
The Effects of Different Carbon Sources on the Production Environment and Breeding Parameters of Litopenaeus vannamei



Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Shrimp and Acclimation
2.2. Experimental Design
2.3. Water Quality
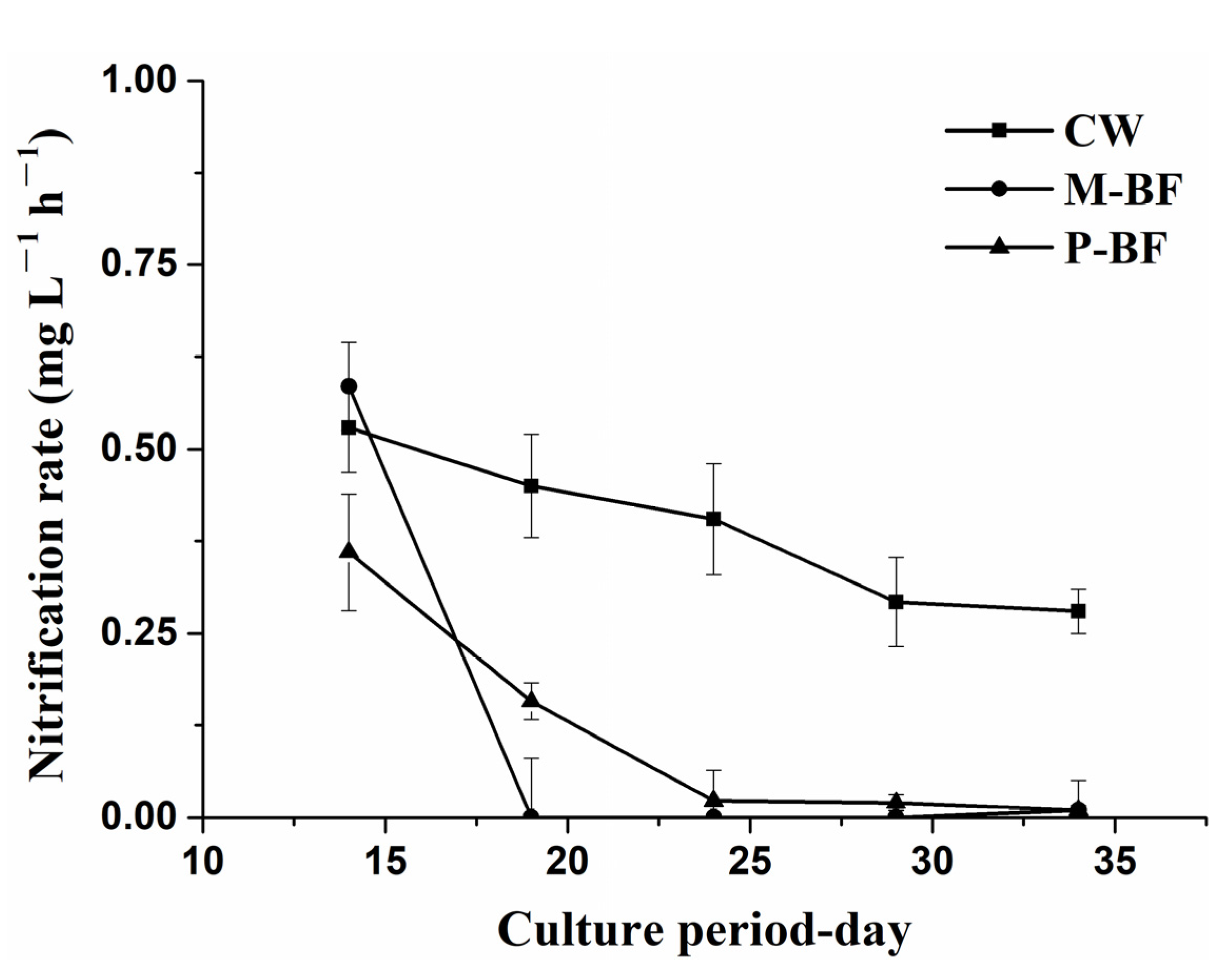
2.4. Nitrification Rate
2.5. 16S rRNA Gene High Throughput Sequencing Analysis
2.6. Shrimp Growth Performance and Survival
2.7. Statistical Analysis
3. Results
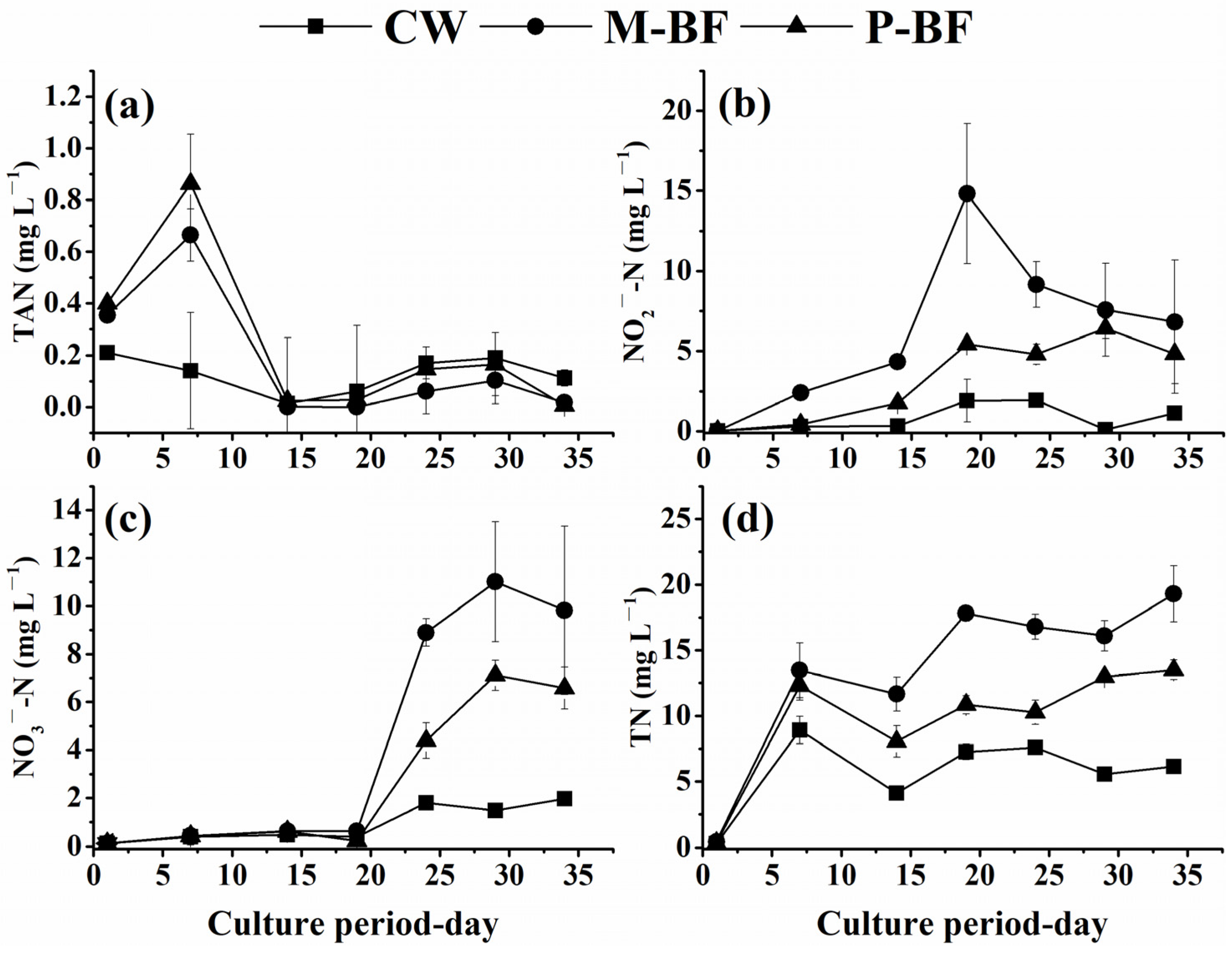
3.1. Water Quality
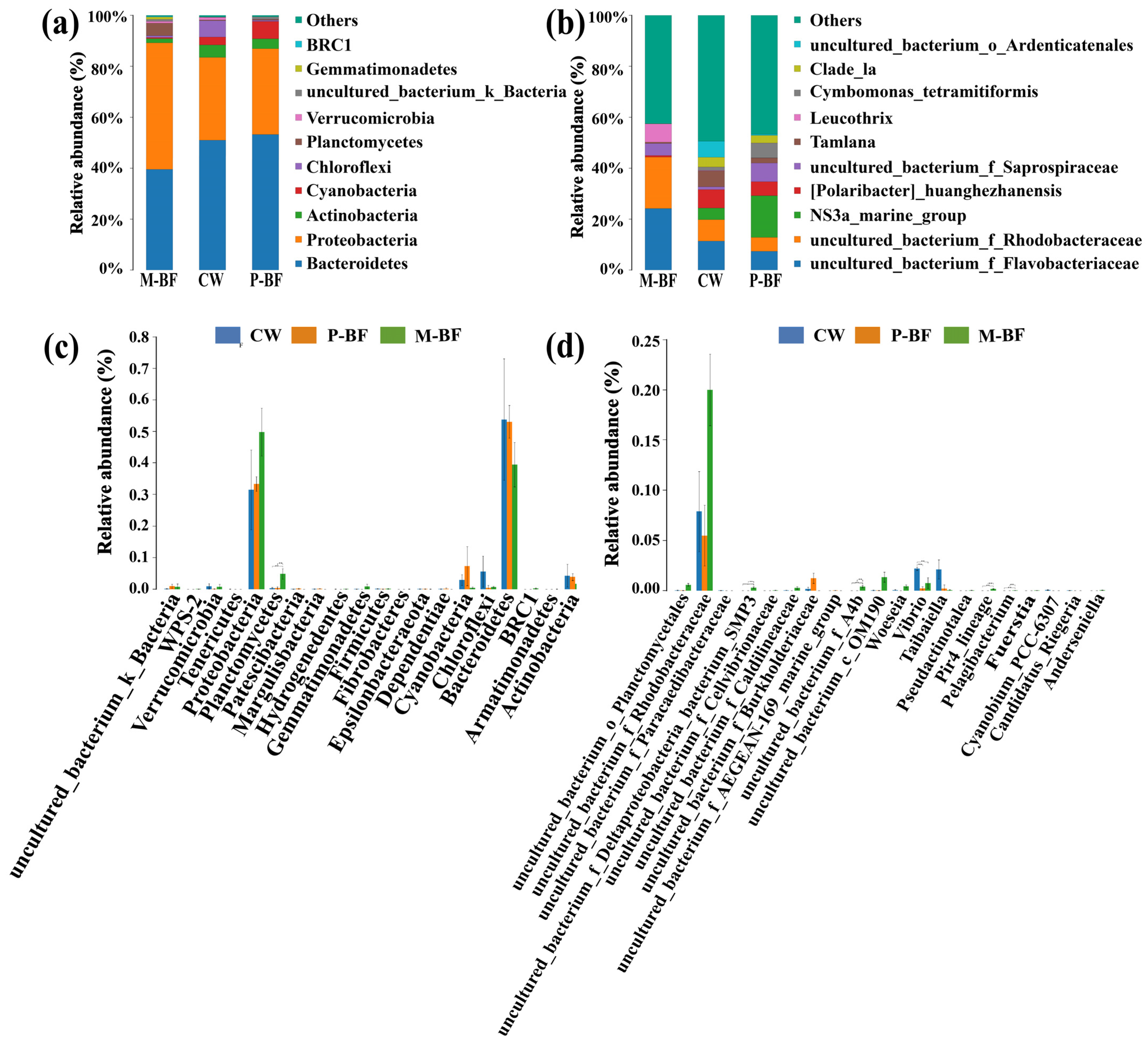
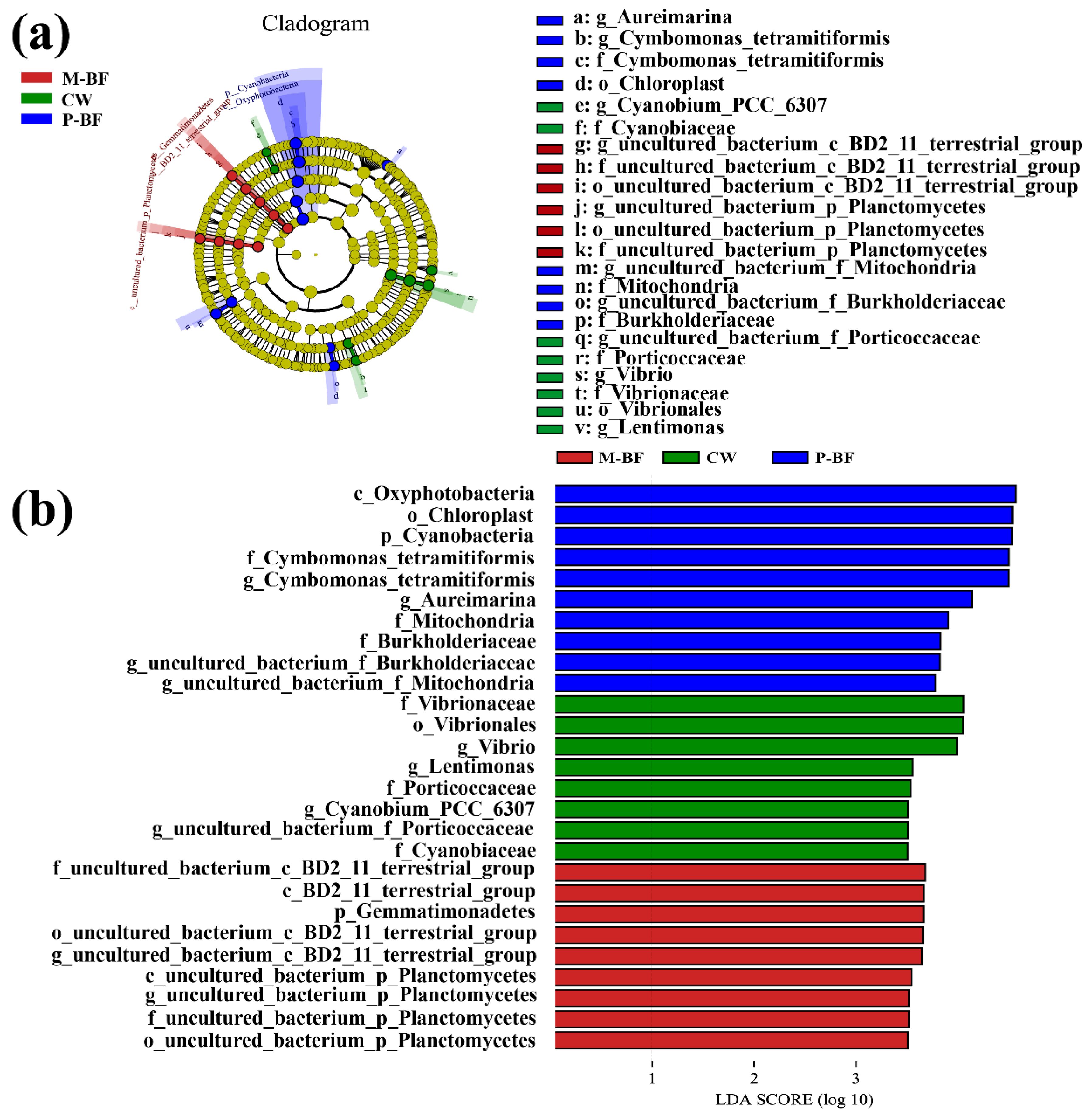
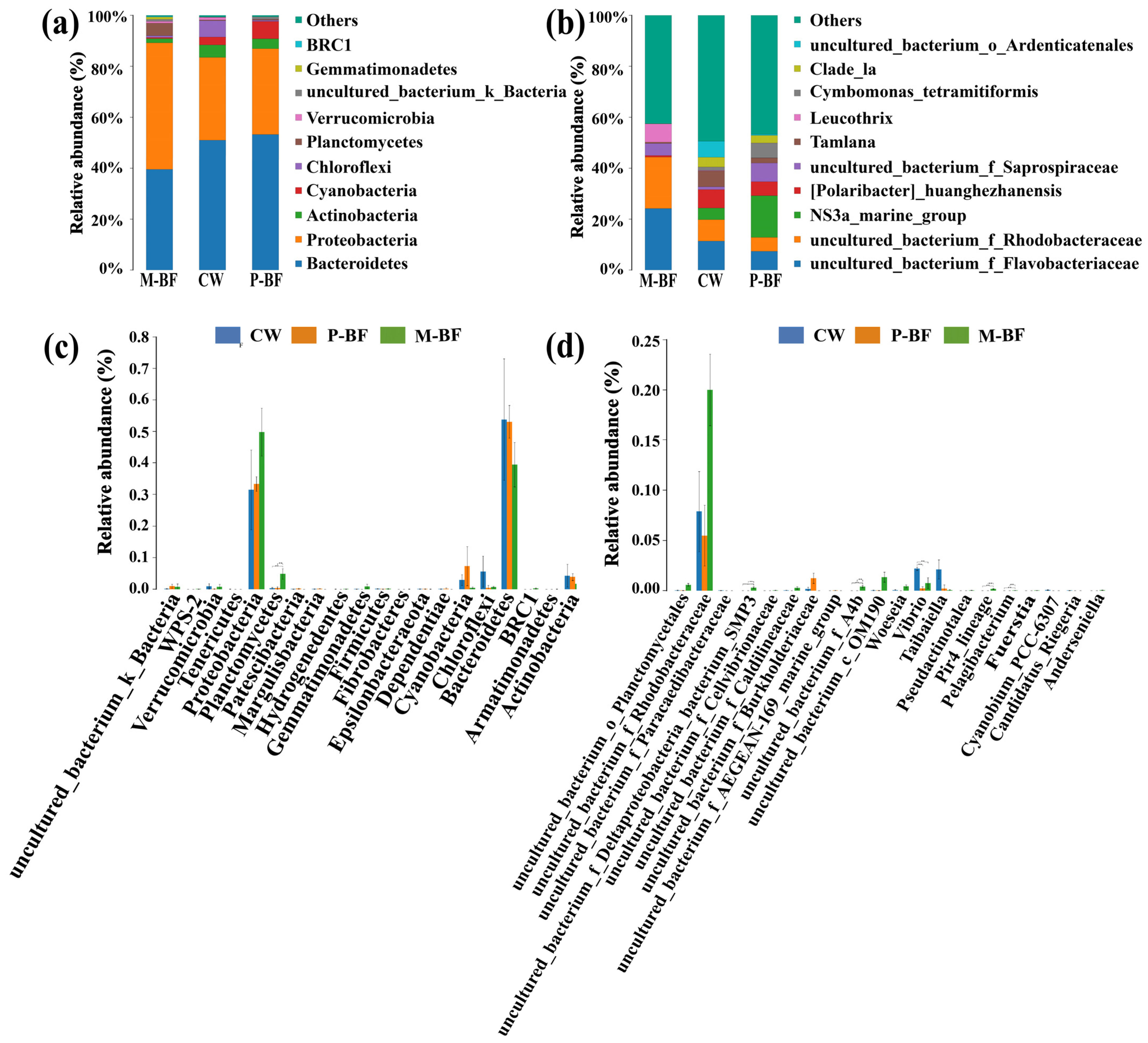
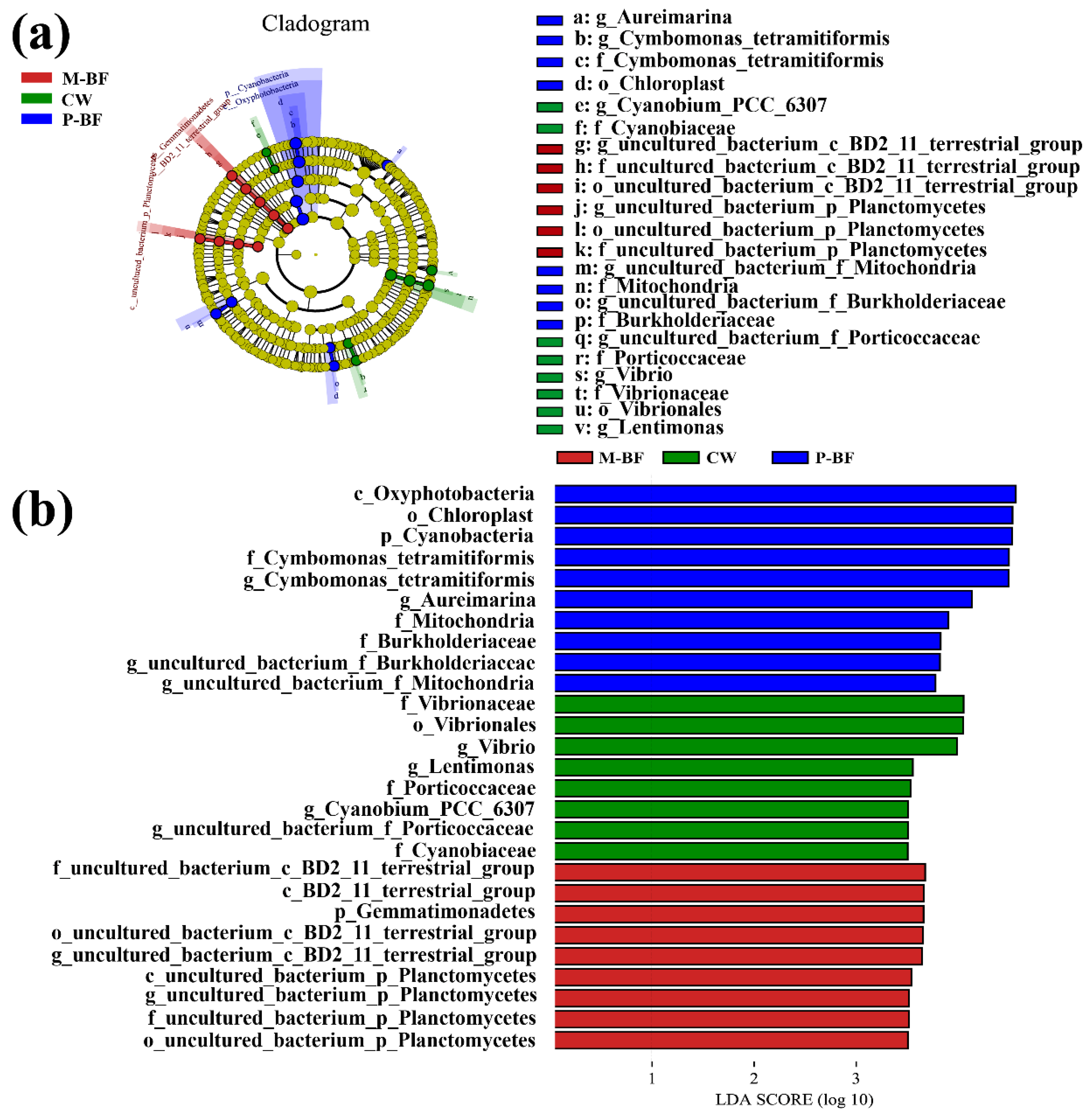
3.2. Characterization of the Bacterial Community
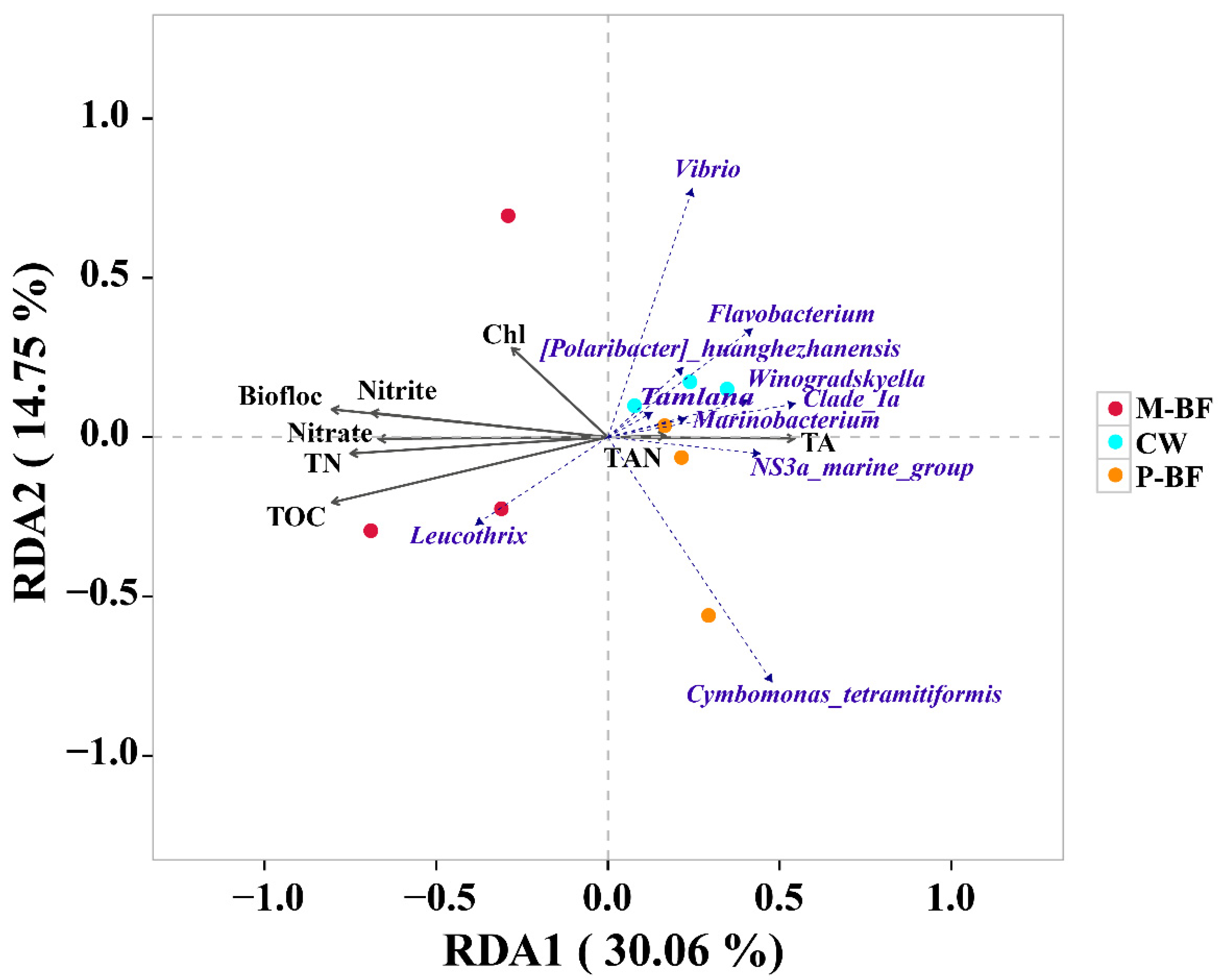
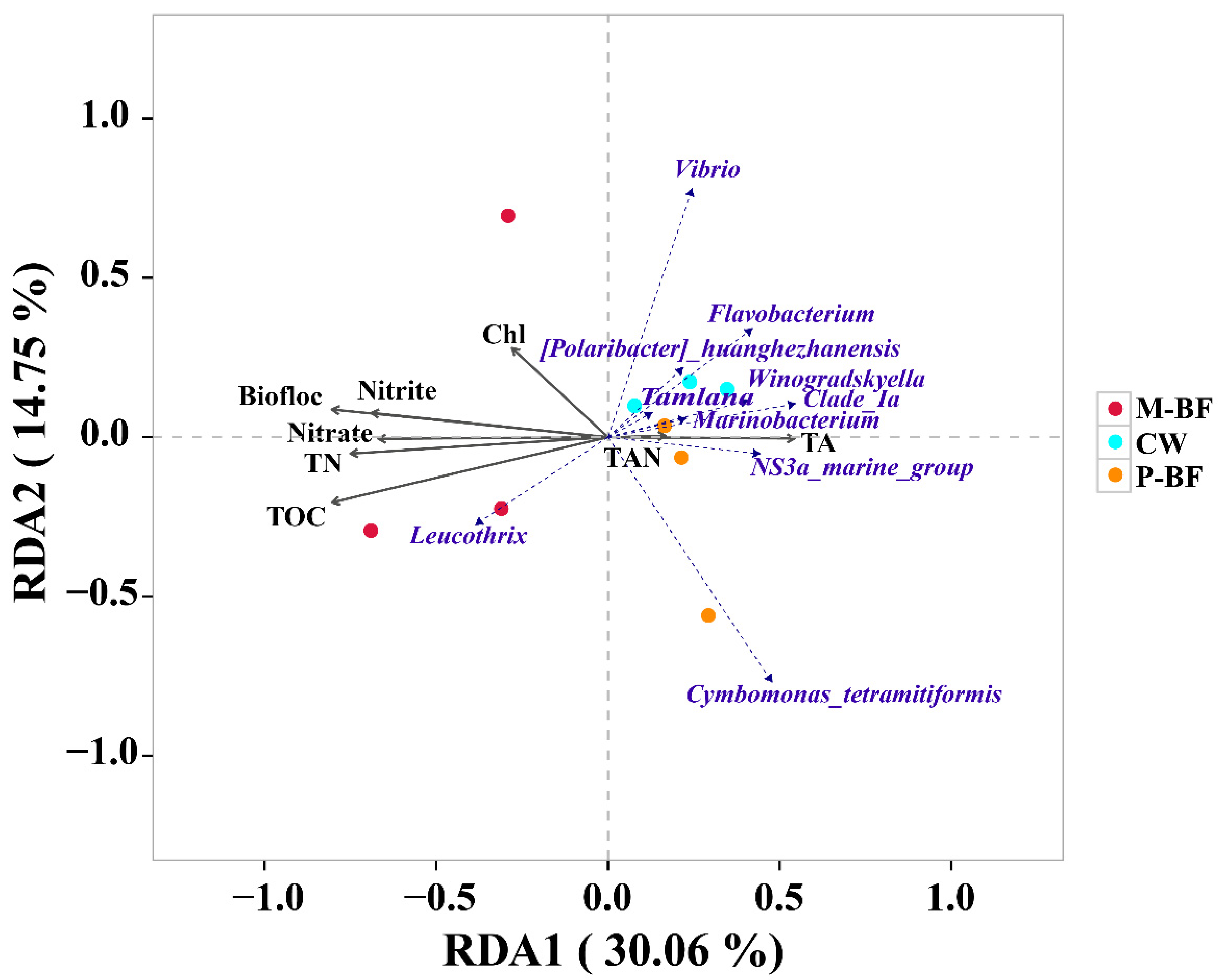
3.3. Correlation Analysis between Environmental Factors and Microbial Community
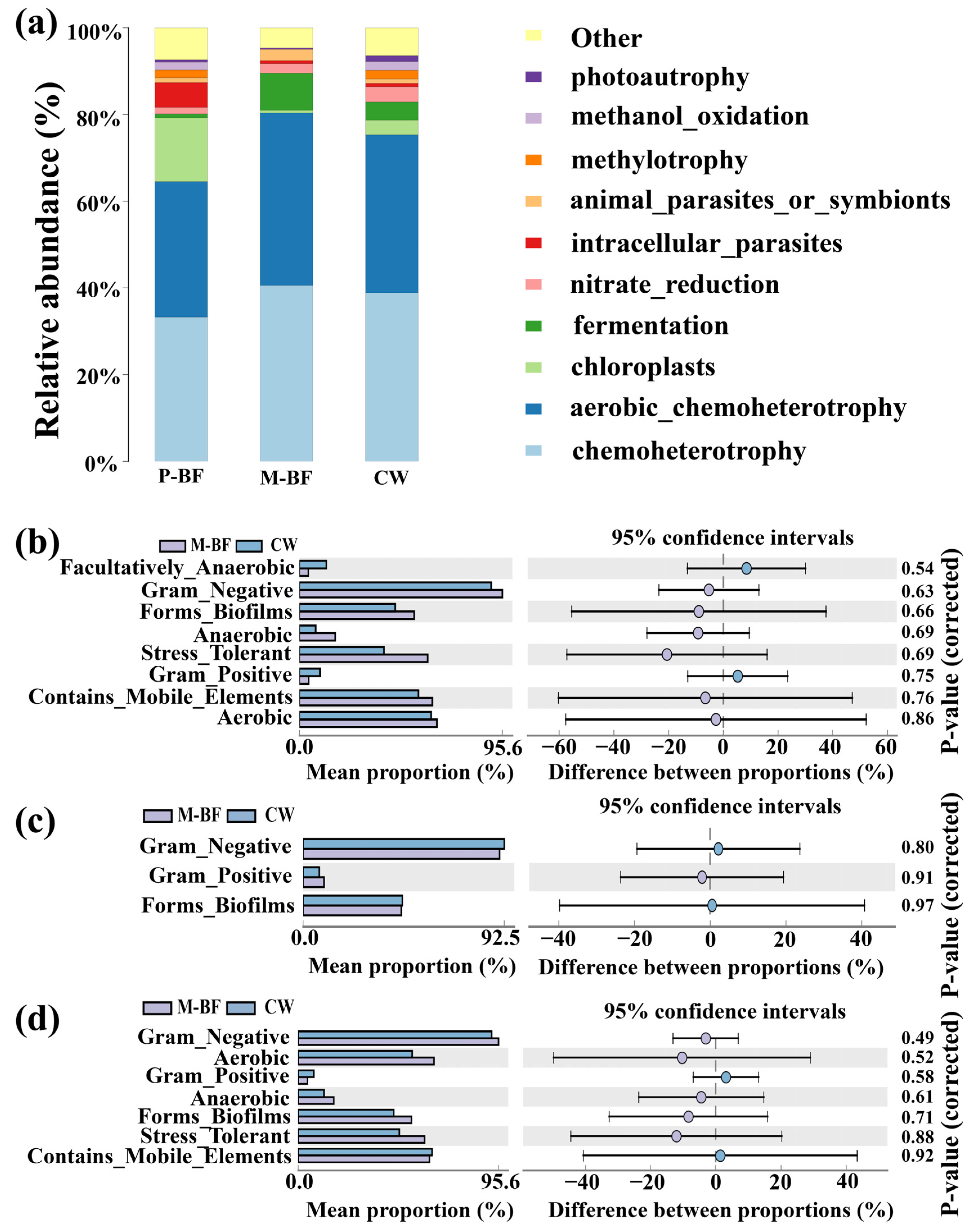
3.4. Functional Prediction in Carbon Adding Environments
3.5. Growth Performance
4. Discussion
4.1. TAN Removal Pathways in Different Systems
4.1.1. Nitrification in Different Systems
4.1.2. Heterotrophic Bacteria Assimilation
4.2. Microbial Composition
4.3. The Correlation between Carbon Sources, Microbial Communities and Environmental Factors
4.4. Practical Value
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Boyd, C.E.; McNevin, A.A. Land Use in Shrimp Aquaculture. World Aquacult. 2018, 49, 28–34. [Google Scholar]
- Haslun, J. Characterization of Bioflocs in a No Water Exchange Super-intensive System for the Production of Food Size Pacific White Shrimp Litopenaeus vannamei. Int. J. Aquac. 2012, 2, 29–38. [Google Scholar] [CrossRef]
- Li, Y.; Boyd, C.E. Laboratory tests of bacterial amendments for accelerating oxidation rates of ammonia, nitrite and organic matter in aquaculture pond water. Aquaculture 2016, 460, 45–58. [Google Scholar] [CrossRef]
- Abu Bakar, N.S.; Mohd Nasir, N.; Lananan, F.; Abdul Hamid, S.H.; Lam, S.S.; Jusoh, A. Optimization of C/N ratios for nutrient removal in aquaculture system culturing African catfish, (Clarias gariepinus) utilizing Bioflocs Technology. Int. Biodeterior. Biodegrad. 2015, 102, 100–106. [Google Scholar] [CrossRef]
- Azim, M.E.; Little, D.C. The biofloc technology (BFT) in indoor tanks: Water quality, biofloc composition, and growth and welfare of Nile tilapia (Oreochromis niloticus). Aquaculture 2008, 283, 29–35. [Google Scholar] [CrossRef]
- Ray, A.J.; Lotz, J.M. Comparing a chemoautotrophic-based biofloc system and three heterotrophic-based systems receiving different carbohydrate sources. Aquac. Eng. 2014, 63, 54–61. [Google Scholar] [CrossRef]
- Ebeling, J.M.; Timmons, M.B.; Bisogni, J.J. Engineering analysis of the stoichiometry of photoautotrophic, autotrophic, and heterotrophic removal of ammonia–nitrogen in aquaculture systems. Aquaculture 2006, 257, 346–358. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.-J.; Morris, T.C.; Samocha, T.M. Effects of C/N ratio on biofloc development, water quality, and performance of Litopenaeus vannamei juveniles in a biofloc-based, high-density, zero-exchange, outdoor tank system. Aquaculture 2016, 453, 169–175. [Google Scholar] [CrossRef]
- Zhu, S.; Chen, S. Effects of organic carbon on nitrification rate in fixed film biofilters. Aquac. Eng. 2001, 25, 1–11. [Google Scholar] [CrossRef]
- Avnimelech, Y. Carbon nitrogen ratio as a control element in aquaculture systems. Aquaculture 1999, 176, 227–235. [Google Scholar] [CrossRef]
- Hargreaves, J.A. Photosynthetic suspended-growth systems in aquaculture. Aquac. Eng. 2006, 34, 344–363. [Google Scholar] [CrossRef]
- Hargreaves, J.A. Biofloc Production Systems for Aquaculture; Southern Regional Aquaculture Center Stoneville: Stoneville, MS, USA, 2013; Volume 4503, pp. 1–11. [Google Scholar]
- Ren, W.; Li, L.; Dong, S.; Tian, X.; Xue, Y. Effects of C/N ratio and light on ammonia nitrogen uptake in Litopenaeus vannamei culture tanks. Aquaculture 2019, 498, 123–131. [Google Scholar] [CrossRef]
- Qiu, T.; Xu, Y.; Gao, M.; Han, M.; Wang, X. Bacterial community dynamics in a biodenitrification reactor packed with polylactic acid/poly (3-hydroxybutyrate-co-3-hydroxyvalerate) blend as the carbon source and biofilm carrier. J. Biosci. Bioeng. 2017, 123, 606–612. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Yang, L.; Wang, J. Denitrification performance and microbial diversity in a packed-bed bioreactor using PCL as carbon source and biofilm carrier. Appl. Microbiol. Biotechnol. 2013, 97, 2725–2733. [Google Scholar] [CrossRef] [PubMed]
- Schneider, O.; Sereti, V.; Eding, E.H.; Verreth, J.A.J. Molasses as C source for heterotrophic bacteria production on solid fish waste. Aquaculture 2006, 261, 1239–1248. [Google Scholar] [CrossRef]
- Fatimah, N.; Pande, G.S.J.; Natrah, F.M.I.; Meritha, W.W.; Widanarni; Sucipto, A.; Ekasari, J. The role of microbial quorum sensing on the characteristics and functionality of bioflocs in aquaculture systems. Aquaculture 2019, 504, 420–426. [Google Scholar] [CrossRef]
- Coyte, K.Z.; Schluter, J.; Foster, K.R. The ecology of the microbiome: Networks, competition, and stability. Science 2015, 350, 663–666. [Google Scholar] [CrossRef]
- Zhang, N.; Luo, G.; Tan, H.; Liu, W.; Hou, Z. Growth, digestive enzyme activity and welfare of tilapia (Oreochromis niloticus) reared in a biofloc-based system with poly-β-hydroxybutyric as a carbon source. Aquaculture 2016, 464, 710–717. [Google Scholar] [CrossRef]
- Kim, M.S.; Min, E.; Kim, J.H.; Koo, J.K.; Kang, J.C. Growth performance and immunological and antioxidant status of Chinese shrimp, Fennerpenaeus chinensis reared in bio-floc culture system using probiotics. Fish Shellfish Immunol. 2015, 47, 141–146. [Google Scholar] [CrossRef]
- Eaton, A. Standard Methods for the Examination of Water and WasteWater; AWWA: Washington DC, USA, 2005. [Google Scholar]
- Gilcreas, F.W. Standard methods for the examination of water and waste water. Am. J. Public Health Nations Health 1966, 56, 387–388. [Google Scholar] [CrossRef]
- Bratvold, D.; Browdy, C.L. Simple electrometric methods for estimating microbial activity in aquaculture ponds. Aquac. Eng. 1998, 19, 29–39. [Google Scholar] [CrossRef]
- Murray, M.G.; Thompson, W.F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 1980, 8, 4321–4325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Haas, B.J.; Gevers, D.; Earl, A.M.; Feldgarden, M.; Ward, D.V.; Giannoukos, G.; Ciulla, D.; Tabbaa, D.; Highlander, S.K.; Sodergren, E.; et al. Chimeric 16S rRNA sequence formation and detection in Sanger and 454-pyrosequenced PCR amplicons. Genome Res. 2011, 21, 494–504. [Google Scholar] [CrossRef] [Green Version]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [Green Version]
- Louca, S.; Parfrey, L.W.; Doebeli, M. Decoupling function and taxonomy in the global ocean microbiome. Science 2016, 353, 1272–1277. [Google Scholar] [CrossRef]
- Ward, T.; Larson, J.; Meulemans, J.; Hillmann, B.; Lynch, J.; Sidiropoulos, D.; Spear, J.R.; Caporaso, G.; Blekhman, R.; Knight, R.; et al. BugBase Predicts Organism Level Microbiome Phenotypes. BioRxiv 2017, 133462. [Google Scholar] [CrossRef]
- Schveitzer, R.; Arantes, R.; Costódio, P.F.S.; do Espírito Santo, C.M.; Arana, L.V.; Seiffert, W.Q.; Andreatta, E.R. Effect of different biofloc levels on microbial activity, water quality and performance of Litopenaeus vannamei in a tank system operated with no water exchange. Aquac. Eng. 2013, 56, 59–70. [Google Scholar] [CrossRef]
- Emilie, C.; Yannick, G.; Kevin, M.; Bénédicte, L.; David, P.; Fabien, P.; Florian, N.; Denis, S. Bacterial community characterization of water and intestine of the shrimp Litopenaeus stylirostrisin a biofloc system. BMC Microbiol. 2016, 16, 157. [Google Scholar] [CrossRef]
- Gerardi, M.H. Wastewater Bacteria; Wiley-Interscience: Hoboken, NJ, USA, 2006. [Google Scholar]
- Martínez-Córdova, L.R.; Vargas-Albores, F.; Garibay-Valdez, E.; Ortíz-Estrada, Á.M.; Porchas-Cornejo, M.A.; Lago-Lestón, A.; Martínez-Porchas, M. Amaranth and wheat grains tested as nucleation sites of microbial communities to produce bioflocs used for shrimp culture. Aquaculture 2018, 497, 503–509. [Google Scholar] [CrossRef]
- Yu, E.; Xie, J.; Wang, J.; Ako, H.; Wang, G.; Chen, Z.; Liu, Y. Surface-attached and suspended bacterial community structure as affected by C/N ratios: Relationship between bacteria and fish production. World J. Microbiol. Biotechnol. 2016, 32, 116. [Google Scholar] [CrossRef]
- Kirchman, D.L. The ecology of Cytophaga-Flavobacteria in aquatic environments. FEMS Microbiol. Ecol. 2002, 39, 91–100. [Google Scholar] [CrossRef]
- Woebken, D.; Fuchs, B.M.; Kuypers, M.M.; Amann, R. Potential interactions of particle-associated anammox bacteria with bacterial and archaeal partners in the Namibian upwelling system. Appl. Environ. Microbiol. 2007, 73, 4648–4657. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Zhu, J.; Zheng, C.; Qiu, H.; Zheng, Z.; Lu, K. Succession of bacterioplankton community in intensive shrimp (Litopenaeus vannamei) aquaculture systems. Aquaculture 2018, 497, 200–213. [Google Scholar] [CrossRef]
- Miranda-Baeza, A.; Mariscal-López, M.d.l.A.; López-Elías, J.A.; Rivas-Vega, M.E.; Emerenciano, M.; Sánchez-Romero, A.; Esquer-Méndez, J.L. Effect of inoculation of the cyanobacteria Oscillatoria sp. on tilapia biofloc culture. Aquacult. Res. 2017, 48, 4725–4734. [Google Scholar] [CrossRef]
- Kim, Y.-S.; Kim, S.-E.; Kim, S.-J.; Jung, H.-K.; Park, J.; Jeon, Y.J.; Kim, D.-H.; Kang, J.-H.; Kim, K.-H. Effects of wheat flour and culture period on bacterial community composition in digestive tracts of Litopenaeus vannamei and rearing water in biofloc aquaculture system. Aquaculture 2021, 531, 735908. [Google Scholar] [CrossRef]
- Goffredi, S.K. Indigenous ectosymbiotic bacteria associated with diverse hydrothermal vent invertebrates. Environ. Microbiol. Rep. 2010, 2, 479–488. [Google Scholar] [CrossRef]
- Hansen, G.H.; Olafsen, J.A. Bacterial Interactions in Early Life Stages of Marine Cold Water Fish. Microb. Ecol. 1999, 38, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Salazar, G.J.; Molina-Garza, Z.J.; Hernández-Acosta, M.; García-Salas, J.A.; Mercado-Hernández, R.; Galaviz-Silva, L. Pathogens in Pacific white shrimp (Litopenaeus vannamei Boone, 1931) and their relationship with physicochemical parameters in three different culture systems in Tamaulipas, Mexico. Aquaculture 2011, 321, 34–40. [Google Scholar] [CrossRef]
- Barcenal, A.; Traifalgar, R.; Jr, V. Anti-Vibrio harveyi Property of Micrococcus luteus Isolated from Rearing Water under Biofloc Technology Culture System. Curr. Res. Bacteriol. 2015, 8, 26–33. [Google Scholar] [CrossRef] [Green Version]
- Dias Schleder, D.; Jatobá, A.; Silva, B.; Ferro, D.; Seiffert, W.; do Nascimento Vieira, F. Soybean protein concentrate in Pacific white shrimp reared in bioflocs: Effect on health and vibrio challenge. Acta Sci. Anim. Sci. 2018, 40, 1–6. [Google Scholar] [CrossRef]
- Jiang, W.; Ren, W.; Li, L.; Dong, S.; Tian, X. Light and carbon sources addition alter microbial community in biofloc-based Litopenaeus vannamei culture systems. Aquaculture 2020, 515, 734572. [Google Scholar] [CrossRef]
- Zhang, H.; Sun, Z.L.; Liu, B.; Xuan, Y.M.; Jiang, M.; Pan, Y.S.; Zhang, Y.M.; Gong, Y.P.; Lu, X.P.; Yu, D.S.; et al. Dynamic changes of microbial communities in Litopenaeus vannamei cultures and the effects of environmental factors. Aquaculture 2016, 455, 97–108. [Google Scholar] [CrossRef]
- Li, J.; Li, F.; Yu, S.; Qin, S.; Wang, G. Impacts of mariculture on the diversity of bacterial communities within intertidal sediments in the Northeast of China. Microb. Ecol. 2013, 66, 861–870. [Google Scholar] [CrossRef]
- Bunse, C.; Pinhassi, J. Marine Bacterioplankton Seasonal Succession Dynamics. Trends Microbiol. 2017, 25, 494–505. [Google Scholar] [CrossRef] [Green Version]
- Lara, G.; Furtado, P.; Hostins, B.; Poersch, L.; Wasielesky Jr, W. Addition of sodium nitrite and biofilm in a Litopenaeus vannamei biofloc culture system. Lat. Am. J. Aquat. Res. 2016, 44, 760–768. [Google Scholar] [CrossRef]
- Xue, Y.; Wei, F.; Jiang, Y.; Li, L.; Dong, S.; Tian, X. Transcriptome signatures of the Pacific white shrimp Litopenaeus vannamei hepatopancreas in response to stress in biofloc culture systems. Fish Shellfish Immunol. 2019, 91, 369–375. [Google Scholar] [CrossRef]
- Emerenciano, M.G.C.; Martínez-Córdova, L.R.; Martínez-Porchas, M.; Miranda-Baeza, A. Biofloc technology (BFT): A tool for water quality management in aquaculture. Water Qual. 2017, 5, 92–109. [Google Scholar] [CrossRef] [Green Version]
- Deng, M.; Chen, J.; Hou, J.; Li, D.; He, X. The effect of different carbon sources on water quality, microbial community and structure of biofloc systems. Aquaculture 2017, 482. [Google Scholar] [CrossRef]
- Wang, G.J.; Yu, E.M.; Xie, J.; Yu, D.G.; Li, Z.F.; Luo, W.; Qiu, L.J.; Zheng, Z.L. Effect of C/N ratio on water quality in zero-water exchange tanks and the biofloc supplementation in feed on the growth performance of crucian carp, Carassius auratus. Aquaculture 2015, 443, 98–104. [Google Scholar] [CrossRef]
- Wei, Y.; Liao, S.-A.; Wang, A.-l. The effect of different carbon sources on the nutritional composition, microbial community and structure of bioflocs. Aquaculture 2016, 465, 88–93. [Google Scholar] [CrossRef]
- Avnimelech, Y. Biofloc Technology: A Practical Guide Book; The World Aquaculture Society: Baton Rouge, LA, USA, 2012; pp. 73–91. [Google Scholar]
- Avnimelech, Y. Feeding with microbial flocs by tilapia in minimal discharge bio-flocs technology ponds. Aquaculture 2007, 264, 140–147. [Google Scholar] [CrossRef]
- Ahmad, I.; Rani, A.M.B.; Verma, A.K.; Maqsood, M. Biofloc technology: An emerging avenue in aquatic animal healthcare and nutrition. Aquacult. Int. 2017, 25, 1215–1226. [Google Scholar] [CrossRef] [Green Version]
- Boley, A.; Muller, W.R.; Haider, G. Biodegradable polymers as solid substrate and biofilm carrier for denitrification in recirculated aquaculture systems. Aquac. Eng. 2000, 22, 75–85. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | CW | M-BF | P-BF |
|---|---|---|---|
| Temperature (°C) | 25.5 ± 1.56 a | 25.5 ± 1.56 a | 25.5 ± 1.56 a |
| DO (mg L−1) | 5.27 ± 0.74 a | 4.83 ± 0.88 a | 4.95 ± 0.84 a |
| pH | 7.7 ± 0.08 a | 7.1 ± 0.07 c | 7.4 ± 0.05 b |
| TA (mg L−1 CaCO3) | 102.67 ± 0.44 a | 81.69 ± 4.98 b | 86.14 ± 1.81 b |
| TOC (mg L−1) | 10.65 ± 3.29 c | 27.01 ± 2.93 a | 18.95 ± 3.15 b |
| Biofloc volume (mL L−1) | 0.00 ± 0.00 b | 3.35 ± 1.10 a | 0.00 ± 0.00 b |
| Chl (mg L−1) | 13.55 ± 1.19 a | 10.48 ± 2.11 a | 3.95 ± 0.57 b |
| TAN ((mg L−1) | 0.13 ± 0.16 a | 0.17 ± 0.04 a | 0.23 ± 0.05 a |
| NO2−-N (mg L−1) | 0.82 ± 0.26 c | 6.44 ± 1.70 a | 3.38 ± 0.49 b |
| NO3−-N (mg L−1) | 0.94 ± 0.09 c | 4.51 ± 0.90 a | 2.79 ± 0.20 b |
| TN (mg L−1) | 5.69 ± 0.30 c | 13.65 ± 0.58 a | 9.75 ± 0.39 b |
| Nitrification rate (mg L−1 h−1) | 0.39 ± 0.06 a | 0.10±0.02 b | 0.12±0.01 b |
| Parameters | CW | M-BF | P-BF |
|---|---|---|---|
| final weight | 5.17 ± 0.81 b | 6.94 ± 1.17 ab | 8.04 ± 0.94 a |
| Specific growth rate (%/day) | 1.84 ± 0.00 b | 2.67 ± 0.00 ab | 3.11 ± 0.00 a |
| Growth rate (g/wk) | 0.49 ± 0.16 b | 0.85 ± 0.23 ab | 1.07 ± 0.19 a |
| Feed conversion ratio (kg/kg) | 1.95 ± 0.54 a | 1.12 ± 0.30 b | 0.86 ± 0.17 b |
| Survival rate (%) | 28.33 ± 0.04 b | 45.56 ± 0.11 ab | 61.11 ± 0.29 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xue, Y.; Li, L.; Dong, S.; Gao, Q.; Tian, X. The Effects of Different Carbon Sources on the Production Environment and Breeding Parameters of Litopenaeus vannamei. Water 2021, 13, 3584. https://doi.org/10.3390/w13243584
Xue Y, Li L, Dong S, Gao Q, Tian X. The Effects of Different Carbon Sources on the Production Environment and Breeding Parameters of Litopenaeus vannamei. Water. 2021; 13(24):3584. https://doi.org/10.3390/w13243584
Chicago/Turabian StyleXue, Yiming, Li Li, Shuanglin Dong, Qinfeng Gao, and Xiangli Tian. 2021. "The Effects of Different Carbon Sources on the Production Environment and Breeding Parameters of Litopenaeus vannamei" Water 13, no. 24: 3584. https://doi.org/10.3390/w13243584
APA StyleXue, Y., Li, L., Dong, S., Gao, Q., & Tian, X. (2021). The Effects of Different Carbon Sources on the Production Environment and Breeding Parameters of Litopenaeus vannamei. Water, 13(24), 3584. https://doi.org/10.3390/w13243584