Abstract
Transitional waters straddle the interface between marine and terrestrial biomes and, among others, include fjords, bays, lagoons, and estuaries. These coastal systems are essential for transport and manufacturing industries and suffer extensive anthropogenic exploitation of their ecosystem services for aquaculture and recreational activities. These activities can have negative effects on the local biota, necessitating investigation and regulation. As a result of this, EcoQS (ecological quality status) assessment has garnered great attention as an essential aspect of governmental bodies’ legislative decision-making process. Assessing EcoQS in transitional water ecosystems is problematic because these systems experience high natural variability and organic enrichment and often lack information about their pre-human impact, baseline, or “pristine” reference conditions, knowledge of which is essential to many commonly used assessment methods. Here, foraminifera can be used as environmental sentinels, providing ecological data such as diversity and sensitivity, which can be used as the basis for EcoQS assessment indices. Fossil shells of foraminifera can also provide a temporal aspect to ecosystem assessment, making it possible to obtain reference conditions from the study site itself. These foraminifera-based indices have been shown to correlate not only with various environmental stressors but also with the most common macrofaunal-based indices currently employed by bodies such as the Water Framework Directive (WFD). In this review, we firstly discuss the development of various foraminifera-based indices and address the challenge of how best to implement these synergistically to understand and regulate human environmental impact, particularly in transitional waters, which have historically suffered disproportionate levels of human impact or are difficult to assess with standard EcoQS methods. Further, we present some case studies to exemplify key issues and discuss potential solutions for those. Such key issues include, for example, the disparate performance of multiple indices applied to the same site and a proper assignment of EcoQS class boundaries (threshold values) for each index. Disparate aptitudes of indices to specific geomorphologic and hydrological regimes can be leveraged via the development of a site characteristics catalogue, which would enable the identification of the most appropriate index to apply, and the integration of multiple indices resulting in more representative EcoQS assessment in heterogenous transitional environments. In addition, the difficulty in assigning threshold values to systems without analogous unimpacted reference sites (a common issue among many transitional waters) can be overcome by recording EcoQS as an ecological quality ratio (EQR). Lastly, we evaluate the current status and future potential of an emerging field, genetic biomonitoring, focusing on how these new techniques can be used to increase the accuracy of EcoQS assessment in transitional systems by supplementing more established morphology-based methods.
1. Introduction
The protection and restoration of continental and marine waters and their essential ecosystems has prompted international regulation and legislature from national governing bodies such as the Clean Water Act (CWA, 33 U.S.C. §1251 et seq. 1972), introduced by the U.S. Environmental Protection Agency; within Europe the Water Framework Directive (WFD, 2000 [1]), and the Marine Strategic Framework Directive (MSFD, 2008 [2]). It involves evaluating the ecological quality status (EcoQS, Figure 1 herein) to assess the health of water systems. Noticeably, transitional water bodies are particularly difficult to assess due to their natural variability and require an in-depth ecological understanding to obtain meaningful results. The term “transitional” was introduced about 20 years ago in the WFD (Water Framework Directive, WFD, 2000 [1]) as a means to complete the continuum between continental and coastal waters. Transitional waters are water bodies influenced by both oceanic and freshwater regimes and include estuaries, deltas, rias, lagoons, and fjords (Table 1). In the WFD, transitional waters are defined as “bodies of surface water in the vicinity of river mouths which are partially saline in character as a result of their proximity to coastal waters but which are substantially influenced by freshwater flows” [1]. These waters may also include mesohaline, poly-euhaline, and hyperhaline lagoons [3]. Human-modified transitional waters are classified as “artificial water bodies” [1].
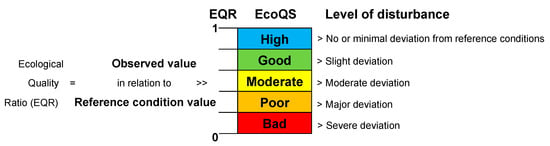
Figure 1.
The basic principles of the classification of ecological status based on the EcoQ ratios assigned, assuming equal intervals between the different classes (adapted and re-drawn from [4]).

Table 1.
Water body types of intertidal areas and transitional waters (modified after [16]).
These ecosystems, located between sea and land, results in a patchwork of highly heterogeneous conditions, which require easily implemented and robust biotic indices. Biotic indices based on the indicator species concept, i.e., on the specific response to organic matter enrichment, for instance, are not fully reliable to assess EcoQS in these water body types (i.e., [5,6,7,8]). In fact, the natural features of these ecosystems make it difficult to disentangle natural and human-induced changes. In particular, silt, clay, and organic matter (OM) sedimentary contents are naturally high in transitional waters, promoting tolerant and opportunistic species, while sensitive species naturally decline [9,10]. Furthermore, total organic carbon (TOC) in transitional waters is a mix of labile and refractory OM with important terrestrial inputs [11]. In transitional waters, benthic communities could therefore be naturally similar to those found in anthropogenically disturbed areas [6,10]. Consequently, pristine, naturally disturbed intertidal areas and transitional waters could easily be misclassified in moderate to bad EcoQS (the “estuarine quality paradox”; [9,12,13]), providing misleading estimates of reference conditions and severely complicating the decision-making process [6,8,14,15].
The EcoQS is a comparative measure of the current condition of a system, compared to that of a reference system free from the negative impact (Figure 1), e.g., heavy metal pollution. It is therefore essential to obtain reference conditions specific to each site from a relatively “pristine” period (before anthropogenic pollution can be detected in the sediment) and compare these to the conditions thereafter. Here lies the central conundrum in EcoQS assessment, as no marine ecosystems on earth today, let alone those at coastal proximity, can be found in “pristine” condition [5,17,18]. Transitional and coastal systems have hosted a disproportionately high human activity for centuries, and as a result, information regarding their pre-impact reference conditions is often missing. Additionally, even if a relatively “pristine” comparison site can be located, the disparate physical and geochemical setting will immediately make it less useful as an analogue [19,20,21]. Hence, for most EcoQS assessment indices, a knowledge gap exists in defining the initial “pristine reference conditions” needed to quantify the change of EcoQS. Without this information to provide a baseline for comparison to a more contemporary EcoQS, the effect of anthropogenic stress cannot be properly determined [19,22].
In this context, palaeoecology can be used to bridge the knowledge gap by providing reference conditions from the exact study site prior to any anthropogenic impact. Having high preservation and fossilization potential makes foraminifera reliable palaeoecological indicators, with the potential to provide reference data from the preindustrial period and beyond [19,20,23,24]. For instance, a more robust assessment of EcoQS for transitional waters was obtained with benthic foraminifera using site-specific local reference conditions in the Oslofjord (Norway) [19], in the Boulogne sur Mer harbor (France) [23], and in the Santos estuary (Brazil) [25]. Furthermore, geochemical analyses of dated sediment cores can be used in combination with benthic foraminifera to determine ecological reference conditions in transitional waters [20,26]. In this review paper, we will describe and discuss how several key characteristics of foraminifera-based indices can be used to address this issue. Particularly how they can be used to provide comparable data from the target site, complete with its unique biogeochemical setting, to set a reliable baseline facilitating the calculation of a representative modern-day EcoQS value.
This paper aims: (1) to detail the scientific background of benthic foraminiferal indices, (2) to list the existing indices, (3) to exemplify their implementation in transitional waters, (4) compare and analyse each index, and (5) present ideas to improve their implementation. Considering that EcoQS assessment with benthic foraminifera is still in its infancy, it appears to be timely in order to ensure accurate application of these indices. In this context, the present work specifically focuses on the relevance of benthic foraminiferal indices to assess EcoQS in transitional waters. First, we review the literature to identify current issues in defining accurate reference conditions and other gaps in EcoQS assessment. Secondly, we identify the strengths of benthic foraminifera to actively bridge these gaps. Thirdly, a critical assessment of existing foraminiferal biotic indices is done in order to evaluate their potential of being implemented in transitional waters. Finally, we discuss future directions that the research on foraminiferal indices may need to follow to reach better monitoring of the health of transitional waters, noticeably the strengths and weaknesses of environmental-DNA metabarcoding.
2. Benthic Foraminifera Are Reliable Indicators of Environmental Conditions
Benthic foraminifera have been shown to respond quickly even to rapid and unprecedented environmental changes in transitional waters; by adapting their species composition and population densities, these protists are able to provide valuable ecological proxies [26]. Since their initial use in the early 1960s by Resig and Watkins [27], benthic foraminifera have proved to be a reliable ecological indicator of various types of anthropogenic stress, including desalination discharge [28], oil-spills [29,30,31,32], aquaculture [33,34], sewage [35,36], heavy metal pollution [22,29,37,38,39,40], and pulp mill effluents (e.g., [41]). At the community and species level, ecological studies have used these protists as a biological quality element (BQE) to decipher the relationship between both natural and anthropogenic environmental variables and the local biota in transitional waters. Foraminiferal assemblages provide additional data (Figure 2) such as changes in population density and species abundance, e.g., [42,43] community compositional shifts (pre-impact sensitive species vs. post-impact opportunists: e.g., [24]) or calcareous vs. agglutinated species: e.g., [44,45,46,47], as well as sensitivity and diversity indices [19,48,49,50], changing depth of habitation in the sediment (observed with living individuals: e.g., [30,51], development of aberrant tests [21,50,51,52,53,54,55] and pollutant accumulation within tests, e.g., [39,56,57]. In fjords [58,59], lagoons [38,49,60], estuaries, and harbours [20,61,62] alike, foraminiferal proxies have been shown to correlate with environmental changes. For instance, certain opportunistic species such as Stainforthia fusiformis [47,58,63], Spiroplectammina biformis [24], Elphidium incertum [64], Ammonia aomoriensis (as A. beccarii: [65]), and Ammonica beccarii [66] have all been linked to adverse environmental conditions caused by OM enrichment and severely hypoxic bottom waters. Possible features behind the superiority of the opportunists in stressful conditions can be related to the presence of specific intracellular complexes in the living cell (S. fusiformis: [67]); ability to denitrify (S. fusiformis: [68]), adjust the depth of habitation in the sediment (E. incertum: [51]), as well as dormancy inside of a protective cyst (or not) during unfavorable conditions such as anoxia (E. incertum: [51,69]), reduced temperature (Ammonia tepida: [68]), temperature and chemical exposure (Amphistegina gibbosa: [70,71]), or extended darkness [72].

Figure 2.
Overview of foraminiferal applications within biomonitoring studies, including (a) EcoQS indices based on species diversity and sensitivity; (b) reconstruction of preindustrial baseline (reference conditions); (c) accumulation of pollutants (e.g., heavy metals and microplastics) within foraminiferal shells; (d) species responses to environmental stress present in situ (e.g., field studies) and (e) simulated in the lab (e.g., culturing experiments); and (f) genomic methods, such as eDNA and sedaDNA.
3. Foraminiferal Biotic Indices
Methods to collect and process benthic foraminifera vary widely in sample acquisition, the sampling gear used, sub-sampling, replication, preservation, preparation, and analysis. In an attempt to address these inconsistencies, Schönfeld et al. [73] proposed using a unified international protocol following a workshop by FOBIMO (Foraminiferal Biomonitoring) group members, which occurred in 2011 in Fribourg, Switzerland. A group of 37 scientists collaborated in the effort to guarantee reproducibility and comparability across studies, with a mind to be adaptable to local conditions [73], inspiring more working groups worldwide to accept foraminifera as a promising BQE to assess EcoQS in contemporary environments. In recent years, however, with the wider acceptance and refinement of the methodology, the momentum of foraminifera index application is gathering rapidly. New foraminiferal indices have been developed, applied, and contrasted between a more extensive range of transitional waters and stressors (Appendix A: Table A1). Below, we provide an overview of foraminiferal indices developed to our knowledge to date based on species diversity and sensitivity and used for EcoQS assessment in transitional waters.
3.1. Foraminiferal Diversity Indices
Using benthic foraminiferal diversity indices, together with associated geochemical parameters, to specifically address human environmental impact was first implemented by Alve et al. [21] investigating changes of ecological status back in time (referred to as paleo-EcoQS) [21]. Although the concept of using foraminiferal diversity had been applied to anthropogenic stress biomonitoring studies earlier [33,35], Alve’s pioneering study was based on sediment archives from Oslofjord, Norway, where the authors used foraminiferal data to obtain the Hurlbert’s ES (100) and Shannon–Wiener H’ (log2) indices, (Table 2), which are the benthic macrofauna diversity indices used by the Norwegian Environmental Protection Agency to assess EcoQS. The work of Alve et al. [21] demonstrated that foraminiferal indices reliably reflect the history of pollution in the region and can be used to obtain the preindustrial reference conditions from “beyond time intervals covered by observational biological time-series”. Since the first use of benthic foraminiferal-based diversity indices, the methodology has been adjusted by Bouchet et al. [74], who used Hill’s number (N1) Exp (H’bc) (1973) and developed novel quantitative threshold values for the EcoQS categories (Table 3) [74].
Table 2.
Threshold values for determining EcoQS classes according to Hlog2 and ES100 [75], Exp (H’bc) [49,74], TSI-Med [50], FSI [76], Foram-AMBI [77], and NQIf [75].
Table 3.
Threshold values for determining EcoQS classes according to Hlog2 and ES100 [75] and Exp (H’bc) [49,74].
The foraminifera-based Exp (H’bc) method has, since then, been successfully tested in a variety of transitional waters, i.e., fjords [74], lagoons [78], harbors [61], bays [78], and estuaries [25,79,80], in both subtidal [49] and intertidal conditions [78]. Furthermore, the index was successfully implemented in different biogeographical regions, i.e., Scandinavia [19], European Atlantic coasts [23,78], the Mediterranean Sea [49], and in South America in Brazil [25]. All these studies have demonstrated several advantages of using benthic foraminifera-based diversity indices. Index Exp (H’bc) has shown a statistical relationship observed between the foraminiferal distribution and environmental parameters, such as organic matter enrichment in the context of aquaculture [71,81], sewage outfall [82], or metal pollutants such as copper (Cu), zinc (Zn), and lead (Pb) [48]. It is, however, important to note that recording such correlations does not necessitate a causal link without considering other key parameters and synergistic effects known to influence benthic foraminifera (e.g., organic carbon flux, salinity, sediment pore-water hypoxia/anoxia), which usually require lab culturing and controlled experimentation [21]. Despite this, studies using the Exp (H’bc) index have been shown to be applicable to different stressors and in disparate biogeographic regions, presenting the potential for wider application, which can be supplemented by more detailed lab-based experimentation. The studies have also found some shortfalls, e.g., the accuracy of the results is dependent on the sampling effort (number of replicates), high seasonal variability may affect EcoQS assessment, and that the method needs to be better adjusted to the naturally low baseline diversity in transitional waters [21,74,83].
3.2. Foraminiferal Sensitivity Indices
Sensitivity indices, strictly speaking, are based on a theoretical succession of species of each sensitivity category along a certain perturbation gradient (Figure 3). Based on ecological preferences, the taxa are assigned to various sensitivity categories, which are given a numerical ranking, varying from 1–5 or even higher depending on the target specificity. The species abundance in each category changes with the amount of OM, other pollution, or stress within the environment; this delineates the biotic state of the site. This approach has some limitations, as it oversimplifies species response to environmental parameters if only considering TOC. Specifically, it is questionable if this method to assign species according to their response to TOC gradients is also applicable in transitional waters, where TOC is reflecting both labile and refractory OM [11]. Noticeably, it was, however, not possible to assign typical salt marsh species from the English Channel, the European Atlantic coast, and the Mediterranean Sea [84] due to the presence of labile and refractory OM that hampers TOC characterization [85,86].

Figure 3.
Theoretical relative abundance patterns for benthic foraminifera species along the main stress gradients (total organic carbon), grouped into the five ecological categories adapted from [77,87].
In this section, some examples of sensitivity indices (Table 2) used to evaluate foraminiferal assemblage response to environmental stressors are presented and discussed, and those include indices such as FIEI [88,89], TSI-Med [51], FSI [76], and FoRAM-Index [86,90,91]. Some indices are designed to evaluate the statuses of warm-water coral reefs. We chose to include those indices herein, as some transitional waters such as fjords in Norway and New Zealand are also known to host cold-water coral reef ecosystems [92]. We also discuss how the natural features of transitional waters may affect the outcome of sensitivity-based foraminiferal indices such as Foram-AMBI and comparable methods [79,93].
3.2.1. Foraminiferal Index of Environmental Impact (FIEI)
In response to drill-cutting disposal taking place in tropical east Atlantic outer shelf environments, Mojtahid et al. [88] designed the Foraminiferal Index of Environmental Impact (FIEI). This index is calculated as the cumulative percentage of all pollution-tolerant and/or opportunistic species observed within a system. Here definition of the “opportunistic species” was based on their patterns of distribution throughout the study area; this was combined with the comparison of the contemporary assemblage, in the uppermost 0–2 cm, with that preserved 2–3 cm below the surface of the sediment. The lower sediment intervals were used as the pre-impact “baseline conditions” [88,89]. This technique has been shown to be more discriminative than macrofaunal indices applied in the same study area, highlighting the benefits of foraminifera in providing a working ecological baseline for biomonitoring [87]. However, such an approach may suffer from circular thinking, as it implies testing the index on the same data set used for the classification of species into tolerant and opportunistic groups. The formula for the FIEI index is given below:
where Nr is the total quantity of pollution-resistant taxa, N0 is the number of individuals of opportunistic taxa, and Ntot. is the total number of counted foraminifera.
3.2.2. Tolerant Species Index (TSI-Med)
Later, working at the French Mediterranean coast, Barras et al. [50] concluded that an index based on the relative proportion of stress-tolerant foraminiferal taxa within a system was the most indicative metric of environmental quality” [50]. Several other parameters were tested in conjunction with stress-tolerance proportion: diversity, indicative species groups, and wall structure proportion, but due to highly variable coastal conditions, all of which were ruled out as lacking significant correlation with coastal ecosystem quality [50]. This index was named the Tolerant Species Index for the Mediterranean (TSI-Med or % TSI) and was developed in response to recent changes to the ecological conditions observed in coastal areas, where there has been strategic clustering of industrial development [50]. The formula for the TSI-Med index is given below:
where %TSx is the percentage of tolerant species and %TSref is the theoretical percentage of tolerant species.
An advantage of the TSI-Med index is that it also integrates a correction for natural eutrophication phenomena involving the definition of reference conditions based on grain size distribution. Although this process is rigorous, the assignment of species to ecological groups often also involves the use of expert judgment to complement the literature review. Unfortunately, species classification, even by the best professional judgment, may fail [94]. Quantifying species response to organic enrichment should have also been used to strengthen the classification [77,84,87]. In fact, there is unlikely to be any single approach that works, and it should be recommended to combine literature review, best professional judgment, and numerical analysis in order to achieve the most accurate species assignment [95].
3.2.3. Foraminiferal Stress Index (FSI)
At the Saronikos Gulf, one of the most anthropogenically impacted coastal areas of Greece, Dimiza et al. [76] developed the novel index FSI (Foram Stress Index). Here, the benthic foraminiferal assemblage was grouped into two ecological groups according to their tolerance to OM enrichment based on a literature review [76]. Similar to TSI-Med, FSI lacks numerical quantification to support species assignments. The relative percentages of the weighted proportion of each group were used to formulate and define five ecological-status classes (similar to that of the AMBI index, see below). The results were promising. At this locality, FSI produced “Poor”, “Moderate”, and “Good” EcoQS rankings, which corresponded well with the level of impact. These were found to correlate strongly with the macrofaunal sensitivity index BENTIX [96], which was applied in the same areas. The formula for the FSI index is given below:
where Sen is sensitive taxa, and Str is stress-tolerant taxa.
3.2.4. Foraminiferal AZTI Marine Biotic Index (Foram-AMBI)
Foram-AMBI has been used in several notable studies [84,87,93] and has been suggested as an additional assessment element to be incorporated into WFD and MSFD’s coordinated approach [77]. Taken from Grall and Glémarec [97] and Borja et al. [98], this index was adapted from the AMBI index (originally used to classify macrofauna). Noticeably, benthic macrofaunal species are traditionally assigned to five groups of sensitivity to OM [96,98] based on the seminal work of Pearson and Rosenberg (1978). Species are meant to be indicative of the prevailing environmental conditions, i.e., the level of organic carbon enrichment where they are found. The following characteristics were used to assign benthic foraminiferal species to the five ecological groups (EGs: Figure 3) considering their response to TOC [84,87,94,97,98]:
Group V (EGV): “sensitive species” are sensitive to TOC enrichment. Their relative abundance is highest at the lowest TOC values and drops to zero as organic carbon concentration increases.
Group II (EGII): “indifferent species” are indifferent to the initial stages of organic carbon enrichment and never dominate the assemblage. They occur in low relative abundance over a broad range of organic carbon concentrations but are absent at very high concentrations.
Group III (EGIII): “tolerant species” are able to endure excess organic carbon enrichment. They may occur at low TOC; their highest frequencies are stimulated by organic carbon enrichment, but they are absent at very high organic carbon concentrations. This group has been termed “third-order opportunistic species” [94].
Group IV (EGIV): “second-order opportunistic species” show a clear positive response to organic carbon enrichment with maximum abundances between the maxima of EGIII and EGV (Figure 3).
Group V (EGV): “first-order opportunistic species” exhibit a clear positive response to excess organic carbon enrichment with maximum abundances at a higher stress level induced by organic load than species belonging to EGIV. At even higher TOC concen- trations, foraminifera are not able to survive.
The abundance of each category of taxa is indicative of the EcoQS of the environment (Figure 3). Here, the Foram-AMBI index is applied to benthic foraminifera; calculated in the same manner as it was previously designed for macrofauna [98]:
Foram-AMBI = 0 × EG1 + 1.5 × EG2 + 3 × EG3 + 4.5 × EG4 + 6 × EG5
Foram-AMBI was first used in the Northeast Atlantic, Arctic fjords, continental shelves, and upper slopes by Alve et al. [77]. In this study, the correlation between TOC and the species sensitivity assignment was based on 19 publications and was tested against independent validation data sets (VDS) to avoid circular arguments [77]. The results confirmed that in organically enriched systems, foraminiferal sensitivity, reflected in assemblage changes, correlates well with TOC gradients (Figure 4). Recently, the assignment of species to the five EGs in transitional waters along the English Channel, European Atlantic coast, and in the Mediterranean Sea allowed for further implementation of Foram-AMBI in transitional waters [84]. The results showed similar correlations to those observed in Alve et al. [77]. A significant positive correlation was observed between Foram-AMBI and organic matter in the two validation data sets. In one of the VDS, the results from Foram-AMBI were also compared to the macrofaunal AMBI index at the same sites, yielding a significant correlation (R2 = 0.56, p < 0.01) between both indices and adding to the support for benthic foraminifera as a reliable BQE. These applications support the implementation of Foram-AMBI. However, recent studies [49,99] already suggested to re-assign some species (originally assigned by Alve et al. [77]) into different groups. Except for the Mediterranean Sea [87], the best professional judgment was not used to support the outcome of the numerical approach to assign foraminiferal species to ecological groups [77,84]. We suggest here that a combination of expert judgment informed by literature review and numerical methods should be used in order to obtain the best possible species assignments.

Figure 4.
Foram-AMBI index plotted against the total organic carbon (%) in the sediments [94] regression line for validation data set 1 from the Norwegian Skagerrak (VDS 1) is shown by the black line and for validation data set 2 from the Oslofjord (VDS 2) is shown by the grey line (raw data from J. Dolven and E. Alve).
3.2.5. Foraminifera in Reef Assessment and Monitoring (FoRAM-Index)
The use of foraminifera-based sensitivity indices for EcoQS goes all the way back to 2003 when Hallock et al. [90] developed the FoRAM-Index (Foraminifera in Reef Assessment and Monitoring) to assess the health of tropical coral reef systems. The FoRAM-Index groups foraminifera associated with coral reefs into three categories: large benthic foraminifera, other small heterotrophic taxa, and stress-tolerant foraminifera. The foraminiferal assemblages typically found in reef environments have been shown to be indicative of both the ambient water quality and reef recovery via nursery transplants or coral recruitment. Additionally, the FoRAM-Index was designed to require only limited computing abilities and hinges on uncomplex calculations, making it user-friendly and easy to implement worldwide for researchers of varying expertise [90]. Since its inception, FoRAM has been successfully applied by 23 separate studies [91] and is able to provide resource managers with data to determine the suitability of benthic habitats for communities dominated by photosymbiont-bearing organisms. The formula for the FoRAM-Index is given below:
where Ps represents the proportion of symbiont-bearing, Po stress-tolerant, and Ph other heterotrophic taxa.
3.3. Foraminiferal Multi-Metric Index
Lastly, Alve et al. [75] proposed a new foraminifer-based multimeric index (NQIf) based on the macrofaunal Norwegian Quality Index (NQI) and tested it in Norwegian fjords on the Skagerrak coast [75]. The study combined paired samples of benthic macrofauna and foraminifera with associated bottom water dissolved oxygen and sediment TOC data. The two BQEs (foraminifera and macrofauna) were intercalibrated via linear regression, and the threshold values for the EcoQS categories were based on those already defined for macrofauna-based indices by the Norwegian governmental guidelines.
This index is composed of the following metrics: (i) a diversity component InS/In (InN), where S is the number of taxa and N is the abundance, (ii) a sensitivity component (AMBI), and (iii) a correction factor to down-weight artificially high diversity values in small samples (few individuals, N/N + 5). Both the macrofaunal and foraminiferal NQI were found to be significantly correlated with bottom water dissolved oxygen concentration; however, in very low oxygen conditions, only foraminifera could be used for EcoQS assessment because macrofauna were absent [75].
The index is an algorithm, where equal weight is given to diversity (50%) and sensitivity (50%) and is formulated as follows:
where AMBIf represents Foram-AMBI (Section 3.2.4) and ES100f represents foraminiferal diversity expressed as Hurlbert’s index.
4. Comparison of EcoQS Indices
4.1. Various Foraminiferal Indices: How Well Do They Perform When Used at the Same Site?
Research shows that foraminifera-based indices have the potential to reveal the ecological status preceding anthropogenic disturbance and are capable of setting informed reference values [26,48,49,91,100,101,102,103]; see Section 5.2 below. To produce meaningful actions, however, the results must be comparable and congruous with each other. At the same time, studies aiming for a comparison of these disparate indices and methodologies reveal some discrepancies in the consensus reached. Below, we provide a case study exemplifying this.
The study by El Kateb et al. [48] assessed EcoQS along the Gulf of Gabes (Tunisia) and incorporated the diversity indices such as Shannon Index (H’), Exp (H’bc), sensitivity indices FSI, % TSstd (using the FSI- assigned species), % TSstd (using Foram-AMBI assigned species), FI’ (Modified FoRAM-Index), and ILS (“long versus short life span” index), which can be considered in combination [48]. Developed by Mateu-Vicens et al. [104], FI’ and ILS rely on foraminifera associated with meadows of the seagrass, present at several locations within the Mediterranean Sea [104]. This study demonstrated that each of these indices assessed EcoQS differently (Figure 5). To understand the reason for this, we must look to the assumptions and criteria considered in each index, as well as the varying geomorphological regimes along the gulf. The indexes that recorded the worst EcoQS values were FI’ and ILS epiphytic foraminiferal indices, FSI, and % TSstd (FSI-assigned species). Conversely the best EcoQS was reported by Foram-AMBI, % TSstd (Foram-AMBI-assigned species), and Shannon Index (H’), while Exp (H’bc) demonstrated intermediate EcoQS values. Begging the question, are the former indices more sensitive to environmental stress than the latter ones? Or perhaps these results reflect the situational aptitude of each index and how compounding environmental variables influence their efficacy? To begin with, FI’ and ILS, both being epiphytic foraminiferal indices, reported predominantly “bad” to “poor” EcoQS along the gulf. These indices rely on the presence of sea grass meadows, which have been in decline due to increasing environmental stress. In the absence of such habitats, there are no epiphytic foraminifera, and the area is recorded as “poor”, meaning these indices reflect the lowest values indicative of unsuitable growth conditions. El Kateb and colleagues concluded that these indices correctly reflect the ecological status of the area due to seagrass Posidonia oceanica’s sensitivity to environmental conditions. Epiphytic indices can, hence, only be used in areas that are known to be suitable for seagrass growth.
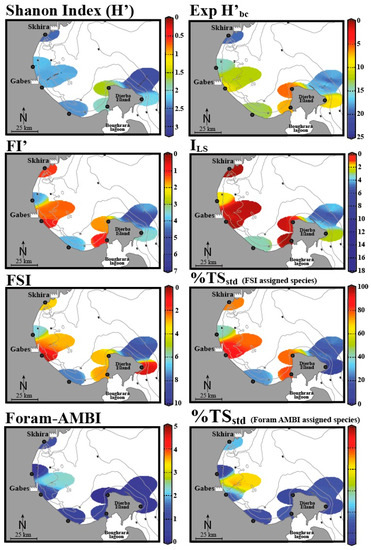
Figure 5.
The EcoQS assessment in the Gulf of Gabes (Tunisia) using various foraminifera-based indices (data is spatially interpolated using DIVA software). Adapted from [48]. Colours indicate various EcoQS classes: blue—high, green—good, orange—moderate, yellow—bad, red—poor.
In contrast, Foram-AMBI, % TSstd (Foram-AMBI assigned species), and the Shannon Index (H’) suggest comparatively unpolluted conditions; to explain this, the authors used two distinct systems sampled from the Gulf region: Gabes, a more polluted area with organic enrichment and fine sediments, and Djerba, with coarser sandy sediments. The near shore silt and mud at Gabes is also reported to be rich in heavy metals (Pb, Li, Zn, Cu) and phosphorous due to industrial waste discharge. Sites with fine sediments appear to be more suitable for application of Foram-AMBI, % TSstd (Foram-AMBI-assigned species), and % TSstd (FSI-assigned species); as the aforementioned indices are all sensitivity-based, the link here is explained by opportunistic/stress-tolerant species dominating systems. In Djerba, with sandy sediments, however, the contrary was observed; here, the diversity indices Exp (H’bc) and H’ as well as the epiphytic indices FI’ and ILS exhibit a stronger relationship. El Kateb et al. further addresses Foram-AMBI alone, pointing out that this index was developed to reflect an increasing OM gradient, relying primarily on sediment mud content, dissolved oxygen, and pollutants, such as metals and polychlorinated biphenyls (PCBs). In the areas of the gulf that do not exhibit these conditions, the index becomes less useful. This highlights that it is essential to understand the components of each index to ensure the results will be representative of the site and that it is advisable to use and compare several indices to reach the soundest conclusion [19].
4.2. Palaeoecological Applications: Toward the Definition of Reference Conditions
In contrast to most benthic macrofauna, the preservation of benthic foraminiferal tests provides information about long-term environmental and biological changes [19,21,23]. Hindcasting approaches (i.e., the use of historical information) are one of the methods recommended by marine legislations to determine reference conditions. Such approaches imply the existence of long-term monitoring or fossil records of taxa used to characterize the EcoQS. Different studies further confirm that fossil benthic foraminifera can enable the reconstruction of in situ local reference conditions in transitional environments with long-term pollution history to determine either contemporary EcoQS or its evolution over time and may contribute to deciphering between natural and anthropogenic stresses. For instance, a more robust assessment of EcoQS for transitional waters was obtained with benthic foraminifera using site-specific local reference conditions in the Oslofjord, Norway [19], in the Boulogne sur Mer harbor, France [23], and in the Santos estuary, Brazil [25]. Furthermore, geochemical analyses of dated sediment cores can be used in combination with benthic foraminifera to determine ecological reference conditions in transitional waters [20,26].
In the Oslofjord (Norway) [19], the results from applying ES100, H’log2, and Exp H’bc diversity indices were found to be consistent with physical and chemical parameters at the site (metals, TOC, and dissolved oxygen). Furthermore, the temporal pattern and scale of change shown by each of the indices are still the same, and this can be used to define deviation from the reference conditions, regardless of the index used. For instance, in Boulogne sur Mer Harbor, local reference conditions from the pre-impacted period allowed the authors to accurately assess EcoQS based on the ecological quality ratio or EQR [23].
The EQR (Figure 1) is the ratio between the value of the observed biological parameter for a given water body and the expected value under reference conditions [4] and is in line with the WFD definition of EcoQS. The ratio ranges as a numerical value between 0 and 1, with high ecological status represented by values close to one and bad ecological status by values close to zero. The EQR boundaries to define Palaeo-EcoQS were determined using the “Reference Conditions Working Group” recommendations [105]. Type-specific reference conditions (including hydromorphological and physicochemical conditions unique to each water body) are the anchor point of EQR based classifications. Hence, the EQR approach can also be used to resolve issues related to the definition of threshold values (see Section 4.3). Using the foraminiferal fossil record, it is possible to determine in situ local reference conditions. For example, in the Santos estuary (Brazil), the highest value of diversity in ~1902, i.e., Exp (H’bc) of 11.66, corresponds to the pre-impacted period and was defined as the reference value. The EQR was calculated accordingly: EQR = Exp (H’bc)observed value/Exp (H’bc)reference value [25]. In that study, the foraminiferal fossil record allowed evaluation of Palaeo-EcoQS based on in situ local reference conditions in accordance with the environmental history of the region.
4.3. Palaeoecological Applications: Defining Correct Threshold Values
EcoQS value uncertainty, due to lack of intercalibration between threshold values, presents some issues for using living foraminiferal assemblages in transitional environments. This can also be observed in Palaeo-EcoQS assessment, which may be exacerbated by preservation bias. By applying the Exp (H’bc) diversity index and threshold values from two separate publications, Bouchet et al. [74] and Bouchet et al. [49], to the foraminiferal assemblage data from Idefjord, the EcoQS of the fjord can be categorized very differently (Table A2) [49,74]. Using threshold values from Bouchet et al. [49], the resulting fjord EcoQS is “Moderate” to “Bad”, with a slight reprise back to “Moderate” observed in the early 2000s. The low natural diversity of the system presents as only “Moderate” EcoQS even back to the 1800s, before the time when the anthropogenic activity began to significantly impact the system, highlighting that indices must be adjusted when applied to transitional waters [24]. Conversely, when the “dry picking” threshold values from Bouchet et al. [74] are applied, the fjord shows mainly “Good” EcoQS values until 1890, where the EcoQS decreases to “Moderate” and “Bad”.
The chosen threshold values from Bouchet et al. [74] were based on foraminiferal assemblages from dry picking the >125 μm sediment fraction, while thresholds from Bouchet et al. [49] are also “dry picking” but “63 μm fraction” boundaries modified after Bouchet et al. [74] based on using the PERSE method (Procedure to Establish a Reference State for Ecosystems [106]); a multivariate non-parametric approach to calculate the relative reference states against which EcoQS fluctuations can be detected and quantified. Table A2 makes it clear that the reconstruction “EcoQS 2” is more realistic based on known pollution history in Idefjord [24] by applying lower diversity thresholds originally obtained for Nordic fjord waters (Bouchet et al. [74]). In contrast, the Exp (H’bc) diversity values (Bouchet et al. [49]) adjusted for the Mediterranean, due to higher species diversity, obviously underestimate changes of EcoQS in Nordic fjord waters. This is an example of issues related to latitudinal diversity gradients and the effect of these on the characterization of transitional water ecosystems across the globe, which makes a strong case against using “universal” threshold values. Among other issues important to consider when setting threshold values or EcoQS boundary classes are sediment size-fraction analyzed for foraminifera, number of replicates, dry versus wet picking method, and sediment characteristics on a site (soft-bottom muddy sediments versus silty and sandy deposits). Hence, future studies should make efforts toward using class boundaries adjusted locally and base these on the above-mentioned characteristics, as well as on local reference conditions. Other possible means to adjust the threshold values can be: indicator equations (a theoretical approach aimed at finding a correspondence between the index equations), reference indicators (a separate indicator used to calibrate other indicators), and indicator distribution laws mathematical properties of the indicator values (such as distribution laws, which are used to obtain the same proportion) [107].
The example presented in Table A2 demonstrates that EcoQS threshold values can have a huge impact on the message displayed by EcoQS assessment index thresholds, which can be exacerbated by preservation bias by natural abiotic and biotic processes (particularly in older sediments), as well as sample preparation methods, such as dry picking vs. wet picking, staining, and sieving. Hence, EcoQS threshold values should be carefully considered upon their application, which makes index calibration and a conversion factor specific for Palaeo-EcoQS assessment in transitional waters a key in obtaining reliable reference conditions.
4.4. Taphonomical Processes and EcoQS Indices
Another key aspect in characterizing contemporary environmental and paleoenvironmental changes accurately is understanding the difference between living and dead benthic foraminiferal assemblages and all the factors that may lead to their formation [60]. Foraminiferal biomonitoring studies based on total assemblages, particularly those aimed at characterizing environments that have undergone recent and extensive habitat modification, either intentionally (e.g., through aquaculture and marina construction) or unintentionally (e.g., agriculture leading to deforestation and subsequent erosion), are susceptible to bias interpretations due to taphonomic (post-mortem) processes affecting the tests (see review in [108])Those processes, including transportation, CaCO3 dissolution, and test breakage, can occur more readily post-mortem and impact species composition of the total assemblages together with seasonal population shifts (different species having peaks during different times of the year) and influxes of allochthonous taxa to the area. In highly stratified fjords, for example, it has been demonstrated that sediment archives may also be heavily biased toward agglutinated species due to seasonal hypoxia and carbonate dissolution [47,109]. Some studies performed in shallow estuaries showed that seasonal changes of redox conditions can cause almost complete dissolution of calcareous component of the assemblages resulting in total assemblages being strongly dominated by agglutinated foraminifera [47,109,110,111], local hydrodynamism in estuaries may hamper the preservation of foraminiferal fauna by sediment resuspension and mixing [112] and bioturbation processes by macro-invertebrates may enhance Fthe homogenization of the upper-sediment layers [107]. Hence, it is important to recognize the conditions that lead to the highest preservation of foraminifera tests and understand how best to adjust data from sub-optimal conditions to most accurately reflect the true ecological conditions of a site. For instance, at the Saquarema lagoon system in Brazil significant difference was found between the living and dead assemblage distributions [60]. These differences were attributed to the hydrodynamic conditions and seasonal variation.
To reduce the effect of taphononomic or post-mortem bias, Belart et al. [60] suggest that in homogenous systems, areas with high living-dead assemblage similarity may be the primary target for palaeo-environmental assessment (to inform the baseline reference conditions) [60], whereas in highly heterogeneous areas, such transitional waters, an alternative approach would be to undertake palaeoecological studies at all stations and adjust the recorded total assemblages by conversion factors inferred from a DCA or PCA plot [113]. Here, the trade-off between effort (time, expertise) input and representative data output must be considered on a site-to-site basis as significantly more extensive data treatment and analysis would be involved.
4.5. Comparison of Foraminifera-Based Ecoqs Indices with Traditional Macrofauna-Based Indices
Increasingly, a multi-metric approach, incorporating several biomonitoring strategies, is being adopted across international management operations; for example, the macrofauna-based Norwegian Quality Index (NQI, see Section 3.3), which employs a sensitivity component with the AMBI index in combination with the H’log2 and ES100 diversity [114,115]. In fjords of Norway, Alve et al. [75] has shown that applying an adapted version of the macrofaunal NQI to foraminifera (NQIf) yields a similar indicator efficiency as its macrofaunal equivalent (Figure 6a); eventually concluding that foraminifera should be included among the biological elements defined by the WFD and used for EcoQS assessment [75]. Another example from Norwegian fjords, where the benthic foraminiferal communities were shown to significantly correlate with benthic macrofaunal ones, comes from the study of Bouchet et al. [84] (Figure 6b). Furthermore, in the oyster farming areas of the Pertuis Charentais (France), Foram-AMBI and macrofaunal AMBI were also significantly correlated [84]. All these examples suggest that other benthic foraminiferal indices may correlate significantly with their macrofaunal counterparts, demonstrating that benthic foraminifera-derived indices can be used to complement the results of other BQEs and can be applied where macrofauna are absent or difficult to sample.

Figure 6.
(a) Significant correlation (R = 0.70, p < 0.0001) between foraminiferal and macrofaunal NQI indices in Norwegian fjords [75]. (b) Significant correlation (procrustes errors analysis m2 = 0.66, p = 0.001) between foraminiferal and macrofaunal communities in Norwegian fjords [83].
5. The Genetic Era
Following the successes of the Foram-AMBI and other indices, foraminifera-based indices are now being considered from a novel perspective. Basing environmental quality assessment on DNA (or RNA) present in environmental samples is a promising technique in working toward the development and implementation of both cost-effective, high-impact, and robust methodologies for the long-term monitoring and management of marine ecosystems. An increase in sophistication and accessibility of genetic technologies has fuelled a surge of research into using these as a biomonitoring tool, prompting some to refer to this period of innovation as the “ecogenomic era” [116] or “biomonitoring 2.0” [117]. Below, we focus on two genetic approaches, eDNA and aDNA, increasingly used in biomonitoring with foraminifera as BQE.
5.1. Environmental DNA
Environmental DNA (eDNA) represents a mixture of the genomic DNA, which was present in the living cells and preserved within cellular organelles, or it is extracellular molecules, which reside in fragments of tissue, secretions, and other organic materials [118]. These eDNA fragments can be found externally from their biotic origin and can be extracted from environmental samples without first isolating the target organism. Surface waters provide a wealth of eDNA in the aqueous or sedimentary form [119]. With the use of high-throughput sequencing (HTS) and bioinformatic pipelines, millions of DNA sequences present in environmental samples can be analysed, and as long as the DNA does not degrade and remains uncontaminated, it is possible to target small or elusive taxa present only at low densities, and sample at the diversity of the existing primers [118,119].
Environmental DNA-based technology is mainly applied in one of three ways: through single-species detection, in biodiversity surveys, or for biotic indices. Depending on the study approach, specific technology and methods are necessary to analyse the resulting molecular information [120]. Firstly, single-species detection is the most simplistic way to use eDNA for biomonitoring purposes. Based on polymerase chain reaction (PCR) amplification, this approach uses species-specific primers to detect endangered, rare, or alien/invasive species. Though its use in marine sediments is limited, studies on alien or harmful species related to industrial activities show promising results [116].
Secondly, using HTS and targeted PCR amplification, eDNA samples can be indicative of the whole biological community. This technique is referred to as eDNA metabarcoding and has been successfully applied in biodiversity surveys as well as bioindication. This technique is often used to investigate marine biodiversity and can shed light on the biogeography and ecology of planktonic and benthic microbes [121], meio- and macrofauna [122]. Increasingly, however, metabarcoding is being used on benthic foraminifera to assess human impacts on the marine environment, ecosystem quality assessment, and biomonitoring.
Several studies have used these new technologies in conjunction with observation-based taxonomic methodologies to verify the use of genetic data in biomonitoring. For instance, the impact of aquaculture, a multitrophic metabarcoding biotic index for monitoring of organic enrichment associated with fish farms has been proposed by Keeley et al. [123] based on benthic foraminifera, bacteria, and general eukaryotes [123]. Bioindicator taxa were identified by metabarcoding eDNA and eRNA for the taxonomic groups and assigning Eco-Groups to the most abundant operational taxonomic units (OTUs); this information was then used to develop a multitrophic metabarcoding biotic index, which showed a strong relationship with the “enrichment stage” (determined by the chemical and biological properties of the site). These techniques have also been successfully employed with benthic foraminifera to assess the environmental effect caused by offshore fossil fuel drilling and production operations [124,125], in monitoring fish farms [126], and in polluted harbors [125]. These studies pointed toward congruence between conventional morphology- and eDNA-based foraminiferal data. Recently, the first application of foraminifera indices on eDNA data showed a good correlation with environmental parameters and congruent results with indices based on morphological data [127], further supporting the implementation of metabarcoding with benthic foraminifera.
The genetic approach is still a new technique and, as a result, is presently regarded as complementary to traditional methodologies rather than a standalone method. This is due to several issues (i) the high number of unassigned OTUs due to low coverage of sequenced species in the GenBank; (ii) presence of propagules (resting stages) of foraminifera in the sediments, which will not necessarily develop into adults, but will be picked up by the eDNA analysis; (iii) high number of monothalamids present in the OTUs but commonly absent as fossils, making a comparison between morphospecies and eDNA complicated; (iv) biases related to the process of DNA extraction and PCR amplification, e.g., underrepresentation of globothalamids compared to monothalamids as the hard shell makes DNA extraction more difficult; and (v) the number of rRNA gene copies, variation nuclei number or ploidy related to alternation of generation in the foraminiferal life-cycle [126,128]. Greater efforts in barcoding research can be used to close some of these knowledge gaps, as well as a taxonomy-free approach, where biotic indices are computed without the taxonomic assignment of sequences, which was first used on benthic diatoms to assess the ecological status of rivers and streams [129]. This process relies on a much higher percentage of OTUs (up to 95%) compared to morphospecies-based taxonomy; indicator values are assigned to particular species or OTUs based on their occurrences within the metabarcoding data. Other studies have proposed a machine learning approach but are seldom used due to the lack of the extensive training data sets needed [128]. Once the methodology has been fine-tuned, a metabarcoding approach could be used to develop EcoQS indices without taxonomic pitfalls associated with erroneous species identification, offering instead the identification of OTUs.
5.2. Ancient DNA
Ancient DNA (aDNA), in a way similar to eDNA, can be recovered even from species that do not leave behind diagnostic microfossils in the sediment, making it a promising approach for refining the reconstruction of palaeo-environmental conditions persisting over millennia to hundreds of thousands of years back in time [130]. Within marine palaeoecology, the term sedimentary DNA (sedaDNA) is usually synonymous with aDNA, as sedaDNA refers to aDNA that is found within the sediment.
Ancient DNA has been shown to be applicable to foraminifera, with the possibility to reveal the presence of species that are otherwise often excluded from species count due to being less readily fossilized, cryptic, or simply too small to detect. For example, previous palaeoecological surveys have shown that the detection of monothalamous and polythalamous foraminifera is possible, and those make a valuable indicator of changes in EcoQS [126,131]. The molecular inclusion of non-fossilized monothalamids in palaeoecological surveys increases the richness observed within the foraminiferal communities compared with that of the fossil record. Monothalamous species, a group of foraminifera characterized by single-chambered, agglutinated, or organic tests, are soft-shelled and often ignored in foraminiferal studies due to their low preservation potential despite their high abundance and diversity, even making up the majority of the species present in some transitional waters, such as fjords in Scotland [126,132]. Hence, the inclusion of this group could potentially improve the overall accuracy of palaeodiversity estimates influencing the assessment of local reference conditions [126,132].
In order to use genetic data to set reference conditions for EcoQS assessment, it must be recoverable from a time before human impact began to negatively affect the environment. Sedimentary aDNA is required from the preindustrial period, but is this feasible?
Foraminiferal DNA can remain contained within the test and be detectable within the sediment [133]. Various studies have shown that plankton-derived DNA, including foraminifera, can be preserved in marine sediments over tens to hundreds of thousands of years [130,131,134]. It has also been reported that environmental DNA was more concentrated in marine sediments than in aqueous form [135,136]. Kirkpatrick et al. [137] reported recovering fossil diatom DNA from up to 1.4 Ma old sediment core samples, the long-term survival of which was attributed to “protection” provided by being surrounded in the diatom microfossils, suggesting that microfossils increase the preservation potential of DNA over time [137]. This points favorably toward the use of foraminifera microfossils as the target organism of similar aDNA studies in the future.
Due to general lack of research in aDNA area, however, Holman et al. [138] have called for more studies to “advance our understanding of how eDNA deposits and degrades in marine sediments in order to temporally contextualize sediment samples”, opting instead to only process the uppermost sediment, only profiling contemporary species composition [138]. The field of genetic biomonitoring, particularly sedimentary or ancient DNA, holds much potential in the effort to constrain reference conditions within the ecological assessment of transitional waters; however, the lack of experience and understanding of post-depositional genetic degradation and taphonomic preservation bias within the sediments renders this methodology at present, only complementary, rather than a viable standalone approach.
6. Foraminiferal Indices: Strengths, Weaknesses and Way Forward
Foraminifera have been shown to be a consistently indicative basis for biomonitoring indices, applied within a wide range of transitional environments. Foraminifera-based indices offer an alternative or complementary methodology to their more widely accepted macrofaunal analogue, particularly for EcoQS assessment in transitional waters, where a unique mosaic of heterogenous habitats converge due to influence from terrestrial, freshwater, and oceanic regimes. The ability to provide reliable ecological information across disparate and often highly variable systems makes foraminifera an ideal BQE for EcoQS assessment in transitional waters. Delineating anthropogenic disturbance versus natural variability is possible due to foraminiferal community composition being comparable directly between sediment layers deposited during contemporary and preindustrial times. Such “preindustrial to modern assemblage composition” can be tracked in sediment archives and directly compared against geochemical changes recorded in the surrounding sediment, which are also able to record ecosystem reprisal and recovery, quantifying the ecological benefits of waste management and industrial regulation. Palaeo-EcoQS assessment can also be instrumental in the determination of pristine reference conditions within the naturally stressed systems of transitional waters. An accurate estimate of EcoQS prevailing long before the anthropogenic economic development and observation-based biomonitoring studies began is necessary to understand the level of human impact and inform counteractive action to ensure proper restoration practices.
6.1. Issues and Resolutions
6.1.1. Differences in Assessed EcoQS Using Different Foraminiferal Indices
In the interest of integrating foraminifera as an official BQE for biomonitoring approved by legislative bodies such as WFD and MSFD, their correlation with macrofaunal indies (the current status quo) has been extensively investigated and shown to be significant. In this pursuit, however, assessment of transitional waters presents an issue; although cross-examination based on both BQEs often shows a similar response to environmental stressors, the various indices based on foraminifera are not always consistent in their EcoQS assessment for the same site. As evidenced by the case studies presented in Section 4, this issue results again from the patchwork nature and naturally stressed conditions found within transitional environments, as well as the individual strengths and weaknesses of the indices within distinct bio-geomorphologic settings. For instance, Foram-AMBI has been developed for fine-grained sediments and is most accurate in OM enriched environments but over-estimates ecological quality in coarse-grained sediments replete with abundant dominant species [48]. Additionally, the threshold values for the EcoQS classes are not easily defined and are usually based on precedent, which can result in the same index, using thresholds applied previously in two different studies, recording different ecological quality values for the same site. To combat these two issues, we firstly suggest an integrated index approach, the rationale in support being that an index appropriate for one environment is not necessarily applicable to the next. To put this into practice, various biotic, abiotic, and geomorphologic features of the site, as well as sampling and sample preparation methods, should be considered as part of the data treatment. In addition, more comparative studies, in which multiple indices are applied to the same system, are needed to provide data on the aptitude of each index and allude to the site characteristics that exhibit the strongest relationship with each index (e.g., Figure 5). Then the most apt index can be chosen and applied to each site to provide the most accurate estimation of ecological health. Although this is more labour-intensive in the present, as the characteristics criteria have yet to be developed and will require some extra diagnostic effort and expert judgment, with an easily accessible checklist (or flow chart) of site characteristics, it would be possible to improve all future biomonitoring assessments within transitional waters and beyond.
6.1.2. Threshold Values
Secondly, when defining threshold values, a similar approach can be considered. It is clear that in such variable systems, such as transitional waters, a given EcoQS evaluation is likely only an indication of the actual status; however, to influence legislation, quantification is key. Using threshold values previously applied to a similar system may be sufficient in some cases; however, as suggested in Section 4.2, a more precise assessment can be achieved by obtaining an EQR value. An exciting benefit of foraminifera-based indices, in situ Palaeo-EcoQS, can be used as the baseline for site-specific valuation, which directly compares the preindustrial and modern-day status of only the target locality. Instead of expressing environmental initiatives as a push to improve water bodies to “good” EcoQS by X year, framing it as achieving an EQR ratio of close to 1, which can also be easily expressed as a percentage allowing for more specific changes to be tracked and targets set.
6.1.3. Intercalibration
Finally, in line with the developments suggested in this review, there has recently been a move toward the intercalibration of benthic foraminiferal indices and macro-faunal indices. This increasingly holistic approach encapsulates more taxa and even entire ecosystems and is being gradually integrated into environmental policy and legislation (e.g., WFD, 2000 [1], MSFD, 2008 [2,124,139,140]). As the application of genetic techniques to marine environments is a recent development, the laboratory and analysis techniques, as well as bioinformatic processing, are not yet fully established or have not yet been standardized [141]. Even less research has been done on the intercalibration of methods across different disciplines with the common goal of achieving a holistic understanding of ecosystem health. In the interest of harmonizing assessment methods for the implementation of the WFD, the EU commissioned the Geographical Intercalibration Work (GIG). The GIG has since inspired several projects, including HARMEX “Harmonisering af Nordiske bundfaunaindeks for økologisk kvalitet i poly- til euhaline områder” (Harmonization of Nordic benthic quality indices in polyhaline and euhaline areas), founded by the Nordic Council of Ministers involving Norway, Denmark, and Sweden [114]. Three Scandinavian multi-metric macrofaunal EcoQS indices (Norwegian NQI and Danish DKI and Swedish BQI) were intercalibrated to assess coastal benthic response to pollution-related stress gradients, using both species diversity and sensitivity. Clear responses to gradients of OM, hypoxia, metals, effluent material, and physical disturbance were observed.
6.2. New Techniques
Metabarcoding has introduced a new potentially valuable tool in ecological assessment studies. Several studies have identified metabarcoding DNA/RNA retrieved from environmental samples as being a useful method for biodiversity assessment and biomonitoring in both past and present, aqueous or sedimentary environments [123,124,142,143]. Studies have instead suggested that the foraminiferal genetic data can provide an additional environmental impact assessment tool where the traditional morphology-based approach is impractical [116]. In addition, the presence of resting stages (propagules) in foraminifera and still relatively low number of sequenced foraminiferal species poses difficulties for the eDNA approach to be used as an independent method. In addition, successfully sequencing the small and often incomplete molecules of DNA/RNA present in sediments is still the biggest barrier to the implementation of this methodology; while it provides some other advantages (see Section 5), the field is still very much in its infancy and will benefit from more years of development and refinement before it can be widely accepted as a standalone alternative. The two methodologies, the morphology-based approach and metabarcoding, are currently regarded as complementary to each other, with disparate scientific trade-offs affecting each, resulting in scenarios where one technique may be more or less appropriate. Here, we advocate for the inclusion of genetic methods into mainstream ecological assessment practices; the additional benefits of this method are clear and require only a small sample volume, which can be collected alongside that used for morphology-based methods. The increased amount of data available about the biological community available at each site will improve assessment accuracy and be available to inform the calibration of indices for future use.
7. Conclusions
Foraminiferal indices and their ability to provide reference conditions beyond observational time series and conventional monitoring have proven their high value for EcoQS assessment even within transitional waters. As evidenced by the range of studies presented here, various aspects of these protists’ living, dead and fossilized assemblages can be used to accurately quantify environmental quality and its temporal changes. Foraminiferal indices can be applied in a variety of environments to obtain meaningful results that can be used to influence legislation; however, they also require careful adjustment, verification, and intercalibration. Gaining an understanding of the characteristics of each index is key in selecting the most appropriate index for a particular region, waterbody, sediment type, assemblage type, and sampling/sample preparation method. Development of a checklist or flowchart to outline the properties of each index and how these relate to site variables would be highly beneficial, helping to both standardize and simplify the process.
Author Contributions
Conceptualization, P.A.J.O. and I.P.A.; methodology P.A.J.O. and I.P.A.; software, P.A.J.O. and V.M.P.B.; investigation, P.A.J.O.; resources, P.A.J.O. and I.P.A.; data curation, P.A.J.O. and I.P.A.; writing—P.A.J.O.; writing—review and editing, P.A.J.O., I.P.A., and V.M.P.B.; visualization, P.A.J.O., I.P.A., and V.M.P.B.; supervision, I.P.A.; project administration, P.A.J.O. and I.P.A.; funding acquisition, I.P.A. All authors have read and agreed to the published version of the manuscript.
Funding
P.A.J.O. and I.P.A received funding from the University of Gothenburg and the Wahlströms Foundation; and V.M.P.B. acknowledges funding from the Agence de l’Eau Artois-Picardie and the Agence de l’eau Seine-Normandie (Foram-INDIC project).
Data Availability Statement
Synthesized data on available literature on the topic are available in Table A1 in the Appendix A. Data used in Table A2. can be found at Polovodova Asteman et al. [24].
Acknowledgments
The authors would like to thank Jane Dolven and Elisabeth Alve for providing the raw data from Alve et al. [94]. Also the two anonymous reviewers and Peter Goethals who provided constructive criticism and feedbacks on the earlier version of the paper.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| aDNA | Ancient DNA |
| AMBI | AZTI’s Marine Biotic Index |
| BQE | Biological quality element |
| CWA | Clean Water Act |
| DCA | Detrended correspondence analysis |
| eDNA | Environmental DNA |
| e.g., | Exempli gratia |
| EQS | Ecological quality status |
| EQR | Ecological quality ratio |
| FI’ | Modified FoRAM-Index |
| FIE | Foraminiferal Index of Environmental Impact |
| FOBIMO | Foraminiferal Biomonitoring group |
| Foram-AMBI | Foraminiferal AZTI’s Marine Biotic Index |
| FSI | Foram Stress Index |
| gAMBI | Genetic AZTI’s Marine Biotic Index |
| GIG | Geographical Intercalibration Work |
| HARMEX | Harmonisering af Nordiske bundfaunaindeks for økologisk kvalitet i poly- til euhaline områder” (Harmonization of Nordic benthic quality indices in polyhaline and euhaline areas) |
| HTS | High-throughput sequencing |
| ILS | “Long Versus Short Life Span” Index |
| MSFD | Marine Strategic Framework Directive |
| nEQR | Normalized ecological quality ratio |
| NQI | Norwegian Quality Index |
| NQIf | Norwegian Quality Index (using Foraminifera) |
| OM | Organic matter |
| OUT | Operational taxonomic unit |
| PERSE | Procedure to Establish a Reference State for Ecosystems |
| PCA | Principal component analysis |
| PCR | Polymerase chain reaction |
| PRISMA | The Preferred Reporting Items for Systematic reviews and Meta-Analyses |
| sedaDNA | Sedimentary DNA |
| TS | Tolerant species |
| TSI-Med | Tolerant Species Index for the Mediterranean |
| TOC | Total organic carbon |
| VDS | Validation data set |
| WFD | Water Framework Directive |
Appendix A
Appendix A.1. Materials and Methods
To exemplify the implementation of foraminiferal indices within transitional environments, a systematic review was conducted and reported according to PRISMA 2020 guidelines.
The eligibility criteria were as follows: Only original peer-reviewed research papers available in full-text pdf format online and presented first-hand research were used; any reviews and commentaries were omitted. Additionally, any papers in which the search term foraminifera or foraminiferal appeared only in the references were also not included.
The database used was Clarivate Analytics’s Web of Science (WoS), an international research repository. The database covers a large quantity of mostly high-impact, English-language publications and is regarded as the world’s leading citation search and analytical information platform [60,144].
The search was not constrained by year and followed a structure similar to that applied in Sousa et al. 2020. The Boolean operators “AND” and “OR” as well as the following search terms were used to collect papers that specifically dealt with the ecological evaluation of transitional waters: (“biotic assessment” OR “biotic index*” OR “biological indices” OR “ecological index*” OR “ecological indices” OR “foraminifera-based index” OR “foraminiferal index*” OR “foraminifera-based indices”) AND (“ecological quality” OR “ecological status” OR “ecological quality status” OR “ecosystem health”) AND (“foraminifera*” OR “foraminiferal” OR “foram” or “forams”) AND (“transitional waters” OR “naturally stressed environments” OR “fjord” OR “fjords” “estuaries” OR “brackish” OR “coastal” OR “lagoon” OR “Delta” OR “Ria”). The addition of quotation marks and asterisks (*) were used on certain search terms such as foraminifera to ensure the search terms are returned exactly as written and increase the precision of the search.
The resulting publications were manually filtered and screened by a single author, and once compiled, the table was checked and approved by the remaining authors. Publications found not to focus on foraminifera-based methods were excluded, as well as those concerning the study of waters that do not fall under the definition of transitional such as coral reefs (unless specified to be nearshore).
Information including the (i) the stressor/environmental parameter studied, (ii) the source of the stressor, (iii) the study site or location, (iv) the type of assemblage (living, fossil, or recent but not living), (v) the index used, and (vi) reference (first author and year) was compiled into Table A1.
We later concluded that the table lacked some key studies. This was due to the WoS repository containing papers primarily from high-impact journals only. As a result, the table was supplemented with articles referenced elsewhere in this publication, which included the search criteria in the title or topic.
The information extracted from each paper concerns only the methods used in each study rather than the outcome observed, and hence, is unlikely to be affected by bias as results, conclusions, and efficacy are not discussed.
The tabulation of the information extracted from each publication was kept as simple as possible and repeated where possible in the same order, so it is easier to identify studies that have used the same index.
Appendix A.2. Results
Using these search strategies, a final yield of 41 articles, the oldest being published in 2009 and the latest 2021. After filtering, seven publications were removed due to not meeting the specified criteria, for example, “Is untreated sewage impacting coral reefs of Cay Caulker, Belize” by Emrich et al. 2017 was excluded due to its focus on coral reefs, which do not fall strictly under the definition of transitional waters.
An additional four publications that met the specific search criteria were included from those referenced elsewhere in this paper (these have been marked with an asterisk next to the reference).
The table has been arranged by grouping firstly those that employ diversity indices, then sensitivity, and lastly, test abnormality or accumulation rates.
Table A1.
Examples of application of benthic foraminifera as BQEs in environmental studies over the last 20 years across a wide range of environmental stress and in various types of transitional and coastal waters.
Table A1.
Examples of application of benthic foraminifera as BQEs in environmental studies over the last 20 years across a wide range of environmental stress and in various types of transitional and coastal waters.
| Stressor/Environmental Parameters | Source of Stress | Study Site | Assemblage Type | Index Used | Reference |
|---|---|---|---|---|---|
| Salinity, oxygen, turbulence, and organic matter inputs | Natural environmental variation | Po coastal plain, Italy | Fossil | Diversity Index Exp (H’bc) | Barbieri et al. 2020 [145] |
| Organic enrichment | Various sources | Mediterranean Sea Coast, France | Living | Diversity indices, wall structure proportion, and sp. tolerance sensitivity | Barras et al. 2014 [50] |
| Chemical pollution: polychlorobiphenyls (PCBs), polycyclic aromatic hydrocarbons (PAHs), barium (Ba), and mercury (Hg) | industries, petrochemical pole | Augusta Harbor, Eastern Sicily, Italy | Living, dead (1960s) | The Foraminiferal Size Index (FSI) and the Lost Species Index (LSI), Sp. richness, diversity, composition, foraminiferal number (FN), and H’ index | Bergamin and Romano et al. 2016 [146] |
| Oxygen depletion, grain size, C/N, TOC, and total nitrogen | Industries and urbanization | Norwegian Skagerrak coastal waters and Fjords, NE North Sea | Living, dead | Exp (H’bc) Diversity Index | Bouchet et al. 2012 [74] |
| Variety of pollution types | Aquaculture/Sewage | Lagoons and lakes, Italy | Living | Exp (H’bc) Diversity Index | Bouchet et al. 2018b [49] |
| Metal pollution, organic enrichment, pollution load index, sediment pollution index | Industries, urbanization, natural variation | Marambaia Cove of the Sepetiba Bay, SE Brazil | Living, fossil | Foram Stress Index (FSI) and Exp (H’bc) Diversity Endex | Castelo et al. 2021 [76] |
| Palaeo-environmental hydrological regime | Natural regime variation | Po Delta, Italy | Living, fossil | Epifaunal vs. Infaunal, Shannon–Wiener Index (H’log 2) Index, Fisher evenness and dominance, indicative groups | Dasgupta et al. 2020 [146] |
| Organic enrichment, metal pollution | Urbanization, industries, shipping activities, domestic pollution | Santos Estuary, Brazil | Living, fossil | Diversity Index Exp(H’bc) based, abundance, diversity, normalized ecological quality ratio (nEQR) | De Jesus et al. 2020 [25] |
| POP and metal pollution | Industries, shipping activities | Hammerfest harbor, Norway | Living | Exp (H’bc) Diversity Index | Dijkstra et al. 2017 * [61] |
| Organic enrichment, metal pollution | Urbanization, industries | Oslofjord, Norway | Fossil, dead | Shannon–Wiener Index (H’ log 2) and the Hurlbert’s Index (ES100), and cluster analyses | Dolven et al. 2013 * [19] |
| Oxygen depletion, salinity fluctuation, temperature, metal pollution | Various Sources | Høgsfjord and Lysefjord, Norway | Living, fossil | Shannon–Wiener Index (H’ log 2) and the Hurlbert’s Index (ES100) | Duffield et al. 2017 * [147] |
| Organic enrichment, metal pollution, phosphates | Industries, urbanization | Gulf of Gabes, Tunisia | Living | “Long versus short life span” index (ILS) and the modified FoRAM-Index (FI’), benthic foraminiferal assemblage composition and diversity, e.g., the Tolerant Species Index (TSstd), the Foram Stress Index (FSI), the Diversity Index Exp(H’bc) and the Newly Developed Foram-AZTI Marine Biotic Index (Foram-AMBI) | El Kateb et al. 2020 [48] |
| Grain size, metal pollution, organic enrichment | Industries, metallurgic factories, shipping activities, urbanization | Hauts de France, English Channel, France | Living | Diversity Exp (H’bc), foraminiferal density (FD), abundance, and indicative groups | Francescangeli et al. 2020 [78] |
| Oxygen depletion | Urbanization, sewage | Bay of Trieste, Italy | Living | Exp (H’bc) Diversity Index | Melis et al. 2019 [80] |
| organic enrichment, metal pollution | Industries, urbanization | Idefjord, Sweden/Norway | Living, fossil | Fisher Alpha Diversity Index, individual rarefaction ES(100), dominance indices (based on Simpsons Index) | Polovodova Asteman et al. 2015 * [24] |
| Chemical pollutants, Oxygen depletion, eutrophication, acidification | Various sources | Bellingham Bay, Washington State, USA | Contemporary | The Shannon Index (H’), Diversity Index | Nesbitt et al. 2014 [148] |
| Oxygen depletion, pH | Industrial effluents | Uppanar Estuary, Tamil Nadu Coast, India | Living | FoRAMFoRAM-Index (FI) ranging, the Foram Stress Index (FSI), Ammonia-Elphidium Index (AEI), indicative groups, abundance, and diversity, species richness | Nagendra and Reddy et al. 2019 [149] |
| pH, temperature | Urbanization, tourism | Zanzibar Archipelago | Living | The FoRAMFoRAM-Index and species richness patterns, Fisher α, | Thissen and Langer et al. 2017 [150] |
| Organic enrichment | Industries, urbanization | Skagerrak coast, Norway | Living, fossil | Foram-AMBI Sensitivity/Tolerance Index with Shannon’s diversity (H′log2) | Alve et al. 2019 [75] |
| Organic enrichment | Cultural eutrophication (Various anthropogenic sources) | Northeast Atlantic and Arctic fjords | Living, fossil | Foram-AMBI Sensitivity/Tolerance Index with Shannon’s diversity (H′log2) | Alve et al. 2016 [94] |
| Organic enrichment, potentially toxic elements, oil | Industries, urbanization | Guanabara Bay, Brazil | Living | Ammonia-Elphidium Index, Foram Stress Index | Alves Martins et al. 2020 [151] |
| Organic enrichment | Oyster Farming | Transitional Waters, English Channel/European Atlantic coast and the Mediterranean Sea | Living | Foram-AMBI Sensitivity/Tolerance Index | Bouchet et al. 2021 [82] |
| Dissolved oxygen, organic enrichment, grain size, total nitrogen, pigments, and depth below threshold | Various sources | 11 Fjords on the Skagerrak Coast, SE Norway | Living | Cluster/indicative species groups, comparison to macrofaunal changes | Bouchet et al. 2018 [81] |
| Grain size (pollutant proxy) | Industries, urbanization, docks, natural variation | 3 Sardinian Coastal areas | Living | Species’ richness, foraminiferal density, and the Fisher-α Diversity Index | Buosi et al. 2013 [152] |
| Organic enrichment, phosphates | Aquaculture (fish farming) | Monastir Bay, Tunisia | Living | AZTI Marine Biotic Index, Foram-AMBI | Damak et al. 2020 [153] |
| Organic enrichment, metal pollution | Domestic and industrial effluents, aquaculture, tourism | Saronikos Gulf, Greece | Living | Foram Stress Index (FSI), tolerance/sensitivity | Dimiza et al. 2016 [88] |
| Low oxygen, nutrient enrichment (eutrophication), run-off | Various sources | Society Archipelago, French Polynesia | Contemporary | Species composition, species richness, patterns of distribution, abundance, indicative groups, Fisher α, FoRAM-Index (FI) | Fajemila et al. 2015 [154] |
| Organic enrichment, metal pollution | Various sources | The Gulf of Manfredonia, southern Adriatic Sea | Living | Foram-AMBI Sensitivity Index and Foram Stress Index (FSI) | Fossile et al. 2021 [155] |
| Organic enrichment, metal pollution | Industries | Boulogne sur Mer Harbor, France | Fossil | Cluster/indicative species groups | Francescangeli et al. 2016 [22] |
| increased nutrient and sediment inputs | Urbanization, tourism | Great Barrier Reef, Australia | Living, fossil | Functional groups, Foraminifera in Reef Assessment and Monitoring (FoRAM) Index (FI) | Johnson et al. 2019 [156] |
| Organic enrichment, metal pollution | Aquaculture (fish farming) | Øksfjord, Norway | Living, fossil | Norwegian Quality Index (fNQI), AZTI’s Marine Biotic Index (fAMBI), fHlog2, ES100, absolute abundances and relative abundances of ecological groups (EGs) | Klootwijk et al. 2021 [26] |
| Metal pollution, oxygen depletion | Urbanization, industries, shipping activities, domestic pollution | Strait of Malacca, Malaysia | Living | Foraminifera Stress Index (FSI) | Minhat et al. 2020 [157] |
| Organic enrichment, nitrogen, carbonate content, phosphorus, mineralogy, grain size | Urbanization, tourism | Abrolhos, Southwest Atlantic, eastern Brazil | Living | Functional Groups for FoRAMFoRAM-Index (FI) | Oliveira-Silva et al. 2012 [158] |
| Organic enrichment | Urbanization, sewage | Levantine basin, Mediterranean | Living | TSI-Med and Foram-AMBI | Parent et al. 2021 [99] |
| Organic enrichment, metal pollution | Trawling, fishing activities, urbanization | Kosterfjord, Sweden | Living, dead | Foram-AMBI and NQIforam Indices | Polovodova Asteman et al. 2021 [45] |
| Hypoxia, metal pollution | Urbanization, industries | Western Coastal Area of Jakarta Bay | Living, fossil | The Ammonia-Elphidium Index, abnormal morphology | Rositasari et al. 2018 [159] |
| Metal pollution | Thermal sources and agricultural activities in | Gulf of Edremit, northeastern Aegean Sea, Turkey | Living, fossil | Abnormal morphology, species diversity | Yümün and Önce et al. 2017 [160] |
| Metal pollution, Organic enrichment, Oxygen depletion | Shipping activities, industries | Horten Inner Harbor, Norway | Living, fossil | Benthic foraminiferal accumulation rate (BFAR) | Hess et al. 2020 [20] |
Table A2.
Two paleo-EcoQS reconstructions based on diversity index Exp (H’bc) applied to the same foraminiferal assemblage data set from site I02-10B in the outer part of the Idefjord [23]. EcoQS threshold values (displayed below) for reconstruction EcoQS 1 are taken from Bouchet et al. [49], and for EcoQS 2 from Bouchet et al. [74], respectively. Years marked with asterisk (*) are interpolated ages based on the average sedimentation rate. EcoQS threshold values (found below) for EcoQS 1 from Bouchet et al. [49], and for EcoQS 2 from Bouchet et al. [74].
Table A2.
Two paleo-EcoQS reconstructions based on diversity index Exp (H’bc) applied to the same foraminiferal assemblage data set from site I02-10B in the outer part of the Idefjord [23]. EcoQS threshold values (displayed below) for reconstruction EcoQS 1 are taken from Bouchet et al. [49], and for EcoQS 2 from Bouchet et al. [74], respectively. Years marked with asterisk (*) are interpolated ages based on the average sedimentation rate. EcoQS threshold values (found below) for EcoQS 1 from Bouchet et al. [49], and for EcoQS 2 from Bouchet et al. [74].
| Depth | EcoQs 1 | EcoQs 2 | Year | ||
|---|---|---|---|---|---|
| 0–1 | 7.71 | 7.71 | 2002,0 | ||
| 2–3 | 4.1 | 4.1 | 1998,0 | ||
| 4–5 | 4.58 | 4.58 | 1994,1 | ||
| 6–7 | 5.5 | 5.5 | 1990,1 | ||
| 8–9 | 4.71 | 4.71 | 1986,1 | ||
| 10–11 | 4.72 | 4.72 | 1982,2 | ||
| 12–13 | 4.96 | 4.96 | 1978,2 | ||
| 14–15 | 6.73 | 6.73 | 1974,2 | ||
| 16–17 | 6.27 | 6.27 | 1970,3 | ||
| 18–19 | 5.71 | 5.71 | 1966,3 | ||
| 20–21 | 4.59 | 4.59 | 1962,3 | ||
| 22–23 | 6.37 | 6.37 | 1958,3 | ||
| 24–25 | 6.12 | 6.12 | 1951,2 | ||
| 26–27 | 6.52 | 6.52 | 1944,1 | ||
| 28–29 | 7.52 | 7.52 | 1936,9 | ||
| 30–31 | 7.45 | 7.45 | 1924.4 * | ||
| 32–33 | 9.15 | 9.15 | 1911.9 * | ||
| 34–35 | 9.06 | 9.06 | 1899,4 * | ||
| 36–37 | 9.59 | 9.59 | 1886,9 * | ||
| 38–39 | 6.4 | 6.4 | 1874,4 * | ||
| 40–41 | 8.05 | 8.05 | 1861,9 * | ||
| 42–44 | 8.51 | 8.51 | 1849,4 * | ||
| 44–45 | 5.44 | 5.44 | 1836,9 * | ||
| 46–47.7 | 7.4 | 7.4 | 1824,4 * | ||
| High | Good | Moderate | Bad | Poor | |
| EcoQS 1 | >15 | 11–15 | 7–11 | 3–7 | <3 |
| EcoQS 2 | >10 | 7.5–10 | 5–7.5 | 2.5–5 | <2.5 |
References
- European Commission, Water framework directive 2000/60/EC. Off. J. Eur. Communities 2000, L 269, 1–15.
- Howarth, W. The marine strategy framework directive. J. Water Law 2008, 19, 95–97. [Google Scholar]
- Reizopolou, S.; Penna, M.; Boix, D.; Buchet, R.; Costas, N.; Derolez, V.; Gascon, S.; Gifre, J.; Martinoy, M.; Pardo, I.; et al. Transitional Waters Mediterranean Geographic Intercalibration Group: Benthic Invertebrates Fauna Ecological Assessment Methods, EUR 29561 EN; Publications Office of the European Union: Luxembourg, 2018. [CrossRef]
- Andersen, M.M. An innovation system approach to eco-innovation—Aligning policy rationales. In Proceedings of the Greening of Policies, Interlinkages and Policy Integration Conference, Berlin, Germany, 3–4 December 2004; pp. 1–28. [Google Scholar]
- Borja, A.; Franco, J.; Muxika, I. Classification tools for marine ecological quality assessment: The usefulness of macro-benthic communities in an area affected by a submarine outfall. In Proceedings of the ICES CM 2003/Session J-02, Tallinn, Estonia, 24–28 September 2003; pp. 1–10. [Google Scholar]
- Blanchet, H.; Lavesque, N.; Ruellet, T.; Dauvin, J.; Sauriau, P.-G.; Desroy, N.; Desclaux, C.; Leconte, M.; Bachelet, G.; Janson, A.-L.; et al. Use of biotic indices in semi-enclosed coastal ecosystems and transitional waters habitats—Implications for the implementation of the European Water Framework Directive. Ecol. Indic. 2008, 8, 360–372. [Google Scholar] [CrossRef] [Green Version]
- Bouchet, V.M.; Sauriau, P.-G. Influence of oyster culture practices and environmental conditions on the ecological status of intertidal mudflats in the Pertuis Charentais (SW France): A multi-index approach. Mar. Pollut. Bull. 2008, 56, 1898–1912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salas, F.; Neto, J.; Borja, A.; Marques, J. Evaluation of the applicability of a marine biotic index to characterize the status of estuarine ecosystems: The case of Mondego estuary (Portugal). Ecol. Indic. 2004, 4, 215–225. [Google Scholar] [CrossRef] [Green Version]
- Elliott, M.; Quintino, V. The estuarine quality paradox, environmental homeostasis and the difficulty of detecting anthropogenic stress in naturally stressed areas. Mar. Pollut. Bull. 2007, 54, 640–645. [Google Scholar] [CrossRef]
- Munari, C.; Mistri, M. Biodiversity of soft-sediment benthic communities from Italian transitional waters. J. Biogeogr. 2008, 35, 1622–1637. [Google Scholar] [CrossRef]
- Pusceddu, A.; Dell’Anno, A.; Danovaro, R.; Manini, E.; Sarà, G.; Fabiano, M. Enzymatically hydrolyzable protein and carbohydrate sedimentary pools as indicators of the trophic state of detritus sink systems: A case study in a Mediterranean coastal lagoon. Estuaries 2003, 26, 641–650. [Google Scholar] [CrossRef]
- Dauvin, J.-C. Paradox of estuarine quality: Benthic indicators and indices, consensus or debate for the future. Mar. Pollut. Bull. 2007, 55, 271–281. [Google Scholar] [CrossRef]
- Dauvin, J.-C.; Ruellet, T. The estuarine quality paradox: Is it possible to define an ecological quality status for specific modified and naturally stressed estuarine ecosystems? Mar. Pollut. Bull. 2009, 59, 38–47. [Google Scholar] [CrossRef]
- Muniz, P.; Venturini, N.; Pires-Vanin, A.M.; Tommasi, L.R.; Borja, Á. Testing the applicability of a Marine Biotic Index (AMBI) to assessing the ecological quality of soft-bottom benthic communities, in the South America Atlantic region. Mar. Pollut. Bull. 2005, 50, 624–637. [Google Scholar] [CrossRef] [PubMed]
- Quintino, V.; Elliott, M.; Rodrigues, A.M. The derivation, performance and role of univariate and multivariate indicators of benthic change: Case studies at differing spatial scales. J. Exp. Mar. Biol. Ecol. 2006, 330, 368–382. [Google Scholar] [CrossRef]
- McLusky, D.S.; Elliott, M. Transitional waters: A new approach, semantics or just muddying the waters? Estuar. Coast. Shelf Sci. 2007, 71, 359–363. [Google Scholar] [CrossRef]
- Halpern, B.S.; Walbridge, S.; Selkoe, K.A.; Kappel, C.V.; Micheli, F.; D’Agrosa, C.; Bruno, J.F.; Casey, K.S.; Ebert, C.; Fox, H.E.; et al. A global map of human impact on marine ecosystems. Science 2008, 319, 948–953. [Google Scholar] [CrossRef] [Green Version]
- Van Hoey, G.; Borja, A.; Birchenough, S.; Buhl-Mortensen, L.; Degraer, S.; Fleischer, D.; Kerckhof, F.; Magni, P.; Muxika, I.; Reiss, H.; et al. The use of benthic indicators in Europe: From the water framework directive to the marine strategy framework directive. Mar. Pollut. Bull. 2010, 60, 2187–2196. [Google Scholar] [CrossRef] [Green Version]
- Dolven, J.K.; Alve, E.; Rygg, B.; Magnusson, J. Defining past ecological status and in situ reference conditions using benthic foraminifera: A case study from the Oslofjord, Norway. Ecol. Indic. 2013, 29, 219–233. [Google Scholar] [CrossRef]
- Hess, S.; Alve, E.; Andersen, T.J.; Joranger, T. Defining ecological reference conditions in naturally stressed environments—How difficult is it? Mar. Environ. Res. 2020, 156. [Google Scholar] [CrossRef]
- Alve, E.; Lepland, A.; Magnusson, J.; Backer-Owe, K. Monitoring strategies for re-establishment of ecological reference conditions: Possibilities and limitations. Mar. Pollut. Bull. 2009, 59, 297–310. [Google Scholar] [CrossRef]
- Alve, E. Foraminifera, climatic change, and pollution: A study of late Holocene sediments in Drammensfiord, southeast Norway. Holocene 1991, 1, 243–261. [Google Scholar] [CrossRef]
- Francescangeli, F.; du Chatelet, E.A.; Billon, G.; Trentesaux, A.; Bouchet, V. Palaeo-ecological quality status based on foraminifera of Boulogne-sur-Mer harbour (Pas-de-Calais, Northeastern France) over the last 200 years. Mar. Environ. Res. 2016, 117, 32–43. [Google Scholar] [CrossRef]
- Polovodova Asteman, I.P.; Hanslik, D.; Nordberg, K. An almost completed pollution-recovery cycle reflected by sediment geochemistry and benthic foraminiferal assemblages in a Swedish–Norwegian Skagerrak fjord. Mar. Pollut. Bull. 2015, 95, 126–140. [Google Scholar] [CrossRef]
- De Jesus, M.S.D.; Frontalini, F.; Bouchet, V.M.P.; Yamashita, C.; Sartoretto, J.R.; Figueira, R.C.L.; Sousa, S.H. Reconstruction of the palaeo-ecological quality status in an impacted estuary using benthic foraminifera: The Santos Estuary (Sao Paulo state, SE Brazil). Mar. Environ. Res. 2020, 162. [Google Scholar] [CrossRef]
- Klootwijk, A.T.; Alve, E.; Hess, S.; Renaud, P.E.; Sørlie, C.; Dolven, J.K. Monitoring environmental impacts of fish farms: Comparing reference conditions of sediment geochemistry and benthic foraminifera with the present. Ecol. Indic. 2021, 120, 106818. [Google Scholar] [CrossRef]
- Resig, J.M. Foraminiferal Ecology around Ocean Outfalls off Southern California; Pergamon Press: London, UK, 1960. [Google Scholar]
- Kenigsberg, C.; Abramovich, S.; Hyams-Kaphzan, O. The effect of long-term brine discharge from desalination plants on benthic foraminifera. PLoS ONE 2020, 15, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Aloulou, F.; Elleuch, B.; Kallel, M. Benthic foraminiferal assemblages as pollution proxies in the northern coast of Gabes Gulf, Tunisia. Environ. Monit. Assess. 2012, 184, 777–795. [Google Scholar] [CrossRef]
- Brunner, C.A.; Yeager, K.M.; Hatch, R.; Simpson, S.; Keim, J.; Briggs, K.B.; Louchouarn, P. Effects of oil from the 2010 Macondo well blowout on marsh foraminifera of Mississippi and Louisiana, USA. Environ. Sci. Technol. 2013, 47, 9115–9123. [Google Scholar] [CrossRef]
- Morvan, J.; Le Cadre, V.; Jorissen, F.; Debenay, J.-P. Foraminifera as potential bio-indicators of the “Erika” oil spill in the Bay of Bourgneuf: Field and experimental studies. Aquat. Living Resour. 2004, 17, 317–322. [Google Scholar] [CrossRef]
- Young, C.M.; Schwing, P.T.; Cotton, L.J. Benthic foraminiferal morphological response to the 2010 Deepwater Horizon oil spill. Mar. Micropaleontol. 2021, 101971. [Google Scholar] [CrossRef]
- Bouchet, V.M.; Debenay, J.-P.; Sauriau, P.-G.; Radford-Knoery, J.; Soletchnik, P. Effects of short-term environmental disturbances on living benthic foraminifera during the Pacific oyster summer mortality in the Marennes-Oléron Bay (France). Mar. Environ. Res. 2007, 64, 358–383. [Google Scholar] [CrossRef] [Green Version]
- Oron, S.; Angel, D.; Goodman-Tchernov, B.; Merkado, G.; Kiflawi, M.; Abramovich, S. Benthic foraminiferal response to the removal of aquaculture fish cages in the Gulf of Aqaba-Eilat, Red Sea. Mar. Micropaleontol. 2014, 107, 8–17. [Google Scholar] [CrossRef]
- Burone, L.; Valente, P.; Pires-Vanin, A.M.S.; De Melloe Sousa, S.H.; Mahiques, M.M.; Braga, E. Benthic foraminiferal variability on a monthly scale in a subtropical bay moderately affected by urban sewage. Sci. Mar. 2007, 71, 775–792. [Google Scholar] [CrossRef] [Green Version]
- Hyams-Kaphzan, O.; Almogi-Labin, A.; Benjamini, C.; Herut, B. Natural oligotrophy vs. pollution-induced eutrophy on the SE Mediterranean shallow shelf (Israel): Environmental parameters and benthic foraminifera. Mar. Pollut. Bull. 2009, 58, 1888–1902. [Google Scholar] [CrossRef]
- Debenay, J.-P.; Tsakiridis, E.; Soulard, R.; Grossel, H. Factors determining the distribution of foraminiferal assemblages in Port Joinville Harbor (Ile d’Yeu, France): The influence of pollution. Mar. Micropaleontol. 2001, 43, 75–118. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Colón, M.; Hallock, P.; Green-Ruíz, C.R.; Smoak, J.M. Benthic foraminifera as bioindicators of potentially toxic element (PTE) pollution: Torrecillas lagoon (San Juan Bay Estuary), Puerto Rico. Ecol. Indic. 2018, 89, 516–527. [Google Scholar] [CrossRef]
- Smith, C.W.; Goldstein, S.T. The effects of selected heavy metal elements (arsenic, cadmium, nickel, zinc) on experimentally grown foraminiferal assemblages from Sapelo Island, Georgia and Little Duck Key, Florida, U.S.A. J. Foraminifer. Res. 2019, 49, 303–317. [Google Scholar] [CrossRef]
- Hart, M.B.; Molina, G.S.; Smart, C.W. Estuarine foraminifera from South West England: Impact of metal pollution in a mining heritage area. J. Sediment. Environ. 2020, 5, 1–16. [Google Scholar] [CrossRef]
- Schafer, C.T.; Collins, E.S.; Smith, J.N. Relationship of Foraminifera and thecamoebian distributions to sediments contaminated by pulp mill effluent: Saguenay Fiord, Quebec, Canada. Mar. Micropaleontol. 1991, 17, 255–283. [Google Scholar] [CrossRef]
- Alve, E. Benthic foraminiferal distribution and recolonization of formerly anoxic environments in Drammensfjord, Southern Norway. Mar. Micropaleontol. 1995, 25, 169–186. [Google Scholar] [CrossRef]
- Scott, D.B.; Tobin, R.; Williamson, M.; Medioli, F.S.; Latimer, J.S.; Boothman, W.A.; Asioli, A.; Haury, V. Pollution monitoring in two North American estuaries: Historical reconstructions using benthic foraminifera. J. Foraminifer. Res. 2005, 35, 65–82. [Google Scholar] [CrossRef]
- Hayward, B.W.; Grenfell, H.R.; Nicholson, K.; Parker, R.; Wilmhurst, J.; Horrocks, M.; Swales, A.; Sabaa, A.T. Foraminiferal record of human impact on intertidal estuarine environments in New Zealand’s largest city. Mar. Micropaleontol. 2004, 53, 37–66. [Google Scholar] [CrossRef]
- Polovodova Asteman, I.; Van Nieuwenhove, N.; Andersen, T.J.; Linders, T.; Nordberg, K. Recent environmental change in the Kosterhavet National Park marine protected area as reflected by hydrography and sediment proxy data. Mar. Environ. Res. 2021, 166. [Google Scholar] [CrossRef] [PubMed]
- Alve, E. Benthic foraminiferal evidence of environmental change in the Skagerrak over the past six decades. NGU Bull. 1996, 430, 85–93. [Google Scholar]
- Nordberg, K.; Asteman, I.P.; Gallagher, T.M.; Robijn, A. Recent oxygen depletion and benthic faunal change in shallow areas of Sannäs Fjord, Swedish west coast. J. Sea Res. 2017, 127, 46–62. [Google Scholar] [CrossRef]
- El Kateb, A.; Stalder, C.; Martínez-Colón, M.; Mateu-Vicens, G.; Francescangeli, F.; Coletti, G.; Stainbank, S.; Spezzaferri, S. Foraminiferal-based biotic indices to assess the ecological quality status of the Gulf of Gabes (Tunisia): Present limitations and future perspectives. Ecol. Indic. 2020, 111, 105962. [Google Scholar] [CrossRef]
- Bouchet, V.; Goberville, E.; Frontalini, F. Benthic foraminifera to assess ecological quality statuses in Italian transitional waters. Ecol. Indic. 2018, 84, 130–139. [Google Scholar] [CrossRef]
- Barras, C.; Jorissen, F.; Labrune, C.; Andral, B.; Boissery, P. Live benthic foraminiferal faunas from the French Mediterranean Coast: Towards a new biotic index of environmental quality. Ecol. Indic. 2014, 36, 719–743. [Google Scholar] [CrossRef] [Green Version]
- Linke, P.; Lutze, G. Microhabitat preferences of benthic foraminifera—A static concept or a dynamic adaptation to optimize food acquisition? Mar. Micropaleontol. 1993, 20, 215–234. [Google Scholar] [CrossRef] [Green Version]
- Yanko, V.; Kronfeld, J.; Flexer, A. Response of benthic foraminifera to various pollution sources; implications for pollution monitoring. J. Foraminifer. Res. 1994, 24, 1–17. [Google Scholar] [CrossRef]
- Samir, A.; El-Din, A. Benthic foraminiferal assemblages and morphological abnormalities as pollution proxies in two Egyptian bays. Mar. Micropaleontol. 2001, 41, 193–227. [Google Scholar] [CrossRef]
- Polovodova, I.; Schönfeld, J. Foraminiferal test abnormalities in the western Baltic Sea. J. Foraminifer. Res. 2008, 38, 318–336. [Google Scholar] [CrossRef]
- Arenillas, I.; Arz, J.A.; Gilabert, V. Blooms of aberrant planktic foraminifera across the K/Pg boundary in the Western Tethys: Causes and evolutionary implications. Paleobiology 2018, 44, 460–489. [Google Scholar] [CrossRef] [Green Version]
- Nardelli, M.; Malferrari, D.; Ferretti, A.; Bartolini, A.; Sabbatini, A.; Negri, A. Zinc incorporation in the miliolid foraminifer Pseudotriloculina rotunda under laboratory conditions. Mar. Micropaleontol. 2016, 126, 42–49. [Google Scholar] [CrossRef]
- Boehnert, S.; Birkelund, A.; Schmiedl, G.; Kuhnert, H.; Kuhn, G.; Hass, H.; Hebbeln, D. Test deformation and chemistry of foraminifera as response to anthropogenic heavy metal input. Mar. Pollut. Bull. 2020, 155, 111112. [Google Scholar] [CrossRef]
- Alve, E.; Bernhard, J. Vertical migratory response of benthic foraminifera to controlled oxygen concentrations in an experimental mesocosm. Mar. Ecol. Prog. Ser. 1995, 116, 137–151. [Google Scholar] [CrossRef]
- Asteman, I.P.; Nordberg, K. A short note on a present-day benthic recovery status in the formerly heavily polluted Idefjord (Sweden/Norway). Mar. Pollut. Bull. 2017, 123, 227–231. [Google Scholar] [CrossRef]
- Belart, P.; Clemente, I.; Raposo, D.; Habib, R.; Volino, E.; Villar, A.; Alves, M.; Fontana, L.; Lorini, M.; Panigai, G.; et al. Living and dead foraminifera as bioindicators in Saquarema lagoon system, Brazil. Lat. Am. J. Aquat. Res. 2018, 46, 1055–1072. [Google Scholar] [CrossRef]
- Dijkstra, N.; Junttila, J.; Skirbekk, K.; Carroll, J.; Husum, K.; Hald, M. Benthic foraminifera as bio-indicators of chemical and physical stressors in Hammerfest harbor (Northern Norway). Mar. Pollut. Bull. 2017, 114, 384–396. [Google Scholar] [CrossRef] [Green Version]
- du Châtelet, E.A.; Debenay, J.-P.; Soulard, R. Foraminiferal proxies for pollution monitoring in moderately polluted harbors. Environ. Pollut. 2004, 127, 27–40. [Google Scholar] [CrossRef]
- Nordberg, K. Decreasing oxygen concentrations in the Gullmar Fjord, Sweden, as confirmed by benthic foraminifera, and the possible association with NAO. J. Mar. Syst. 2000, 23, 303–316. [Google Scholar] [CrossRef]
- Polovodova, I.; Nikulina, A.; Schönfeld, J.; Dullo, W.-C. Recent benthic foraminifera in the Flensburg Fjord (Western Baltic Sea). J. Micropalaeontol. 2009, 28, 131–142. [Google Scholar] [CrossRef] [Green Version]
- Nikulina, A.; Polovodova, I.; Schönfeld, J. Foraminiferal response to environmental changes in Kiel Fjord, SW Baltic Sea. eEarth 2008, 3, 37–49. [Google Scholar] [CrossRef] [Green Version]
- Thomas, E.; Gapotchenko, T.; Varekamp, J.C.; Mecray, E.L.; Buchholtz ten Brink, M.R. Benthic foraminifera and environmental changes in Long Island Sound. J. Coast. Res. 2000, 16, 641–655. [Google Scholar]
- Bernhard, J.; Alve, E. Survival, ATP pool, and ultrastructural characterization of benthic foraminifera from Drammensfjord (Norway): Response to anoxia. Mar. Micropaleontol. 1996, 28, 5–17. [Google Scholar] [CrossRef]
- Risgaard-Petersen, N.; Langezaal, A.M.; Ingvardsen, S.; Schmid, M.C.; Jetten, M.S.M.; Camp, H.O.D.; Derksen, J.W.M.; Piña-Ochoa, E.; Eriksson, S.P.; Nielsen, L.P.; et al. Evidence for complete denitrification in a benthic foraminifer. Nature 2006, 443, 93–96. [Google Scholar] [CrossRef]
- Gustafsson, M.; Nordberg, K. Benthic foraminifera and their response to hydrography, periodic hypoxic conditions and primary production in the Koljö fjord on the Swedish west coast. J. Sea Res. 1999, 41, 163–178. [Google Scholar] [CrossRef]
- Bradshaw, J.S. Laboratory studies on the rate of growth of the foraminifer, “Streblus beccarii (Linné) var. tepida (Cushman)”. J. Paleontol. 1957, 31, 1138–1147. [Google Scholar]
- Ross, B.J.; Hallock, P. Survival and recovery of the foraminifer Amphistegina gibbosa and associated diatom endosymbionts following up to 20 months in aphotic conditions. Mar. Micropaleontol. 2019, 149, 35–43. [Google Scholar] [CrossRef]
- Ross, B.J.; Hallock, P. Dormancy in the foraminifera: A review. J. Foraminifer. Res. 2016, 46, 358–368. [Google Scholar] [CrossRef]
- Schönfeld, J.; Alve, E.; Geslin, E.; Jorissen, F.; Korsun, S.; Spezzaferri, S. The FOBIMO (FOraminiferal BIo-MOnitoring) initiative—Towards a standardised protocol for soft-bottom benthic foraminiferal monitoring studies. Mar. Micropaleontol. 2012, 94–95, 1–13. [Google Scholar] [CrossRef]
- Bouchet, V.; Alve, E.; Rygg, B.; Telford, R. Benthic foraminifera provide a promising tool for ecological quality assessment of marine waters. Ecol. Indic. 2012, 23, 66–75. [Google Scholar] [CrossRef]
- Alve, E.; Hess, S.; Bouchet, V.; Dolven, J.K.; Rygg, B. Intercalibration of benthic foraminiferal and macrofaunal biotic indices: An example from the Norwegian Skagerrak coast (NE North Sea). Ecol. Indic. 2019, 96, 107–115. [Google Scholar] [CrossRef]
- Dimiza, M.D.; Triantaphyllou, M.V.; Koukousioura, O.; Hallock, P.; Simboura, N.; Karageorgis, A.P.; Papathanasiou, E. The Foram Stress Index: A new tool for environmental assessment of soft-bottom environments using benthic foraminifera. A case study from the Saronikos Gulf, Greece, Eastern Mediterranean. Ecol. Indic. 2016, 60, 611–621. [Google Scholar] [CrossRef]
- Alve, E.; Korsun, S.; Schönfeld, J.; Dijkstra, N.; Golikova, E.; Hess, S.; Husum, K.; Panieri, G. Foram-AMBI: A sensitivity index based on benthic foraminiferal faunas from North-East Atlantic and Arctic fjords, continental shelves and slopes. Mar. Micropaleontol. 2016, 122, 1–12. [Google Scholar] [CrossRef]
- Castelo, W.F.L.; Martins, M.V.A.; Martinez-Colon, M.; Guerra, J.V.; Dadalto, T.P.; Terroso, D.; Soares, M.F.; Frontalini, F.; Duleba, W.; Socorro, O.A.A.; et al. Disentangling natural vs. anthropogenic induced environmental variability during the Holocene: Marambaia Cove, SW sector of the Sepetiba Bay (SE Brazil). Environ. Sci. Pollut. Res. 2021, 28. [Google Scholar] [CrossRef]
- Punniyamoorthy, R.; Sarathy, P.P.; Mahadevan, G.; Selvaraj, P.; Bharathidasan, V.; Murugesan, P. Benthic foraminifera to assess ecological quality status of Kaduvaiyar and Uppanar estuaries, Southeast coast of India. J. Mar. Biol. Assoc. India 2020, 61, 52–62. [Google Scholar] [CrossRef]
- Francescangeli, F.; Quijada, M.; du Châtelet, E.A.; Frontalini, F.; Trentesaux, A.; Billon, G.; Bouchet, V. Multidisciplinary study to monitor consequences of pollution on intertidal benthic ecosystems (Hauts de France, English Channel, France): Comparison with natural areas. Mar. Environ. Res. 2020, 160. [Google Scholar] [CrossRef]
- Bouchet, V.M.; Deldicq, N.; Baux, N.; Dauvin, J.-C.; Pezy, J.-P.; Seuront, L.; Méar, Y. Benthic foraminifera to assess ecological quality statuses: The case of salmon fish farming. Ecol. Indic. 2020, 117. [Google Scholar] [CrossRef]
- Melis, R.; Celio, M.; Bouchet, V.; Varagona, G.; Bazzaro, M.; Crosera, M.; Pugliese, N. Seasonal response of benthic foraminifera to anthropogenic pressure in two stations of the Gulf of Trieste (northern Adriatic Sea, Italy): The marine protected area of Miramare versus the Servola water sewage outfall. Mediterr. Mar. Sci. 2019, 20, 120–141. [Google Scholar] [CrossRef]
- Bouchet, V.M.P.; Telford, R.J.; Rygg, B.; Oug, E.; Alve, E. Can benthic foraminifera serve as proxies for changes in benthic macrofaunal community structure? Implications for the definition of reference conditions. Mar. Environ. Res. 2018, 137, 24–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouchet, V.M.P.; Frontalini, F.; Francescangeli, F.; Sauriau, P.-G.; Geslin, E.; Martins, M.V.A.; Almogi-Labin, A.; Avnaim-Katav, S.; Di Bella, L.; Cearreta, A.; et al. Indicative value of benthic foraminifera for biomonitoring: Assignment to ecological groups of sensitivity to total organic carbon of species from European intertidal areas and transitional waters. Mar. Pollut. Bull. 2021, 164. [Google Scholar] [CrossRef] [PubMed]
- Armynot du Châtelet, E.A.; Bout-Roumazeilles, V.; Riboulleau, A.; Trentesaux, A. Sediment (grain size and clay mineralogy) and organic matter quality control on living benthic foraminifera. Rev. Micropaléontol. 2009, 52, 75–84. [Google Scholar] [CrossRef]
- Leorri, E.; Zimmerman, A.R.; Mitra, S.; Christian, R.R.; Fatela, F.; Mallinson, D.J. Refractory organic matter in coastal salt marshes-effect on C sequestration calculations. Sci. Total Environ. 2018, 633, 391–398. [Google Scholar] [CrossRef]
- Jorissen, F.; Nardelli, M.P.; Almogi-Labin, A.; Barras, C.; Bergamin, L.; Bicchi, E.; El Kateb, A.; Ferraro, L.; McGann, M.; Morigi, C.; et al. Developing Foram-AMBI for biomonitoring in the Mediterranean: Species assignments to ecological categories. Mar. Micropaleontol. 2018, 140, 33–45. [Google Scholar] [CrossRef] [Green Version]
- Mojtahid, M.; Jorissen, F.; Durrieu, J.; Galgani, F.; Howa, H.; Redois, F.; Camps, R. Benthic foraminifera as bio-indicators of drill cutting disposal in tropical east Atlantic outer shelf environments. Mar. Micropaleontol. 2006, 61, 58–75. [Google Scholar] [CrossRef] [Green Version]
- Denoyelle, M.; Jorissen, F.J.; Martin, D.; Galgani, F.; Miné, J. Comparison of benthic foraminifera and macrofaunal indicators of the impact of oil-based drill mud disposal. Mar. Pollut. Bull. 2010, 60, 2007–2021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hallock, P.; Lidz, B.H.; Cockey-Burkhard, E.M.; Donnelly, K.B. Foraminifera as bioindicators in coral reef assessment and monitoring: The FORAM index. Environ. Monit. Assess. 2003, 81, 221–238. [Google Scholar] [CrossRef]
- Prazeres, M.; Martínez-Colón, M.; Hallock, P. Foraminifera as bioindicators of water quality: The FoRAM Index revisited. Environ. Pollut. 2020, 257. [Google Scholar] [CrossRef] [PubMed]
- Roberts, J.M.; Davies, A.J.; Henry, L.A.; Dodds, L.A.; Duineveld, G.C.A.; Lavaleye, M.S.S.; Maier, C.; Van Soest, R.W.M.; Bergman, M.J.N.; Hühnerbach, V.; et al. Mingulay reef complex: An interdisciplinary study of cold-water coral habitat, hydrography and biodiversity. Mar. Ecol. Prog. Ser. 2009, 397, 139–151. [Google Scholar] [CrossRef] [Green Version]
- Parent, B. Développement d’un Indice Biotique Basé sur les Foraminiféres Benthiques: Application sur la Façade Méditerranéenne Française. Ph.D. Thesis, University of Angers, Angers, France, 2019. [Google Scholar]
- Thompson, B.; Weisberg, S.B.; Melwani, A.; Lowe, S.; Ranasinghe, J.A.; Cadien, D.B.; Dauer, D.M.; Diaz, R.J.; Fields, W.; Kellogg, M.; et al. Low levels of agreement among experts using best professional judgment to assess benthic condition in the San Francisco Estuary and Delta. Ecol. Indic. 2012, 12, 167–173. [Google Scholar] [CrossRef]
- Zettler, M.L.; Proffitt, C.E.; Darr, A.; Degraer, S.; Devriese, L.; Greathead, C.; Kotta, J.; Magni, P.; Martin, G.; Reiss, H.; et al. On the myths of indicator species: Issues and further consideration in the use of static concepts for ecological applications. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [Green Version]
- Simboura, N.; Zenetos, A. Benthic indicators to use in Ecological Quality classification of Mediterranean soft bottom marine ecosystems, including a new Biotic Index. Mediterr. Mar. Sci. 2002, 3, 77–111. [Google Scholar] [CrossRef] [Green Version]
- Grall, J.; Glémarec, M. Using biotic indices to estimate macrobenthic community perturbations in the Bay of Brest. Estuar. Coast. Shelf Sci. 1997, 44, 43–53. [Google Scholar] [CrossRef]
- Borja, A.; Franco, J.; Pérez, V. A marine biotic index to establish the ecological quality of soft-bottom benthos within European estuarine and coastal environments. Mar. Pollut. Bull. 2000, 40, 1100–1114. [Google Scholar] [CrossRef]
- Parent, B.; Hyams-Kaphzan, O.; Barras, C.; Lubinevsky, H.; Jorissen, F. Testing foraminiferal environmental quality indices along a well-defined organic matter gradient in the Eastern Mediterranean. Ecol. Indic. 2021, 125, 107498. [Google Scholar] [CrossRef]
- Bergamin, L.; Di Bella, L.; Ferraro, L.; Frezza, V.; Pierfranceschi, G.; Romano, E. Benthic foraminifera in a coastal marine area of the eastern Ligurian Sea (Italy): Response to environmental stress. Ecol. Indic. 2019, 96, 16–31. [Google Scholar] [CrossRef]
- Martínez-Colón, M.; Bouchet, V. Benthic Foraminifera as Ecological Sentinels of Marine Systems Health. SCOR Working Group Proposal: FORAM-ECO. 2020. Available online: https://scor-int.org/wp-content/uploads/2020/05/FORAM-ECO_SCOR_v7.pdf (accessed on 6 April 2021).
- Sousa, S.H.M.; Members of the BIOFOM Group; Yamashita, C.; Semensatto, D.L.; Santarosa, A.C.A.; Iwai, F.S.; Omachi, C.Y.; Disaró, S.T.; Martins, M.V.A.; Barbosa, C.F.; et al. Opportunities and challenges in incorporating benthic foraminifera in marine and coastal environmental biomonitoring of soft sediments: From science to regulation and practice. J. Sediment. Environ. 2020, 5. [Google Scholar] [CrossRef]
- Zeppilli, D.; Sarrazin, J.; Leduc, D.; Arbizu, P.M.; Fontaneto, D.; Fontanier, C.; Gooday, A.J.; Kristensen, R.M.; Ivanenko, V.; Sørensen, M.; et al. Is the meiofauna a good indicator for climate change and anthropogenic impacts? Mar. Biodivers. 2015, 45, 505–535. [Google Scholar] [CrossRef] [Green Version]
- Mateu-Vicens, G.; Khokhlova, A.; Sebastián-Pastor, T. Epiphytic foraminiferal indices as bioindicators in Mediterranean seagrass meadows. J. Foraminifer. Res. 2014, 44, 325–339. [Google Scholar] [CrossRef]
- Refcond. Guidance on Establishing Reference Conditions and Ecological Status Class Boundaries for Inland Surface Waters; Produced by Working Group 2.31, Reference Conditions for Inland Surface (REFCOND), Common Implementation Strategy; European Commission: Brussels, Belgium, 2003; p. 86. [Google Scholar]
- Rombouts, I.; Beaugrand, G.; Artigas, L.F.; Dauvin, J.-C.; Gevaert, F.; Goberville, E.; Kopp, D.; Lefebvre, S.; Luczak, C.; Spilmont, N.; et al. Evaluating marine ecosystem health: Case studies of indicators using direct observations and modelling methods. Ecol. Indic. 2013, 24, 353–365. [Google Scholar] [CrossRef]
- Ruellet, T.; Dauvin, J.-C. Benthic indicators: Analysis of the threshold values of ecological quality classifications for transitional waters. Mar. Pollut. Bull. 2007, 54, 1707–1714. [Google Scholar] [CrossRef] [PubMed]
- Berkeley, A.; Perry, C.T.; Smithers, S.G.; Horton, B.P.; Taylor, K.G. A review of the ecological and taphonomic controls on foraminiferal assemblage development in intertidal environments. Earth-Sci. Rev. 2007, 83, 205–230. [Google Scholar] [CrossRef]
- Murray, J.W.; Alve, E.; Cundy, A. The origin of modern agglutinated foraminiferal assemblages: Evidence from a stratified fjord. Estuar. Coast. Shelf Sci. 2003, 58, 677–697. [Google Scholar] [CrossRef]
- Murray, J.W.; Alve, E. Natural dissolution of modern shallow water benthic foraminifera: Taphonomic effects on the palaeoecological record. Palaeogeogr. Palaeoclim. Palaeoecol. 1999, 146, 195–209. [Google Scholar] [CrossRef]
- Murray, J.W.; Alve, E. Taphonomic experiments on marginal marine foraminiferal assemblages: How much ecological information is preserved? Palaeogeogr. Palaeoclim. Palaeoecol. 1999, 149, 183–197. [Google Scholar] [CrossRef]
- Francescangeli, F.; Portela, M.; Du Chatelet, E.A.; Billon, G.; Andersen, T.; Bouchet, V.; Trentesaux, A. Infilling of the Canche Estuary (Eastern English Channel, France): Insight from benthic foraminifera and historical pictures. Mar. Micropaleontol. 2018, 142, 1–12. [Google Scholar] [CrossRef]
- Mulik, J.; Sukumaran, S.; Srinivas, T.; Vijapure, T. Comparative efficacy of benthic biotic indices in assessing the Ecological Quality Status (EcoQS) of the stressed Ulhas estuary, India. Mar. Pollut. Bull. 2017, 120, 192–202. [Google Scholar] [CrossRef]
- Josefson, A.B.; Blomqvist, M.; Hansen, J.L.; Rosenberg, R.; Rygg, B. Assessment of marine benthic quality change in gradients of disturbance: Comparison of different Scandinavian multi-metric indices. Mar. Pollut. Bull. 2009, 44, 1689–1699. [Google Scholar] [CrossRef]
- Borja, A.; Josefson, A.B.; Miles, A.; Muxika, I.; Olsgard, F.; Phillips, G.; Rodríguez, J.G.; Rygg, B. An approach to the intercalibration of benthic ecological status assessment in the North Atlantic ecoregion, according to the European Water Framework Directive. Mar. Pollut. Bull. 2007, 55, 42–52. [Google Scholar] [CrossRef]
- Pawlowski, J.; Kelly-Quinn, M.; Altermatt, F.; Apothéloz-Perret-Gentil, L.; Beja, P.; Boggero, A.; Borja, A.; Bouchez, A.; Cordier, T.; Domaizon, I.; et al. The future of biotic indices in the ecogenomic era: Integrating (e)DNA metabarcoding in biological assessment of aquatic ecosystems. Sci. Total. Environ. 2018, 637–638, 1295–1310. [Google Scholar] [CrossRef]
- Baird, D.J.; Hajibabaei, M. Biomonitoring 2.0: A new paradigm in ecosystem assessment made possible by next-generation DNA sequencing. Mol. Ecol. 2012, 21, 2039–2044. [Google Scholar] [CrossRef] [PubMed]
- Cordier, T.; Frontalini, F.; Cermakova, K.; Apothéloz-Perret-Gentil, L.; Treglia, M.; Scantamburlo, E.; Bonamin, V.; Pawlowski, J. Multi-marker eDNA metabarcoding survey to assess the environmental impact of three offshore gas platforms in the North Adriatic Sea (Italy). Mar. Environ. Res. 2019, 146, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Cordier, T.; Alonso-Sáez, L.; Apothéloz-Perret-Gentil, L.; Aylagas, E.; Bohan, D.A.; Bouchez, A.; Chariton, A.; Creer, S.; Frühe, L.; Keck, F.; et al. Ecosystems monitoring powered by environmental genomics: A review of current strategies with an implementation roadmap. Mol. Ecol. 2020, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Pawlowski, J.; Levin, L.; Recio-Blanco, X.; Le, J.; Lejzerowicz, F.; Cordier, T.; Mulsow, S.; Gjerde, K.; Mengerink, K.; Gooday, A.J. Enabling the Use of Environmental DNA for Regulatory Policymaking in the Marine Industrial Revolution; SNIS Project Working Paper; SNIS: Sydney, Australia, 2018; pp. 1–20. [Google Scholar]
- Forster, D.; Dunthorn, M.; Mahé, F.; Dolan, J.; Audic, S.; Bass, D.; Bittner, L.; Boutte, C.; Christen, R.; Claverie, J.-M.; et al. Benthic protists: The under-charted majority. FEMS Microbiol. Ecol. 2016, 92, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, V.; Carvalho, G.R.; Sung, W.; Johnson, H.F.; Power, D.; Neill, S.; Packer, M.; Blaxter, M.; Lambshead, P.J.D.; Thomas, W.K.; et al. Second-generation environmental sequencing unmasks marine metazoan biodiversity. Nat. Commun. 2010, 1. [Google Scholar] [CrossRef] [Green Version]
- Keeley, N.; Wood, S.A.; Pochon, X. Development and preliminary validation of a multi-trophic metabarcoding biotic index for monitoring benthic organic enrichment. Ecol. Indic. 2018, 85, 1044–1057. [Google Scholar] [CrossRef]
- Laroche, O.; Wood, S.A.; Tremblay, L.A.; Ellis, J.; Lear, G.; Pochon, X. A cross-taxa study using environmental DNA/RNA metabarcoding to measure biological impacts of offshore oil and gas drilling and production operations. Mar. Pollut. Bull. 2018, 127, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Mauffrey, F.; Cordier, T.; Apothéloz-Perret-Gentil, L.; Cermakova, K.; Merzi, T.; Delefosse, M.; Blanc, P.; Pawlowski, J. Benthic monitoring of oil and gas offshore platforms in the North Sea using environmental DNA metabarcoding. Mol. Ecol. 2020, 1–16. [Google Scholar] [CrossRef]
- Pawlowski, J.; Esling, P.; Lejzerowicz, F.; Cedhagen, T.; Wilding, T.A. Environmental monitoring through protist next-generation sequencing metabarcoding: Assessing the impact of fish farming on benthic foraminifera communities. Mol. Ecol. Resour. 2014, 14, 1129–1140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavaliere, M.; Angeles, I.B.; Montresor, M.; Bucci, C.; Brocani, L.; Balassi, E.; Margiotta, F.; Francescangeli, F.; Bouchet, V.; Pawlowski, J.; et al. Assessing the ecological quality status of the highly polluted Bagnoli area (Tyrrhenian Sea, Italy) using foraminiferal eDNA metabarcoding. Sci. Total. Environ. 2021, 790, 147871. [Google Scholar] [CrossRef]
- Frontalini, F.; Cordier, T.; Balassi, E.; du Chatelet, E.A.; Cermakova, K.; Apothéloz-Perret-Gentil, L.; Martins, M.V.A.; Bucci, C.; Scantamburlo, E.; Treglia, M.; et al. Benthic foraminiferal metabarcoding and morphology-based assessment around three offshore gas platforms: Congruence and complementarity. Environ. Int. 2020, 144, 106049. [Google Scholar] [CrossRef]
- Apothéloz-Perret-Gentil, L.; Cordonier, A.; Straub, F.; Iseli, J.; Esling, P.; Pawlowski, J. Taxonomy-free molecular diatom index for high-throughput eDNA biomonitoring. Mol. Ecol. Resour. 2017, 17, 1231–1242. [Google Scholar] [CrossRef]
- Boere, A.C.; Rijpstra, W.I.C.; De Lange, G.; Damsté, J.S.S.; Coolen, M.J.L. Preservation potential of ancient plankton DNA in Pleistocene marine sediments. Geobiology 2011, 9, 377–393. [Google Scholar] [CrossRef] [Green Version]
- Lejzerowicz, F.; Esling, P.; Majewski, W.; Szczuciński, W.; Decelle, J.; Obadia, C.; Martinez-Arbizu, P.; Pawlowski, J. Ancient DNA complements microfossil record in deep-sea subsurface sediments. Biol. Lett. 2013, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pawłowska, J.; Zajaczkowski, M.; Łącka, M.; Lejzerowicz, F.; Esling, P.; Pawlowski, J. Palaeoceanographic changes in Hornsund Fjord (Spitsbergen, Svalbard) over the last millennium: New insights from ancient DNA. Clim. Past 2016, 12, 1459–1472. [Google Scholar] [CrossRef] [Green Version]
- Vuillemin, A.; Horn, F.; Alawi, M.; Henny, C.; Wagner, D.; Crowe, S.A.; Kallmeyer, J. Preservation and significance of extracellular DNA in ferruginous sediments from Lake Towuti, Indonesia. Front. Microbiol. 2017, 8, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Pawłowska, J.; Wollenburg, J.E.; Zajączkowski, M.; Pawlowski, J. Planktonic foraminifera genomic variations reflect paleoceanographic changes in the Arctic: Evidence from sedimentary ancient DNA. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Turner, C.R.; Uy, K.L.; Everhart, R.C. Fish environmental DNA is more concentrated in aquatic sediments than surface water. Biol. Conserv. 2015, 183, 93–102. [Google Scholar] [CrossRef] [Green Version]
- Sakata, M.K.; Yamamoto, S.; Gotoh, R.O.; Miya, M.; Yamanaka, H.; Minamoto, T. Sedimentary eDNA provides different information on timescale and fish species composition compared with aqueous eDNA. Environ. DNA 2020, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Kirkpatrick, J.B.; Walsh, E.A.; D’Hondt, S. Fossil DNA persistence and decay in marine sediment over hundred-thousand-year to million-year time scales. Geology 2016, 44, 615–618. [Google Scholar] [CrossRef] [Green Version]
- Holman, L.E.; De Bruyn, M.; Creer, S.; Carvalho, G.; Robidart, J.; Rius, M. Detection of introduced and resident marine species using environmental DNA metabarcoding of sediment and water. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borja, Á.; Elliott, M.; Carstensen, J.; Heiskanen, A.-S.; van de Bund, W. Marine management—Towards an integrated implementation of the European Marine Strategy Framework and the Water Framework Directives. Mar. Pollut. Bull. 2010, 60, 2175–2186. [Google Scholar] [CrossRef] [PubMed]
- Mueller, M.; Geist, J. Conceptual guidelines for the implementation of the ecosystem approach in biodiversity monitoring. Ecosphere 2016, 7, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Armbrecht, L. The potential of sedimentary ancient DNA to reconstruct past ocean ecosystems. Oceanography 2020, 33, 116–123. [Google Scholar] [CrossRef]
- Gibson, J.; Shokralla, S.; Porter, T.M.; King, I.; van Konynenburg, S.; Janzen, D.H.; Hallwachs, W.; Hajibabaei, M. Simultaneous assessment of the macrobiome and microbiome in a bulk sample of tropical arthropods through DNA metasystematics. Proc. Natl. Acad. Sci. USA 2014, 111, 8007–8012. [Google Scholar] [CrossRef] [Green Version]
- Ruppert, K.M.; Kline, R.J.; Rahman, S. Past, present, and future perspectives of environmental DNA (eDNA) metabarcoding: A systematic review in methods, monitoring, and applications of global eDNA. Glob. Ecol. Conserv. 2019, 17, e00547. [Google Scholar] [CrossRef]
- Li, K.; Rollins, J.; Yan, E. Web of Science use in published research and review papers 1997–2017: A selective, dynamic, cross-domain, content-based analysis. Scientometrics 2018, 115, 1–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbieri, G.; Rossi, V.; Ghosh, A.; Vaiani, S.C. Conservation paleobiology as a tool to define reference conditions in naturally stressed transitional settings: Micropaleontological insights from the Holocene of the Po coastal plain (Italy). Water 2020, 12, 3420. [Google Scholar] [CrossRef]
- Bergamin, L.; Romano, E. Suitable sediment fraction for paleoenvironmental reconstruction and assessment of contaminated coastal areas based on benthic foraminifera: A case study from Augusta Harbour (Eastern Sicily, Italy). Ecol. Indic. 2016, 71, 66–78. [Google Scholar] [CrossRef]
- Duffield, C.; Alve, E.; Andersen, N.; Andersen, T.; Hess, S.; Strohmeier, T. Spatial and temporal organic carbon burial along a fjord to coast transect: A case study from Western Norway. Holocene 2017, 27, 1325–1339. [Google Scholar] [CrossRef] [Green Version]
- Nesbitt, E.A.; Martin, R.A.; Martin, D.E.; Apple, J. Rapid deterioration of sediment surface habitats in Bellingham Bay, Washington State, as indicated by benthic foraminifera. Mar. Pollut. Bull. 2015, 97, 273–284. [Google Scholar] [CrossRef]
- Nagendra, R.; Reddy, A.N. benthic foraminifera response to ecosystem pollution in the Uppanar estuary, Tamil Nadu coast, India. J. Geol. Soc. India 2019, 93, 555–566. [Google Scholar] [CrossRef]
- Thissen, J.M.; Langer, M.R. Spatial patterns and structural composition of foraminiferal assemblages from the Zanzibar Archipelago (Tanzania). Palaeontogr. Abt. A 2017, 308, 1–67. [Google Scholar] [CrossRef]
- Martins, M.V.A.; Hohenegger, J.; Martínez-Colón, M.; Frontalini, F.; Bergamashi, S.; Laut, L.; Belart, P.; Mahiques, M.; Pereira, E.; Rodrigues, R.; et al. Ecological quality status of the NE sector of the Guanabara Bay (Brazil): A case of living benthic foraminiferal resilience. Mar. Pollut. Bull. 2020, 158. [Google Scholar] [CrossRef] [PubMed]
- Buosi, C.; Cherchi, A.; Ibba, A.; Marras, B.; Marrucci, A.; Schintu, M. Preliminary data on benthic foraminiferal assemblages and sedimentological characterisation from some polluted and unpolluted coastal areas of Sardinia (Italy). Boll. Soc. Paleontol. Ital. 2013, 52, 35–44. [Google Scholar]
- Damak, M.; Fourati, R.; Elleuch, B.; Kallel, M. Environmental quality assessment of the fish farms’ impact in the Monastir Bay (eastern of Tunisia, Central Mediterranean): A benthic foraminiferal perspective. Environ. Sci. Pollut. Res. 2020, 27, 9059–9074. [Google Scholar] [CrossRef]
- Fajemila, O.T.; Langer, M.R.; Lipps, J. Spatial Patterns in the distribution, diversity and abundance of benthic foraminifera around Moorea (Society Archipelago, French Polynesia). PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fossile, E.; Sabbatini, A.; Spagnoli, F.; Caridi, F.; Dell’Anno, A.; De Marco, R.; Dinelli, E.; Droghini, E.; Tramontana, M.; Negri, A. Sensitivity of foraminiferal-based indices to evaluate the ecological quality status of marine coastal benthic systems: A case study of the Gulf of Manfredonia (southern Adriatic Sea). Mar. Pollut. Bull. 2021, 163, 111933. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.; Perry, C.; Smithers, S.; Morgan, K.; Woodroffe, S. Reef shallowing is a critical control on benthic foraminiferal assemblage composition on nearshore turbid coral reefs. Palaeogeogr. Palaeoclim. Palaeoecol. 2019, 533. [Google Scholar] [CrossRef]
- Minhat, F.I.; Shaari, H.; Razak, N.S.A.; Satyanarayana, B.; Saelan, W.N.W.; Yusoff, N.M.; Husain, M.L. Evaluating performance of foraminifera stress index as tropical-water monitoring tool in Strait of Malacca. Ecol. Indic. 2020, 111. [Google Scholar] [CrossRef]
- Oliveira-Silva, P.; Barbosa, C.; De Almeida, C.M.; Seoane, J.C.S.; Cordeiro, R.; Turcq, B.J.; Soares-Gomes, A. Sedimentary geochemistry and foraminiferal assemblages in coral reef assessment of Abrolhos, Southwest Atlantic. Mar. Micropaleontol. 2012, 94–95, 14–24. [Google Scholar] [CrossRef]
- Rositasari, R.; Puspitasari, R.; Budiyanto, F.; Lestari, L. Ecological changes over a century in the western coastal area of Jakarta Bay: Based on a short core sample. In Proceedings of the 2nd International Conference on Chemistry, Chemical Process and Engineering, Yogyakarta, Indonesia, 14–15 August 2018; Fatimah, I., Kawasaki, H., Kamari, A., Chuenchom, L., Kurniawan, S.M.A., Sahroni, I., Musawwa, M., Eds.; AIP Publishing: Melville, NY, USA, 2018; Volume 2026. [Google Scholar]
- Yümün, Z.Ü.; Önce, M. Monitoring heavy metal pollution in foraminifera from the Gulf of Edremit (northeastern Aegean Sea) between Izmir, Balıkesir and Çanakkale (Turkey). J. Afr. Earth Sci. 2017, 130, 110–124. [Google Scholar] [CrossRef]
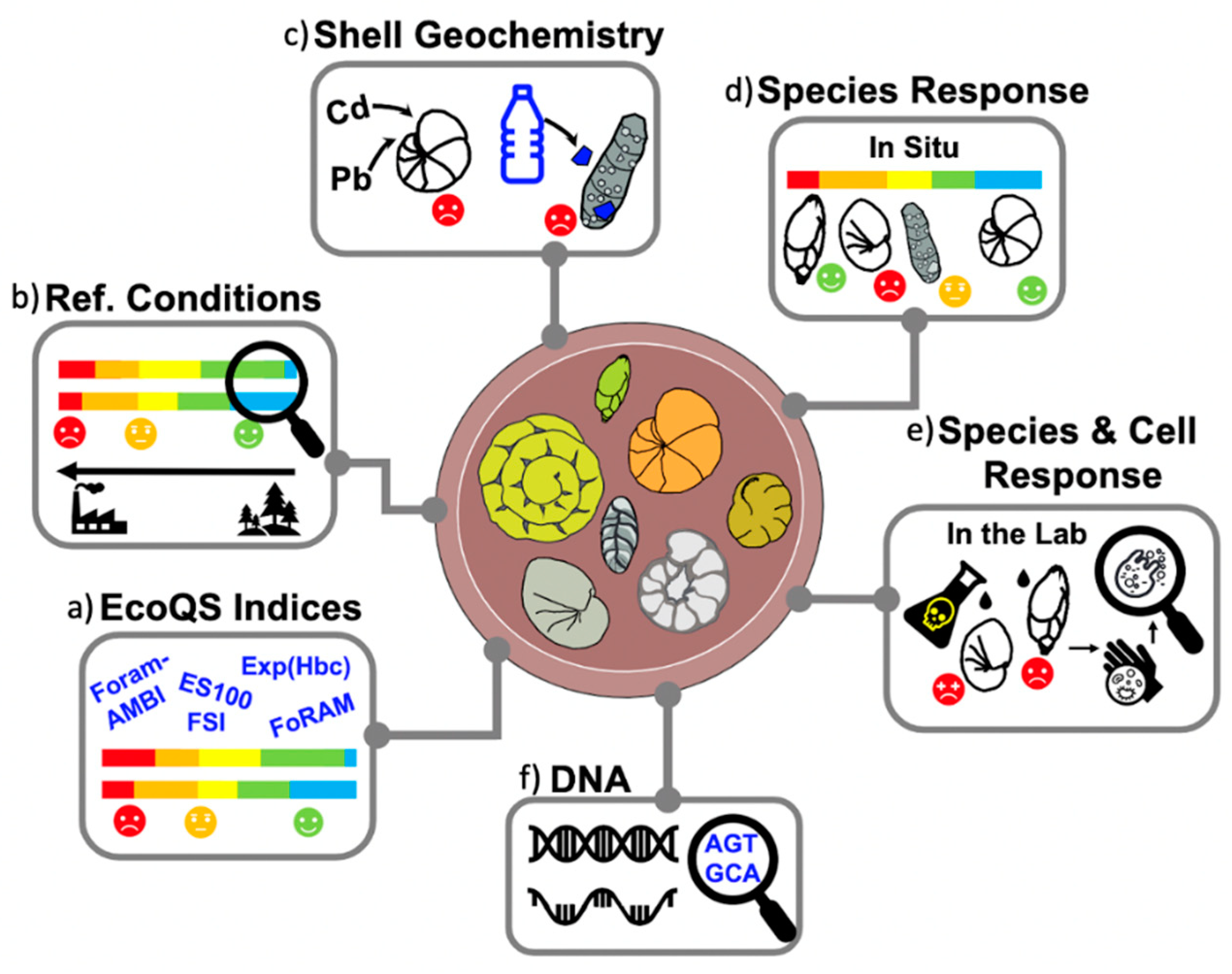
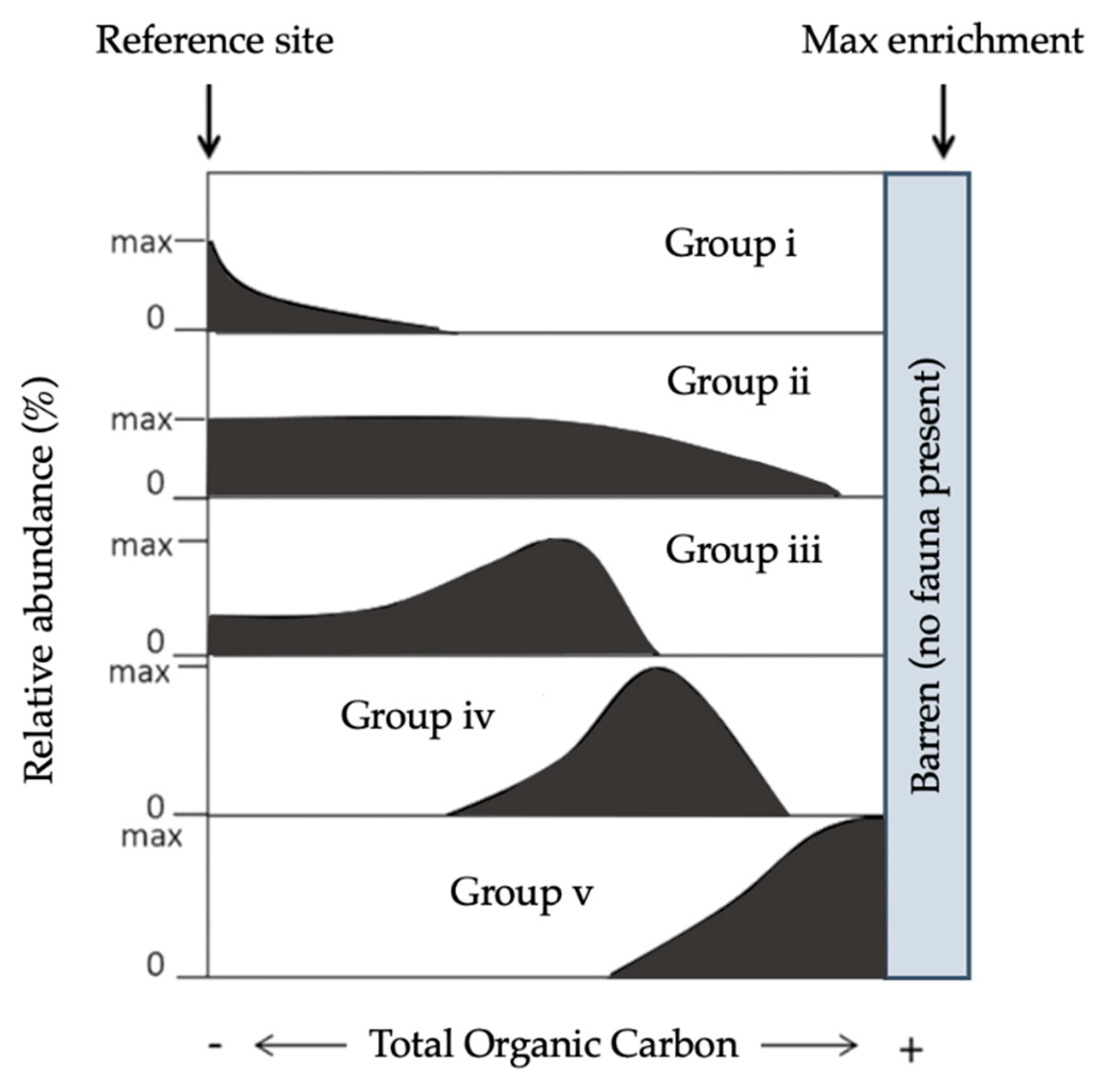
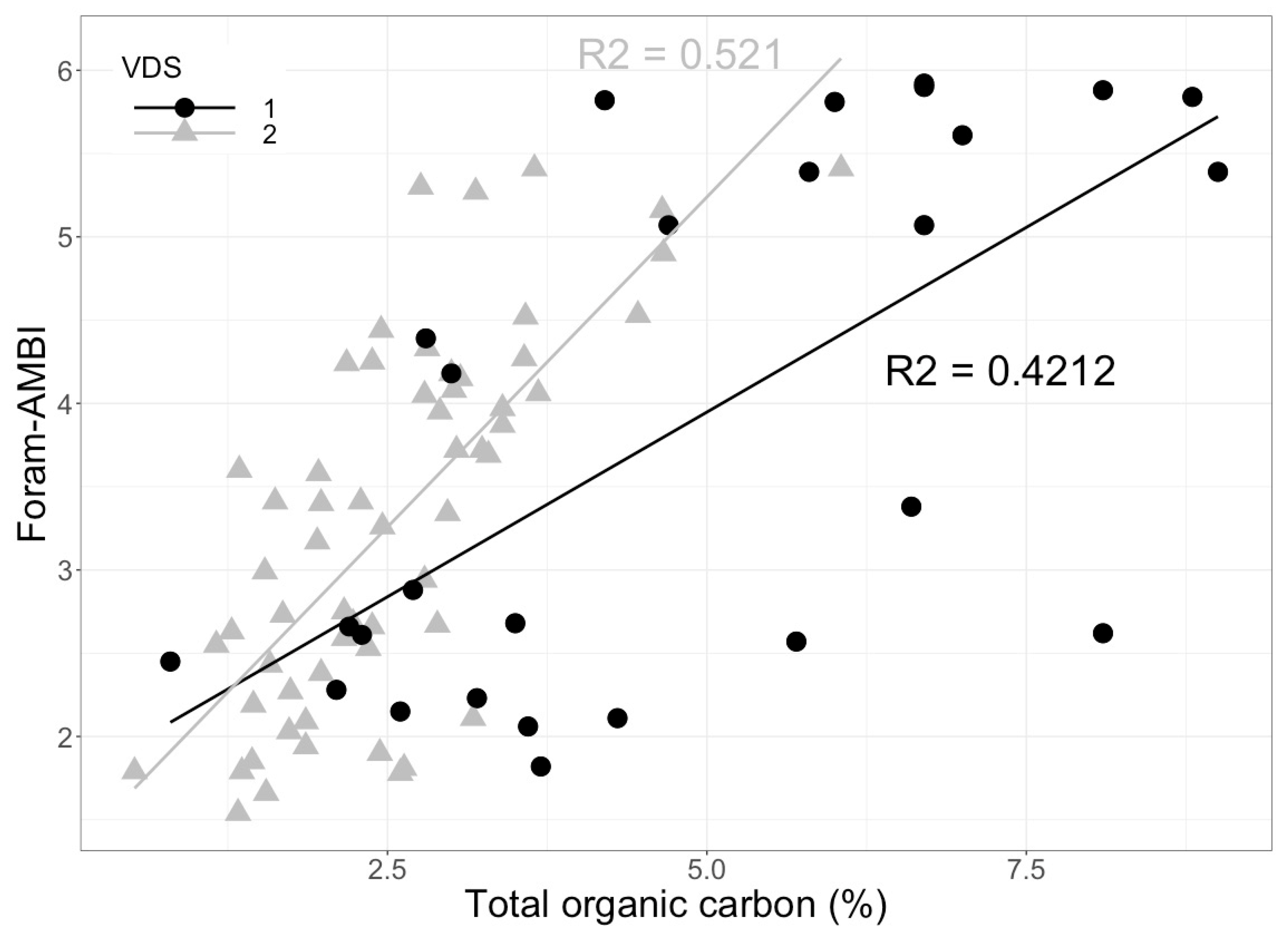
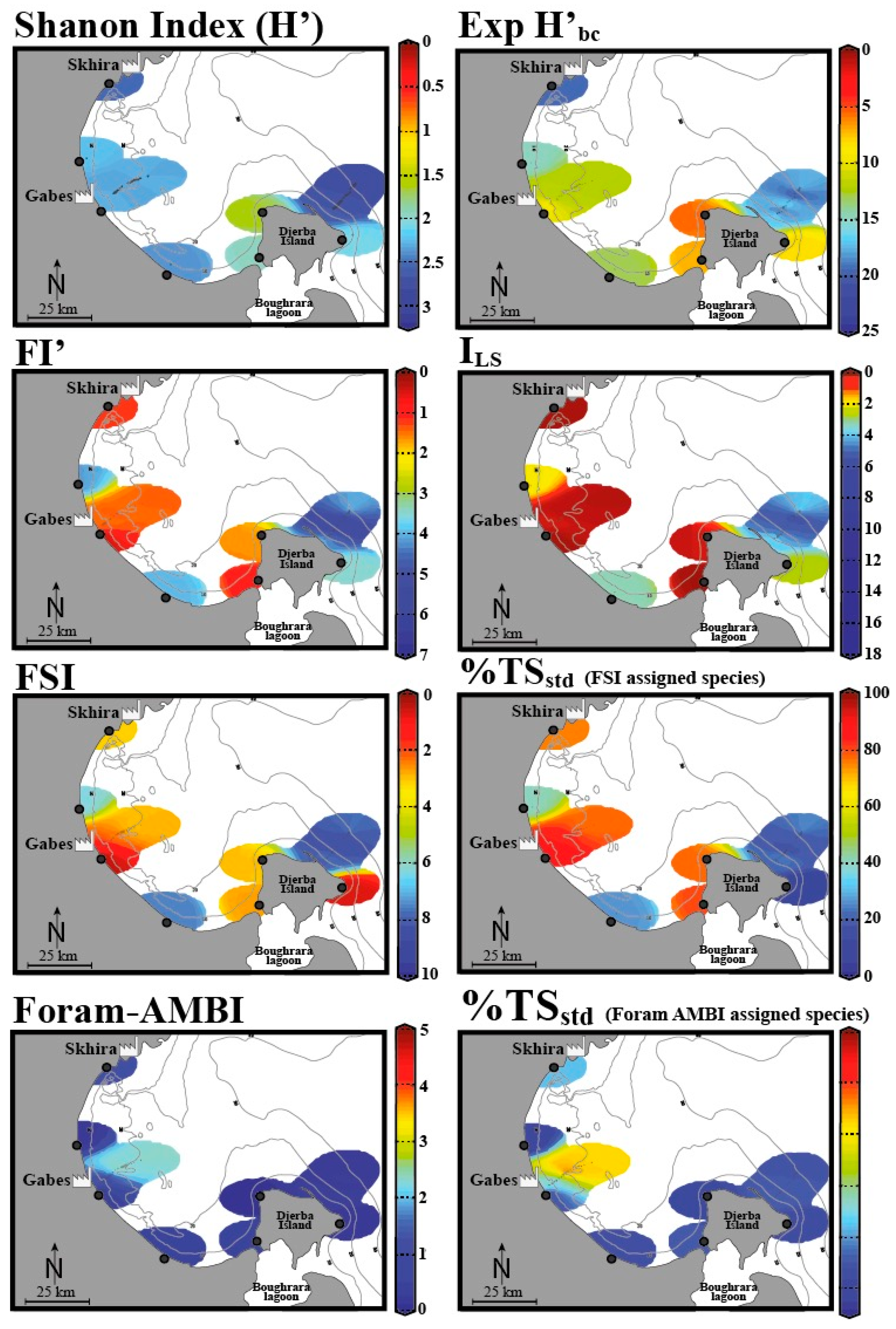
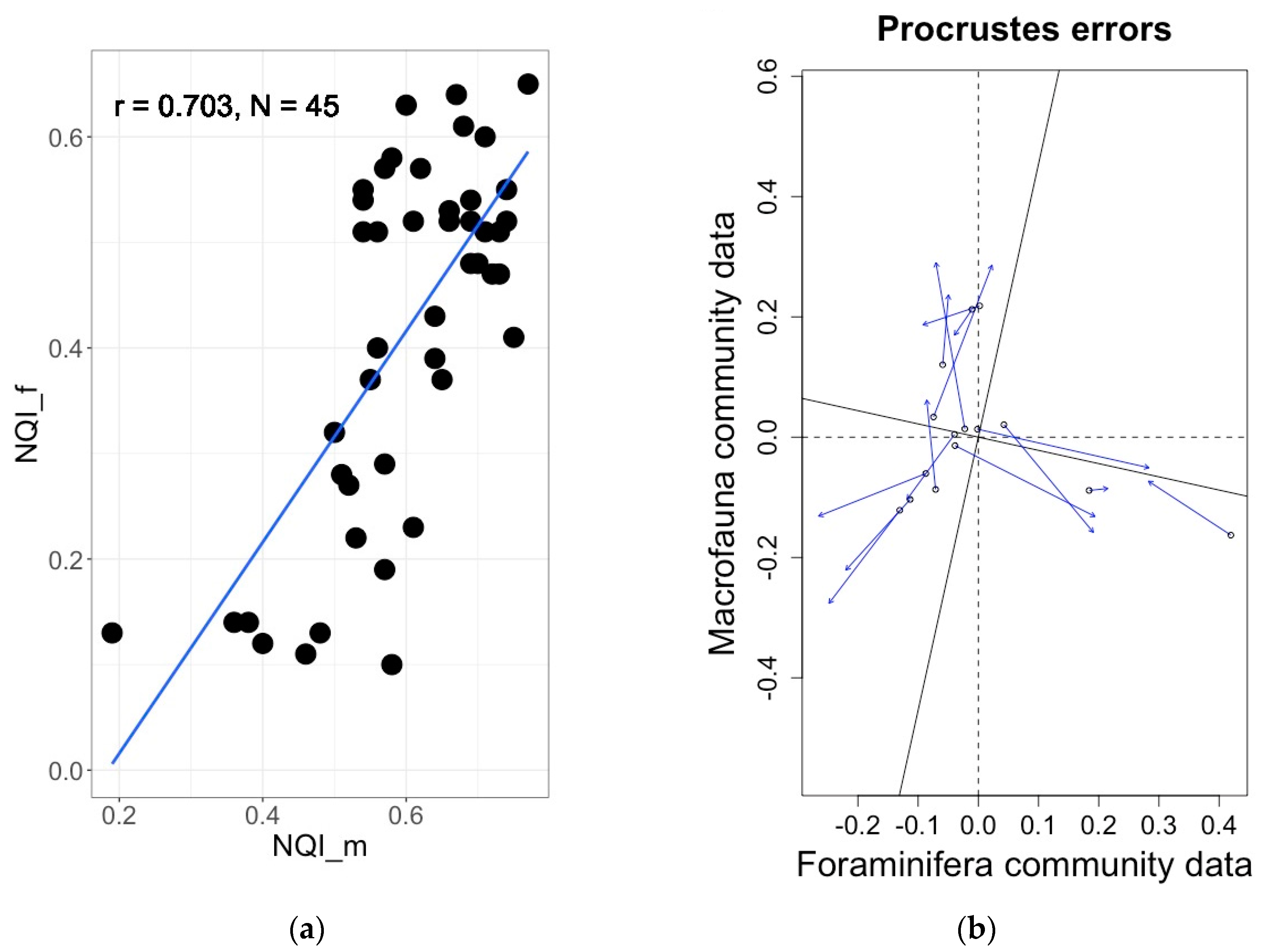
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).