
Riparian Land Cover, Water Temperature Variability, and Thermal Stress for Aquatic Species in Urban Streams



Abstract
1. Introduction
2. Materials and Methods
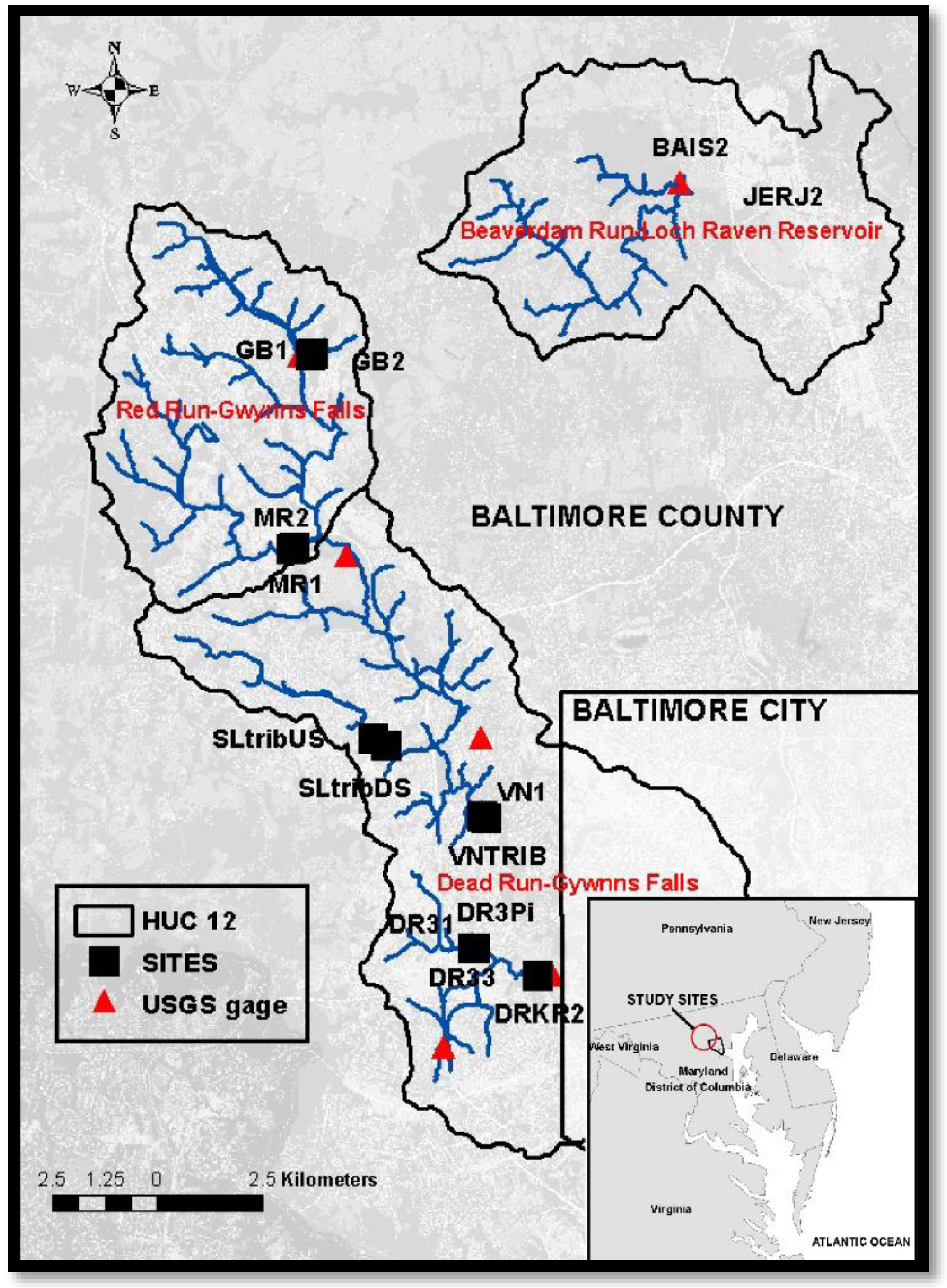
2.1. Study Design
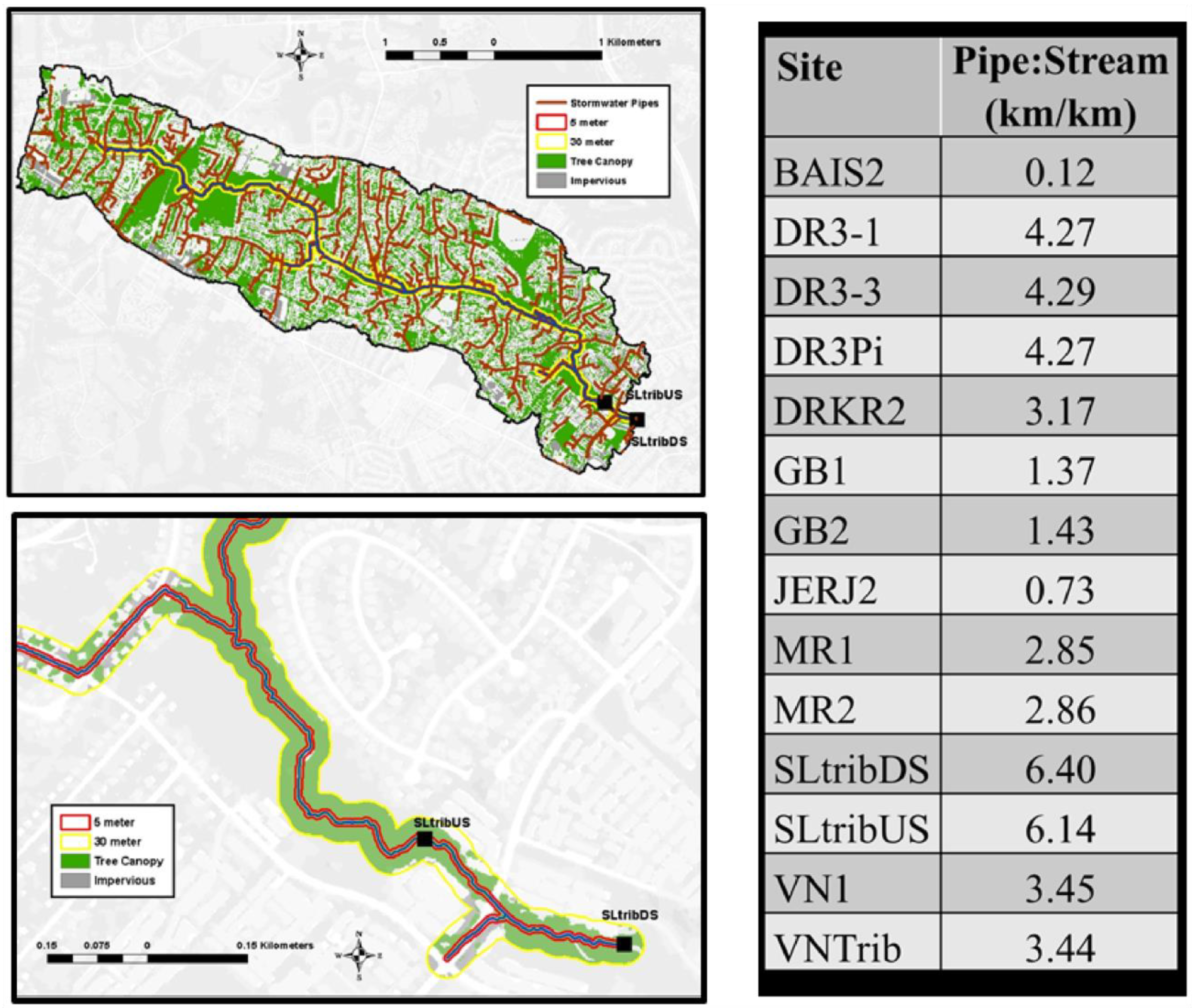
2.2. Loggershed and GIS Data
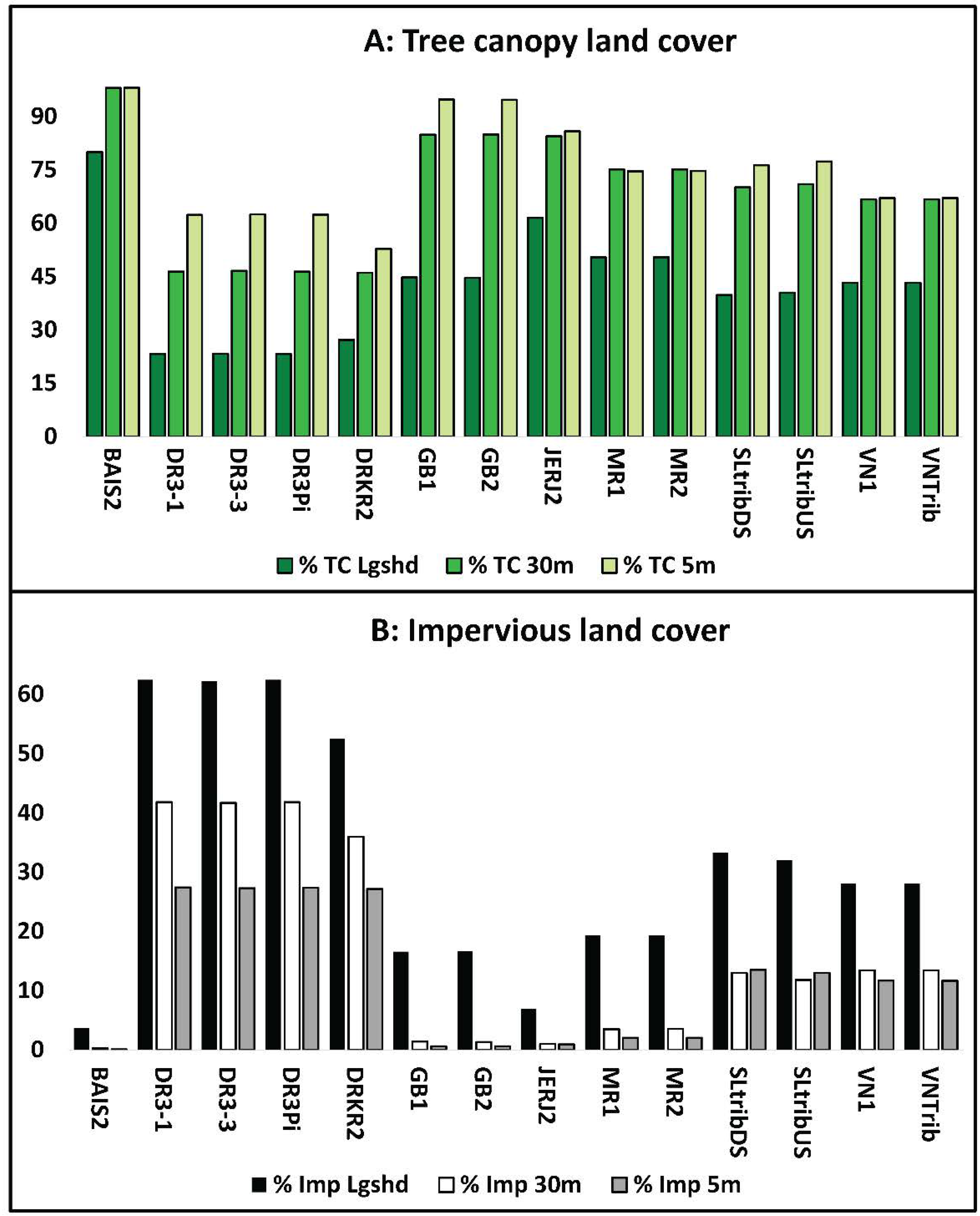
2.2.1. Land Cover Data
2.2.2. Hydrology Data
2.3. Water Temperature Data
2.3.1. Logger Data
2.3.2. Water Temperature Metrics
2.4. Data Analysis
2.4.1. Correlation Analysis
2.4.2. Mixed-Effects Models
2.4.3. Candidate Model Approach
3. Results
3.1. Site Variables and Correlations
3.1.1. Site Characteristics
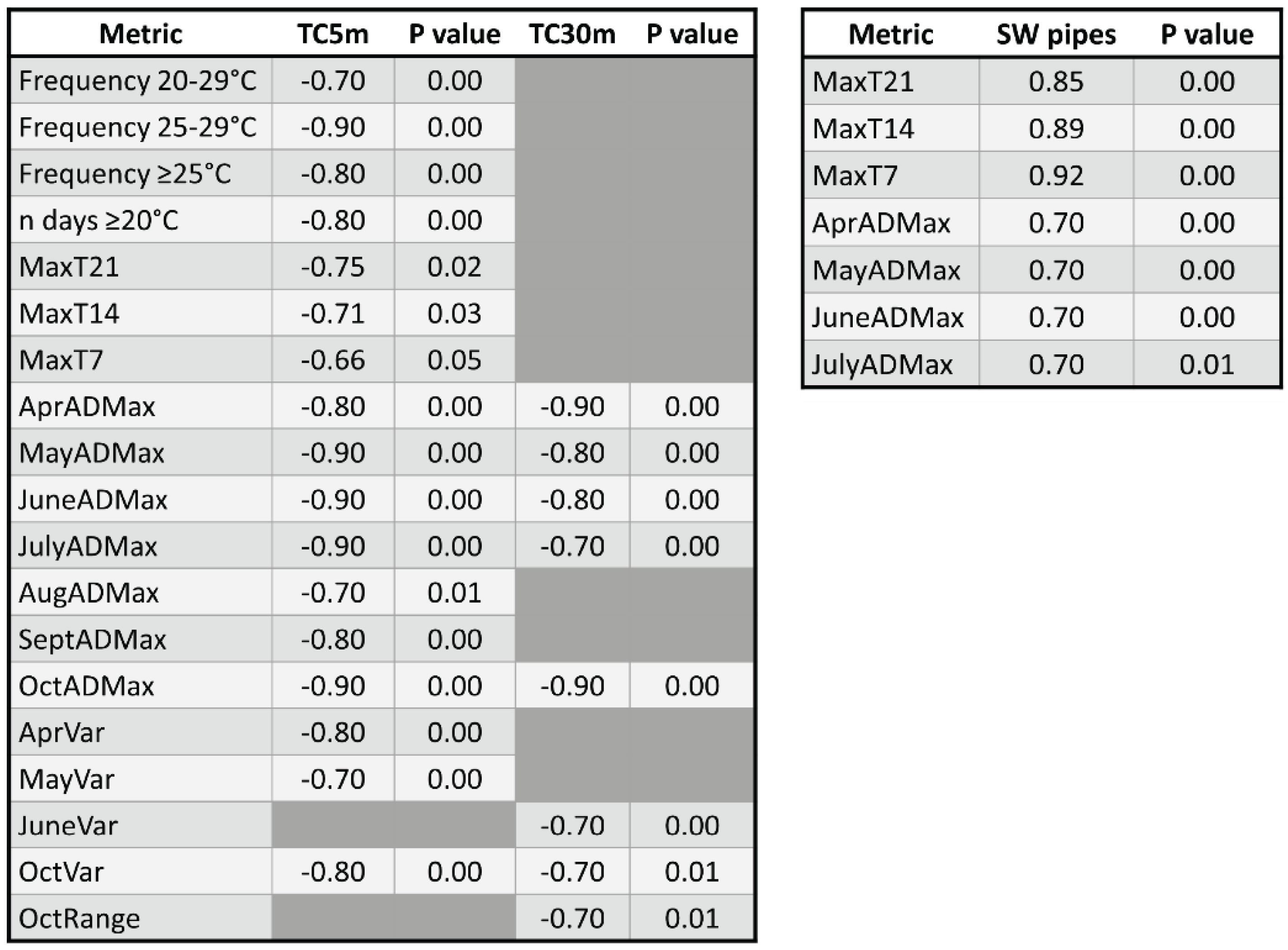
3.1.2. Overall Variable Correlations
3.2. Frequency and Duration of Exceedance
3.2.1. Metrics
3.2.2. Significant Correlations and Best Fit Mixed-Effects Models
3.2.3. Mixed-Effects Model Interpretation
3.3. Annual Magnitude of Change
3.3.1. Metrics
3.3.2. Significant Correlations and Best Fit Mixed-Effects Models
3.3.3. Mixed-Effects Model Interpretation
3.4. Monthly Magnitude of Change
3.4.1. Metrics
3.4.2. Significant Correlations and Best Fit Mixed-Effects Models
3.4.3. Mixed-Effects Model Interpretation
3.5. Monthly Variability
3.5.1. Metrics
3.5.2. Significant Correlations and Best Fit Mixed-Effects Models
3.5.3. Mixed-Effects Model Interpretation
4. Discussion
Urbanization and Thermal Degradation Mitigation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Caissie, D. The thermal regime of rivers: A review. Freshw. Biol. 2006, 51, 1389–1406. [Google Scholar] [CrossRef]
- Ouellet, V.; Gibson, E.E.; Daniels, M.D.; Watson, N.A. Riparian and geomorphic controls on thermal habitat dynamics of pools in a temperate headwater stream. Ecohydrology 2017, 10, 1–10. [Google Scholar] [CrossRef]
- Comte, L.; Olden, J.D.; Tedesco, P.A.; Ruhi, A.; Giam, X. Climate and land-use changes interact to drive long-term reorganization of riverine fish communities globally. Proc. Natl. Acad. Sci. USA 2021, 118, e2011639118. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.E.; Isaak, D.J.; Imhof, J.; Hendrickson, D.A.; McMillan, J.R. Cold-Water Fishes and Climate Change in North America. Ref. Modul. Earth Syst. Environ. Sci. 2015, 1–10. [Google Scholar] [CrossRef]
- Ouellet, V.; St-Hilaire, A.; Dugdale, S.J.; Hannah, D.M.; Krause, S.; Proulx-Ouellet, S. River temperature research and practice: Recent challenges and emerging opportunities for managing thermal habitat conditions in stream ecosystems. Sci. Total Environ. 2020, 736, 139679. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, J.B.; Fullerton, A.H.; Jordan, C.E.; Ebersole, J.L.; Bellmore, J.R.; Arismendi, I.; Penaluna, B.E.; Reeves, G.H. The importance of warm habitat to the growth regime of cold-water fishes. Nat. Clim. Chang. 2021, 11, 354–361. [Google Scholar] [CrossRef]
- Poole, G.C.; Dunham, J.B.; Keenan, D.M.; Sauter, S.T.; McCullough, D.A.; Mebane, C.; Lockwood, J.C.; Essig, D.A.; Hicks, M.P.; Sturdevant, D.J.; et al. The Case for Regime-based Water Quality Standards. Bioscience 2004, 54, 155. [Google Scholar] [CrossRef]
- Hirsch, R.M.; De Cicco, L. User guide to Exploration and Graphics for RivEr Trends (EGRET) and dataRetrieval: R packages for hydrologic data. Tech. Methods B. 2015, 4, 93. [Google Scholar] [CrossRef]
- Kingsolver, J.G.; Woods, H.A. Beyond thermal performance curves: Modeling time-dependent effects of thermal stress on ectotherm growth rates. Am. Nat. 2016, 187, 283–294. [Google Scholar] [CrossRef]
- Whitney, J.E.; Al-Chokhachy, R.; Bunnell, D.B.; Caldwell, C.A.; Cooke, S.J.; Eliason, E.J.; Rogers, M.; Lynch, A.J.; Paukert, C.P. Bases physiologiques des impacts des changements climatiques sur les poissons continentaux d’Amérique du Nord. Fisheries 2016, 41, 332–345. [Google Scholar] [CrossRef]
- Culumber, Z.W.; Monks, S. Resilience to extreme temperature events: Acclimation capacity and body condition of a polymorphic fish in response to thermal stress. Biol. J. Linn. Soc. 2014, 111, 504–510. [Google Scholar] [CrossRef]
- Durance, I.; Ormerod, S.J. Trends in water quality and discharge confound long-term warming effects on river macroinvertebrates. Freshw. Biol. 2009, 54, 388–405. [Google Scholar] [CrossRef]
- Woods, T.; Kaz, A.; Giam, X. Phenology in freshwaters: A review and recommendations for future research. Ecography 2021, 1–14. [Google Scholar] [CrossRef]
- Crozier, L.G.; Scheuerell, M.D.; Zabel, R.W. Using time series analysis to characterize evolutionary and plastic responses to environmental change: A case study of a shift toward earlier migration date in sockeye salmon. Am. Nat. 2011, 178, 755–773. [Google Scholar] [CrossRef]
- Keefer, M.L.; Clabough, T.S.; Jepson, M.A.; Johnson, E.L.; Peery, C.A.; Caudill, C.C. Thermal exposure of adult Chinook salmon and steelhead: Diverse behavioral strategies in a large and warming river system. PLoS ONE 2018, 13, e0204274. [Google Scholar] [CrossRef] [PubMed]
- Kaylor, M.J.; Justice, C.; Armstrong, J.B.; Staton, B.A.; Burns, L.A.; Sedell, E.; White, S.M. Temperature, emergence phenology and consumption drive seasonal shifts in fish growth and production across riverscapes. J. Anim. Ecol. 2021, 90, 1727–1741. [Google Scholar] [CrossRef] [PubMed]
- Campbell, E.Y.; Dunham, J.B.; Reeves, G.H.; Wondzell, S.M. Phenology of hatching, emergence, and end-of-season body size in young-of-year coho salmon in thermally contrasting streams draining the Copper River Delta, Alaska. Can. J. Fish. Aquat. Sci. 2019, 76, 185–191. [Google Scholar] [CrossRef]
- Flitcroft, R.L.; Lewis, S.L.; Arismendi, I.; LovellFord, R.; Santelmann, M.V.; Safeeq, M.; Grant, G. Linking hydroclimate to fish phenology and habitat use with ichthyographs. PLoS ONE 2016, 11, e0168831. [Google Scholar] [CrossRef]
- Fuhrman, A.E.; Larsen, D.A.; Steel, E.A.; Young, G.; Beckman, B.R. Chinook salmon emergence phenotypes: Describing the relationships between temperature, emergence timing and condition factor in a reaction norm framework. Ecol. Freshw. Fish 2018, 27, 350–362. [Google Scholar] [CrossRef]
- Hebert, C.; Caissie, D.; Satish, M.G.; El-Jabi, N. Modeling of hourly river water temperatures using artificial neural networks. Water Qual. Res. J. Can. 2014, 49, 144–162. [Google Scholar] [CrossRef][Green Version]
- Hannah, D.M.; Garner, G. River water temperature in the United Kingdom: Changes over the 20th century and possible changes over the 21st century. Prog. Phys. Geogr. 2015, 39, 68–92. [Google Scholar] [CrossRef]
- Arismendi, I.; Johnson, S.L.; Dunham, J.B.; Haggerty, R. Descriptors of natural thermal regimes in streams and their responsiveness to change in the Pacific Northwest of North America. Freshw. Biol. 2013, 58, 880–894. [Google Scholar] [CrossRef]
- Arismendi, I.; Johnson, S.L.; Dunham, J.B.; Haggerty, R.; Hockman-Wert, D. The paradox of cooling streams in a warming world: Regional climate trends do not parallel variable local trends in stream temperature in the Pacific continental United States. Geophys. Res. Lett. 2012, 39, 1–7. [Google Scholar] [CrossRef]
- Steel, E.A.; Beechie, T.J.; Torgersen, C.E.; Fullerton, A.H. Envisioning, Quantifying, and Managing Thermal Regimes on River Networks. Bioscience 2017, 67, 506–522. [Google Scholar] [CrossRef]
- Dugdale, S.J.; Malcolm, I.A.; Kantola, K.; Hannah, D.M. Stream temperature under contrasting riparian forest cover: Understanding thermal dynamics and heat exchange processes. Sci. Total Environ. 2018, 610–611, 1375–1389. [Google Scholar] [CrossRef]
- Broadmeadow, S.B.; Jones, J.G.; Langford, T.E.L.; Shaw, P.J.; Nisbet, T.R. The influence of riparian shade on lowland stream water temperatures in southern England and their viability for brown trout. River Res. Appl. 2011, 27, 226–237. [Google Scholar] [CrossRef]
- Kanno, Y.; Vokoun, J.C.; Letcher, B.H. Paired stream-air temperature measurements reveal fine-scale thermal heterogeneity within headwater brook trout stream networks. River Res. Appl. 2014, 30, 745–755. [Google Scholar] [CrossRef]
- Ebersole, J.L.; Liss, W.J.; Frissell, C.A. Cold water patches in warm streams: Physicochemical characteristics and the influence of shading. J. Am. Water Resour. Assoc. 2003, 39, 355–368. [Google Scholar] [CrossRef]
- Taylor, C.A.; Stefan, H.G. Shallow groundwater temperature response to climate change and urbanization. J. Hydrol. 2009, 375, 601–612. [Google Scholar] [CrossRef]
- Gunawardhana, L.N.; Kazama, S. Statistical and numerical analyses of the influence of climate variability on aquifer water levels and groundwater temperatures: The impacts of climate change on aquifer thermal regimes. Glob. Planet. Chang. 2012, 86–87, 66–78. [Google Scholar] [CrossRef]
- Hathaway, J.M.; Winston, R.J.; Brown, R.A.; Hunt, W.F.; McCarthy, D.T. Temperature dynamics of stormwater runoff in Australia and the USA. Sci. Total Environ. 2016, 559, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Kaushal, S.S.; Likens, G.E.; Jaworski, N.A.; Pace, M.L.; Sides, A.M.; Seekell, D.; Belt, K.T.; Secor, D.H.; Wingate, R.L. Rising stream and river temperatures in the United States. Front. Ecol. Environ. 2010, 8, 461–466. [Google Scholar] [CrossRef]
- Bhaskar, A.S.; Hopkins, K.G.; Smith, B.K.; Stephens, T.A.; Miller, A.J. Hydrologic Signals and Surprises in U.S. Streamflow Records During Urbanization. Water Resour. Res. 2020, 56, 1–22. [Google Scholar] [CrossRef]
- Timm, A.; Ouellet, V.; Daniels, M. Swimming through the urban heat island: Can thermal mitigation practices reduce the stress? River Res. Appl. 2020, 36, 1973–1984. [Google Scholar] [CrossRef]
- Kaushal, S.S.; Belt, K.T. The urban watershed continuum: Evolving spatial and temporal dimensions. Urban Ecosyst. 2012, 15, 409–435. [Google Scholar] [CrossRef]
- Kalnay, E.; Cai, M. Impact of urbanization and land-use. Nature 2003, 425, 102. [Google Scholar] [CrossRef]
- Rose, S.; Peters, N.E. Effects of urbanization on streamflow in the Atlanta area (Georgia, USA): A comparative hydrological approach. Hydrol. Process. 2001, 15, 1441–1457. [Google Scholar] [CrossRef]
- Palmer, M.A.; Nelson, K.C. Stream Temperature Surges Under Urbanization and Climate Change. J. Am. Water Resour. Assoc. 2007, 43, 440–452. [Google Scholar] [CrossRef]
- Moore, R.D.; Spittlehouse, D.L.; Story, A. Riparian microclimate and stream temperature response to forest harvesting: A review. J. Am. Water Resour. Assoc. 2005, 7, 813–834. [Google Scholar] [CrossRef]
- Garner, G.; Malcolm, I.A.; Sadler, J.P.; Hannah, D.M. The role of riparian vegetation density, channel orientation and water velocity in determining river temperature dynamics. J. Hydrol. 2017, 553, 471–485. [Google Scholar] [CrossRef]
- Garner, G.; Malcolm, I.A.; Sadler, J.P.; Millar, C.P.; Hannah, D.M. Inter-annual variability in the effects of riparian woodland on micro-climate, energy exchanges and water temperature of an upland Scottish stream. Hydrol. Process. 2015, 29, 1080–1095. [Google Scholar] [CrossRef]
- Garner, G.; Malcolm, I.A.; Sadler, J.P.; Hannah, D.M. What causes cooling water temperature gradients in a forested stream reach? Hydrol. Earth Syst. Sci. 2014, 18, 5361–5376. [Google Scholar] [CrossRef]
- Johnson, M.F.; Wilby, R.L. Seeing the landscape for the trees: Metrics to guide riparian shade management in river catchments. Water Resour. Res. 2015, 51, 3754–3769. [Google Scholar] [CrossRef]
- Kalny, G.; Laaha, G.; Melcher, A.; Trimmel, H.; Weihs, P.; Rauch, H.P. The influence of riparian vegetation shading on water temperature during low flow conditions in a medium sized river. Knowl. Manag. Aquat. Ecosyst. 2017, 1–14. [Google Scholar] [CrossRef]
- Bowler, D.E.; Mant, R.; Orr, H.; Hannah, D.M.; Pullin, A.S. What are the effects of wooded riparian zones on stream temperature? Environ. Evid. 2012, 1, 1–9. [Google Scholar] [CrossRef]
- Sweeney, B.W.; Newbold, J.D. Streamside forest buffer width needed to protect stream water quality, habitat, and organisms: A literature review. J. Am. Water Resour. Assoc. 2014, 50, 560–584. [Google Scholar] [CrossRef]
- Schueler, T. The importance of imperviousness. Watershed Prot. Tech. 1994, 1, 100–111. [Google Scholar]
- Hession, W.C.; Johnson, T.E.; Charles, D.F.; Hart, D.D.; Horwitz, R.J.; Kreeger, D.A.; Pizzuto, J.E.; Velinski, D.J.; Newbold, J.D.; Cianfrani, C.; et al. Ecological benefits of riparian reforestation in urban watersheds: Study design and preliminary results. Environ. Monit. Assess. 2000, 63, 211–222. [Google Scholar] [CrossRef]
- Roy, A.H.; Faust, C.L.; Freeman, M.C.; Meyer, J.L. Reach-scale effects of riparian forest cover on urban stream ecosystems. Can. J. Fish. Aquat. Sci. 2005, 62, 2312–2329. [Google Scholar] [CrossRef]
- Horwitz, R.J.; Johnson, T.E.; Overbeck, P.F.; O’Donnell, T.K.; Hession, W.C.; Sweeney, B.W. Effects of riparian vegetation and watershed urbanization on fishes in streams of the mid-atlantic piedmont (USA). J. Am. Water Resour. Assoc. 2008, 44, 724–741. [Google Scholar] [CrossRef]
- U.S. Geological Survey, U.S. Department of Agriculture Natural Resources Conservation Service Federal. Standards and Procedures for the National Watershed Boundary Dataset (WBD); Technics and Methods 11–A3; U.S. Geological Survey: Reston, VA, USA, 2013; p. 63.
- Buffington, J.M.; Montgomery, D.R. Geomorphic Classification of Rivers. In Treatise on Geomorphology; Fluvial Geomorphology; Academic Press: San Diego, CA, USA, 2013; Volume 9, ISBN 9780123747396. [Google Scholar]
- Flynn, K.M.; Kirby, W.H.; Hummel, P.R. User’s Manual for Program PeakFQ; Annual Flood-Frequency Analysis Using Bulletin 17B Guidelines; U.S. Geological Survey: Reston, VA, USA, 2006.
- Rutledge, A.T. Computer Programs for Describing the Recession of Ground-Water Discharge and for Estimating Mean Ground-Water Recharge and Discharge from Streamflow Records-Update; Water-Resources Investigations Report 98-4148; U.S. Geological Survey: Reston, VA, USA, 1998; p. 52.
- O’Neil-Dunne, J.; MacFaden, S.; Royar, A. A versatile, production-oriented approach to high-resolution tree-canopy mapping in urban and suburban landscapes using GEOBIA and data fusion. Remote Sens. 2014, 6, 12837–12865. [Google Scholar] [CrossRef]
- O’Neil-Dunne, J.P.M.; MacFaden, S.W.; Royar, A.R.; Pelletier, K.C. An object-based system for LiDAR data fusion and feature extraction. Geocarto Int. 2013, 28, 227–242. [Google Scholar] [CrossRef]
- Chesapeake Conservancy Land Cover Data Project 2013/2014. Available online: https://www.chesapeakeconservancy.org/conservation-innovation-center/high-resolution-data/land-cover-data-project/ (accessed on 1 April 2017).
- Caissie, D.; El-Jabi, N. The importance of cross-calibration and protecting water temperature sensors against direct solar radiation heating in hydrological studies. Hydrol. Sci. J. 2020, 65, 102–111. [Google Scholar] [CrossRef]
- Albertson, L.K.; Ouellet, V.; Daniels, M.D. Impacts of stream riparian buffer land use on water temperature and food availability for fish. J. Freshw. Ecol. 2018, 33, 195–210. [Google Scholar] [CrossRef]
- Crozier, L.G.; Hendry, A.P.; Lawson, P.W.; Quinn, T.P.; Mantua, N.J.; Battin, J.; Shaw, R.G.; Huey, R.B. Potential responses to climate change in organisms with complex life histories: Evolution and plasticity in Pacific salmon. Evol. Appl. 2008, 1, 252–270. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, F.J. The breeding behavior of the southern blacknose dace, Rhinichthys atratulus obtusus Agassiz. Copeia 1958, 2, 141–143. [Google Scholar] [CrossRef]
- Tsang, Y.P.; Infante, D.M.; Stewart, J.; Wang, L.; Tingly, R.W.; Thornbrugh, D.; Cooper, A.R.; Daniel, W.M. StreamThermal: Un progiciel de calcul des valeurs thermiques à partir des données de température des cours d’eau. Fisheries 2016, 41, 548–554. [Google Scholar] [CrossRef]
- Beitinger, T.L.; Bennett, W.A.; McCauley, R.W. Temperature tolerances of North American freshwater fishes exposed to dynamic changes in temperature. Environ. Biol. Fishes 2000, 58, 237–275. [Google Scholar] [CrossRef]
- Westhoff, J.T.; Rosenberger, A.E. A global review of freshwater crayfish temperature tolerance, preference, and optimal growth. Rev. Fish Biol. Fish. 2016, 26, 329–349. [Google Scholar] [CrossRef]
- Lutterschmidt, W.I.; Hutchison, V.H. The critical thermal maximum: History and critique. Can. J. Zool. 1997, 75, 1561–1574. [Google Scholar] [CrossRef]
- Becker, C.D.; Genoway, R.G. Evaluation of the critical thermal maximum for determining thermal tolerance of freshwater fish. Environ. Biol. Fishes 1979, 4, 245–256. [Google Scholar] [CrossRef]
- Chadwick, J.G.; Nislow, K.H.; McCormick, S.D. Thermal onset of cellular and endocrine stress responses correspond to ecological limits in brook trout, an iconic cold-water fish. Conserv. Physiol. 2015, 3, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.M.; Rinne, J.N. Critical Thermal Maxima of Five Trout Species in the Southwestern United States. Trans. Am. Fish. Soc. 1980, 109, 632–635. [Google Scholar] [CrossRef]
- Hartman, K.J.; Porto, M.A. Thermal Performance of Three Rainbow Trout Strains at Above-Optimal Temperatures. Trans. Am. Fish. Soc. 2014, 143, 1445–1454. [Google Scholar] [CrossRef]
- Hart, J.S. Geographic Variations of Some Physiological and Morphological Characters in Certain Freshwater Fish; The University of Toronto Press: Toronto, ON, Canada, 1952. [Google Scholar]
- Wetzel, J.E.; Brown, P.B. Growth and Survival of Juvenile Orconectes virilis and Orconectes immunis at Different Temperatures. J. World Aquac. Soc. 1993, 24, 339–343. [Google Scholar] [CrossRef]
- Whitledge, G.W.; Rabeni, C.F. Maximum daily consumption and respiration rates at four temperatures for five species of crayfish from missouri, U.S.A. (Decapoda, Orconectes spp.). Crustaceana 2002, 75, 1119–1132. [Google Scholar] [CrossRef]
- Heck, M.P.; Schultz, L.D.; Hockman-Wert, D.; Dinger, E.C.; Dunham, J.B. Monitoring Stream Temperatures—A Guide for Non-Specialists; U.S. Geological Survey: Reston, VA, USA, 2018.
- Jones, N.E.; Schmidt, B.J. Thermal regime metrics and quantifying their uncertainty for North American streams. River Res. Appl. 2018, 34, 382–393. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.M.; Walker, S.C. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Burnham, K.; Anderson, D. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 2002; p. 488. [Google Scholar]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014. [Google Scholar]
- Turschwell, M.P.; Balcombe, S.R.; Steel, E.A.; Sheldon, F.; Peterson, E.E. Thermal habitat restricts patterns of occurrence in multiple life-stages of a headwater fish. Freshw. Sci. 2017, 36, 402–414. [Google Scholar] [CrossRef]
- Ketabchy, M.; Sample, D.J.; Wynn-Thompson, T.; Nayeb Yazdi, M. Thermal evaluation of urbanization using a hybrid approach. J. Environ. Manag. 2018, 226, 457–475. [Google Scholar] [CrossRef]
- Ketabchy, M.; Sample, D.J.; Wynn-Thompson, T.; Yazdi, M.N. Simulation of watershed-scale practices for mitigating stream thermal pollution due to urbanization. Sci. Total Environ. 2019, 671, 215–231. [Google Scholar] [CrossRef] [PubMed]
- Raney, E.C. Comparison of the breeding habitats of two subspecies of the blacknosed dace, Rhinichthys atratulus (Herman). Am. Midl. Nat. 1940, 23, 399–403. [Google Scholar] [CrossRef]
- Atkinson, J.B. Shenandoah National Park Fisheries Monitoring Program Annual Report for 2004; U.S. Geological Survey: Luray, VA, USA, 2005.
- Anderson, H.E.; Albertson, L.K.; Walters, D.M. Water temperature drives variability in salmonfly abundance, emergence timing, and body size. River Res. Appl. 2019, 35, 1013–1022. [Google Scholar] [CrossRef]
- Villalobos-Jiménez, G.; Hassall, C. Effects of the urban heat island on the phenology of Odonata in London, UK. Int. J. Biometeorol. 2017, 61, 1337–1346. [Google Scholar] [CrossRef]
- Baldock, J.R.; Armstrong, J.B.; Schindler, D.E.; Carter, J.L. Juvenile coho salmon track a seasonally shifting thermal mosaic across a river floodplain. Freshw. Biol. 2016, 61, 1454–1465. [Google Scholar] [CrossRef]
- Benjamin, J.R.; Heltzel, J.M.; Dunham, J.B.; Heck, M.; Banish, N. Thermal Regimes, Nonnative Trout, and Their Influences on Native Bull Trout in the Upper Klamath River Basin, Oregon. Trans. Am. Fish. Soc. 2016, 145, 1318–1330. [Google Scholar] [CrossRef]
- Laub, B.G.; Baker, D.W.; Bledsoe, B.P.; Palmer, M.A. Range of variability of channel complexity in urban, restored and forested reference streams. Freshw. Biol. 2012, 57, 1076–1095. [Google Scholar] [CrossRef]
- Wenger, S.J.; Peterson, J.T.; Freeman, M.C.; Freeman, B.J.; Homans, D.D. Stream fish occurrence in response to impervious cover, historic land use, and hydrogeomorphic factors. Can. J. Fish. Aquat. Sci. 2008, 65, 1250–1264. [Google Scholar] [CrossRef]
- Alberti, M.; Booth, D.; Hill, K.; Coburn, B.; Avolio, C.; Coe, S.; Spirandelli, D. The impact of urban patterns on aquatic ecosystems: An empirical analysis in Puget lowland sub-basins. Landsc. Urban Plan. 2007, 80, 345–361. [Google Scholar] [CrossRef]
- Roy, A.H.; Freeman, M.C.; Freeman, B.J.; Wenger, S.J.; Ensign, W.E.; Meyer, J.L. Investigating hydrologic alteration as a mechanism of fish assemblage shifts in urbanizing streams. J. N. Am. Benthol. Soc. 2005, 24, 656–678. [Google Scholar] [CrossRef]
- FWPCA. Federal Water Pollution Control Act (Clean Water Act). 33 U.S.C. §§ 1251-1387. 18 October 1972. Available online: http://www.epa.gov/lawsregs/laws/cwa.html (accessed on 1 April 2013).
- Food Security Act, 1985. Farm Bill. US Public Law No. 99-198, 99 Stat. 1354. Available online: https://www.govinfo.gov/content/pkg/STATUTE-99/pdf/STATUTE-99-Pg1354.pdf (accessed on 13 September 2021).
- Welsch, D. Riparian Forest Buffers. Function and Design for Protection and Enhancement of Water Resources; USDA Forest Service, Northeastern Area State & Private Forestry: Radnor, PA, USA, 1991.
- Baruch, E.M.; Voss, K.A.; Blaszczak, J.R.; Delesantro, J.; Urban, D.L.; Bernhardt, E.S. Not all pavements lead to streams: Variation in impervious surface connectivity affects urban stream ecosystems. Freshw. Sci. 2018, 37, 673–684. [Google Scholar] [CrossRef]
- Booth, D. Challenges and prospects for restoring urban streams: A perspective from the Pacific Northwest of North America: BRIDGES is a recurring feature of J-NABS intended. J. N. Am. Benthol. Soc. 2005, 24, 724–737. [Google Scholar] [CrossRef]
- Sabouri, F.; Gharabaghi, B.; Mahboubi, A.A.; McBean, E.A. Impervious surfaces and sewer pipe effects on stormwater runoff temperature. J. Hydrol. 2013, 502, 10–17. [Google Scholar] [CrossRef]
- Lim, T.C.; Welty, C. Effects of spatial configuration of imperviousness and green infrastructure networks on hydrologic response in a residential sewershed. Water Resour. Res. 2017, 53, 8084–8104. [Google Scholar] [CrossRef]
- Wullschleger, S.D.; Meinzer, F.C.; Vertessy, R.A. A review of whole-plant water use studies in tree. Tree Physiol. 1998, 18, 499–512. [Google Scholar] [CrossRef] [PubMed]
- Torgersen, C.E.; Price, D.M.; Li, H.W.; McIntosh, B.A. Multiscale thermal refugia and stream habitat associations of chinook salmon in northeastern Oregon. Ecol. Appl. 1999, 9, 301–319. [Google Scholar] [CrossRef]
- Ebersole, J.; Liss, W.; Frissell, C. Relationship between stream temperature, thermal refugia and rainbow trout Oncorhynchus mykiss abundance in arid-land streams in the northwestern United States. Ecol. Freshw. Fish 2001, 9, 864–879. [Google Scholar] [CrossRef]
- Isaak, D.J.; Young, M.K.; Nagel, D.E.; Horan, D.L.; Groce, M.C. The cold-water climate shield: Delineating refugia for preserving salmonid fishes through the 21st century. Glob. Chang. Biol. 2015, 21, 2540–2553. [Google Scholar] [CrossRef]
- Steffy, L.Y.; Kilham, S.S. Effects of urbanization and land use on fish communities in Valley Creek watershed, Chester County, Pennsylvania. Urban Ecosyst. 2006, 9, 119–133. [Google Scholar] [CrossRef]
- Steffy, L.Y.; McGinty, A.L.; Welty, C.; Kilham, S.S. Connecting groundwater influxes with fish species diversity in an urbanized watershed. J. Am. Water Resour. Assoc. 2004, 40, 1269–1275. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Area (km2) | Stream (km) | Pipe (km) | Baseflow | Elevation | Slope | Aspect |
|---|---|---|---|---|---|---|---|
| BAIS2 | 3.78 | 5.24 | 0.63 | 0.0435 | 170.51 | 2.72 | 76.88 |
| DR3-1 | 4.87 | 6.30 | 26.94 | 0.0245 | 132.35 | 1.37 | 86.88 |
| DR3-3 | 4.90 | 6.33 | 27.14 | 0.0246 | 132.23 | 1.38 | 8.54 |
| DR3Pi | 4.88 | 6.32 | 26.94 | 0.0245 | 132.35 | 1.37 | −1.78 |
| DRKR2 | 13.93 | 20.12 | 63.73 | 0.0642 | 130.59 | 1.42 | 10.64 |
| GB1 | 1.42 | 1.84 | 2.51 | 0.0105 | 195.71 | 1.72 | 18.11 |
| GB2 | 1.41 | 1.75 | 2.51 | 0.0105 | 195.84 | 1.72 | 65.16 |
| JERJ2 | 22.82 | 31.55 | 23.05 | 0.2626 | 175.71 | 2.14 | −15.23 |
| MR1 | 17.75 | 24.64 | 70.25 | 0.0013 | 184.44 | 1.83 | −3.81 |
| MR2 | 17.73 | 24.56 | 70.23 | 0.0013 | 184.47 | 1.83 | 86.61 |
| SLtribDS | 8.61 | 9.84 | 63.02 | 0.0441 | 164.51 | 1.30 | 16.24 |
| SLtribUS | 7.98 | 9.34 | 57.41 | 0.0408 | 165.53 | 1.28 | 18.85 |
| VN1 | 81.86 | 116.71 | 402.24 | 0.6401 | 170.04 | 1.72 | 75.42 |
| VNTrib | 81.89 | 116.82 | 402.24 | 0.6404 | 170.02 | 1.72 | 54.32 |
| Scientific Name | Common Name | Temperature | Reference |
|---|---|---|---|
| Salvelinus fontinalis | Brook Trout | 21–24 °C (stress) | [67,68] |
| 30 °C (CTM) | |||
| Onchorynchus mykiss | Rainbow Trout | 20 to 22 °C (stress) | [68,69] |
| 29 °C (CTM) | |||
| Rhinichthys atratulus | Blacknose Dace | 29.3 °C (CTM) | [70] |
| Faxonius virilis | Virile Crayfish | 25 °C (stress) | [71,72] |
| 26 °C (stress) |
| Metric | Description | Reference |
|---|---|---|
| Frequency (FmaxcT) | Number days max temperature ≥ 20–30 °C | [62] |
| Duration (n) | Maximum consecutive days per event ≥ 20 °C | [70] |
| MovingAMaxT | Max of 21, 14, 7 day moving average of max | [62] |
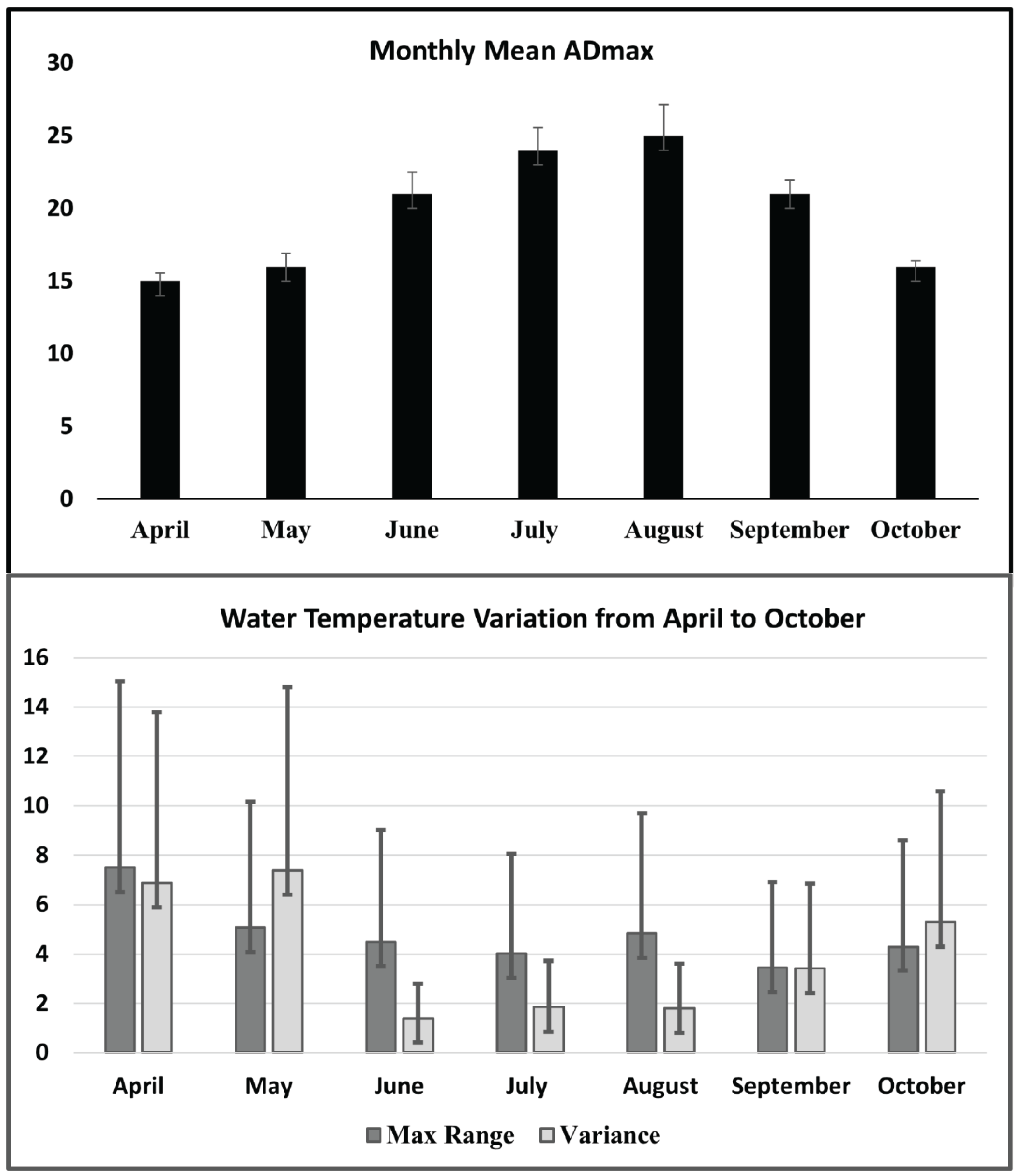
| Monthly ADmax | Average daily max, April to October | [62] |
| Max range per month | Greatest value for difference between daily max and daily min temperature per month | [70] |
| Monthly variance | Variance of mean daily temperatures | [22] |
| Water Temperature Metric (x) | Independent Variables (y) |
|---|---|
| Frequency, Duration | Baseflow + SWPipes + TCLgshd |
| (FmaxcT, Events, n days per event) | Baseflow + SWPipes + TC30 |
| Magnitude | Baseflow + SWPipes + TC5m |
| (MaxMovingAMaxT, ADmax) | Baseflow + SWPipes + ImpLgshd |
| Variability | Baseflow + SWPipes + Imp30m |
| (Max range, Variance of mean) | Baseflow + SWPipes + Imp5m |
| Metric | Model (Y) | t Value | Pr(>|t|) | Model |
|---|---|---|---|---|
| Frequency 20–29 °C | Intercept | 7.54 | 0.00 | 25–29 °C = 139.97–5.73(TC5m) |
| TC5m | −5.73 | 0.00 | Adjusted R-squared: 0.71 | |
| Model: F statistic 32.79, p-value: 0.00 | ||||
| Frequency ≥ 25 °C | Intercept | 6.35 | 0.00 | 25–41 °C = 131.20–4.99(TC5m) + 2.37(BF) |
| TC5m | −4.99 | 0.00 | Adjusted R-squared: 0.73 | |
| BF | 2.37 | 0.04 | Model: F statistic 18.40, p-value: 0.00 | |
| Duration (n) days ≥ 20 °C | Intercept | 8.62 | 0.00 | Max Days = 182.20–4.93(TC5m) + 2.33(SWPipes) + 1.51(BF) |
| TC5m | −4.93 | 0.00 | Adjusted R-squared: 0.73 | |
| SWPipes | 2.33 | 0.04 | Model: F statistic 12.40, p-value: 0.00 | |
| BF | 1.51 | 0.16 | ||
| MaxT21 | Intercept | 45.67 | 0.00 | MaxT21 = 21.6586–0.0528(TC5m) + 0.0084(SWPipes) |
| TC5m | −8.85 | 0.00 | Adjusted R-squared: 0.97 | |
| SWPipes | 11.41 | 0.00 | Model p-value: 0.00 | |
| MaxT14 | Intercept | 78.67 | 0.00 | MaxT14 = 21.7167–0.051(TC5m) + 0.0097(SWPipes) |
| TC5m | −14.68 | 0.00 | Adjusted R-squared: 0.99 | |
| SWPipes | 22.74 | 0.00 | Model p-value: 0.00 | |
| MaxT7 | Intercept | 134.11 | 0.00 | MaxT7 = 23.1676–0.0505(TC5m) + 0.0117(SWPipes) |
| TC5m | −23.26 | 0.00 | Adjusted R-squared: 0.99 | |
| SWPipes | 43.68 | 0.00 | Model p-value: 0.00 |
| Metric | Model (Y) | t Value | Pr(>|t|) | Model |
|---|---|---|---|---|
| AprADMax | Intercept | 91.95 | 0.00 | AprADmax = 14.23 + 4.39(Imp30m) + 2.61(SWPipes) |
| Imp30m | 4.39 | 0.00 | Adjusted R-squared: 0.79 | |
| SWPipes | 2.61 | 0.02 | Model: F statistic 25.48, p-value: 0.00 | |
| MayADMax | Intercept | 27.28 | 0.00 | MayADmax = 19.78–6.72(TC5m) + 2.43(SWPipes) |
| TC5m | −6.72 | 0.00 | Adjusted R-squared: 0.89 | |
| SWPipes | 2.43 | 0.03 | Model: F statistic 54.86, p-value: 0.00 | |
| JuneADMax | Intercept | 19.40 | 0.00 | JuneADmax = 25.94–5.28(TC5m) + 2.63(SWPipes) + 2.08(BF) |
| TC5m | −5.28 | 0.00 | Adjusted R-squared: 0.88 | |
| SWPipes | 2.63 | 0.03 | Model: F statistic 31.79, p-value: 0.00 | |
| BF | 2.08 | 0.06 | ||
| JulyADMax | Intercept | 17.56 | 0.00 | JulyADmax = 28.73–4.25(TC5m) + 2.28(SW Pipes) + 1.75(BF) |
| TC5m | −4.25 | 0.00 | Adjusted R-squared: 0.83 | |
| SWPipes | 2.28 | 0.05 | Model: F statistic 21.58, p-value: 0.00 | |
| BF | 1.75 | 0.11 | ||
| AugADMax | Intercept | 15.40 | 0.00 | AugADmax = 30.87–3.64(TC5m) + 3.22(BF) |
| TC5m | −3.64 | 0.00 | Adjusted R-squared: 0.68 | |
| BF | 3.22 | 0.01 | Model: F statistic 14.72, p-value: 0.00 | |
| OctADMax | Intercept | 92.43 | 0.00 | OctADmax = 17.26–9.43(TC30m) + 2.36(BF) |
| TC30m | −9.43 | 0.00 | Adjusted R-squared: 0.88 | |
| BF | 2.36 | 0.04 | Model: F statistic 45.89, p-value: 0.00 | |
| JuneVar | Intercept | 10.34 | 0.00 | JuneVar = 0.89 + 5.38(Imp5m) + 3.58(SWPipes) |
| Imp5m | 5.38 | 0.00 | Adjusted R-squared: 0.79 | |
| SWPipes | 3.58 | 0.00 | Model: F statistic 27.4, p-value: 0.00 | |
| OctRange | Intercept | 15.35 | 0.00 | OctRange = 6.37– 5.15(TC) |
| TC | −5.15 | 0.00 | Adjusted R-squared: 0.70 | |
| Model: F statistic 26.56, p-value: 0.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Timm, A.; Ouellet, V.; Daniels, M. Riparian Land Cover, Water Temperature Variability, and Thermal Stress for Aquatic Species in Urban Streams. Water 2021, 13, 2732. https://doi.org/10.3390/w13192732
Timm A, Ouellet V, Daniels M. Riparian Land Cover, Water Temperature Variability, and Thermal Stress for Aquatic Species in Urban Streams. Water. 2021; 13(19):2732. https://doi.org/10.3390/w13192732
Chicago/Turabian StyleTimm, Anne, Valerie Ouellet, and Melinda Daniels. 2021. "Riparian Land Cover, Water Temperature Variability, and Thermal Stress for Aquatic Species in Urban Streams" Water 13, no. 19: 2732. https://doi.org/10.3390/w13192732
APA StyleTimm, A., Ouellet, V., & Daniels, M. (2021). Riparian Land Cover, Water Temperature Variability, and Thermal Stress for Aquatic Species in Urban Streams. Water, 13(19), 2732. https://doi.org/10.3390/w13192732