
A Perspective for Best Governance of the Bari Canyon Deep-Sea Ecosystems

 ,
, 

Abstract
1. Introduction
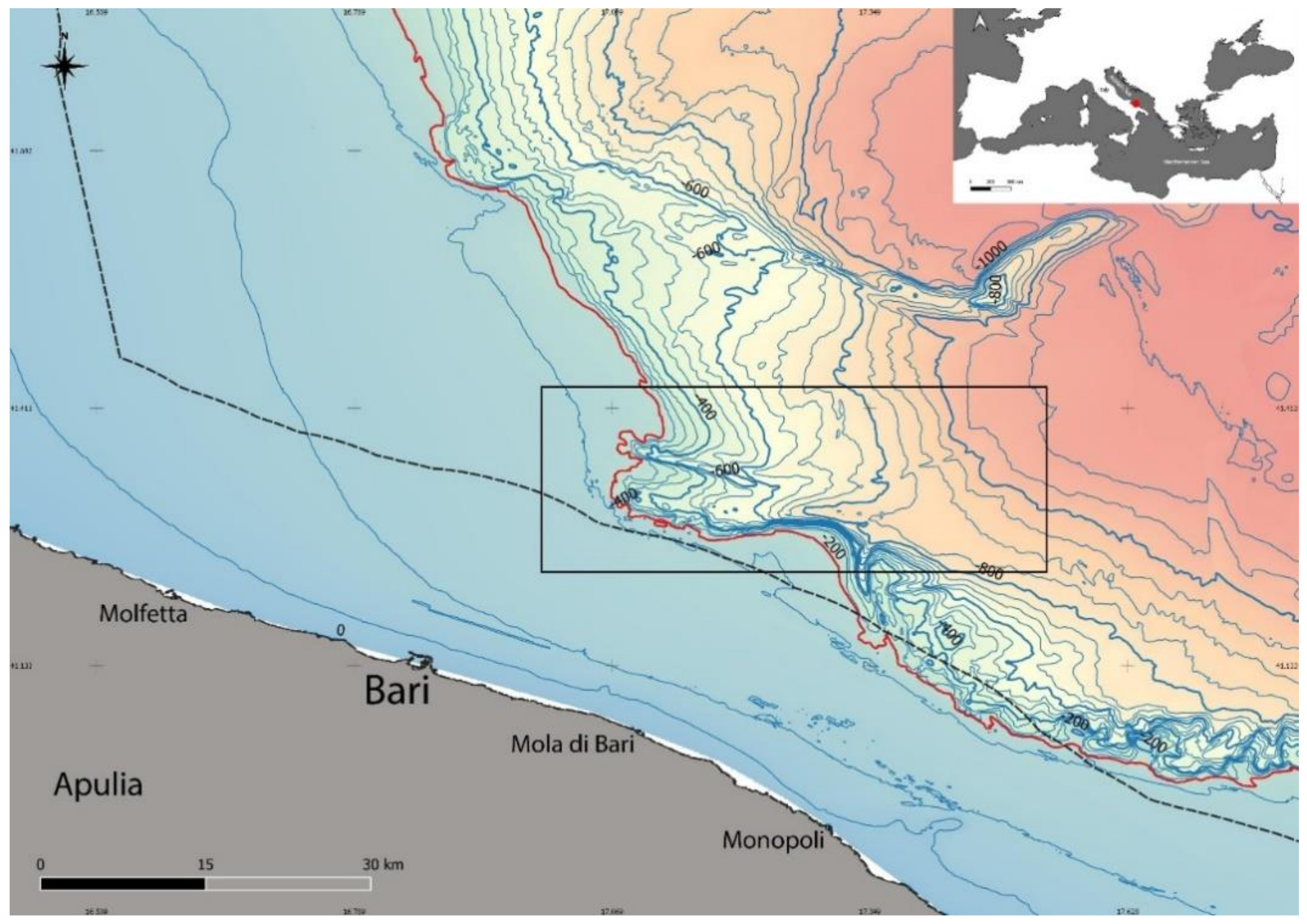
Setting of the Area
2. Materials and Methods
2.1. Benthic and Demersal Fauna: Sampling and Visual Analysis
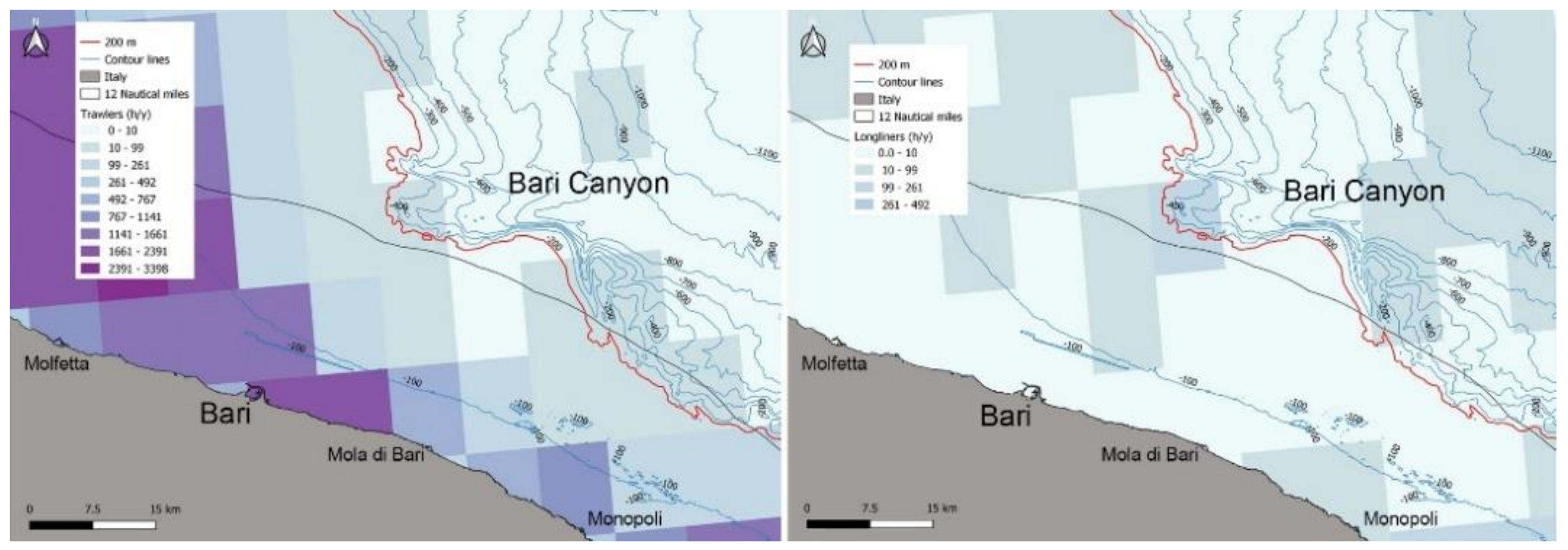
2.2. Fishers, Fisheries, Fishing Pressures
3. Results
3.1. Biodiversity
3.2. Anthropic Impact
3.3. Fishers, Fisheries, Fishing Pressures
3.4. Vulnerable Ecosystems
4. Discussion
4.1. Biodiversity, Anthropic Impact, and Fishing Pressure
4.2. Governance and Protection
- (i)
- Fishing activities could be completely banned, at least for the most impactful gear such as trawls, or only allowed under strict regulations.
- (ii)
- A no-take area could be useful for the protection of the densest CWCs and sponge grounds, as well as EFHs.
- (iii)
- The institution of an offshore protection zone in the Bari Canyon through conservation and management measures will ensure sustainable management of its ecosystems. The establishment of no-take zones within it will protect CWC communities and assist in replenishing overfished stocks. Moreover, it will not strongly affect existing fishing activities in this area negatively, despite restrictions and limitations being defined, but will bring a positive effect to the surroundings and will enhance the goods and services (i.e., ecological and economical value) the canyon can provide. In fact, even if the economic value is difficult to estimate, taking into account other examples of protected areas [119], catches increase almost instantaneously after beginning the protection process [120,121].
- (iv)
- The development of an offshore protection zone should balance short-term and long-term actions, as suggested by [9]. The long-term measures include the more sustainable use of the natural capital and sources for ecosystem services they provide (e.g., hydrodynamic processes of canyons enhancing nutrient supply to the deep-ocean ecosystem, carbon sequestration and storage, refuge sites for other marine life, rich source of genetic resources and chemical compounds, stock recovery, redistribution of commercial species in the nearby areas) by also increasing more sustainable fishing activity in surrounding areas.
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hughes, K.M.; Kaiser, M.J.; Jennings, S.; McConnaughey, R.A.; Pitcher, R.; Hilborn, R.; Amoroso, R.O.; Collie, J.; Hiddink, J.G.; Parma, A.M.; et al. Investigating the effects of mobile bottom fishing on benthic biota: A systematic review protocol. Environ. Evid. 2014, 3, 23. [Google Scholar] [CrossRef]
- Ragnarsson, S.Á.; Burgos, J.M.; Kutti, T.; van den Beld, I.; Egilsdóttir, H.; Arnaud-Haond, S.; Grehan, A. The impact of anthropogenic activity on cold-water corals. In Marine Animal Forests: The Ecology of Benthic Biodiversity Hotspots; Rossi, S., Bramanti, L., Gori, A., Orejas, C., Eds.; Springer International Publishing AG: Berlin/Heidelberg, Germany, 2017; pp. 989–1023. [Google Scholar] [CrossRef]
- Caddell, R. Deep-Sea Bottom Fisheries and the Protection of Seabed Ecosystems: Problems, Progress and Prospects. In The Law of the Seabed: Access, Uses, and Protection of Seabed Resources; Banet, C., Ed.; Brill Nijhoff: Leiden, The Netherlands, 2020; pp. 255–284. [Google Scholar] [CrossRef]
- FAO. Review of the State of World Marine Fishery Resources; FAO Fisheries and Aquaculture Technical Paper No. 569; Food and Agriculture Organization of the United Nations: Rome, Italy, 2011; pp. 1–354. [Google Scholar]
- FAO. The State of World Fisheries and Aquaculture 2020. Sustainability in Action; Food and Agriculture Organization of the United Nations: Rome, Italy, 2020; pp. 1–224. [Google Scholar] [CrossRef]
- UNEP. Ecosystems and Biodiversity in Deep Waters and High Seas; UNEP Regional Seas Reports and Studies No. 178; UNEP/IUCN: Gland, Switzerland, 2006. [Google Scholar]
- Buhl-Mortensen, L.; Neat, F.; Koen-Alonso, M.; Hvingel, C.; Holte, B. Fishing impacts on benthic ecosystems: An introduction to the 2014 ICES symposium special issue. ICES J. Mar. Sci. 2016, 73, i1–i4. [Google Scholar] [CrossRef][Green Version]
- Clark, M.R.; Rowden, A.A. Effect of deep water trawling on the macro-invertebrate assemblages of seamounts on the Chatham Rise, New Zealand. Deep Sea Res. I 2009, 56, 1540–1554. [Google Scholar] [CrossRef]
- Clark, M.R.; Althaus, F.; Schlacher, T.A.; Williams, A.; Bowden, D.A.; Rowden, A.A. The impacts of deep-sea fisheries on benthic communities: A review. ICES J. Mar. Sci. 2016, 73, 51–69. [Google Scholar] [CrossRef]
- Pitcher, T.J.; Clark, M.R.; Morato, T.; Watson, R. Seamount fisheries: Do they have a future? Oceanography 2010, 23, 134–144. [Google Scholar] [CrossRef]
- Norse, E.A.; Brooke, S.; Cheung, W.W.L.; Clark, M.R.; Ekeland, L.; Froese, R.; Gjerde, K.M.; Haedrich, R.L.; Heppell, S.S.; Morato, T.; et al. Sustainability of deep-sea fisheries. Mar. Policy 2012, 36, 307–320. [Google Scholar] [CrossRef]
- Würtz, M. Submarine canyons and their role in the Mediterranean ecosystem. In IUCN Mediterranean Submarine Canyons: Ecology and Governance; Würtz, M., Ed.; IUCN: Gland, Switzerland, 2012; pp. 11–26. [Google Scholar]
- Fernandez-Arcaya, U.; Ramirez-Llodra, E.; Aguzzi, J.; Allcock, A.L.; Davies, J.S.; Dissanayake, A.; Harris, P.; Howell, K.; Huvenne, V.A.I.; Macmillan-Lawler, M.; et al. Ecological role of submarine canyons and need for canyon conservation: A review. Front. Mar. Sci. 2017, 4, 5. [Google Scholar] [CrossRef]
- Koslow, J.A.; Boehlert, G.W.; Gordon, J.D.M.; Haedrich, R.L.; Lorance, P.; Parin, N. Continental slope and deep-sea fisheries: Implications for a fragile ecosystem. ICES J. Mar. Sci. 2000, 57, 548–557. [Google Scholar] [CrossRef]
- Pusceddu, A.; Bianchelli, S.; Martín, J.; Puig, P.; Palanques, A.; Masqué, P.; Danovaro, R. Chronic and intensive bottom trawling impairs deep-sea biodiversity and ecosystem functioning. Proc. Natl. Acad. Sci. USA 2014, 111, 8861–8866. [Google Scholar] [CrossRef]
- European Commission. EU Biodiversity Strategy for 2030. Bringing Nature Back to Our Lives. 2020. Available online: https://ec.europa.eu/info/sites/info/files/communication-annex-eu-biodiversity-strategy-2030_en.pdf (accessed on 10 April 2021).
- FAO. Report of the Technical Consultation on International Guidelines for the Management of Deep-Sea Fisheries in the High Seas; Food and Agriculture Organization of the United Nations: Rome, Italy, 2009. [Google Scholar]
- FAO. Vulnerable Marine Ecosystems: Processes and Practices in the High Seas; Food and Agriculture Organization of the United Nations: Rome, Italy, 2016. [Google Scholar]
- FAO. Report of the FAO Workshop on Deep-Sea Fisheries and Vulnerable Marine Ecosystems of the Mediterranean; Food and Agriculture Organization of the United Nations: Rome, Italy, 2016. [Google Scholar]
- D’Onghia, G.; Calculli, C.; Capezzuto, F.; Carlucci, R.; Carluccio, A.; Grehan, A.; Indennidate, A.; Maiorano, P.; Mastrototaro, F.; Pollice, A.; et al. Anthropogenic impact in the Santa Maria di Leuca cold-water coral province (Mediterranean Sea): Observations and conservation straits. Deep Sea Res. II 2017, 145, 87–101. [Google Scholar] [CrossRef]
- Grehan, A.J.; Arnaud-Haond, S.; D’Onghia, G.; Savini, A.; Yesson, C. Towards ecosystem based management and monitoring of the deep Mediterranean, North-East Atlantic and Beyond. Deep Sea Res. II 2017, 145, 1–7. [Google Scholar] [CrossRef]
- GFCM. Establishment of Geographical Sub-Areas in the GFCM Area Amending the Resolution GFCM/31/2007/2; GFCM Resolution RES-GFCM/33/2009/2; GFCM: Rome, Italy, 2009. [Google Scholar]
- FAO. Major Fishing Areas. Mediterranean and Black Sea (Major Fishing Area 37). CWP Data Collection. FAO Fisheries Division. Available online: http://www.fao.org/fishery/area/Area37/en (accessed on 18 May 2021).
- FAO-GFCM. Report of the Twenty-Sixth Session of The General Fisheries Commission for the Mediterranean; GFCM Report No. 26; FAO: Rome, Italy, 2001; 27p. [Google Scholar]
- Mannini, A.; Sabatella, R.F. Annuario sullo stato delle risorse e sulle strutture produttive dei mari italiani. Biol. Mar. Mediterr. 2015, 22 (Suppl. S1), 358. [Google Scholar]
- Spedicato, M.T.; Zupa, W.; Carbonara, P.; Casciaro, L.; Bitetto, I.; Facchini, M.T.; Gaudio, P.; Palmisano, M.; Lembo, G. Lo stato delle risorse biologiche e della pesca nel Basso Adriatico e nello Ionio nord occidentale. In Il Mare Adriatico e le Sue Risorse; Marini, M., Bombace, G., Iacobone, G., Eds.; Carlo Saladino Editore: Palermo, Italy, 2017; pp. 179–207. [Google Scholar]
- D’Onghia, G.; Sion, L.; Capezzuto, F. Cold-water coral habitats benefit adjacent fisheries along the Apulian margin (central Mediterranean). Fish. Res. 2019, 213, 172–179. [Google Scholar] [CrossRef]
- Oceana. Oceana MedNet, MPA Proposal for the Mediterranean Sea. 100 Reasons to Reach 10%; Oceana: Madrid, Spain, 2011. [Google Scholar]
- Marin, P.; Aguilar, R. Mediterranean submarine canyons 2012: Pending protection. In IUCN Mediterranean Submarine Canyons: Ecology and Governance; Würtz, M., Ed.; IUCN: Gland, Switzerland, 2012; pp. 191–206. [Google Scholar]
- Capezzuto, F.; Ancona, F.; Carlucci, R.; Carluccio, A.; Cornacchia, L.; Maiorano, P.; Ricci, P.; Sion, L.; Tursi, A.; D’Onghia, G. Cold-water coral communities in the Central Mediterranean: Aspects on megafauna diversity, fishery resources and conservation perspectives. Rendiconti Lincei. Scienze Fisiche Nat. 2018, 29, 589–597. [Google Scholar] [CrossRef]
- IUCN. Thematic Report—Conservation Overview of Mediterranean Deep-Sea Biodiversity: A Strategic Assessment; IUCN: Gland, Switzerland, 2019; 122p. [Google Scholar]
- Freiwald, A.; Beuck, L.; Rüggeberg, A.; Taviani, M.; Hebbeln, D.; R/V METEOR Cruise M70-1 Participants. The white coral community in the central Mediterranean Sea revealed by ROV surveys. Oceanography 2009, 22, 58–74. [Google Scholar] [CrossRef]
- Bo, M.; Bertolino, M.; Bavestrello, G.; Canese, S.; Giusti, M.; Angiolillo, M.; Pansini, M.; Taviani, M. Role of deep sponge grounds in the Mediterranean Sea: A case study in southern Italy. Hydrobiologia 2012, 687, 163–177. [Google Scholar] [CrossRef]
- Angeletti, L.; Taviani, M.; Canese, S.; Foglini, F.; Mastrototaro, F.; Argnani, A.; Trincardi, F.; Bakran-Petricioli, T.; Ceregato, A.; Chimienti, G.; et al. New deep-water cnidarian sites in the southern Adriatic Sea. Mediterr. Mar. Sci. 2014, 15, 263–273. [Google Scholar] [CrossRef]
- Angeletti, L.; Prampolini, M.; Foglini, F.; Grande, V.; Taviani, M. Cold-water coral habitat in the Bari Canyon System, Southern Adriatic Sea (Mediterranean Sea). In Seafloor Geomorphology as Benthic Habitat; Harris, P.T., Baker, E., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 811–824. [Google Scholar] [CrossRef]
- D’Onghia, G.; Capezzuto, F.; Cardone, F.; Carlucci, R.; Carluccio, A.; Chimienti, G.; Corriero, G.; Longo, C.; Maiorano, P.; Mastrototaro, F.; et al. Macro- and megafauna recorded in the submarine Bari Canyon (southern Adriatic, Mediterranean Sea) using different tools. Mediterr. Mar. Sci. 2015, 16, 180–196. [Google Scholar] [CrossRef]
- D’Onghia, G.; Calculli, E.; Capezzuto, F.; Carlucci, R.; Carluccio, A.; Maiorano, P.; Pollice, A.; Ricci, P.; Sion, L.; Tursi, A. New records of cold-water coral sites and fish fauna characterization of a potential network existing in the Mediterranean Sea. Mar. Ecol. 2016, 37, 1398–1422. [Google Scholar] [CrossRef]
- Taviani, M.; Angeletti, L.; Beuck, L.; Campiani, E.; Canese, S.; Foglini, F.; Freiwald, A.; Montagna, P.; Trincardi, F. Reprint of “On and off the beaten track: Megafaunal sessile life and Adriatic cascading processes”. Mar. Geol. 2016, 375, 146–160. [Google Scholar] [CrossRef]
- Capezzuto, F.; Sion, L.; Ancona, F.; Carlucci, R.; Carluccio, A.; Cornacchia, L.; Maiorano, P.; Ricci, P.; Tursi, A.; D’Onghia, G. Cold-water coral habitats and canyons as Essential Fish Habitats in the southern Adriatic and northern Ionian Sea (central Mediterranean). Ecol. Quest. 2018, 29, 9–23. [Google Scholar] [CrossRef]
- Carlucci, R.; Lembo, G.; Maiorano, P.; Capezzuto, F.; Marano, C.A.; Sion, L.; Spedicato, M.T.; Ungaro, N.; Tursi, A.; D’Onghia, G. Nursery areas of red mullet (Mullus barbatus), hake (Merluccius merluccius) and deep-water rose shrimp (Parapenaeus longirostris) in Eastern-Central Mediterranean Sea. Estuar. Coast. Shelf Sci. 2009, 83, 529–538. [Google Scholar] [CrossRef]
- Giannoulaki, M.A.; Belluscio, F.; Colloca, S.; Fraschetti, S.; Scardi, M.; Smith, C.; Panayotidis, P.; Valavanis, V.; Spedicato, M.T. Mediterranean Sensitive Habitats, Specific Contract (SI2.600741); Final Report; Hellenic Centre for Marine Research: Attiki, Greece, 2013; 557p. [Google Scholar]
- Druon, J.N.; Fiorentino, F.; Murenu, M.; Knittweis, L.; Colloca, F.; Osio, C.; Merigot, B.; Garofalo, G.; Mannini, A.; Jadaud, A.; et al. Modelling of European hake nurseries in the Mediterranean Sea: An ecological niche approach. Prog. Oceanogr. 2015, 130, 188–204. [Google Scholar] [CrossRef]
- Trincardi, F.; Campiani, E.; Correggiari, A.; Foglini, F.; Maselli, V.; Remia, A. Bathymetry of the Adriatic Sea: The legacy of the last eustatic cycle and the impact of modern sediment dispersal. J. Maps 2014, 10, 151–158. [Google Scholar] [CrossRef]
- Dalla Valle, G.; Gamberi, F.; Foglini, F.; Trincardi, F. The Gondola Slide: A mass transport complex controlled by margin topography (South-western Adriatic Margin, Mediterranean Sea). Mar. Geol. 2015, 366, 97–113. [Google Scholar] [CrossRef]
- Gamberi, F.; Dalla Valle, G.; Foglini, F.; Rovere, M.; Trincardi, F. Submarine Landslides on the Seafloor: Hints on Subaqueous Mass-Transport Processes From the Italian Continental Margins (Adriatic and Tyrrhenian Seas, Offshore Italy). In Submarine Landslides: Subaqueous Mass Transport Deposits from Outcrops to Seismic Profiles; Ogata, K., Festa, A., Pini, G.A., Eds.; American Geophysical Union: Washington, DC, USA, 2019. [Google Scholar] [CrossRef]
- Würtz, M.; Rovere, M. Atlas of the Mediterranean Seamounts and Seamount-Like Structures; IUCN: Gland, Switzerland, 2015; 276p, Available online: https://www.iucn.org/content/atlas-mediterraneanseamounts-and-seamount-structures (accessed on 20 April 2021).
- Foglini, F.; Campiani, E.; Trincardi, F. The reshaping of the SouthWest Adriatic Margin by cascading of dense shelf waters. Mar. Geol. 2016, 375, 64–81. [Google Scholar] [CrossRef]
- Trincardi, F.; Foglini, F.; Verdicchio, G.; Asioli, A.; Correggiari, A.; Minisini, D.; Piva, A.; Remia, A.; Ridente, D.; Taviani, M. The impact of cascading currents on the Bari Canyon System, SW-Adriatic margin (central Mediterranean). Mar. Geol. 2007, 246, 208–230. [Google Scholar] [CrossRef]
- Turchetto, M.; Boldrin, A.; Langone, L.; Miserocchi, S.; Tesi, T.; Foglini, F. Particle transport in the Bari Canyon (southern Adriatic Sea). Mar. Geol. 2007, 246, 231–247. [Google Scholar] [CrossRef]
- Lascaratos, A.; Roether, W.; Nittis, K.; Klein, B. Recent changes in deep water formation and spreading in the Mediterranean Sea: A review. Progr. Oceanogr. 1999, 44, 5–36. [Google Scholar] [CrossRef]
- Cushman-Roisin, B.; Gacic, M.; Poulain, P.M.; Artegiani, A. Physical Oceanography of the Adriatic Sea: Past, Present and Future; Springer: Berlin/Heidelberg, Germany, 2001. [Google Scholar] [CrossRef]
- Cardin, V.; Bensi, M.; Pacciaroni, M. Variability of water mass properties in the last two decades in the south Adriatic Sea with emphasis on the period 2006–2009. Cont. Shelf Res. 2011, 31, 951–965. [Google Scholar] [CrossRef]
- Vilibić, I.; Supić, N. Dense water generation on a shelf: The case of the Adriatic Sea. Ocean Dyn. 2005, 55, 403–415. [Google Scholar] [CrossRef]
- Bonaldo, D.; Orlić, M.; Carniel, S. Framing Continental Shelf Waves in the southern Adriatic Sea, a further flushing factor beyond dense water cascading. Sci. Rep. 2018, 8, 660. [Google Scholar] [CrossRef]
- Taviani, M.; Angeletti, L.; Foglini, F.; Corselli, C.; Nasto, I.; Pons-Brachu, E.; Montagna, P. U/Th dating records of cold-water coral colonization in submarine canyons and adjacent sectors of the southern Adriatic Sea since the Last Glacial Maximum. Prog. Oceanogr. 2019, 175, 300308. [Google Scholar] [CrossRef]
- Bargain, A.; Foglini, F.; Pairaud, I.; Bonaldo, D.; Carniel, S.; Angeletti, L.; Taviani, M.; Rochette, S.; Fabri, M.C. Predictive habitat modelling in two Mediterranean canyons including hydrodynamic variables. Prog. Oceanogr. 2018, 169, 151168. [Google Scholar] [CrossRef]
- Angeletti, L.; Bargain, A.; Campiani, E.; Foglini, F.; Grande, V.; Leidi, E.; Mercorella, A.; Prampolini, M.; Taviani, M. Cold-water coral multiscale habitat mapping: Methodologies and perspectives. In Mediterranean Cold-Water Corals: Past, Present and Future; Orejas, C., Jiménez, C., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2019; pp. 173–189. [Google Scholar]
- Taviani, M.; Angeletti, L.; Antolini, B.; Ceregato, A.; Froglia, C.; Lòpez Correa, M.; Montagna, P.; Remia, A.; Trincardi, F.; Vertino, A. Geo-biology of Mediterranean deep-water coral ecosystems. In DTA/06-2011—Marine Research at CNR–6—Fishery and Sea Resources; Beatrici, D., Braico, P., Cappelletto, M., Gallo, E., Mazari Villanova, L., Moretti, P.F., Eds.; Dipartimento Scienze del Sistema Terra e Tecnologie per l’Ambiente: Rome, Italy, 2011; pp. 705–720. [Google Scholar]
- Castellan, G.; Angeletti, L.; Correggiari, A.; Foglini, F.; Grande, V.; Taviani, M. Visual Methods for Monitoring Mesophotic-to-Deep Reefs and Animal Forests: Finding a Compromise Between Analytical Effort and Result Quality. In Perspectives on the Marine Animal Forests of the World; Rossi, S., Bramanti, L., Eds.; Springer: Berlin/Heidelberg, Germany, 2021; pp. 487–514. [Google Scholar] [CrossRef]
- Bell, J.J.; Barnes, D.K.A. Sponge morphological diversity: A qualitative predictor of species diversity? Aquat. Conserv. Mar. Freshw. Ecosyst. 2001, 11, 109–121. [Google Scholar] [CrossRef]
- D’Onghia, G.; Capezzuto, F.; Carluccio, A.; Carlucci, R.; Giove, A.; Mastrototaro, F.; Panza, M.; Sion, L.; Tursi, A.; Maiorano, P. Exploring composition and behaviour of fish fauna by in situ observations in the Bari Canyon (Southern Adriatic Sea, Central Mediterranean). Mar. Ecol. 2015, 36, 541–556. [Google Scholar] [CrossRef]
- Sion, L.; Calculli, C.; Capezzuto, F.; Carlucci, R.; Carluccio, A.; Cornacchia, L.; Maiorano, P.; Pollice, A.; Ricci, P.; Tursi, A.; et al. Does the Bari Canyon (Central Mediterranean) influence the fish distribution and abundance? Progr. Oceanogr. 2019, 170, 81–92. [Google Scholar] [CrossRef]
- WoRMS Editorial Board. World Register of Marine Species. VLIZ. 2021. Available online: https://www.marinespecies.org (accessed on 3 March 2021).
- Lembo, G.; Carbonara, P.; Casciaro, L.; Bitetto, I.; Facchini, M.T.; Divanocić, D.; Joksimović, A.; Marcović, Z.; Ikica, Z.; Hoxha, A.; et al. Review of 2015 Medits Survey in the GSA18-South Adriatic Sea. Medits. Annual Report. 2016. Available online: https://www.sibm.it/MEDITS%202011/principalereports.htm (accessed on 16 April 2021).
- Piano di Lavoro Raccolta Dati Alieutici REG. (UE) N. 508/2014 Relativo al Fondo Europeo per gli Affari Marittimi e la Pesca (FEAMP) e REG. (EU) N. 2017/1004. Available online: https://dcf-italia.cnr.it/web/ (accessed on 18 May 2021).
- Kroodsma, D.A.; Mayorga, J.; Hochberg, T.; Miller, N.A.; Boerder, K.; Ferretti, F.; Wilson, A.; Bergman, B.; White, T.D.; Block, B.A.; et al. Tracking the global footprint of fisheries. Science 2018, 359, 904–908. [Google Scholar] [CrossRef] [PubMed]
- Natale, F.; Gibin, M.; Alessandrini, A.; Vespe, M.; Paulrud, A. Mapping Fishing Effort through AIS Data. PLoS ONE 2015, 10, e0130746. [Google Scholar] [CrossRef] [PubMed]
- Bertrand, J.A.; de Sola, L.G.; Papaconstantinou, C.; Relini, G.; Souplet, A. The general specifications of the MEDITS surveys. Sci. Mar. 2002, 66, 9–17. [Google Scholar] [CrossRef]
- Lembo, G. Programma Nazionale Italiano per la Raccolta di Dati Alieutici 2015. Modulo H “Campionamento Biologico delle Catture”; Rapporto Finale GSA18-Adriatico Meridionale; COISPA Tecnologia e Ricerca: Bari, Italy, 2016; 110p. [Google Scholar]
- Marini, M.; Bombace, G.; Iacobone, G. Il Mare Adriatico e le Sue Risorse; Carlo Saladino Editore: Palermo, Italy, 2017; pp. 1–267. [Google Scholar]
- Pinello, D.; Gee, J.; Dimech, M. Handbook for Fisheries Socio-Economic Sample Survey Principles and Practice; FAO Fisheries and Aquaculture Technical Paper No. 613; FAO: Rome, Italy, 2017. [Google Scholar]
- IUCN. Socio-Economic Study on the Fisheries and Fleets Potentially Involved in the Introduction of a Fishery Restricted Area (FRA) in the Bari Canyon; Internal Report; IUCN: Gland, Switzerland, 2019; pp. 1–17. [Google Scholar]
- Sanfilippo, R.; Vertino, A.; Rosso, A.; Beuck, L.; Freiwald, A.; Taviani, M. Serpula-aggregates and their role in deep-sea coral communities in the southern Adriatic Sea. Facies 2013, 59, 663677. [Google Scholar] [CrossRef]
- Castellan, G.; Angeletti, L.; Taviani, M.; Montagna, P. The yellow coral Dendrophyllia cornigera in a warming ocean. Front. Mar. Sci. 2019, 6, 692. [Google Scholar] [CrossRef]
- Chimienti, G.; Bo, M.; Taviani, M.; Mastrototaro, F. Occurrence and biogeography of Mediterranean CWCs. In Mediterranean Cold-Water Corals: Past, Present and Future; Orejas, C., Jiménez, C., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2019; p. 213243. [Google Scholar]
- Foglini, F.; Grande, V.; Marchese, F.; Bracchi, V.A.; Prampolini, M.; Angeletti, L.; Castellan, G.; Chimienti, G.; Hansen, I.M.; Gudmundsen, M.; et al. Application of Hyperspectral Imaging to Underwater Habitat Mapping, Southern Adriatic Sea. Sensors 2019, 19, 2261. [Google Scholar] [CrossRef] [PubMed]
- Rueda, J.L.; Urra, J.; Aguilar, R.; Angeletti, L.; Bo, M.; García-Ruiz, C.; Gonzalez-Duarte, M.M.; Lopez, E.; Madurell, T.; Maldonaldo, M.; et al. Cold-water coral associated fauna in the Mediterranean Sea and Adjacent Areas. In Mediterranean Cold-Water Corals: Past, Present and Future; Orejas, C., Jiménez, C., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2019; pp. 295–333. [Google Scholar] [CrossRef]
- Addamo, A.M.; Vertino, A.; Stolarski, J.; Garcia-Jiménez, R.; Taviani, M.; Machordom, A. Merging scleractinian genera: The overwhelming genetic similarity between solitary Desmophyllum and colonial Lophelia. BMC Evol. Biol. 2016, 16, 108. [Google Scholar] [CrossRef]
- IUCN. The IUCN Red List of Threatened Species. Version 2021-1. 2021. Available online: https://www.iucnredlist.org (accessed on 1 March 2021).
- Lembo, G.; Donnaloia, L. Osservatorio Regionale Pesca e Acquacoltura. Puglia 2007; COISPA: Bari, Italy, 2007; pp. 1–89. [Google Scholar]
- Lembo, G.; Spedicato, M.T. Lo stato delle risorse demersali nei mari italiani. GSA 18—Adriatico meridionale. In The State of Italian Marine Fisheries and Aquaculture in Italy; Cataudella, S., Spagnolo, M., Eds.; Ministero delle Politiche Agricole Alimentari e Forestali (MiPAAF): Rome, Italy, 2011; pp. 79–87. [Google Scholar]
- Lembo, G.; Spedicato, M.T. Caratterizzazione ambientale delle aree di pesca. GSA 18—Adriatico meridionale. In The State of Italian Marine Fisheries and Aquaculture in Italy; Cataudella, S., Spagnolo, M., Eds.; Ministero delle Politiche Agricole Alimentari e Forestali (MiPAAF): Rome, Italy, 2011; pp. 159–170. [Google Scholar]
- STECF. Small Pelagic Stocks in the Adriatic Sea. Mediterranean Assessments Part 1 (STECF-15-14); Publications Office of the European Union: Luxembourg, 2015. [Google Scholar]
- GFCM. Forty-Second Session of the Commission. Final Report; FAO: Rome, Italy, 2018; 129p. [Google Scholar]
- EUNIS Marine Habitat Classification. 2019. Available online: https://www.eea.europa.eu/data-and-maps/data/eunis-habitat-classification (accessed on 1 March 2021).
- Capezzuto, F.; Ancona, F.; Calculli, C.; Sion, L.; Maiorano, P.; D’Onghia, G. Feeding of the deep-water fish Helicolenus dactylopterus (Delaroche, 1809) in different habitats: From muddy bottoms to cold-water coral habitats. Deep Sea Res. I 2020, 159, 103252. [Google Scholar] [CrossRef]
- Capezzuto, F.; Ancona, F.; Calculli, C.; Carlucci, R.; Sion, L.; Maiorano, P.; D’Onghia, G. Comparison of trophic spectrum in the blackspot seabream, Pagellus bogaraveo (Brünnich, 1768), between cold-water coral habitats and muddy bottoms in the central Mediterranean. Deep Sea Res. I 2021, 169, 103474. [Google Scholar] [CrossRef]
- Taviani, M.; Angeletti, L.; Canese, S.; Cannas, R.; Cardone, F.; Cau, A.; Cau, A.B.; Follesa, M.C.; Marchese, F.; Montagna, P.; et al. The “Sardinian cold-water coral province” in the context of the Mediterranean coral ecosystems. Deep Sea Res. II 2017, 145, 61–78. [Google Scholar] [CrossRef]
- Fanelli, E.; Delbono, I.; Ivaldi, R.; Pratellesi, M.; Cocito, S.; Peirano, A. Cold-water coral Madrepora oculata in the eastern Ligurian Sea (NW Mediterranean): Historical and recent findings. Aquat. Conserv. Mar. Freshw. Ecosyst. 2016, 27, 965–975. [Google Scholar] [CrossRef]
- Fabri, M.C.; Pedel, L.; Beuck, L.; Galgani, F.; Hebbeln, D.; Freiwald, A. Megafauna of vulnerable marine ecosystems in French Mediterranean submarine canyons: Spatial distribution and anthropogenic impacts. Deep Sea Res. II 2014, 104, 184–207. [Google Scholar] [CrossRef]
- Orejas, C.; Gori, A.; Lo Iacono, C.; Puig, P.; Gili, J.-M.; Dale, M.R. Cold-water corals in the Cap de Creus canyon, northwestern Mediterranean: Spatial distribution, density and anthropogenic impact. Mar. Ecol. Prog. Ser. 2009, 397, 37–51. [Google Scholar] [CrossRef]
- Mastrototaro, F.; D’Onghia, G.; Corriero, G.; Matarrese, A.; Maiorano, P.; Panetta, P.; Gherardi, M.; Longo, C.; Rosso, A.; Sciuto, F.; et al. Biodiversity of the white coral bank off Cape Santa Maria di Leuca (Mediterranean Sea): An update. Deep Sea Res. II 2010, 57, 412–430. [Google Scholar] [CrossRef]
- Taviani, M.; Angeletti, L.; Cardone, F.; Montagna, P.; Danovaro, R. A unique and threatened deep water coral-bivalve biotope new to the Mediterranean Sea offshore the Naples megalopolis. Sci. Rep. 2019, 9, 3411. [Google Scholar] [CrossRef]
- Angeletti, L.; Castellan, G.; Montagna, P.; Remia, A.; Taviani, M. The “Corsica Channel Cold-Water Coral Province” (Mediterranean Sea). Front. Mar. Sci. 2020, 7, 661. [Google Scholar] [CrossRef]
- Longo, C.; Mastrototaro, F.; Corriero, G. Sponge fauna associated with a Mediterranean deep-sea coral bank. J. Mar. Biol. Assoc. UK 2005, 85, 1341–1352. [Google Scholar] [CrossRef][Green Version]
- Ramirez-Llodra, E.; Brandt, A.; Danovaro, R.; De Mol, B.; Escobar, E.; German, C.R.; Levin, L.A.; Martinez Arbizu, P.; Menot, L.; Buhl-Mortensen, P.; et al. Deep, diverse and definitely different: Unique attributes of the world’s largest ecosystem. Biogeosciences 2010, 7, 2851–2899. [Google Scholar] [CrossRef]
- Giusti, M.; Canese, S.; Fourt, M.; Bo, M.; Innocenti, C.; Goujard, A.; Daniel, B.; Angeletti, L.; Taviani, M.; Aquilina, L.; et al. Coral forests and derelict fishing gears in submarine canyon systems of the Ligurian Sea. Prog. Oceanogr. 2019, 178, 102186. [Google Scholar] [CrossRef]
- Enrichetti, F.; Dominguez-Carrió, C.; Toma, M.; Bavestrello, G.; Canese, S.; Bo, M. Assessment and distribution of seafloor litter on the deep Ligurian continental shelf and shelf break (NW Mediterranean Sea). Mar. Pollut. Bull. 2020, 151, 110872. [Google Scholar] [CrossRef]
- Ramirez-Llodra, E.; De Mol, B.; Company, J.B.; Coll, M.; Sardà, F. Effects of natural and anthropogenic processes in the distribution of marine litter in the deep Mediterranean Sea. Prog. Oceanogr. 2013, 118, 273–287. [Google Scholar] [CrossRef]
- Tubau, X.; Canals, M.; Lastras, G.; Rayo, X.; Rivera, J.; Amblas, D. Marine litter on the floor of deep submarine canyons of the Northwestern Mediterranean Sea: The role of hydrodynamic processes. Prog. Oceanogr. 2015, 134, 379–403. [Google Scholar] [CrossRef]
- Prampolini, M.; Angeletti, L.; Grande, V.; Taviani, M.; Foglini, F. Tricase Submarine Canyon: Cold-water coral habitats in the southwesternmost Adriatic Sea (Mediterranean Sea). In Seafloor Geomorphology as Benthic Habitat; Harris, P.T., Baker, E., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 793–810. [Google Scholar] [CrossRef]
- UNEP-MAP-RAC/SPA. Status and Conservation of Fisheries in the Adriatic Sea; Draft Internal Report for the Purposes of the Mediterranean Regional Workshop to Facilitate the Description of Ecologically or Biologically Significant Marine Areas, Malaga, Spain, 7–11 April 2014; UNEP-MAP-RAC/SPA: Athens, Greece, 2014. [Google Scholar]
- Otero, M.; Marin, P. Conservation of cold-water corals in the Mediterranean: Current status and future prospects for improvement. In Mediterranean Cold-Water Corals: Past, Present and Future; Orejas, C., Jiménez, C., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2019. [Google Scholar]
- De Juan, S.; Lleonart, J. A conceptual framework for the protection of vulnerable habitats impacted by fishing activities in the Mediterranean high seas. Ocean Coast. Manag. 2010, 53, 717–723. [Google Scholar] [CrossRef]
- Ardron, J.A.; Clark, M.R.; Penney, A.J.; Hourigan, T.F.; Rowden, A.A.; Dunstan, P.K.; Watling, L.; Shank, T.M.; Tracey, M.; Dunn, M.R.; et al. A systematic approach towards the identification and protection of vulnerable marine ecosystems. Mar. Policy 2014, 49, 146–154. [Google Scholar] [CrossRef]
- Blasiak, R.; Yagi, N. Shaping an international agreement on marine biodiversity beyond areas of national jurisdiction: Lessons from high seas fisheries. Mar. Policy 2016, 71, 210–216. [Google Scholar] [CrossRef]
- Reed, J.R.; Lombard, A.T.; Sink, K.J. A diversity of spatial management instruments can support integration of fisheries management and marine spatial planning. Mar. Policy 2020, 119, 104089. [Google Scholar] [CrossRef]
- D’Onghia, G. Cold-water corals as shelter, feeding and life-history critical habitats for fish species: Ecological interactions and fishing impact. In Mediterranean Cold-Water Corals: Past, Present and Future; Orejas, C., Jiménez, C., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2019. [Google Scholar]
- Wagner, D.; Friedlander, A.M.; Pyle, R.L.; Brooks, C.; Gjerde, K.M.; Wilhelm, T.A. Coral Reefs of the High Seas: Hidden Biodiversity Hotspots in Need of Protection. Front. Mar. Sci. 2020, 7, 567428. [Google Scholar] [CrossRef]
- Rice, J.; Houston, K. Representativity and networks of marine protected areas. Aquat. Conserv. Mar. Freshw. Ecosyst. 2011, 21, 649–657. [Google Scholar] [CrossRef]
- Dunn, D.C.; Ardron, J.A.; Bax, N.; Bernal, P.; Cleary, J.; Cresswell, I.; Donnelly, B.; Dunstan, P.; Gjerde, K.; Johnson, D.; et al. The convention on biological diversity’s ecologically or biologically significant areas: Origins, development, and current status. Mar. Policy 2013, 49, 137–145. [Google Scholar] [CrossRef]
- Gjerde, K.M.; Currie, D.; Wowk, K.; Scak, K. Ocean in peril: Reforming the management of global ocean living resources in areas beyond national jurisdiction. Mar. Pollut. Bull. 2013, 74, 540–551. [Google Scholar] [CrossRef] [PubMed]
- CBD. Report of the Mediterranean Regional Workshop to Facilitate the Description of Ecologically or Biologically Significant Marine Areas. UNEP/CBD/EBSA/WS/2014/3/4. 2014. Available online: http://www.cbd.int/doc/meetings/mar/ebsaws-2014-03/official/ebsaws-2014-03-04-en.pdf (accessed on 16 April 2021).
- Druel, E.; Gjerdem, K.M. Sustaining marine life beyond boundaries: Options for an implementing agreement for marine biodiversity beyond national jurisdiction under the United Nations Convention on the Law of the Sea. Mar. Policy 2014, 49, 90–97. [Google Scholar] [CrossRef]
- Rees, S.E.; Foster, N.L.; Langmead, O.; Pittmann, S.; Johnson, D.E. Defining the qualitative elements of Aichi Biodiversity Target 11 with regard to the marine and coastal environment in order to strengthen global efforts for marine biodiversity conservation outlined in the United Nations sustainable development goal 14. Mar. Policy 2018, 93, 241–250. [Google Scholar] [CrossRef]
- Danovaro, R.; Fanelli, E.; Canals, M.; Ciuffardi, T.; Fabri, M.-C.; Taviani, M.; Argyrou, M.; Azzurro, E.; Bianchelli, S.; Cantafaro, A.; et al. Towards a marine strategy for the deep Mediterranean Sea: Analysis of current ecological status. Mar. Policy 2020, 112, 103781. [Google Scholar] [CrossRef]
- Stojanovic, T.; Gee, K. Governance as a framework to theorise and evaluate marine planning. Mar. Policy 2020, 120, 104115. [Google Scholar] [CrossRef]
- de la Torriente, A.; Gonzàlez-Irusta, J.M.; Aguilar, R.; Fernàndez-Salas, L.M.; Punzòn, A.; Serrano, A. Benthic habitat modelling and mapping as a conservation tool for marine protected areas: A seamount in the western Mediterranean. Aquat. Conserv. Mar. Freshw. Ecosyst. 2019, 29, 732–750. [Google Scholar] [CrossRef]
- GFCM-RAC/SPA. Report of the Transversal Workshop on Marine Protected Areas (MPAs). Salammbô, Tunisia; GFCM-RAC/SPA: Rome, Italy, 2007; p. 34. [Google Scholar]
- ICF. Study on the Economic Benefits of Marine Protected Areas; European Commission: Brussels, Belgium, 2018; pp. 1–147. Available online: https://www.msp-platform.eu/sites/default/files/ea0318223enn.en__0.pdf (accessed on 1 March 2021). [CrossRef]
- Huvenne, V.A.I.; Bett, B.J.; Masson, D.G.; Le Bas, T.P.; Wheeler, A.J. Effectiveness of a deep-sea cold-water coral Marin Protected Area, following 8 years of fisheries closure. Biol. Conserv. 2016, 200, 60–69. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Station | Start Lat N—Long E (GG°mm.xx’) | End Lat N—Long E (GG°mm.xx’) | Start–End Depth (m) | Length (m) | Frames |
|---|---|---|---|---|---|
| A71 | 41°17.26′—17°16.61′ | 41°17.27′—17°16.63′ | 428–434 | 72 | 589 |
| A77 | 41°14.66′—17°16.61′ | 41°12.01′—16°56.91′ | 422–449 | 55 | 149 |
| A91 | 41°16.99′—17°17.55′ | 41°17.00′—17°17.53′ | 309–306 | 67 | 583 |
| A207 | 41°17.27′—17°16.60′ | 41°17.18′—17°16,75′ | 460–330 | 1414 | 122 |
| A208 | 41°14.24′—17°17.05′ | 41°17.15′—17°16.66′ | 412–382 | 654 | 418 |
| A210 | 41°17.31′—17°17.12′ | 41°16.77′—17°15.94′ | 478–479 | 1075 | 478 |
| MS15_20 | 41°17.71′—17°07.87′ | 41°17.58′—17°08.24′ | 284–205 | 2064 | 263 |
| MS15_22 | 41°19.05′—17°05.02′ | 41°19.03′—17°04.96′ | 293–245 | 1092 | 149 |
| MS15_24 | 41°16.92′—17°17.92′ | 41°16.76′—17°17.66′ | 284–222 | 1287 | 89 |
| MS15_25 | 41°15.13′—17°19.55′ | 41°15.13′—17°19.14′ | 299–219 | 1133 | 87 |
| MS17_II_140 | 41°17.32′—17°17.12′ | 41°17.27′—17°17.03′ | 490–385 | 860 | 508 |
| MS17_II_141 | 41°22.69′—17°06.92′ | 41°22.97′—17°06.41′ | 274–201 | 1417 | 788 |
| Total frames | 4223 |
| Taxon | Status | Legal Instrument |
|---|---|---|
| Porifera | ||
| Biemna parthenopea * | ||
| Biemna tenuisigma * | ||
| Eurypon topsenti * | ||
| Hexadella pruvoti * | ||
| Poecillastra compressa | VME indicator taxa | |
| Pachastrella monilifera | VME indicator taxa | |
| Cnidaria | ||
| Dendrophyllia cornigera | Endangered | IUCN, Annex II of SPA/BD, VME indicator taxa |
| Desmophyllum dianthus | Endangered | IUCN, Annex II of SPA/BD, VME indicator taxa |
| Desmophyllum pertusum | Endangered | IUCN, Annex II of SPA/BD, VME indicator taxa |
| Leiopathes glaberrima | Endangered | IUCN, Annex II of SPA/BD, VME indicator taxa |
| Madrepora oculata | Endangered | IUCN, Annex II of SPA/BD, VME indicator taxa |
| Polychaeta | ||
| Hyalopomatus madreporae * | ||
| Vermiliopsis monodiscus * | ||
| Chondrichthyes | ||
| Centrophorus granulosus | Critically endangered | IUCN |
| Chimaera monstrosa | Near threatened | IUCN |
| Dalatias licha | Vulnerable | IUCN |
| Hexanchus griseus | Least concern | IUCN |
| Actinopterygii | ||
| Helicolenus dactylopterus | Least concern | IUCN |
| Merluccius merluccius | Vulnerable | IUCN, GFCM Species priority |
| Polyprion americanus | Data deficient | IUCN |
| Phycis phycis | Least concern | IUCN |
| Pagellus bogaraveo | Least concern | IUCN, GFCM species priority |
| Station | Longlines | Plastic Net | Plastic Bag | Plastic Can | Plastic Tube | Aluminum Barrel/Can | Glass | Total |
|---|---|---|---|---|---|---|---|---|
| A71 | 1 | 1 | ||||||
| A77 | 1 | 1 | ||||||
| A91 | 0 | |||||||
| A208 | 2 | 1 | 3 | |||||
| A207 | 1 | 1 | 2 | |||||
| A210 | 2 | 1 | 3 | |||||
| MS15_20 | 2 | 2 | ||||||
| MS15_22 | 25 | 2 | 1 | 2 | 30 | |||
| MS15_24 | 0 | |||||||
| MS15_25 | 0 | |||||||
| MS17_II_140 | 1 | 1 | 2 | 1 | 5 | |||
| MS17_II_141 | 8 | 1 | 9 | |||||
| Total | 41 | 1 | 5 | 3 | 1 | 3 | 2 | 56 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Angeletti, L.; D’Onghia, G.; Otero, M.d.M.; Settanni, A.; Spedicato, M.T.; Taviani, M. A Perspective for Best Governance of the Bari Canyon Deep-Sea Ecosystems. Water 2021, 13, 1646. https://doi.org/10.3390/w13121646
Angeletti L, D’Onghia G, Otero MdM, Settanni A, Spedicato MT, Taviani M. A Perspective for Best Governance of the Bari Canyon Deep-Sea Ecosystems. Water. 2021; 13(12):1646. https://doi.org/10.3390/w13121646
Chicago/Turabian StyleAngeletti, Lorenzo, Gianfranco D’Onghia, Maria del Mar Otero, Antonio Settanni, Maria Teresa Spedicato, and Marco Taviani. 2021. "A Perspective for Best Governance of the Bari Canyon Deep-Sea Ecosystems" Water 13, no. 12: 1646. https://doi.org/10.3390/w13121646
APA StyleAngeletti, L., D’Onghia, G., Otero, M. d. M., Settanni, A., Spedicato, M. T., & Taviani, M. (2021). A Perspective for Best Governance of the Bari Canyon Deep-Sea Ecosystems. Water, 13(12), 1646. https://doi.org/10.3390/w13121646