
Abiotic Aggregation of Organic Matter in Coastal and Estuarine Waters: Cases in the Eastern Long Island Sound, USA

Abstract
1. Introduction
2. Materials and Methods
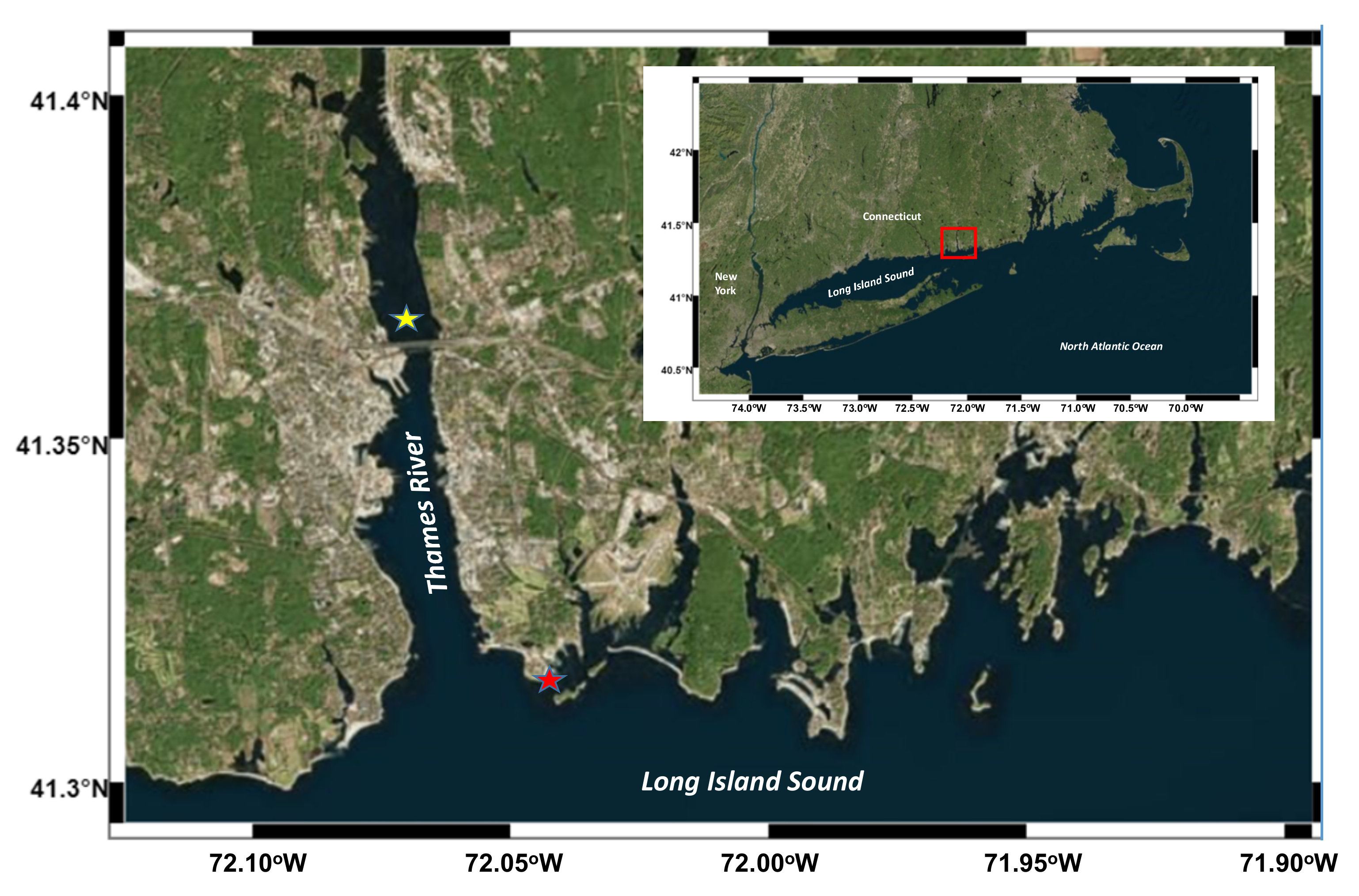
2.1. Study Sites and Field Sampling
2.2. Roller Table Experiments
2.3. Chemical and Biological Determinations
2.4. Data Analysis
3. Results
3.1. Environmental Conditions
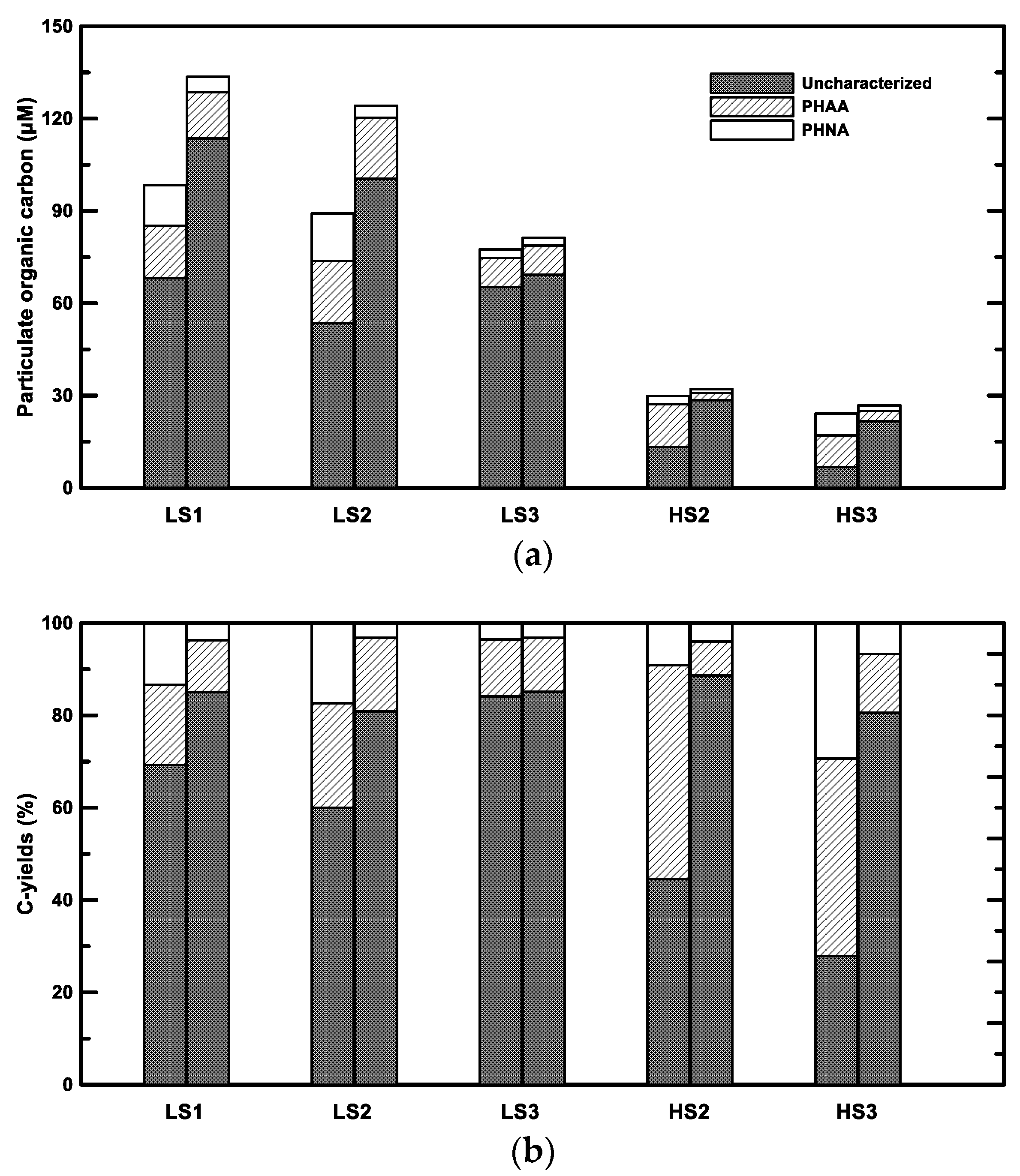
3.2. Concentrations of Bulk Organic Matter (OM) and Bacterial Abundance
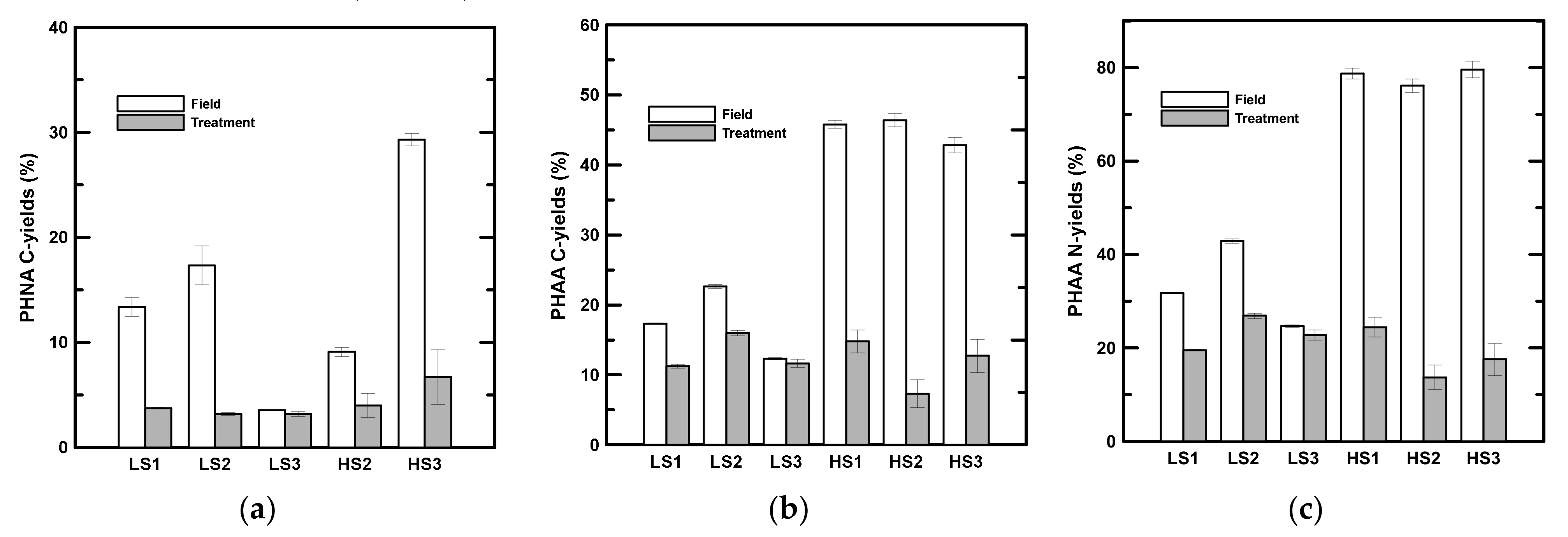
3.3. Concentrations of Bulk Neutral Aldose and Amino Acid
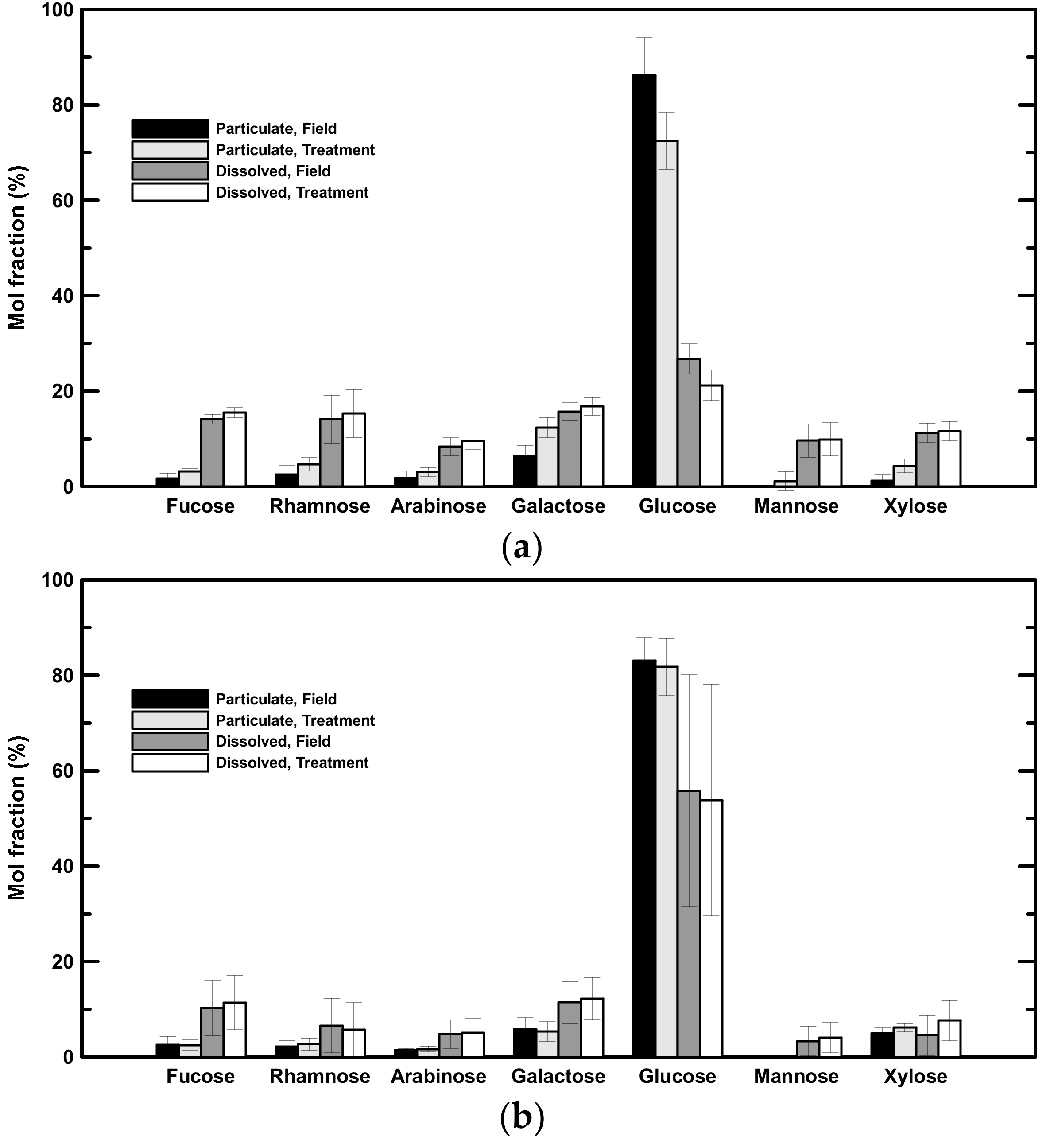
3.4. Concentrations of Compound-Specific Neutral Aldose and Amino Acid
4. Discussion
4.1. Aggregation of Bulk Organic Matter
4.2. Changes in Biologically-Labile Components
4.3. Changes in Uncharacterized Component
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kepkay, P.E. Colloids and the ocean carbon cycle. In Marine Chemistry; Wangersky, P.J., Ed.; Springer: New York, NY, USA, 2000; pp. 35–56. [Google Scholar]
- Eppley, R.W.; Peterson, B.J. Particulate organic matter flux and planktonic new production in the deep ocean. Nature 1979, 282, 677–680. [Google Scholar] [CrossRef]
- Thornton, D.C.O. Diatom aggregation in the sea: Mechanisms and ecological implications. Eur. J. Phycol. 2002, 37, 149–167. [Google Scholar] [CrossRef]
- Simon, M.; Grossart, H.P.; Schweitzer, B.; Ploug, H. Microbial ecology of organic aggregates in aquatic ecosystems. Aquat. Microb. Ecol. 2002, 28, 175–211. [Google Scholar] [CrossRef]
- Kerner, M.; Hohenberg, H.; Ertl, S.; Reckermann, M.; Spitzy, A. Self-organization of dissolved organic matter to micelle-like microparticles in river water. Nature 2003, 422, 150–154. [Google Scholar] [CrossRef] [PubMed]
- Kepkay, P.E. Particle aggregation and biological reactivity of colloids. Mar. Ecol. Prog. Ser. 1994, 109, 293–304. [Google Scholar] [CrossRef]
- Chin, W.-C.; Orellana, M.V.; Verdugo, P. Spontaneous assembly of marine dissolved organic matter into polymer gels. Nature 1998, 391, 568–572. [Google Scholar] [CrossRef]
- Alldredge, A.L.; Silver, M.W. Characteristics, dynamics and significance of marine snow. Prog. Oceanogr. 1988, 20, 41–82. [Google Scholar] [CrossRef]
- Shanks, A.L.; Edmondson, E.W. Laboratory-made marine snow: A biological model of the real thing. Mar. Biol. 1989, 101, 463–470. [Google Scholar] [CrossRef]
- Drapeau, D.T.; Dam, H.G.; Grenier, G. An improved flocculator design for use in particle aggregation experiments. Limnol. Oceanogr. 1994, 39, 723–729. [Google Scholar] [CrossRef]
- Mopper, K.; Zhou, J.; Sri Ramana, K.; Passow, U.; Dam, H.G.; Drapeau, D.T. The role of surface-active carbohydrates in the flocculation of a diatom bloom in a mesocosm. Deep.-Sea Res. Part II 1995, 42, 47–73. [Google Scholar] [CrossRef]
- Alldredge, A.L. The carbon, nitrogen, and mass content of marine snow as a function of aggregate size. Deep.-Sea Res. 1998, 45, 529–541. [Google Scholar] [CrossRef]
- Jackson, G.A.; Lochmann, S. Modeling coagulation of algae in marine ecosystem. In Environmental Particles; Buffle, J., van Leeuwen, H.P., Eds.; Lewis Publisher: Boca Raton, FL, USA, 1993; Volume 2, pp. 387–414. [Google Scholar]
- Kiørboe, T.; Andersen, K.P.; Dam, H.G. Coagulation efficiency and aggregate formation in marine phytoplankton. Mar. Biol. 1990, 107, 235–245. [Google Scholar] [CrossRef]
- Grossart, H.-P.; Berman, T.; Simon, M.; Pohlmann, K. Occurrence and microbial dynamics of macroscopic organic aggregates (lake snow) in Lake Kinneret, Israel, in fall. Aquat. Microb. Ecol. 1998, 14, 59–67. [Google Scholar] [CrossRef]
- Artolozaga, I.; Santamaría, E.; López, A.; Ayo, B.; Iriberri, J. Succession of bacterivorous protists on laboratory-made marine snow. J. Plankton Res. 1997, 19, 1429–1440. [Google Scholar] [CrossRef]
- Knoll, S.; Zwisler, W.; Simon, M. Bacterial colonization of early stages of limnetic diatom microaggregates. Aquat. Microb. Ecol. 2001, 25, 141–150. [Google Scholar] [CrossRef]
- Engel, A.; Meyerhöfer, M.; von Bröckel, K. Chemical and biological composition of suspended particles and aggregates in the Baltic Sea in summer (1999). Estuar. Coast. Shelf Sci. 2002, 55, 729–741. [Google Scholar] [CrossRef]
- Unanue, M.A.; Azua, I.; Arrieta, J.M.; Herndl, G.J.; Iriberri, J. Laboratory-made particles as a useful approach to analyse microbial processes in marine aggregates. FEMS Microbiol. Ecol. 1998, 26, 325–334. [Google Scholar] [CrossRef][Green Version]
- Quigg, A.; Santschi, P.H.; Burd, A.; Chin, W.-C.; Kamalanathan, M.; Xu, C.; Ziervogel, K. From nano-gels to marine snow: A synthesis of gel formation processes and modeling efforts involved with particle flux in the ocean. Gels 2021, 7, 114. [Google Scholar] [CrossRef] [PubMed]
- Welsh, B.L.; Eller, F.C. Mechanisms controlling summertime oxygen depletion in western Long Island Sound. Estuaries 1991, 14, 265–278. [Google Scholar] [CrossRef]
- Benner, R.; Strom, M. A critical evaluation of the analytical blank associated with DOC measurements by high-temperature catalytic oxidation. Mar. Chem. 1993, 41, 153–160. [Google Scholar] [CrossRef]
- Svensson, E.; Skoog, A.; Amend, J.P. Concentration and distribution of dissolved amino acids in a shallow hydrothermal system, Vulcano Island (Italy). Org. Geochem. 2004, 35, 1001–1014. [Google Scholar] [CrossRef]
- Skoog, A.; Benner, R. Aldoses in various size fractions of marine organic matter: Implications for carbon cycling. Limnol. Oceanogr. 1997, 42, 1803–1813. [Google Scholar] [CrossRef]
- Parsons, T.R.; Maita, Y.; Lalli, C.M. A Manual of Chemical and Biological Methods for Seawater Analysis; Pergamon Press: New York, NY, USA, 1984; p. 173. [Google Scholar]
- Hobbie, J.E.; Daley, R.J.; Jasper, S. Use of nuclepore filters for counting bacteria by fluorescence microscopy. Appl. Environ. Microb. 1977, 33, 1225–1228. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Fuhrman, J.A. Relationships between biovolume and biomass of naturally derived marine bacterioplankton. Appl. Environ. Microb. 1987, 53, 1298–1303. [Google Scholar] [CrossRef] [PubMed]
- Rorabacher, D.B. Statistical treatment for rejection of deviant values: Critical values of Dixon’s "Q" parameter and related subrange ratios at the 95% confidence level. Anal. Chem. 1991, 63, 139–146. [Google Scholar] [CrossRef]
- Ortiz, V.L.; Mason, R.P.; Ward, J.E. An examination of the factors influencing mercury and methylmercury particulate distributions, methylation and demethylation rates in laboratory-generated marine snow. Mar. Chem. 2015, 177, 753–762. [Google Scholar] [CrossRef] [PubMed]
- Jackson, G.A. Particle trajectories in a rotating cylinder: Implications for aggregation incubations. Deep.-Sea Res. I 1994, 41, 429–437. [Google Scholar] [CrossRef]
- Burban, P.-Y.; Lick, W.; Lick, J. The flocculation of grained sediments in estuarine waters. J. Geophys. Res. 1989, 94, 8323–8330. [Google Scholar] [CrossRef]
- Spicer, P.T.; Pratsinis, S. Shear-induced floccultion: The evolution of floc structure and the shape of the size distribution at steady state. Water Res. 1996, 30, 1049–1056. [Google Scholar] [CrossRef]
- Tsai, C.-H.; Iacobellis, S.; Lick, W. Flocculation of fine-grained lake sediments due to a uniform shear stress. J. Great Lakes Res. 1987, 13, 135–146. [Google Scholar] [CrossRef]
- Kepkay, P.E.; Johnson, B.D. Microbial response to organic particle generation by surface coagulation in seawater. Mar. Ecol. Prog. Ser. 1988, 48, 193–198. [Google Scholar] [CrossRef]
- Kepkay, P.E.; Johnson, B.D. Coagulation on bubbles allows the microbial respiration of oceanic dissolved organic carbon. Nature 1989, 385, 63–65. [Google Scholar] [CrossRef]
- Ducklow, H.W.; Hill, S.M.; Gardner, W.D. Bacterial growth and the decomposition of particulate organic carbon collected in sediment traps. Cont. Shelf Res. 1985, 4, 445–464. [Google Scholar] [CrossRef]
- Sholkovitz, E.R. Flocculation of dissolved organic and inorganic matter during the mixing of river water and seawater. Geochim. Cosmochim. Acta 1976, 40, 831–845. [Google Scholar] [CrossRef]
- Amon, R.M.W.; Benner, R. Bacterial utilization of different size classes of dissolved organic matter. Limnol. Oceanogr. 1996, 41, 41–51. [Google Scholar] [CrossRef]
- Chen, T.-Y.; Skoog, A. Aggregation of organic matter in coastal waters: A dilemma of using a Couette flocculator. Cont. Shelf Res. 2017, 139, 62–70. [Google Scholar] [CrossRef]
- Cowie, G.L.; Hedges, J.I. Biochemical indicators of diagenetic alteration in natural organic matter mixtures. Nature 1994, 369, 304–307. [Google Scholar] [CrossRef]
- Hernes, P.J.; Hedges, J.I.; Peterson, M.L.; Wakeham, S.G.; Lee, C. Neutral carbohydrate geochemistry of particulate material in the central equatorial Pacific. Deep.-Sea Res. II 1996, 43, 1181–1204. [Google Scholar] [CrossRef]
- Skoog, A.; Alldredge, A.; Passow, U.; Dunne, J.; Murray, J. Neutral aldoses as source indicators for marine snow. Mar. Chem. 2008, 108, 195–206. [Google Scholar] [CrossRef][Green Version]
- Benner, R.; Kaiser, K. Abundance of amino sugars and peptidoglycan in marine particulate and dissolved organic matter. Limnol. Oceanogr. 2003, 48, 118–128. [Google Scholar] [CrossRef]
- Alldredge, A.L.; Passow, U.; Logan, B.E. The abundance and significance of a class of large, transparent organic particles in the ocean. Deep.-Sea Res. I 1993, 40, 1131–1140. [Google Scholar] [CrossRef]
- Ogawa, H.; Amagai, Y.; Koike, I.; Kaiser, K.; Benner, R. Production of refractory dissolved organic matter by bacteria. Science 2001, 292, 917–920. [Google Scholar] [CrossRef] [PubMed]
- Sholkovitz, E.R.; Boyle, E.A.; Price, N.B. The removal of dissolved humic acids and iron during esturine mixing. Earth Planet. Sci. Lett. 1978, 40, 130–136. [Google Scholar] [CrossRef]
- Fox, L.E. The removal of dissolved humic acid during esturine mixing. Estuar. Coast. Shelf Sci. 1983, 16, 431–440. [Google Scholar] [CrossRef]
- Tremblay, L.; Gagné, J.-P. Organic matter distribution and reactivity in the waters of a large estuarine system. Mar. Chem. 2009, 116, 1–12. [Google Scholar] [CrossRef]
- Flores-Cervantes, D.X.; Plata, D.; MacFarlane, J.K.; Reddy, C.M.; Gschwend, P.M. Black carbon in marine particulate organic carbon: Inputs and cycling of highly recalcitrant organic carbon in the Gulf of Maine. Mar. Chem. 2009, 113, 172–181. [Google Scholar] [CrossRef]
- Santschi, P.H.; Xu, C.; Schwehr, K.A.; Lin, P.; Sun, L.; Chin, W.-C.; Kamalanathan, M.; Bacosa, H.P.; Quigg, A. Can the protein/carbohydrate (P/C) ratio of exopolymeric substances (EPS) be used as a proxy for their ‘stickiness’ and aggregation propensity? Mar. Chem. 2020, 218, 103734. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Low-Salinity Samples | High-Salinity Samples | ||||||
|---|---|---|---|---|---|---|---|
| Parameter | Unit | LS1 | LS2 | LS3 | HS1 | HS2 | HS3 |
| Salinity | 6.3 | 6.3 | 6.8 | 26.7 | 26.2 | 21.4 | |
| NOx, Field | μM | 27.0 | 27.0 | 17.1 | 0.88 | 1.90 | 1.52 |
| PO4, Field | μM | 0.53 | 0.56 | 0.88 | 0.58 | 0.60 | 0.97 |
| N/P ratio Field | mol/mol | 51 | 48 | 19 | 1.5 | 3.2 | 1.6 |
| DOC Field | μM | 310.3 | 313.4 | 425.0 | 141.1 | 141.4 | 170.6 |
| POC Field | μM | 98.3 | 89.2 | 77.5 | 27.3 | 29.9 | 24.1 |
| POC Treatment | μM | 133.6 | 124.2 | 81.3 | 29.7 | 32.1 | 26.8 |
| ΔPOC | μM | 35.3 | 35.0 | 3.8 | 2.4 | 2.2 | 2.7 |
| ΔPOC | % | 35.9 | 39.2 | 4.9 | 8.8 | 7.4 | 11.2 |
| PON Field | μM | 14.8 | 13.0 | 10.7 | 4.32 | 4.99 | 3.58 |
| PON Treatment | μM | 21.2 | 20.4 | 11.6 | 4.93 | 4.80 | 5.29 |
| ΔPON | μM | 6.4 | 7.4 | 0.9 | 0.61 | −0.19 | 1.71 |
| ΔPON | % | 43.2 | 56.9 | 8.4 | 14.1 | −3.8 | 47.8 |
| Part. C/N Field | mol/mol | 6.65 | 6.89 | 7.25 | 6.32 | 5.99 | 6.72 |
| Part. C/N Treatment | mol/mol | 6.41 | 6.09 | 7.15 | 6.04 | 6.67 | 5.50 |
| Bact. # Field | 105 cells ml−1 | 7.52 | 11.8 | 14.6 | 9.64 | 14.1 | 13.0 |
| Bact. # Treatment | 105 cells ml−1 | 28.1 | 68.2 | 21.9 | 30.2 | 31.8 | 27.2 |
| BBC Field | μM C | 1.25 | 1.97 | 2.43 | 1.61 | 2.34 | 2.17 |
| BBC Treatment | μM C | 4.69 | 11.37 | 3.65 | 5.02 | 5.31 | 4.53 |
| ΔBBC | μM C | 3.44 | 9.40 | 1.22 | 3.41 | 2.97 | 2.36 |
| BBN Field | μM N | 0.25 | 0.40 | 0.49 | 0.32 | 0.47 | 0.43 |
| BBN Treatment | μM N | 0.94 | 2.28 | 0.73 | 1.00 | 1.06 | 0.91 |
| ΔBBN | μM N | 0.69 | 1.88 | 0.24 | 0.68 | 0.59 | 0.47 |
| Low-Salinity Samples | High-Salinity Samples | |||||
|---|---|---|---|---|---|---|
| Parameter | LS1 | LS2 | LS3 | HS1 | HS2 | HS3 |
| (POC) | + | + | + | + | + | + |
| (DOC) | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ |
| BB | + | + | + | + | + | + |
| C/Np | ‒ | ‒ | ‒ | × | + | ‒ |
| (PHNA) | ‒ | ‒ | × | n.a. | ‒ | ‒ |
| PHNA-CY | ‒ | ‒ | × | n.a. | ‒ | ‒ |
| (DHNA) | × | ‒ | × | × | × | ‒ |
| DHNA-CY | × | ‒ | × | × | × | ‒ |
| (PHAA) | × | × | × | ‒ | ‒ | ‒ |
| PHAA-CY | ‒ | ‒ | × | ‒ | ‒ | ‒ |
| PHAA-NY | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ |
| (DHAA) | × | × | × | × | × | × |
| DHAA-CY | × | × | × | × | × | × |
| DHAA-NY | × | × | × | × | × | × |
| Fucose | Rhamnose | Arabinose | Galactose | Glucose | Mannose | Xylose | Total | |||
|---|---|---|---|---|---|---|---|---|---|---|
| Exp | Sample | Group | (nM) | (nM) | (nM) | (nM) | (nM) | (nM) | (nM) | (nM) |
| LS1 | Particulate | Field | 25.9 | 39.7 | 24.8 | 141.1 | 1949.4 | - | 14.7 | 2195.7 |
| Particulate | Treatment | 21.7 | 30.0 | 19.6 | 110.1 | 627.3 | - | 29.1 | 837.8 | |
| Dissolved | Field | 185.6 | 113.6 | 89.7 | 238.9 | 367.7 | 184.5 | 182.5 | 1362.4 | |
| Dissolved | Treatment | 196.5 | 151.1 | 113.0 | 238.6 | 278.2 | 168.8 | 181.7 | 1328.0 | |
| LS2 | Particulate | Field | 25.5 | 33.0 | 18.9 | 110.3 | 2386.7 | - | 6.3 | 2580.7 |
| Particulate | Treatment | 19.6 | 27.5 | 17.2 | 66.7 | 510.8 | - | 23.1 | 664.9 | |
| Dissolved | Field | 182.5 | 225.5 | 112.9 | 187.0 | 404.7 | 116.1 | 127.7 | 1356.5 | |
| Dissolved | Treatment | 192.6 | 204.5 | 117.4 | 178.4 | 234.4 | 109.1 | 125.5 | 1162.0 | |
| LS3 | Particulate | Field | 14.8 | 22.6 | 17.1 | 42.6 | 379.4 | - | 13.4 | 489.9 |
| Particulate | Treatment | 16.9 | 26.8 | 18.1 | 60.6 | 282.9 | - | 25.8 | 431.0 | |
| Dissolved | Field | 203.4 | 231.5 | 136.4 | 209.8 | 311.7 | 90.8 | 145.4 | 1328.9 | |
| Dissolved | Treatment | 194.0 | 219.3 | 131.1 | 220.2 | 289.9 | 96.5 | 133.6 | 1284.6 | |
| HS1 | Particulate | Field | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. |
| Particulate | Treatment | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | |
| Dissolved | Field | 147.0 | 116.4 | 67.0 | 134.8 | 285.7 | 55.3 | 82.1 | 888.2 | |
| Dissolved | Treatment | 163.8 | 74.3 | 84.3 | 125.2 | 415.2 | 85.5 | 100.5 | 1048.7 | |
| HS2 | Particulate | Field | 17.4 | 14.2 | 7.8 | 34.5 | 364.5 | - | 18.9 | 457.3 |
| Particulate | Treatment | 7.0 | 7.8 | 4.5 | 14.7 | 167.7 | - | 14.7 | 216.3 | |
| Dissolved | Field | 226.2 | 95.8 | 68.0 | 280.1 | 3417.1 | - | 148.7 | 4236.0 | |
| Dissolved | Treatment | 140.2 | 102.4 | 77.6 | 244.5 | 2227.0 | - | 134.9 | 2926.5 | |
| HS3 | Particulate | Field | 14.8 | 14.8 | 14.6 | 48.0 | 1030.5 | - | 68.7 | 1191.4 |
| Particulate | Treatment | 5.1 | 5.6 | 3.8 | 11.8 | 260.0 | - | 16.1 | 302.5 | |
| Dissolved | Field | 141.1 | 69.7 | 81.1 | 197.3 | 858.4 | 60.3 | 165.6 | 1573.5 | |
| Dissolved | Treatment | 123.0 | 57.4 | 40.0 | 146.6 | 407.6 | 35.4 | 77.5 | 887.4 |
| . | ALA | ARG | ASP | GLU | GLY | HIS | ILE | LEU | LYS | MET | ORN | PHE | SER | THR | TYR | VAL | BALA | GABA | Total | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Exp | Sample | Group | (nM) | (nM) | (nM) | (nM) | (nM) | (nM) | (nM) | (nM) | (nM) | (nM) | (nM) | (nM) | (nM) | (nM) | (nM) | (nM) | (nM) | (nM) | (nM) |
| LS1 | Particulate | Field | 497.0 | 192.0 | 387.4 | 420.4 | 405.2 | 55.5 | 211.0 | 334.1 | 286.0 | 16.9 | - | 151.4 | 232.2 | 225.5 | 5.9 | 300.0 | - | 4.7 | 3725.2 |
| Particulate | Treatment | 449.6 | 188.9 | 366.6 | 399.0 | 432.4 | 45.6 | 181.1 | 301.8 | 260.4 | 23.7 | - | 138.0 | 263.8 | 229.4 | 57.2 | 259.5 | 15.3 | 4.9 | 3617.2 | |
| Dissolved | Field | 90.6 | 26.3 | 183.4 | 117.0 | 185.7 | - | 0.5 | - | - | 2.0 | - | - | 83.9 | 22.4 | - | 82.6 | - | - | 794.5 | |
| Dissolved | Treatment | 49.4 | 37.3 | 230.7 | 136.5 | 228.0 | - | 26.6 | - | - | 26.6 | - | - | 101.9 | 42.7 | - | 90.9 | - | - | 970.7 | |
| LS2 | Particulate | Field | 579.6 | 220.3 | 366.4 | 407.8 | 556.0 | 55.7 | 213.0 | 392.9 | 364.1 | 13.0 | - | 184.8 | 294.3 | 365.1 | 80.9 | 302.7 | 16.7 | 3.2 | 4416.3 |
| Particulate | Treatment | 647.7 | 255.3 | 437.0 | 504.4 | 606.7 | 69.0 | 254.2 | 440.2 | 417.7 | 39.0 | - | 207.0 | 303.4 | 400.3 | 71.1 | 361.6 | 5.8 | 6.4 | 5026.8 | |
| Dissolved | Field | - | - | 139.6 | 108.0 | - | - | - | - | - | 0.0 | - | - | 415.3 | 280.9 | 56.7 | - | - | - | 1100.4 | |
| Dissolved | Treatment | - | - | 170.8 | 97.2 | 36.8 | - | - | - | - | 0.0 | - | - | 422.5 | 336.7 | 50.3 | - | - | - | 1114.2 | |
| LS3 | Particulate | Field | 277.2 | 115.7 | 242.7 | 243.9 | 245.2 | 28.5 | 110.1 | 178.9 | 122.1 | 6.8 | - | 79.5 | 141.3 | 130.3 | 24.6 | 156.1 | - | 3.5 | 2106.5 |
| Particulate | Treatment | 272.8 | 114.2 | 254.3 | 250.2 | 253.5 | 28.1 | 109.7 | 172.7 | 126.6 | 14.0 | - | 79.1 | 145.0 | 137.3 | 1.2 | 156.3 | - | 4.7 | 2119.8 | |
| Dissolved | Field | - | 88.5 | 188.7 | 113.0 | 14.1 | - | - | - | - | 0.0 | - | - | 379.1 | 324.2 | 70.3 | - | - | - | 1177.9 | |
| Dissolved | Treatment | - | 114.5 | 171.5 | 91.6 | 7.8 | - | - | - | - | 0.0 | - | - | 393.7 | 303.2 | 51.1 | - | - | - | 1187.0 | |
| HS1 | Particulate | Field | 307.2 | 137.4 | 286.1 | 311.2 | 362.4 | 40.8 | 134.6 | 217.0 | 167.9 | 43.4 | - | 101.3 | 207.7 | 160.3 | 72.3 | 181.6 | - | 3.7 | 2734.8 |
| Particulate | Treatment | 113.5 | 45.4 | 125.3 | 123.3 | 129.7 | 14.0 | 51.5 | 78.5 | 56.8 | 4.6 | - | 38.0 | 66.8 | 62.1 | 2.7 | 69.8 | - | 2.4 | 984.2 | |
| Dissolved | Field | - | 17.9 | 121.9 | 115.3 | 8.5 | - | - | - | - | 0.0 | - | - | 5.9 | 100.2 | 15.1 | - | - | - | 384.9 | |
| Dissolved | Treatment | - | 7.0 | 86.8 | 46.0 | - | - | - | - | - | 0.0 | - | - | 4.9 | 60.2 | 18.6 | - | - | - | 223.4 | |
| HS2 | Particulate | Field | 381.4 | 146.7 | 328.3 | 341.1 | 364.6 | 37.9 | 152.4 | 245.9 | 235.2 | 12.3 | - | 119.4 | 179.3 | 248.6 | 36.4 | 211.1 | - | 4.5 | 3045.0 |
| Particulate | Treatment | 54.6 | 21.3 | 56.7 | 53.2 | 62.6 | 27.0 | 23.6 | 36.9 | 34.5 | 3.2 | - | 19.1 | 29.9 | 35.4 | 13.2 | 32.4 | 3.2 | 0.6 | 507.3 | |
| Dissolved | Field | 100.7 | - | 111.7 | 57.9 | 220.0 | - | - | - | - | 84.2 | - | - | - | - | - | 41.8 | - | - | 616.2 | |
| Dissolved | Treatment | 107.5 | - | 106.8 | 48.7 | 205.0 | - | - | - | - | 92.8 | - | - | - | - | - | 33.8 | - | - | 594.6 | |
| HS3 | Particulate | Field | 267.2 | 109.0 | 249.1 | 252.5 | 317.2 | 24.2 | 106.8 | 179.5 | 164.9 | 7.5 | - | 87.8 | 163.2 | 183.3 | 30.1 | 147.9 | 14.1 | 4.1 | 2308.7 |
| Particulate | Treatment | 85.6 | 33.0 | 91.6 | 83.1 | 111.4 | 9.4 | 37.7 | 60.0 | 46.4 | 3.3 | - | 29.7 | 54.0 | 51.1 | 12.9 | 51.8 | 1.6 | 1.1 | 763.6 | |
| Dissolved | Field | 92.3 | - | 94.6 | 59.7 | 117.1 | - | - | - | - | 66.5 | - | - | - | - | - | 29.2 | - | - | 459.4 | |
| Dissolved | Treatment | 80.0 | - | 77.0 | 32.3 | 98.8 | - | - | - | - | 53.7 | - | - | - | - | - | 13.0 | - | - | 354.7 |
| NA-C | NA-CY | AA-C | AA-CY | AA-N | AA-NY | |||
|---|---|---|---|---|---|---|---|---|
| Exp | Sample | Group | (μM C) | (%) | (μM C) | (%) | (μM N) | (%) |
| LS1 | Particulate | Field | 13.13 | 13.4% | 17.04 | 17.3% | 4.70 | 31.8% |
| Particulate | Treatment | 4.98 | 3.7% | 15.03 | 11.3% | 4.14 | 19.6% | |
| Dissolved | Field | 7.90 | 2.5% | 2.89 | 0.9% | 0.87 | 2.6% | |
| Dissolved | Treatment | 7.67 | 2.9% | 3.66 | 1.4% | 1.01 | 3.9% | |
| LS2 | Particulate | Field | 15.46 | 17.3% | 20.21 | 22.7% | 5.55 | 42.9% |
| Particulate | Treatment | 3.95 | 3.2% | 19.84 | 16.0% | 5.48 | 26.9% | |
| Dissolved | Field | 7.90 | 2.5% | 3.98 | 1.3% | 1.00 | 3.0% | |
| Dissolved | Treatment | 6.73 | 2.6% | 4.31 | 1.7% | 1.11 | 4.0% | |
| LS3 | Particulate | Field | 2.73 | 3.5% | 9.55 | 12.3% | 2.63 | 24.7% |
| Particulate | Treatment | 2.59 | 3.2% | 9.48 | 11.7% | 2.65 | 22.7% | |
| Dissolved | Field | 7.69 | 1.8% | 4.95 | 1.2% | 1.44 | 4.1% | |
| Dissolved | Treatment | 7.44 | 2.0% | 4.70 | 1.3% | 1.48 | 4.3% | |
| HS1 | Particulate | Field | n.a. | n.a. | 12.50 | 45.8% | 3.40 | 78.7% |
| Particulate | Treatment | n.a. | n.a. | 4.39 | 14.8% | 1.21 | 24.5% | |
| Dissolved | Field | 5.18 | 3.7% | 1.74 | 1.2% | 0.44 | 3.1% | |
| Dissolved | Treatment | 6.11 | 4.5% | 1.04 | 0.8% | 0.24 | 1.7% | |
| HS2 | Particulate | Field | 2.72 | 9.1% | 13.87 | 46.4% | 3.80 | 76.1% |
| Particulate | Treatment | 1.28 | 4.0% | 2.35 | 7.3% | 0.66 | 13.7% | |
| Dissolved | Field | 25.20 | 14.8% | 2.11 | 1.2% | 0.62 | 4.5% | |
| Dissolved | Treatment | 17.35 | 11.2% | 2.04 | 1.3% | 0.59 | 4.4% | |
| HS3 | Particulate | Field | 7.07 | 29.3% | 10.32 | 42.8% | 2.85 | 79.6% |
| Particulate | Treatment | 1.80 | 6.7% | 3.41 | 12.7% | 0.93 | 17.6% | |
| Dissolved | Field | 9.19 | 6.5% | 1.67 | 1.2% | 0.46 | 4.0% | |
| Dissolved | Treatment | 5.21 | 3.9% | 1.24 | 0.9% | 0.35 | 3.3% |
| LS1 | LS2 | LS3 | HS2 | HS3 | |
|---|---|---|---|---|---|
| Fucose | 0.84 ± 0.04 | 0.77 ± 0.05 | 1.14 ± 0.22 | 0.40 ± 0.12 | 0.35 ± 0.08 |
| Rhamnose | 0.75 ± 0.02 | 0.83 ± 0.13 | 1.18 ± 0.19 | 0.55 ± 0.11 | 0.38 ± 0.06 |
| Arabinose | 0.79 ± 0.02 | 0.91 ± 0.10 | 1.06 ± 0.13 | 0.58 ± 0.19 | 0.26 ± 0.11 |
| Galactose | 0.78 ± 0.09 | 0.61 ± 0.14 | 1.42 ± 0.20 | 0.43 ± 0.06 | 0.25 ± 0.03 |
| Glucose | 0.32 ± 0.01 | 0.21 ± 0.01 | 0.75 ± 0.16 | 0.46 ± 0.16 | 0.25 ± 0.11 |
| Mannose | n.a. | n.a. | n.a. | n.a. | n.a. |
| Xylose | 1.32 ± 0.05 | 1.21 ± 0.22 | 1.28 ± 0.16 | 0.78 ± 0.22 | 0.23 ± 0.07 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, T.-Y.; Skoog, A. Abiotic Aggregation of Organic Matter in Coastal and Estuarine Waters: Cases in the Eastern Long Island Sound, USA. Water 2021, 13, 3077. https://doi.org/10.3390/w13213077
Chen T-Y, Skoog A. Abiotic Aggregation of Organic Matter in Coastal and Estuarine Waters: Cases in the Eastern Long Island Sound, USA. Water. 2021; 13(21):3077. https://doi.org/10.3390/w13213077
Chicago/Turabian StyleChen, Tzong-Yueh, and Annelie Skoog. 2021. "Abiotic Aggregation of Organic Matter in Coastal and Estuarine Waters: Cases in the Eastern Long Island Sound, USA" Water 13, no. 21: 3077. https://doi.org/10.3390/w13213077
APA StyleChen, T.-Y., & Skoog, A. (2021). Abiotic Aggregation of Organic Matter in Coastal and Estuarine Waters: Cases in the Eastern Long Island Sound, USA. Water, 13(21), 3077. https://doi.org/10.3390/w13213077