2010, 3R, 716–724. [Google Scholar]
- Christensen, S.C.B.; Nissen, E.; Arvin, E.; Albrechtsen, H.-J. Distribution of Asellus aquaticus and microinvertebrates in a non-chlorinated drinking water supply system—Effects of pipe material and sedimentation. Water Res. 2011, 45, 3215–3224. [Google Scholar] [CrossRef] [Green Version]
- van Lieverloo, J.H.M.; Hoogenboezem, W.; Veenendaal, G.; van der Kooij, D. Variability of invertebrate abundance in drinking water distribution systems in the Netherlands in relation to biostability and sediment volumes. Wat. Res. 2012, 46, 4918–4932. [Google Scholar] [CrossRef] [PubMed]
- Hijnen, W.A.M.; Schurer, R.; Bahlman, J.A.; Ketelaars, H.A.M.; Italiaander, R.; van der Wal, A.; van der Wielen, P.W.J.J. Slowly biodegradable organic compounds impact the biostability of non-chlorinated drinking water produced from surface water. Water Res. 2018, 129, 240–251. [Google Scholar] [CrossRef] [PubMed]
- van Bel, N.; van der Wielen, P.; Wulloings, B.; van Rijn, J.; van der Mark, E.; Ketelaars, H.A.M.; Hijnen, W. Aeromonas species from non-chlorinated distribution systems and their competitive planktonic growth in drinking water. Appl. Environ. Microbiol. 2020. [Google Scholar] [CrossRef] [PubMed]
- WHO. Safe Piped Water: Managing Microbial Water Quality in Piped Distribution Systems; Ainsworth, R., Ed.; World Health Organization, IWA Publishing: London, UK, 2004; 147p, ISBN 1-84339-039-6. [Google Scholar]
- Prest, E.I.; Hammes, F.; van Loosdrecht, M.C.M.; Vrouwenvelder, J.S. Biological Stability of Drinking Water: Controlling Factors, Methods, and Challenges. Front. Microbiol. 2016, 7, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Michels, U.; Gunkel, G.; Scheideler, M. Im Fokus der Wasserversorger—Die biologische Trinkwasserqualität. Vom Wasser 2014, 112(3), 93–95. [Google Scholar]
- WHO. Guidelines for Drinking-Water Quality Incorporating First Addendum, 3rd ed.; World Health Organization: Geneva, Sitzerland, 2006; 515p. [Google Scholar]
- WHO. Guidelines for Drinking-Water Quality, 4th ed.; World Health Organisation: Geneva, Switzerland, 2011; p. 564. [Google Scholar]
- Wingender, J.; Flemming, H.-C. Biofilms in drinking water and their role as reservoir for pathogens. Int. J. Hyg. Environ. Health 2011, 214, 417–423. [Google Scholar] [CrossRef]
- Michels, U. Invertebraten in Trinkwasserverteilungssystemen. Lebensraum, Verbreitung, Nahrungsbeziehungen. Ph.D. Thesis, Technische Universität Berlin, Berlin, Germany, 2018. [Google Scholar]
- Szewzyk, U.; Szewzyk, R.; Manz, W.; Schleifer, K.-H. Microbiological safety of drinking water. Annu. Rev. Microbiol. 2000, 54, 81–127. [Google Scholar] [CrossRef]
- Ripl, K.; Slavik, I.; Edel, T.; Oltmann, J.; Uhl, W. Modellbasierte Spülplanoptimierung für Trinkwasserverteilungsnetze am Beispiel des Netzes eines niedersächsischen Wasserverbandes. gwf Wasser Abwasser 2014, 11, 1194–1203. [Google Scholar]
- Korth, A.; Donath, O. Spülverfahren für Trinkwasserleitungen problemspezifisch auswählen. Energie | Wasser-Praxis 2015, 1, 36–41. [Google Scholar]
- Brielmann, H.; Lueders, T.; Schreglmann, K.; Ferraro, F.; Avramov, M.; Hammerl, V.; Blum, P.; Bayer, P.; Griebler, C. Oberflächennahe Geothermie und ihre potenziellen Auswirkungen auf Grundwasserökosysteme. Grundwasser 2011, 16, 77–91. [Google Scholar] [CrossRef]
- Berkhoff, S.E.; Bork, J.; Hahn, H.J. Grundwasserfauna als Indikator für Oberflächenwasser-Grundwasser-Interaktionen im Bereich einer Uferfiltrationsanlage. Grundwasser 2009, 14, 3–10. [Google Scholar] [CrossRef]
- Ludwig, U.; Grischek, T.; Nestler, W.; Neumann, V. Behavior of Different Molecular-weight Fractions of DOC of Elbe River Water during River Bank Infiltration. Acta Hydrochim. Hydrobiol. 2006, 25, 145–150. [Google Scholar] [CrossRef]
- Dole-Olivier, M.-J.; Galassi, D.M.P.; Marmonier, P.; Creuzé des Châtelliers, M. The biology and ecology of lotic microcrustaceans. Freshw. Biol. 2001, 44, 63–91. [Google Scholar] [CrossRef]
- DWGV. Invertebraten in Wasserversorgungsanlagen: Vorkommen und Empfehlungen zum Umgang; Technische Regel—Arbeitsblatt W 271 (A); Deutscher Verein des Gas- und Wasserfaches (DVGW): Bonn, Germany, April 2018. [Google Scholar]
- AWWA. Problem Organisms in Water: Identification and Treatment, 3rd ed.; American Water Works Association: Denver, CO, USA, 2011; p. 145. [Google Scholar]
- Olsen, A.; Leadbeater, B.S.C.; Callow, M.E.; Holden, J.B.; Bale, J.S. The origin and population dynamics of annually re-occurring Paratanytarus grimmii (Diptera: Chironomidae) colonising granular activated carbon (GAC) adsorbers used in potable water treatment. Bull. Entomol. Res. 2009, 99, 643–651. [Google Scholar] [CrossRef] [Green Version]
- de Vries, H. Die Pflanzen und Thiere in den dunklen Räumen der Rotterdamer Wasserleitung; Staats- und Universitätsbibliothek Hamburg Carl von Ossietzky: Hamburg, Germany, 1887. [Google Scholar]
- Laviad, S.; Halpern, M. Chironomids’ Relationship with Aeromonas Species. Front. Microbiol. 2016, 7, 736. [Google Scholar] [CrossRef]
- Locas, A.; Barbeau, B.; Gauthier, V. Nematodes as a source of total coliforms in a distribution system. Can. J. Microbiol. 2007, 53, 580–585. [Google Scholar] [CrossRef]
- Bouyer, S.; Imbert, C.; Rodier, M.-H.; Héchard, Y. Long-term survival of Legionella pneumophila associated with Acanthamoeba castellanii vesicles. Environm. Micorbiol. 2007, 9, 1341–1344. [Google Scholar] [CrossRef]
- Wolmarans, E.; du Preez, H.H.; de Wet, C.M.E.; Venter, S.N. Significance of bacteria associated with invertebrates in drinking water distribution networks. Water Sci. Technol. 2005, 52, 171–175. [Google Scholar] [CrossRef]
- Bichai, F.; Payment, P.; Barbeau, B. Protection of waterborne pathogens by higher organisms in drinking water: A review. Can. J. Microbiol. 2008, 54, 509–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saxena, G.; Bharagava, R.N.; Kaithwas, G.; Raj, A. Microbial indicators, pathogens and methods for their monitoring in water environment. J. Water Health 2015, 13, 319–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gunkel, G.; Michels, U.; Scheideler, M. Erfahrungen und Effizienz der Rohrnetzpflege bei Anwendung des CO2-Spülverfahren—Vorkommen der Wasserasseln in Trinkwassernetzen. gfw Wasser Abwasser 2018, 159, 77–84. [Google Scholar]
- Gunkel, G.; Michels, U.; Scheideler, M. Invertebrates in drinking water distribution systems—Filter System for Hydrant Sampling and CO2-flushing for elimination of water bugs. gwf Wasser Abwasser 2015, 156, 52–56. [Google Scholar]
- Huber, S.A.; Frimmel, F.H. Gelchromatographie mit Kohlenstoffdetektion (LC/OCD): Ein rasches und aussagekräftiges Verfahren zur Charakterisierung hydrophiler organischer Wasserinhaltsstoffe. Vom Wasser 1996, 86, 277–290. [Google Scholar]
- Nakano, D.; Strayer, D.L. Biofouling animals in fresh water: Biology, impacts, and ecosystem engineering. Front Ecol Environ. 2014, 12, 167–175. [Google Scholar] [CrossRef]
- Økland, K.A. Life history and growth of Asellus aquaticus (L.) in relation to the environment in a eutrophic lake in Norway. Hydrobiologia 1978, 59, 243–259. [Google Scholar] [CrossRef]
- Marcus, J.H.; Sutcliffe, D.W.; Willoughby, L.G. Feeding and growth of Asellus aquaticus (Isopoda) on food items from the littoral of Windermere, including green leaves of Elodea canadensis. Freshw. Biol. 1978, 8, 505–519. [Google Scholar] [CrossRef]
- Evins, C. Small animals in drinking water distribution systems. In Safe Piped Water: Managing Microbial Water Quality in Piped Distribution Systems; Ainsworth, R., Ed.; World Health Organization, IWA Publishing: London, UK, 2004; pp. 101–120. [Google Scholar]
- Michels, U. Die Wasserassel (Asellus aquaticus) in Wasserversorgungsanlagen. Available online: https://www.yumpu.com/de/document/read/9828848/die-wasserassel-asellus-aquaticus-in-wasserversorgungs-anlagen (accessed on 12 January 2021).
- Gunkel, G. Trinkwasserqualität und Klimawandel—eine Herausforderung für die Wasserversorger—Was sind die Folgen? unpublished.
- Chambers, M.R. A comparison of the population ecology of Asellus aquaticus (L.) and Asellus meridianus rac. In the reed beds of the Tjeukemeer. Hydrobiologia 1977, 53, 147–154. [Google Scholar] [CrossRef]
- Servais, P.; Billen, G.; Laurent, P.; Levi, Y.; Randon, G. Studies of BDOC and bacterial dynamics in the drinking water distribution system of the Northern Parisian suburbs. Rev. Des Sci. De L’Eau 1992, 5, 69–89. [Google Scholar] [CrossRef] [Green Version]
- Arakelova, K.S. The evaluation of individual production and scope for growth in aquatic sow bugs (Asellus aquaticus). Aquat. Ecol. 2001, 35, 31–42. [Google Scholar] [CrossRef]
- Christensen, S.C.B.; Arvin, E.; Nissen, E.; Albrechtsen, H.-J. Asellus aquaticus as a potential carrier of Escherichia coli and other coliform bacteria into drinking water distribution systems. Int. J. Environ. Res. Public Health 2013, 10, 845–855. [Google Scholar] [CrossRef] [PubMed]
- King, C.H.; Shotts, E.B., Jr.; Wooley, R.E.; Porter, K.G. Survival of coliforms and bacterial pathogens within protozoa during chlorination. Appl. Environ. Microbiol. 1988, 54, 3023–3033. [Google Scholar] [PubMed]
- Bichai, F.; Barbeau, B.; Payment, P. Protection against UV disinfection of E. coli bacteria and B. subtilis spores ingested by C. elegans nematodes. Wat. Res. 2009, 43, 3397–3406. [Google Scholar] [CrossRef]
- Graça, M.A.S.; Maltby, L.; Calow, P. Comparative ecology of Gammarus pulex (L.) and Asellus aquaticus (L.) II: Fungal preferences. Hydrobiologia 1994, 281, 163–170. [Google Scholar] [CrossRef]
- Mayer, M. Zur Ernährungsweise von Isopoden in Trinkwasserverteilungssystemen. Ph.D. Thesis, Technical University of Berlin, Berlin, Germany, September 2013. [Google Scholar]
- Halpern, M.; Senderovich, Y. Chironomid microbiome. Microb. Ecol. 2015, 70, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Figueras, M.J.; Beaz-Hidalgo, R.; Senderovich, Y.; Laviad, S.; Halpern, M. Re-identification of Aeromonas isolates from chironomid egg masses as the potential pathogenic bacteria Aeromonas aquariorum. Environ Microbiol Rep. 2011, 3, 239–244. [Google Scholar] [CrossRef]
- Senderovich, Y.; Gershtein, Y.; Halewa, E.; Halpern, M. Vibrio cholerae and Aeromonas: Do they share a mutual host? ISME J. 2008, 2, 276–283. [Google Scholar] [CrossRef]
- Broza, M.; Gancz, H.; Halpern, M.; Kashi, Y. Adult non-biting midges: Possible windborne carriers of Vibrio cholerae non-O1 non-O139. Environ. Microbiol. 2005, 7, 576–585. [Google Scholar] [CrossRef]
- Lehtola, M.J.; Nissinen, T.K.; Miettinen, I.T.; Martikainen, P.J.; Vartianinen, T. Removal of soft deposits from distribution system improves the drinking water quality. Water Res. 2004, 38, 601–610. [Google Scholar]
- Liu, G.; Zhang, Y.; Knibbe, W.-J.; Feng, C.; Liu, W.; Medema, G.; van der Meer, W. Potential impacts of changing supply-water quality on drinking water distribution: A Review. Water Res. 2017, 116, 135–148. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Animal Groups and Species | Size Range (Length) (mm) | Occurrence Probability (%) | n (Positive Samples) | Median (Ind. m−3) | Percentile (10%) (Ind. m−3) | Percentile (90%) (Ind. m−3) | Maximum (Ind. m−3) |
|---|---|---|---|---|---|---|---|
| Macroinvertebrates (total) | >2 | 96.5 | 1003 | 15.9 | 2.0 | 135 | 4764 |
| Isopods Water louse (Asellus aquaticus) | 0.5–11 | 79.3 | 824 | 15.6 | 1.0 | 61 | 869 |
| Cave water louse (Proasellus cavaticus) ** | 1–6 | sporadic | 34 | 8.0 | 1.0 | 34 | 89 |
| Amphipoda Freshwater amphipod (Niphargus aquilex) | 0.4–6.5 | 2.4 | 20 | 1.9 | 0.9 | 14.6 | 40 |
| Midges Simuliide, adults | 1–4 | sporadic | |||||
| Chironomide, larvae Paratanytarsus grimmii | 5 | rare | 66 * | 27 | 1.9 | 154 | 1834 |
| Chironomide, adults, larvae (Limnophyes asquamatus) | 2 | sporadic | |||||
| Oligochaete earthworms (Oligochaeta) | 0.5–40 | 74.9 | 778 | 6.0 | 1.0 | 92.3 | 4723 |
| Springtails (Collembola) | 1–5 | sporadic | |||||
| Snails Nautilus ramshorn (Gyraulus crista) | 1–2 | rare | 12 * | 6 | 1599 | ||
| Bladder snail (Physella acuta) | 1.5–5 | rare | |||||
| Bryozoa (Plumatella spec.) | 20 | rare |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gunkel, G.; Michels, U.; Scheideler, M. Water Lice and Other Macroinvertebrates in Drinking Water Pipes: Diversity, Abundance and Health Risk. Water 2021, 13, 276. https://doi.org/10.3390/w13030276
Gunkel G, Michels U, Scheideler M. Water Lice and Other Macroinvertebrates in Drinking Water Pipes: Diversity, Abundance and Health Risk. Water. 2021; 13(3):276. https://doi.org/10.3390/w13030276
Chicago/Turabian StyleGunkel, Günter, Ute Michels, and Michael Scheideler. 2021. "Water Lice and Other Macroinvertebrates in Drinking Water Pipes: Diversity, Abundance and Health Risk" Water 13, no. 3: 276. https://doi.org/10.3390/w13030276
APA StyleGunkel, G., Michels, U., & Scheideler, M. (2021). Water Lice and Other Macroinvertebrates in Drinking Water Pipes: Diversity, Abundance and Health Risk. Water, 13(3), 276. https://doi.org/10.3390/w13030276