Physiological, Developmental, and Biomarker Responses of Zebrafish Embryos to Sub-Lethal Exposure of Bendiocarb


Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Fish Husbandry
2.3. Zebrafish Embryo Toxicity Tests
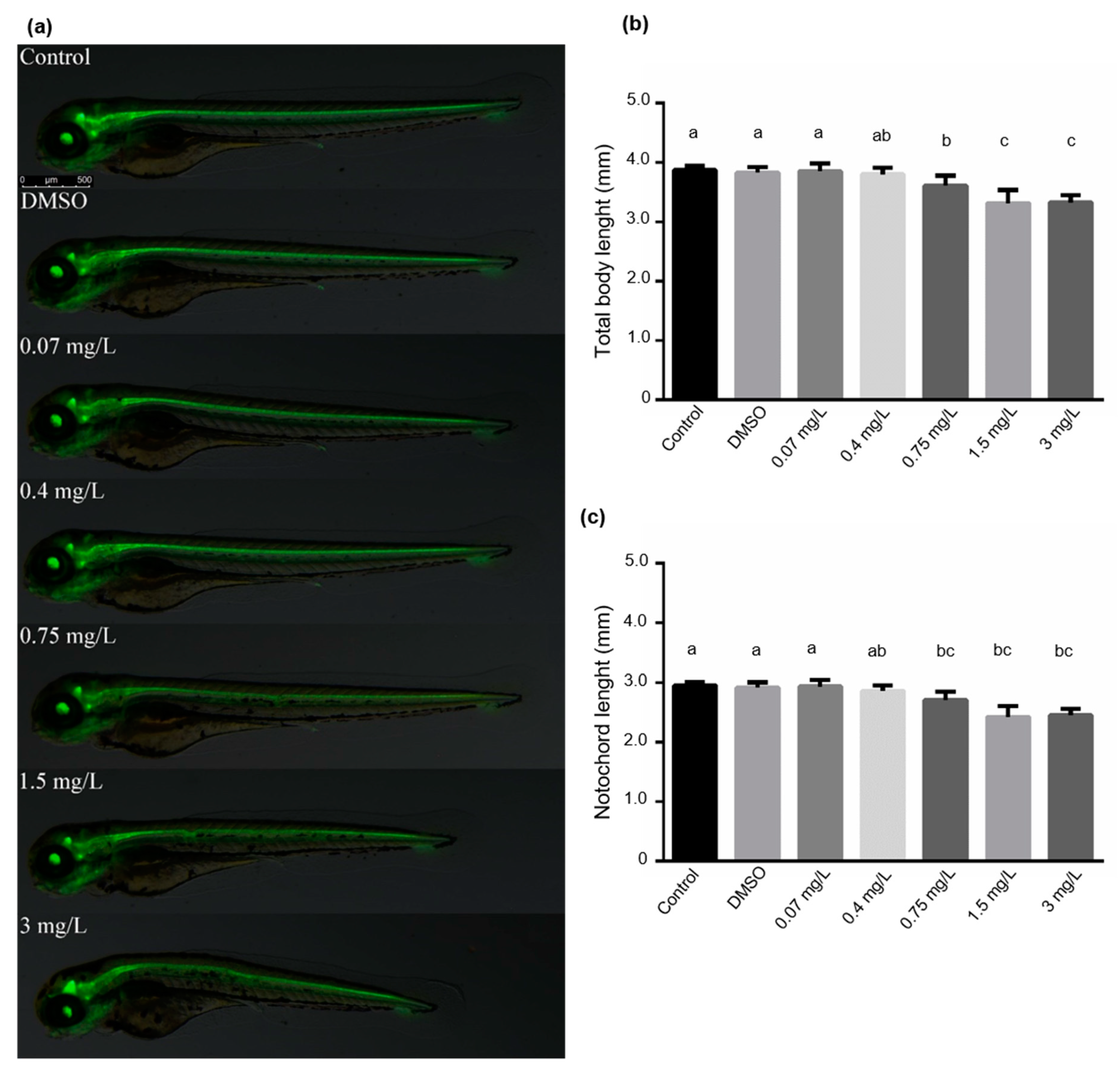
2.4. Heart Rate, Total Body and Notochord Length Determinations
2.5. Tissue Preparation and Biochemical Marker Measurements
2.5.1. Lipid Peroxidation Measurement
2.5.2. AChE Activity Measurement
2.5.3. CAT Activity Measurement
2.5.4. GST Activity Measurement
2.5.5. Glutathione Peroxide Activity Measurement
2.5.6. SOD Activity Measurement
2.6. Statistical Analyses
3. Results
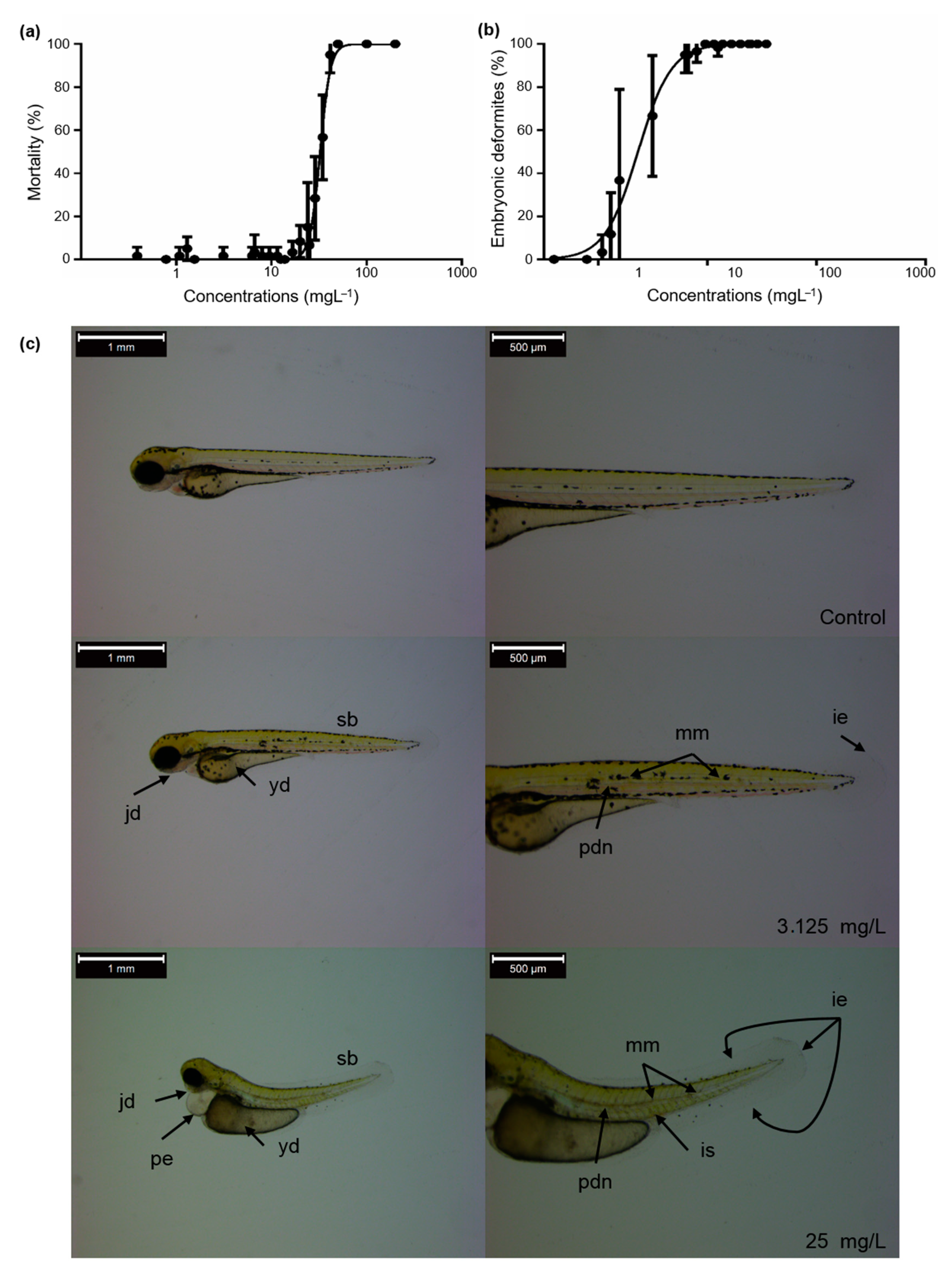
3.1. Zebrafish Embryotoxicity Induced by Bendiocarb Exposure
3.2. Acetylcholinesterase Activity and Heart Rate Assessments
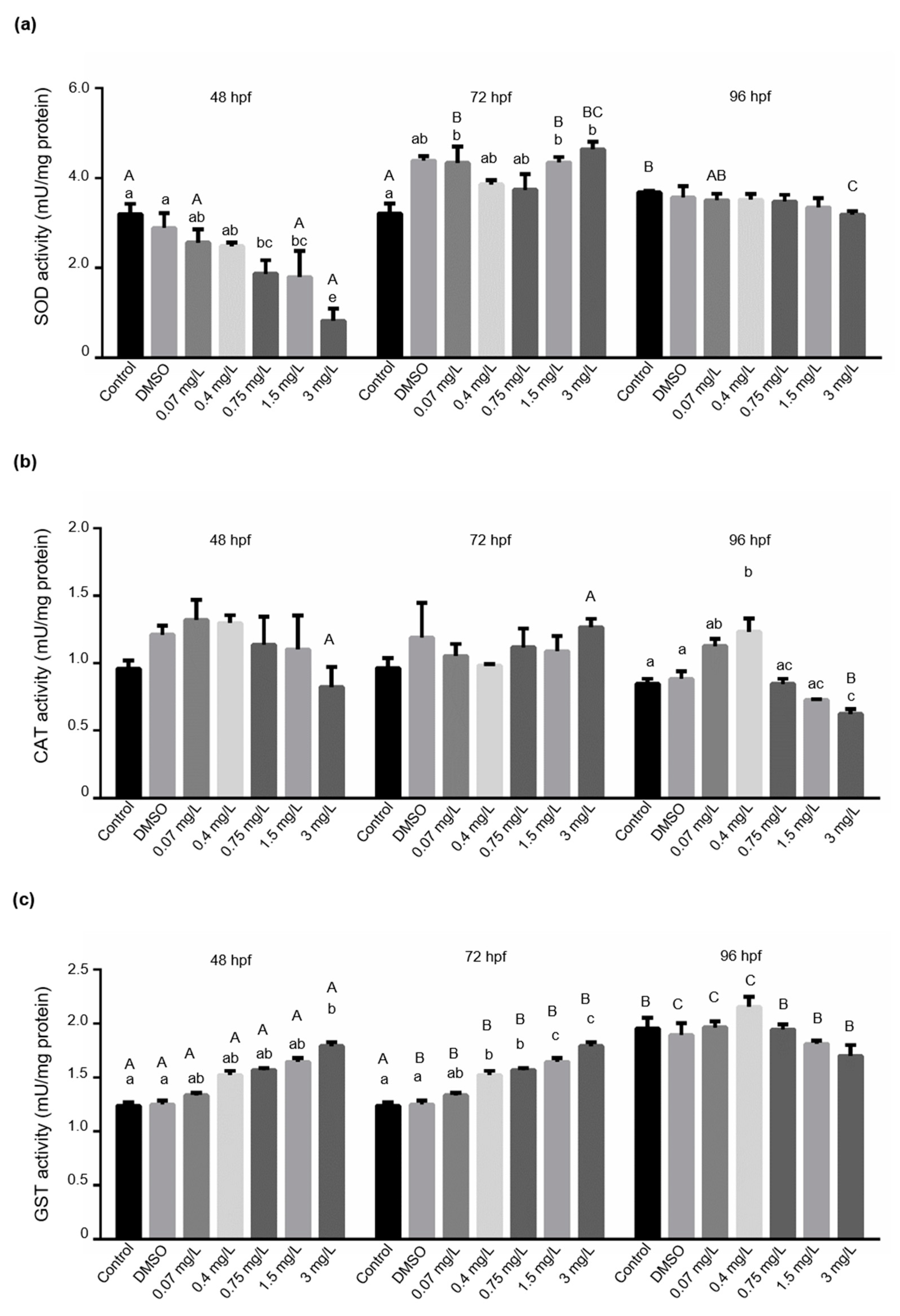
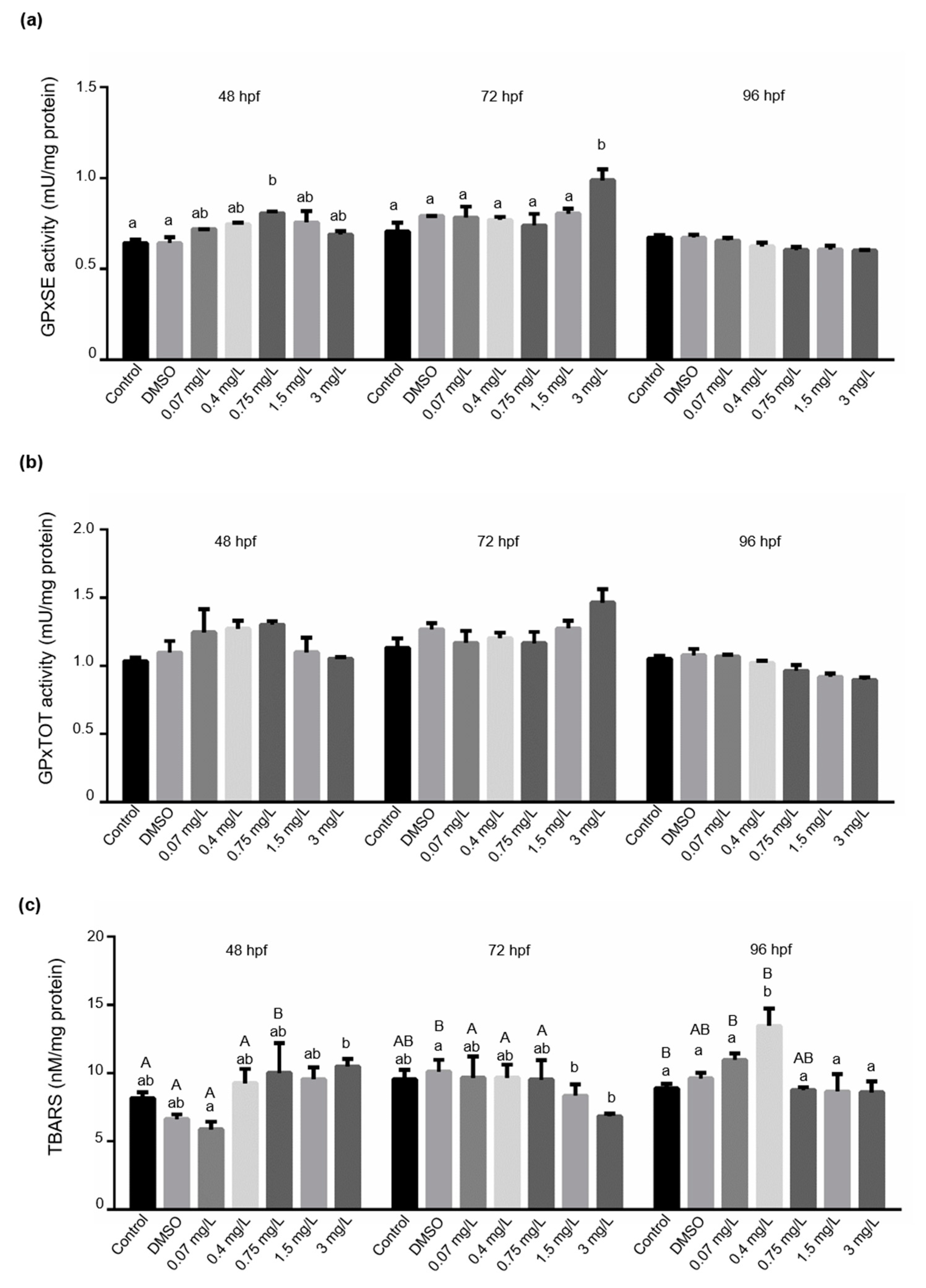
3.3. Determination of Biochemical Markers in Larval Fish
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sogorb, A.M.; Vilanova, E. Enzymes involved in the detoxification of organophosphorus, carbamate and pyrethroid insecticides through hydrolysis. Toxicol. Lett. 2002, 128, 215–228. [Google Scholar] [CrossRef]
- Krockova, J.; Massányi, P.; Toman, R.; Danko, J.; Roychoudhury, S. In vivo and in vitro effect of bendiocarb on rabbit testicular structure and spermatozoa motility. J. Environ. Sci. Health A 2012, 47, 1301–1311. [Google Scholar] [CrossRef] [PubMed]
- Leong, K.H.; Benjamin Tan, L.L.; Mustafa, A.M. Contamination levels of selected organochlorine and organophosphate pesticides in the Selangor River, Malaysia between 2002 and 2003. Chemosphere 2006, 66, 1153–1159. [Google Scholar] [CrossRef] [PubMed]
- Hallenbeck, W.H.; Cunningham-Burns, K.M. Pesticides and Human Health; Springer: New York, NY, USA, 1985. [Google Scholar]
- Holovska, K.; Almasiova, V.; Cigankova, V. Ultrastructural changes in the rabbit liver induced by carbamate insecticide bendiocarb. J. Environ. Sci. Health B 2014, 49, 616–623. [Google Scholar] [CrossRef]
- Polláková, J.; Kovalkovičová, N.; Csank, T.; Pistl, J.; Kočišová, A.; Legáth, J. Evaluation of bendiocarb cytotoxicity in mammalian and insect cell cultures. J. Environ. Sci. Health. B 2012, 47, 538–543. [Google Scholar] [CrossRef]
- Sadasivaiah, S.; Tozan, Y.; Breman, J.G. Dichlorodiphenyltrichloroethane (DDT) for indoor residual spraying in Africa: How can it be used formalaria control? Am. J. Trop. Med. Hyg. 2007, 77, 249–263. [Google Scholar] [CrossRef]
- Hayes, W.J.; Laws, E.R. Classes of pesticides. In Handbook of Pesticide Toxicology; Academic Press: New York, NY, USA, 1990; Volume 3, p. 1576. [Google Scholar]
- Chambers, H.W.; Boone, J.S.; Carr, R.L.; Chanbers, J.E. Chemistry of organophosphorus insecticides. In Handbook of Pesticide Toxicology, 2nd ed.; Robert, I.K., Ed.; Academic Press: San Diego, CA, USA, 2001; pp. 913–917. [Google Scholar]
- Berman, T.; Hochner-Celnikier, D.; Barr, D.B.; Needham, L.L.; Amitai, Y.; Wormser, U.; Richter, E. Pesticide exposure among pregnant women in Jerusalem, Israel: Results of a pilot study. Environ. Int. 2011, 37, 198–203. [Google Scholar] [CrossRef]
- Capcarova, M.; Petrovova, E.; Flesarova, S.; Dankova, M.; Massanyi, P.; Danko, J. Bendiocarbamate induced alterations in selected parameters of rabbithomeostasis after experimental peroral administration. Pestic. Biochem. Physiol. 2010, 98, 213–218. [Google Scholar] [CrossRef]
- Whyatt, R.M.; Barr, D.B.; Camann, D.E.; Kinney, P.L.; Barr, J.R.; Andrews, H.F.; Hoepner, L.A.; Garfinkel, R.; Hazi, Y.; Reyes, A.; et al. Contemporary-use pesticides in personal air samples duringpregnancy and blood samples at delivery among urban minority mothers andnewborns. Environ. Health Perspect. 2003, 111, 749–756. [Google Scholar] [CrossRef]
- Almasiova, V.; Holovska, K.; Tarabova, L.; Cigankova, V.; Lukacinova, A.; Nistiar, F. Structural and ultrastructural study of rabbit testes exposed tocarbamate insecticide. J. Environ. Sci. Health Part B 2012, 47, 1319–1328. [Google Scholar] [CrossRef]
- Apaydin, F.G.; Pandir, D.; Kalender, S.; Baş, H.; Kalender, Y. Hematoprotective effect of vitamins C and E against subchronic toxicity of bendiocarb: Biochemical evidences. J. Food Biochem. 2018, 126–159. [Google Scholar]
- Petrovova, E.; Massanyi, P.; Capcarova, M.; Zivcak, J.; Stodola, L. Structural alterations in rabbit spleen after bendiocarb administration. J. Environ. Sci. Health B 2011, 46, 788–792. [Google Scholar] [PubMed]
- Petrovova, E.; Purzyc, H.; Mazensky, D.; Luptakova, L.; Torma, N.; Sopoliga, I.; Sedmera, D. Morphometric alterations, steatosis, fibrosis and activecaspase-3 detection in carbamate bendiocarb treated rabbit liver. Environ. Toxicol. 2015, 30, 212–222. [Google Scholar] [CrossRef] [PubMed]
- Flesarova, S.; Lukac, N.; Danko, J.; Massanyi, P. Bendiocarbamate inducedstructural alterations in rabbit thymus after experimental peroraladministration. J. Environ. Sci. Health Part B 2007, 42, 329–334. [Google Scholar] [CrossRef]
- Baron, R.L. Carbamate insecticides. In Handbook of Pesticide Toxicology; Hayes, W.J., Laws, E.R., Eds.; Academic Press: New York, NY, USA, 1991; pp. 3–6. ISBN 978-0123341600. [Google Scholar]
- Lazartigues, E.; Freslon, J.L.; Tellioglu, T. Pressor and bradycardic effects of tacrine and other acetylcholinesterase inhibitors in the rat. Eur. J. Pharmacol. 1998, 361, 61–71. [Google Scholar] [CrossRef]
- Masuda, Y.; Kawamura, A. Acetylcholinesterase inhibitor (donepezil hydrochloride) reduces heart rate variability. J. Cardiovasc. Pharmacol. 2003, 41 (Suppl. 1), S67–S71. [Google Scholar]
- Sobeková, A.; Holovská, K.; Lenártová, V.; Flešárová, S.; Javorský, P. The another toxic effect of carbamate insecticides. Acta Biol. Hung. 2009, 60, 45–54. [Google Scholar] [CrossRef]
- Lahr, J.; Diallo, A.O.; Gadji, B.; Diouf, P.S.; Bedaux, J.J.M.; Badji, A.; Ndour, K.B.; Andreasen, J.E.; Straalen, N.M.V. Ecological effects of experimental insecticide applications on invertebrates in sahelian temporary ponds. Environ. Toxicol. Chem. 2000, 19, 1278–1289. [Google Scholar] [CrossRef]
- Bendiocarb (Technical Fact Sheet) NPIC Technical Fact Sheets. 2002. Available online: http://npic.orst.edu/factsheets/archive/bendiotech.pdf (accessed on 30 December 2020).
- Tatarazako, N.; Iguchi, T. Evaluation of toxicities of herbicides using short-term chronic tests of alga, daphnid and fish, herbicides. In Herbicides—Environmental Impact Studies and Management Approaches; Alvarez-Fernandez, R., Ed.; InTech: Rijeka, Croatia, 2012; pp. 135–166. [Google Scholar]
- Scott, G.I.; Fulton, M.H.; Moore, D.W. Agricultural Insecticide Runoff Effects on Estuarine Organisms: Correlating Laboratory and Field Toxicity Testing with Ecotoxicological Biomonitoring; Report #CR813138-01-1; U.S. National Marine Fisheries Service, Southeast Fisheries Science Center, Charleston Laboratory: Charleston, SC, USA, 1990. [Google Scholar]
- Teles, M.; Pacheco, M.; Santos, M.A. Endocrine and metabolic responses of Anguilla anguilla L. caged in a freshwater-wetland (Pateira de Fermentelos-Portugal). Sci. Total Environ. 2007, 372, 562–570. [Google Scholar] [CrossRef]
- El-Saeid, M.H.; Al-Turki, A.M.; Al-Wable, M.I.; Abdel-Nasser, G. Evaluation of pesticide residues in saudi arabia ground water. Res. J. Environ. Sci. 2011, 5, 171–178. [Google Scholar]
- Spalding, R.F.; Exner, M.E.; Snow, D.D.; Cassada, D.A.; Burbach, M.E.; Monson, S.J. Herbicides in ground water beneath Nebraska’s management systems evaluation area. J. Environ. Qual. 2003, 32, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Leu, C.; Singer, H.; Müller, S.R.; Schwarzenbach, R.P.; Stamm, C. Comparison of atrazine losses in three small headwater catchments. J. Environ. Qual. 2005, 34, 1873–1882. [Google Scholar] [CrossRef] [PubMed]
- Senger, M.R.; Rico, E.P.; de Bem Arizi, M.; Rosemberg, D.B.; Dias, R.D.; Bogo, M.R. Carbofuran and malathion inhibit nucleotide hydrolysis in zebrafish (Danio rerio) brain membranes. Toxicology 2005, 212, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Hadzhiev, Y.; Lele, Z.; Schindler, S.; Wilson, S.W.; Ahlberg, P.; Strähle, U.; Müller, F. Hedgehog signaling patterns the outgrowth of unpaired skeletal appendages in zebrafish. BMC Dev. Biol. 2007, 27, 7–75. [Google Scholar] [CrossRef] [PubMed]
- Müller, F.; Chang, B.; Albert, S.; Fischer, N.; Tora, L.; Strahle, U. Intronic enhancers control expression of zebrafish sonic hedgehog in floor plate and notochord. Development 1999, 126, 2103–2116. [Google Scholar]
- Shkumatava, A.; Fischer, S.; Muller, F.; Straehle, U.; Neumann, C.J. Sonic Hedgehog, secreted by amacrine cells, acts as a short range signal to direct differentiation and lamination in the zebrafish retina. Development 2004, 131, 3849–3858. [Google Scholar] [CrossRef]
- OECD. OECD Guidelines for the Testing of Chemicals, Section 2; Test No. 236: Fish Embryo Acute Toxicity (FET) Test; OECD Publishing: Paris, France, 2013. [Google Scholar]
- Rasband, W.S.; ImageJ, U.S. National Institutes of Health, Bethesda, Maryland, USA, 1997–2018. Available online: https://imagej.nih.gov/ij/ (accessed on 30 December 2020).
- Bradford, M. A rapid and sensitive assay of protein utilizing the principle of dye binding. Anal. Biochem. 1976, 772, 242–264. [Google Scholar]
- Wills, E.D. Evaluation of lipid peroxidation in lipids and biological membranes. In Biochemical Toxicology: A Practical Approach; Snell, K., Mullock, B., Eds.; IRL Press: Washington, DC, USA, 1987; pp. 127–152. [Google Scholar]
- Ellman, G.L.; Courtney, K.D.; Andres, V.; Featherstone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Guilhermino, L.; Lopes, M.C.; Carvalho, A.P.; Soares, A.M.V.M. Inhibition of acetylcholinesterase activity as effect criterion in acute tests with juvenile Daphnia magna. Chemosphere 1996, 32, 727–738. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in Vitro. Meth. Enzymol. 1984, 105, 121–126. [Google Scholar]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione S-transferases—The first enzymatic step in mercapturic acid formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar] [CrossRef]
- Paglia, D.E.; Valentin, W.N. Studies on quantitative and qualitative characterization of erythrocyte glutathione peroxidase. J. Lab. Clin. Med. 1967, 70, 158–162. [Google Scholar] [PubMed]
- Lawrence, R.A.; Burk, R.F. Glutathione peroxidase activity in selenium deficient rat liver. Biochem. Biophys. Res. Commun. 1976, 71, 952–958. [Google Scholar] [CrossRef]
- Faria, M.; Carrasco, L.; Diez, S.; Riva, M.C.; Bayona, J.M.; Barata, C. Multi-biomarker responses in the freshwater mussel Dreissena polymorpha exposed to polychlorobiphenyls and metals. Comp. Biochem. Phys. C 2009, 149, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Crapo, J.D.; McCord, J.M.; Fridovich, I. Preparation and assay of superoxide dismutases. Methods Enzymol. 1978, 53, 382–393. [Google Scholar] [PubMed]
- Visser, J.T.; Linders, J. Bendiocarb. Milieu-Fiche; Advisory Report 90/670101/007; College voor de Toelating van Bestrijdingsmiddelen: Wageningen, The Netherlands, 1999. [Google Scholar]
- Fischer, A.; Wolman, M.; Granato, M.; Parsons, M.; McCallion, A.-S.; Proescher, J.; English, E. Carbamate nerve agent prophylatics exhibit distinct toxicological effects in the zebrafish embryo model. Neurotoxicol. Teratol. 2015, 50, 1–10. [Google Scholar] [CrossRef]
- Qian, L.; Qi, S.; Cao, F.; Zhang, J.; Li, C.; Song, M.; Wang, C. Effects of penthiopyrad on the development and behaviour of zebrafish in early-life stages. Chemosphere 2019, 214, 184–194. [Google Scholar] [CrossRef]
- Kashiwada, S.; Tatsuta, H.; Kameshiro, M.; Sugaya, Y.; Sabo-Attwood, T.; Chandler, G.T.; Ferguson, P.L.; Goka, K. Stage-dependent differences in effects of carbaryl on population growth rate in japanese medaka (Oryzias latipes). Environ. Toxicol. Chem. 2008, 27, 2397–2402. [Google Scholar] [CrossRef]
- Mensah, P.K.; Okuthe, G.H.; Onani, M. Sublethal effects of carbaryl on embryonic and gonadal developments of zebrafish Danio rerio. Afr. J. Aquat. Sci. 2012, 37, 271–275. [Google Scholar] [CrossRef]
- Meng, S.L.; Chen, J.Z.; Hu, G.D.; Song, C.; Fan, L.M.; Qiu, L.P.; Xu, P. Effects of chronic exposure of methomyl on the antioxidant system in liver of Nile tilapia (Oreochromis niloticus). Ecotoxicol. Environ. Saf. 2014, 101, 1–6. [Google Scholar] [CrossRef]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of embryonic development of the zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef] [PubMed]
- Barrionuevo, W.R.; Burggren, W.W. O2 consumption and heart rate in developing zebrafish (Danio rerio): Influence of temperature and ambient O2. Am. J. Physiol. 1999, 276, R505–R513. [Google Scholar] [CrossRef] [PubMed]
- Watson, F.L.; Schmidt, H.; Turman, Z.K.; Hole, N.; Garcia, H.; Gregg, J.; Tilghman, J.; Fradinger, E.A. Organophosphate pesticides induce morphological abnormalities and decrease locomotor activity and heart rate in Danio rerio and Xenopus laevis. Environ. Toxicol. Chem. 2014, 33, 1337–1345. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.C.; Hui, M.N.; Cheng, S.H. Toxicity and cardiac effects of carbaryl in early developing zebrafish (Danio rerio) embryos. Toxicol. Appl. Pharmacol. 2007, 222, 159–168. [Google Scholar] [CrossRef]
- Hsieh, D.J.; Liao, C.F. Zebrafish M2 muscarinic acetylcholine receptor: Cloning, pharmacological characterization, expression patterns and roles in embryonic bradycardia. Br. J. Pharmacol. 2002, 137, 782–792. [Google Scholar] [CrossRef]
- Ecobichon, D.J. Carbamate insecticides. In Handbook of Pesticide Toxicology-Agents, 2nd ed.; Krieger, R., Ed.; Academic Press: San Diego, CA, USA, 2001; pp. 1087–1106. [Google Scholar]
- Küster, E.; Altenburger, R. Suborganismic and organismic effects of aldicarb and its metabolite aldicarb-sulfoxide to the zebrafish embryo (Danio rerio). Chemosphere 2007, 68, 751–760. [Google Scholar] [CrossRef]
- WHO/IPCS. Environmental Health Criteria 155. Biomarkers and Risk Assessment: Concepts and Principles; IPCS/World Health Organization: Geneva, Switzerland, 1993. [Google Scholar]
- Filho, D.W. Fish antioxidant defences—A comparative approach. Braz. J. Med. Biol. Res. 1996, 29, 1735–1742. [Google Scholar]
- Apaydin, F.G.; Bas, H.; Kalender, S.; Kalender, Y. Bendiocarb induced histopathological and biochemical alterations inr at liver and preventive role of vitamins C and E. Environ. Toxicol. Pharmacol. 2017, 49, 148–155. [Google Scholar] [CrossRef]
- Karami-Mohajeri, S.; Abdollahi, M. Toxic influence of organophosphate, carbamate, and organochlorine pesticides on cellular metabolism of lipids, proteins, and carbohydrates: A systematic review. Hum. Exp. Toxicol. 2011, 30, 1119–1140. [Google Scholar] [CrossRef]
- Peterman, E.M.; Sullivan, C.; Goody, M.F.; Rodriguez-Nunez, I.; Yoder, J.A.; Kim, C.H. Neutralization of mitochondrial superoxide by superoxide dismutase 2 promotes bacterial clearance and regulates phagocyte numbers in zebrafish. Infect. Immun. 2015, 83, 430–440. [Google Scholar] [CrossRef]
- Meng, S.L.; Chen, J.Z.; Xu, P.; Qu, J.H.; Fan, L.M.; Song, C.; Qiu, L.P. Hepatic antioxidant enzymes SOD and CAT of Nile tilapia (Oreochromis niloticus) in response to pesticide methomyl and recovery pattern. Bull. Environ. Contam. Toxicol. 2014, 92, 388–392. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, A.; Venturino, A.; Pechén de D’Angelo, A.M. Effects of carbaryl and azinphos methyl on juvenile rainbow trout (Oncorhynchus mykiss) detoxifying enzymes. Pestic. Biochem. Physiol. 2007, 88, 134–142. [Google Scholar] [CrossRef]
- Li, H.; Jiang, H.; Gao, X.; Wang, X.; Qu, W.; Lin, R.; Chen, J. Acute toxicity of the pesticide methomyl on the topmouth gudgeon ((Pseudorasbora parva): Mortality and effects on four biomarkers. Fish. Physiol. Biochem. 2008, 34, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Rendón-von Osten, J.; Ortíz-Arana, A.; Guilhermino, L.; Soares, A.M. In vivo evaluation of three biomarkers in the mosquitofish (Gambusia yucatana) exposed to pesticides. Chemosphere 2005, 58, 627–636. [Google Scholar] [CrossRef]
- Hernández-Moreno, D.; Soler, F.; Míguez, M.P.; Pérez-López, M. Brain acetylcholinesterase, malondialdehyde and reduced glutathione as biomarkers of continuous exposure of tench, Tinca tinca, to carbofuran or deltamethrin. Sci. Total Environ. 2010, 408, 4976–4983. [Google Scholar] [CrossRef]
- Hernández-Moreno, D.; Pérez-López, M.; Soler, F.; Gravato, C.; Guilhermino, L. Effects of carbofuran on the sea bass (Dicentrarchus labrax L.): Study of biomarkers and behaviour alterations. Ecotoxicol. Environ. Saf. 2011, 74, 1905–1912. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 96 h LC/EC Values | ||||
|---|---|---|---|---|
| 90 | 50 | 10 | 1 | |
| Mortality | 42.73 mgL−1 | 32.52 mgL−1 | 24.74 mgL−1 | 18.36 mgL−1 |
| Embryonic deformities | 4.98 mgL−1 | 2.30 mgL−1 | 1.06 mgL−1 | 0.46 mgL−1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gazsi, G.; Czimmerer, Z.; Ivánovics, B.; Berta, I.R.; Urbányi, B.; Csenki-Bakos, Z.; Ács, A. Physiological, Developmental, and Biomarker Responses of Zebrafish Embryos to Sub-Lethal Exposure of Bendiocarb. Water 2021, 13, 204. https://doi.org/10.3390/w13020204
Gazsi G, Czimmerer Z, Ivánovics B, Berta IR, Urbányi B, Csenki-Bakos Z, Ács A. Physiological, Developmental, and Biomarker Responses of Zebrafish Embryos to Sub-Lethal Exposure of Bendiocarb. Water. 2021; 13(2):204. https://doi.org/10.3390/w13020204
Chicago/Turabian StyleGazsi, Gyöngyi, Zsolt Czimmerer, Bence Ivánovics, Izabella Roberta Berta, Béla Urbányi, Zsolt Csenki-Bakos, and András Ács. 2021. "Physiological, Developmental, and Biomarker Responses of Zebrafish Embryos to Sub-Lethal Exposure of Bendiocarb" Water 13, no. 2: 204. https://doi.org/10.3390/w13020204
APA StyleGazsi, G., Czimmerer, Z., Ivánovics, B., Berta, I. R., Urbányi, B., Csenki-Bakos, Z., & Ács, A. (2021). Physiological, Developmental, and Biomarker Responses of Zebrafish Embryos to Sub-Lethal Exposure of Bendiocarb. Water, 13(2), 204. https://doi.org/10.3390/w13020204