Detection of Cyanotoxin-Producing Genes in a Eutrophic Reservoir (Billings Reservoir, São Paulo, Brazil)

Abstract
1. Introduction
2. Materials and Methods
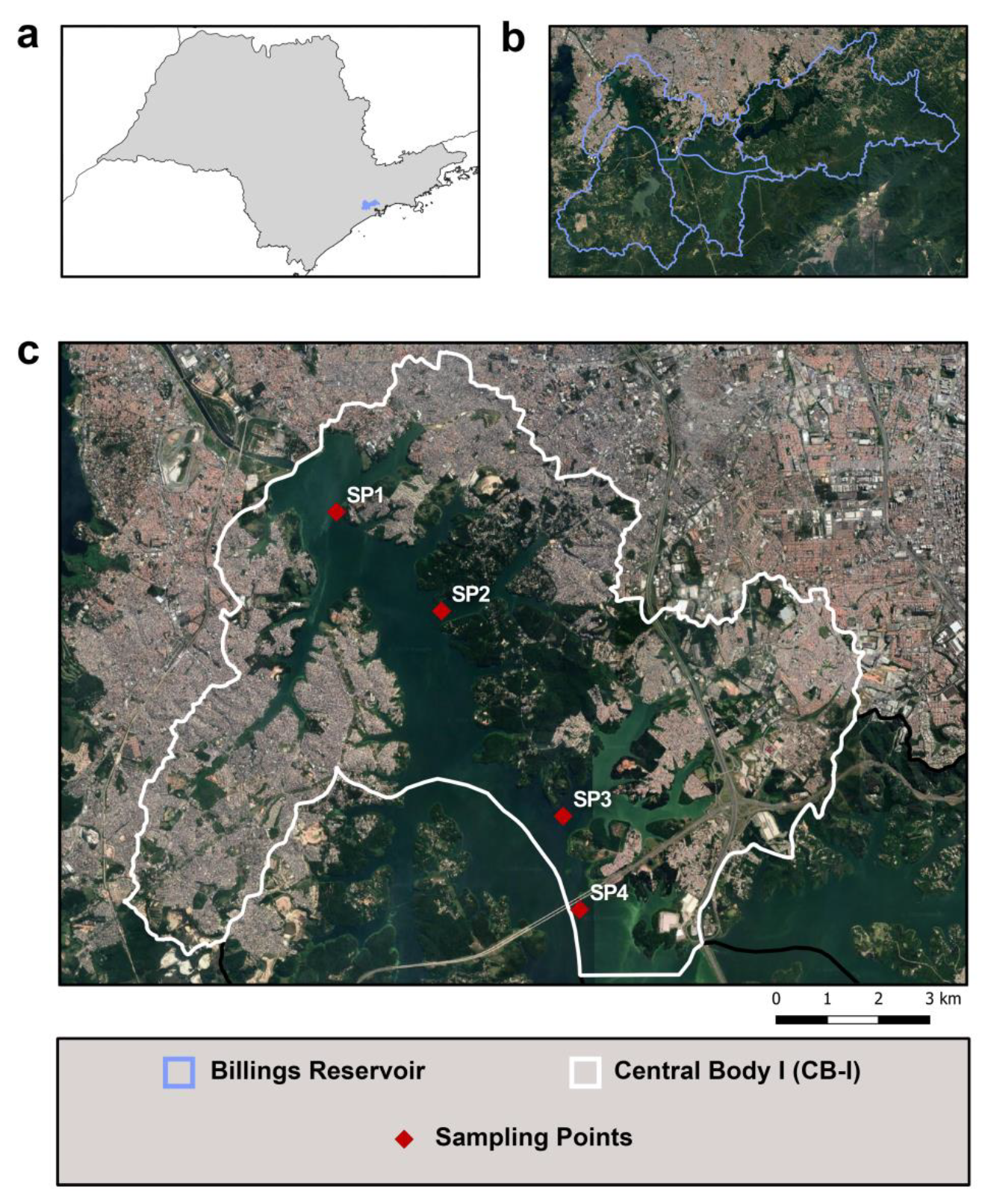
2.1. Area of Study
2.2. Sampling
2.3. DNA Extraction from the Microbial Community
2.4. Amplification of the Cyanotoxin Genes
2.5. Cyanobacteria Strains
2.6. Cyanobacteria and Eukaryotic Microalgae Identification and Density
3. Results
3.1. Amplification of Cyanotoxin Genes
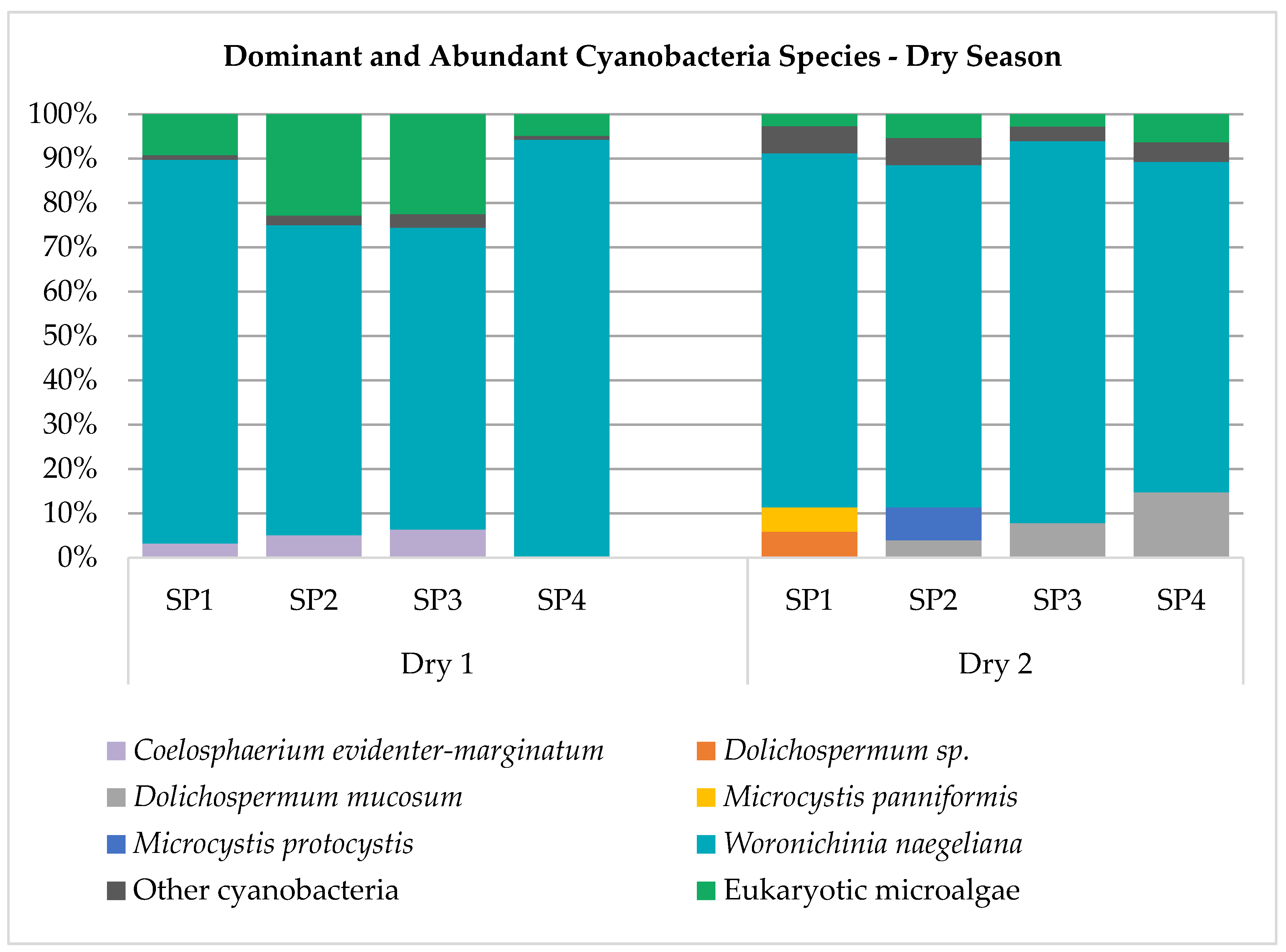
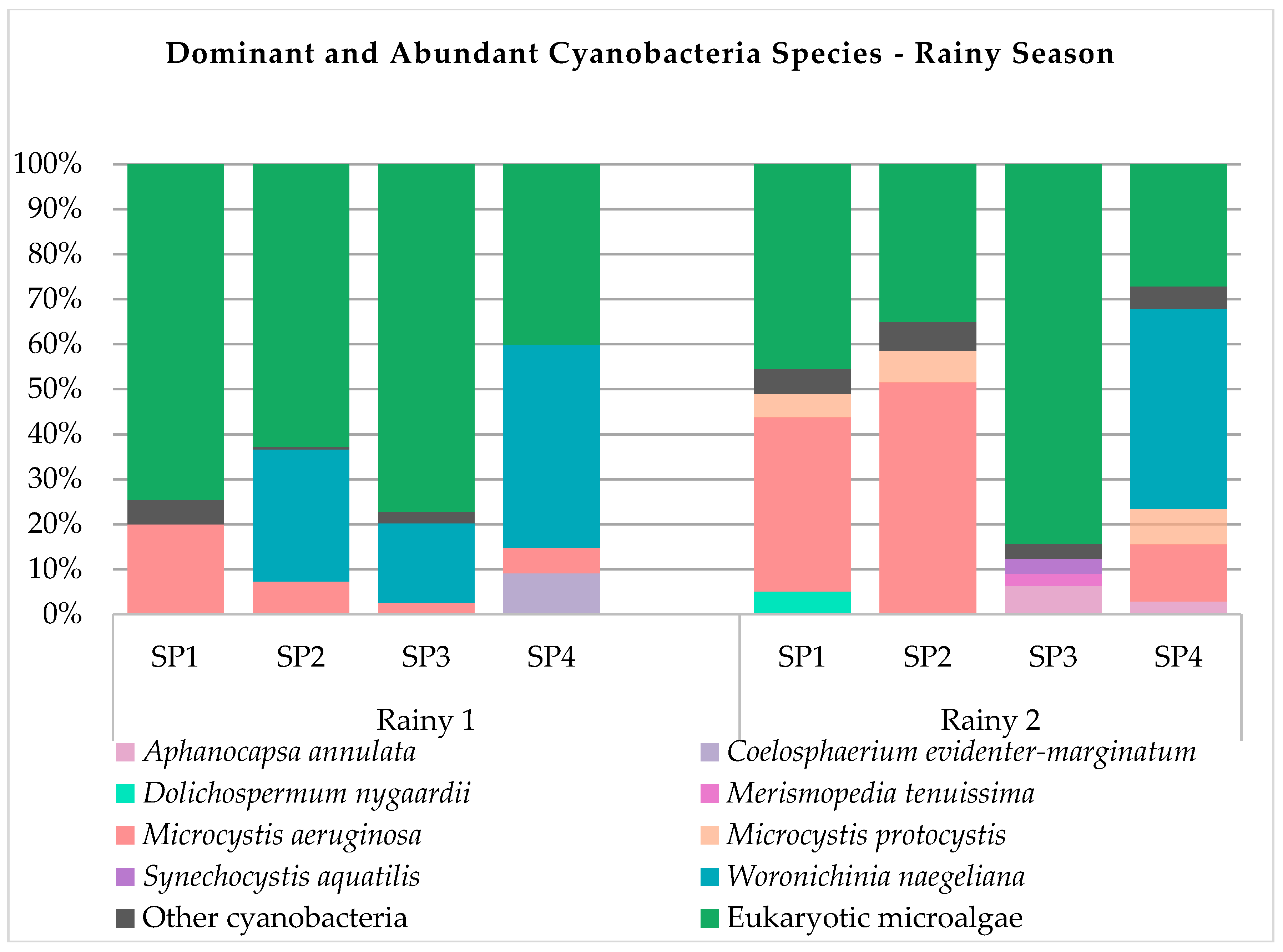
3.2. Cyanobacteria Composition
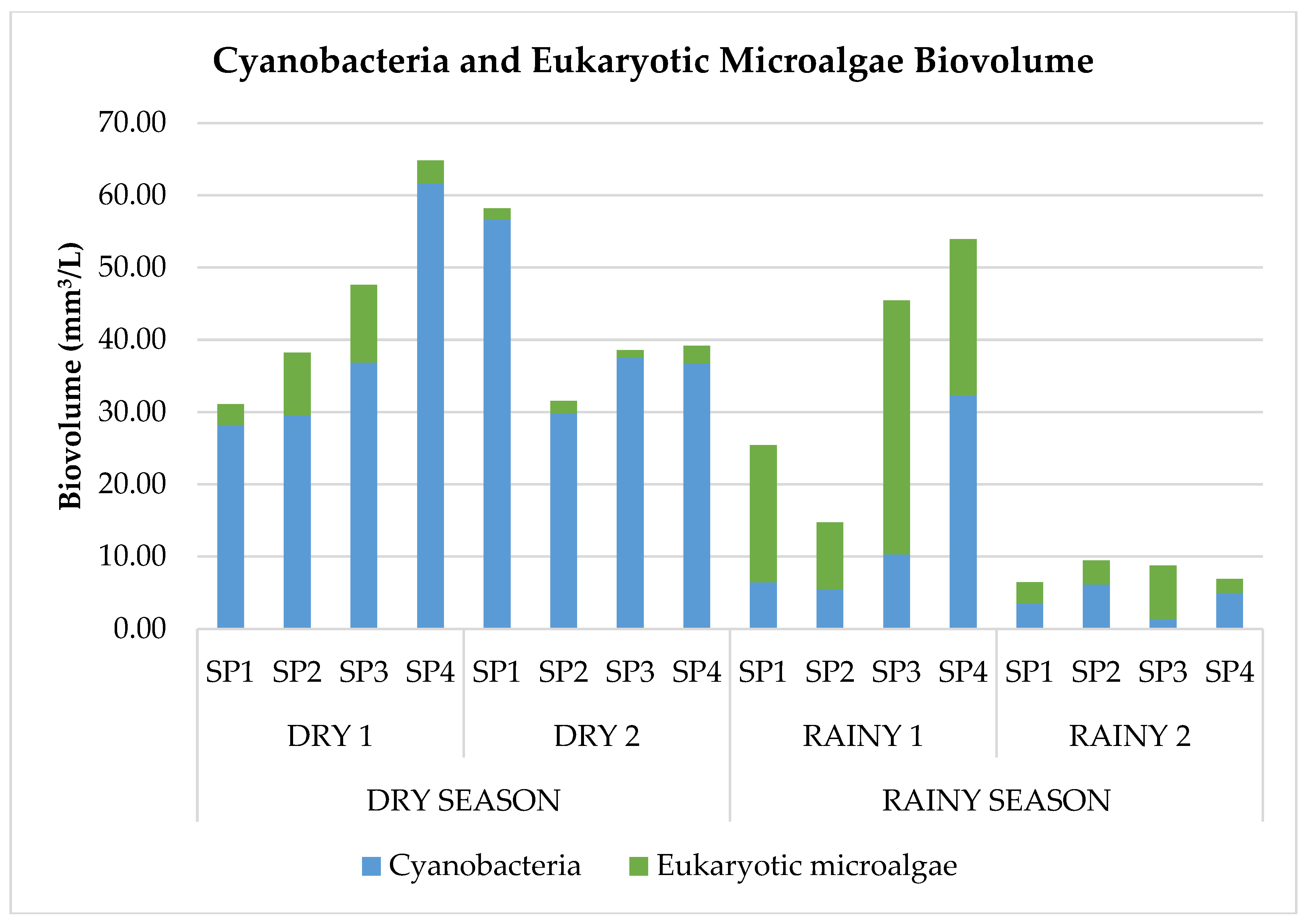
3.3. Cyanobacteria Biovolume
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Saad, A.; Atia, A. Review on freshwater blue-green algae (cyanobacteria): Occurrence, classification and toxicology. Biosci. Biotechnol. Res. Asia 2014, 11, 1319–1325. [Google Scholar] [CrossRef]
- Rastogi, R.P.; Madamwar, D.; Incharoensakdi, A. Bloom Dynamics of Cyanobacteria and Their Toxins: Environmental Health Impacts and Mitigation Strategies. Front. Microbiol. 2015, 6, 1254. [Google Scholar] [CrossRef] [PubMed]
- Backer, L.C. Cyanobacterial Harmful Algal Blooms (CyanoHABs): Developing a Public Health Response. Lake Reserv. Manag. 2002, 18, 20–31. [Google Scholar] [CrossRef]
- Carmichael, W.W.; Azevedo, S.M.; An, J.S.; Molica, R.J.; Jochimsen, E.M.; Lau, S.; Rinehart, K.L.; Shaw, G.R.; Eaglesham, G.K. Human fatalities from cyanobacteria: Chemical and biological evidence for cyanotoxins. Environ. Health Perspect. 2001, 109, 663–668. [Google Scholar] [CrossRef]
- Chun, S.-J.; Cui, Y.; Lee, C.S.; Cho, A.R.; Baek, K.; Choi, A.; Ko, S.-R.; Lee, H.-G.; Hwang, S.; Oh, H.-M.; et al. Characterization of Distinct CyanoHABs-Related Modules in Microbial Recurrent Association Network. Front. Microbiol. 2019, 10, 1637. [Google Scholar] [CrossRef]
- Drobac, D.; Tokodi, N.; Simeunović, J.; Baltić, V.; Stanić, D.; Svirčev, Z. Human Exposure to Cyanotoxins and their Effects on Health. Arch. Ind. Hyg. Toxicol. 2013, 64, 305–316. [Google Scholar] [CrossRef]
- Buratti, F.M.; Manganelli, M.; Vichi, S.; Stefanelli, M.; Scardala, S.; Testai, E.; Funari, E. Cyanotoxins: Producing organisms, occurrence, toxicity, mechanism of action and human health toxicological risk evaluation. Arch. Toxicol. 2017, 91, 1049–1130. [Google Scholar] [CrossRef]
- He, X.; Liu, Y.L.; Conklin, A.; Westrick, J.; Weavers, L.K.; Dionysiou, D.D.; Lenhart, J.J.; Mouser, P.J.; Szlag, D.; Walker, H.W. Toxic cyanobacteria and drinking water: Impacts, detection, and treatment. Harmful Algae 2016, 54, 174–193. [Google Scholar] [CrossRef]
- Aguilera, A.; Gómez, E.B.; Kaštovský, J.; Echenique, R.O.; Salerno, G.L. The polyphasic analysis of two native Raphidiopsis isolates supports the unification of the genera Raphidiopsis and Cylindrospermopsis (Nostocales, Cyanobacteria). Phycologia 2018, 57, 130–146. [Google Scholar] [CrossRef]
- Bittencourt-Oliveira, M.D.C.; Piccin-Santos, V.; Moura, A.N.; Aragão-Tavares, N.K.C.; Cordeiro-Araújo, M.K. Cyanobacteria, microcystins and cylindrospermopsin in public drinking supply reservoirs of Brazil. Anais da Academia Brasileira de Ciências 2014, 86, 297–309. [Google Scholar] [CrossRef]
- Park, H.-K.; Kwon, M.-A.; Lee, H.-J.; Oh, J.; Lee, S.-H.; Kim, I.-S.; Park, H.-K.; Kwon, M.-A.; Lee, H.-J.; Oh, J.; et al. Molecular Verification of Bloom-forming Aphanizomenon flos-aquae and Their Secondary Metabolites in the Nakdong River. Int. J. Environ. Res. Public Health 2018, 15, 1739. [Google Scholar] [CrossRef] [PubMed]
- Svirčev, Z.; Drobac, D.; Tokodi, N.; Mijović, B.; Codd, G.A.; Meriluoto, J. Toxicology of microcystins with reference to cases of human intoxications and epidemiological investigations of exposures to cyanobacteria and cyanotoxins. Arch. Toxicol. 2017, 91, 621–650. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring, and Management; Chorus, I., Bartram, J., Eds.; CRC Press: Boca Raton, FL, USA, 1999; ISBN 0419239308. [Google Scholar]
- Conselho Nacional do Meio Ambiente. Resolução n° 357, de 17 de março de 2005; Resolução n° 357, DOU n° 053; Conselho Nacional Do Meio Ambiente: Brasília, Brazil, 2005. [Google Scholar]
- World Health Organization. Guidelines for Drinking-Water Quality, 4th ed.; World Health Organization, Ed.; World Health Organization: Geneva, Switzerland, 2017; ISBN 9789241549950. [Google Scholar]
- Ministério da Saúde. Portaria de Consolidação n° 5, de 28 de Setembro de 2017; Ministério da Saúde: Bebedouro, Brazil, 2017. [Google Scholar]
- Janse, I.; Kardinaal, W.E.A.; Meima, M.; Fastner, J.; Visser, P.M.; Zwart, G. Toxic and nontoxic microcystis colonies in natural populations can be differentiated on the basis of rRNA gene internal transcribed spacer diversity. Appl. Environ. Microbiol. 2004, 70, 3979–3987. [Google Scholar] [CrossRef] [PubMed]
- Al-Tebrineh, J.; Mihali, T.K.; Pomati, F.; Neilan, B.A. Detection of saxitoxin-producing cyanobacteria and Anabaena circinalis in environmental water blooms by quantitative PCR. Appl. Environ. Microbiol. 2010, 76, 7836–7842. [Google Scholar] [CrossRef]
- Al-Tebrineh, J.; Merrick, C.; Ryan, D.; Humpage, A.; Bowling, L.; Neilan, B.A. Community composition, toxigenicity, and environmental conditions during a cyanobacterial bloom occurring along 1100 kilometers of the Murray River. Appl. Environ. Microbiol. 2012, 78, 263–272. [Google Scholar] [CrossRef]
- dos Anjos, F.M.; do Bittencourt-Oliveira, M.C.; Zajac, M.P.; Hiller, S.; Christian, B.; Erler, K.; Luckas, B.; Pinto, E. Detection of harmful cyanobacteria and their toxins by both PCR amplification and LC-MS during a bloom event. Toxicon 2006, 48, 239–245. [Google Scholar] [CrossRef]
- Baker, J.A.; Entsch, B.; Neilan, B.A.; McKay, D.B. Monitoring changing toxigenicity of a cyanobacterial bloom by molecular methods. Appl. Environ. Microbiol. 2002, 68, 6070–6076. [Google Scholar] [CrossRef]
- Belykh, O.I.; Gladkikh, A.S.; Sorokovikova, E.G.; Tikhonova, I.V.; Potapov, S.A.; Butina, T.V. Saxitoxin-Producing cyanobacteria in Lake Baikal. Contemp. Probl. Ecol. 2015, 8, 186–192. [Google Scholar] [CrossRef]
- Sidelev, S.I.; Golokolenova, T.B.; Chernova, E.N.; Russkikh, Y.V. Analysis of phytoplankton in Tsimlyansk Reservoir (Russia) for the presence of cyanobacterial hepato- and neurotoxins. Microbiology 2015, 84, 828–837. [Google Scholar] [CrossRef]
- Kurmayer, R.; Sivonen, K.; Wilmotte, A.; Salmaso, N. Molecular Tools for the Detection and Quantification of Toxigenic Cyanobacteria, 1st ed.; Wiley: Hoboken, NJ, USA, 2017; pp. 163–203. [Google Scholar]
- do Bittencourt-Oliveira, M.C.; Piccin-Santos, V.; Kujbida, P.; do Moura, A.N. Cylindrospermopsin in Water Supply Reservoirs in Brazil Determined by Immunochemical and Molecular Methods. J. Water Resour. Prot. 2011, 03, 349–355. [Google Scholar] [CrossRef]
- Hoff-Risseti, C.; Dörr, F.A.; Schaker, P.D.C.; Pinto, E.; Werner, V.R.; Fiore, M.F. Cylindrospermopsin and Saxitoxin Synthetase Genes in Cylindrospermopsis raciborskii Strains from Brazilian Freshwater. PLoS ONE 2013, 8, e74238. [Google Scholar] [CrossRef] [PubMed]
- Barón-Sola, Á.; Ouahid, Y.; del Campo, F.F. Detection of potentially producing cylindrospermopsin and microcystin strains in mixed populations of cyanobacteria by simultaneous amplification of cylindrospermopsin and microcystin gene regions. Ecotoxicol. Environ. Saf. 2012, 75, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Jia, R.; Liu, Y.; Zhang, H. Detection of Microcystin-Producing Cyanobacteria in a Reservoir by Whole Cell Quantitative PCR. Procedia Environ. Sci. 2011, 10, 2272–2279. [Google Scholar] [CrossRef][Green Version]
- Macário, I.P.E.; Castro, B.B.; Nunes, I.M.S.; Pizarro, C.; Coelho, C.; Gonçalves, F.; de Figueiredo, D.R. Stepwise strategy for monitoring toxic cyanobacterial blooms in lentic water bodies. Environ. Monit. Assess. 2017, 189, 620. [Google Scholar] [CrossRef] [PubMed]
- Legrand, B.; Lesobre, J.; Colombet, J.; Latour, D.; Sabart, M. Molecular tools to detect anatoxin-a genes in aquatic ecosystems: Toward a new nested PCR-based method. Harmful Algae 2016, 58, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Lorenzi, A.S.; Chia, M.A.; Piccin-Santos, V.; Do, M.; Bittencourt-Oliveira, C. Microcystins and cylindrospermopsins molecular markers for the detection of toxic cyanobacteria: A case study of northeastern Brazilian reservoirs. Limnetica 2011, 29, 269–282. [Google Scholar]
- Zhang, W.; Lou, I.; Ung, W.K.; Kong, Y.; Mok, K.M. Analysis of cylindrospermopsin- and microcystin-producing genotypes and cyanotoxin concentrations in the Macau storage reservoir. Hydrobiologia 2014, 741, 51–68. [Google Scholar] [CrossRef]
- Gkelis, S.; Papadimitriou, T.; Zaoutsos, N.; Leonardos, I. Anthropogenic and climate-induced change favors toxic cyanobacteria blooms: Evidence from monitoring a highly eutrophic, urban Mediterranean lake. Harmful Algae 2014, 39, 322–333. [Google Scholar] [CrossRef]
- Gkelis, S.; Zaoutsos, N. Cyanotoxin occurrence and potentially toxin producing cyanobacteria in freshwaters of Greece: A multi-disciplinary approach. Toxicon 2014, 78, 1–9. [Google Scholar] [CrossRef]
- Magana Arachchi, D.N.; Liyanage, H.M. Determining the presence of cyanotoxins in water reservoirs of anuradhapura, using molecular and bioassay methods. J. Natl. Sci. Found. Sri Lanka 2012, 40, 156–166. [Google Scholar] [CrossRef]
- De Souza, R.C.R.; Carvalho, M.C.; Truzzi, A.C. Cylindrospermopsis raciborskii (Wolosz.) Seenaya and Subba Raju (Cyanophyceae) dominance and a contribution to the knowledge of Rio Pequeno Arm, Billings Reservoir, Brazil. Environ. Toxicol. Water Qual. 1998, 13, 73–81. [Google Scholar] [CrossRef]
- de Carvalho, L.R.; Sant’Anna, C.L.; Gemelgo, M.C.P.; de P. Azevedo, M.T. Cyanobacterial occurrence and detection of microcystin by planar chromatography in surface water of Billings and Guarapiranga Reservoirs, SP, Brazil. Rev. Bras. Botânica 2007, 30, 141–148. [Google Scholar] [CrossRef][Green Version]
- Gemelgo, M.; Mucci, J.; Navas-Pereira, D. Population dynamics: Seasonal variation of phytoplankton functional groups in brazilian reservoirs (Billings and Guarapiranga, São Paulo). Braz. J. Biol. 2009, 69, 1001–1013. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Gemelgo, M.C.P.; Sant’Anna, C.L.; Tucci, A.; Barbosa, H.R. Population dynamics of Cylindrospermopsis raciborskii (Woloszynska) Seenayya & Subba Raju, a Cyanobacteria toxic species, in watersupply reservoirs in São Paulo, Brazil. Hoehnea 2008, 35, 297–307. [Google Scholar]
- Piccin-Santos, V.; do Bittencourt-Oliveira, M.C. Toxic Cyanobacteria in Four Brazilian Water Supply Reservoirs. J. Environ. Prot. 2012, 3, 68–73. [Google Scholar] [CrossRef]
- Sant’Anna, C.L.; Melcher, S.S.; Carvalho, M.D.; Gelmego, M.P.; Azevedo, M.T. Planktic cyanobacteria from upper Tietê basin reservoirs, SP, Brazil. Rev. Bras. Bot. 2007, 30, 1–17. [Google Scholar] [CrossRef]
- Companhia Ambiental do Estado de São Paulo. Qualidade das Águas Interiores no Estado de São Paulo 2018; São Paulo State Inland Water Quality Report; CETESB: São Paulo, Brazil, 2019. [Google Scholar]
- Frias, H.V.; Mendes, M.A.; Cardozo, K.H.M.; Carvalho, V.M.; Tomazela, D.; Colepicolo, P.; Pinto, E. Use of electrospray tandem mass spectrometry for identification of microcystins during a cyanobacterial bloom event. Biochem. Biophys. Res. Commun. 2006, 344, 741–746. [Google Scholar] [CrossRef]
- Moschini-Carlos, V.; Bortoli, S.; Pinto, E.; Nishimura, P.Y.; De Freitas, L.G.; Pompêo, M.L.M.; Dörr, F. Cyanobacteria and cyanotoxin in the billings reservoir (São Paulo, SP, Brazil). Limnetica 2009, 28, 273–282. [Google Scholar]
- Assembléia Legislativa do Estado de São Paulo. Lei No 13.579, de 13 de Julho de 2009; Assembléia Legislativa do Estado de São Paulo: São Paulo, Brazil, 2009; pp. 1–30.
- Companhia Ambiental do Estado de São Paulo. Qualidade das Águas Interiores no Estado de São Paulo 2016; São Paulo State Inland Water Quality Report; CETESB: São Paulo, Brazil, 2017. [Google Scholar]
- Companhia Ambiental do Estado de São Paulo. Qualidade das Águas Interiores no Estado de São Paulo 2010; São Paulo State Inland Water Quality Report; CETESB: São Paulo, Brazil, 2011. [Google Scholar]
- Companhia Ambiental do Estado de São Paulo. Qualidade das Águas Interiores no Estado de São Paulo 2011; São Paulo State Inland Water Quality Report; CETESB: São Paulo, Brazil, 2012. [Google Scholar]
- Companhia Ambiental do Estado de São Paulo. Qualidade das Águas Interiores no Estado de São Paulo 2012; São Paulo State Inland Water Quality Report; CETESB: São Paulo, Brazil, 2013. [Google Scholar]
- Companhia Ambiental do Estado de São Paulo. Qualidade das Águas Interiores no Estado de São Paulo 2013; São Paulo State Inland Water Quality Report; CETESB: São Paulo, Brazil, 2014. [Google Scholar]
- Companhia Ambiental do Estado de São Paulo. Qualidade das Águas Interiores no Estado de São Paulo 2014; São Paulo State Inland Water Quality Report; CETESB: São Paulo, Brazil, 2015. [Google Scholar]
- Companhia Ambiental do Estado de São Paulo. Qualidade das Águas Interiores no Estado de São Paulo 2015; São Paulo State Inland Water Quality Report; CETESB: São Paulo, Brazil, 2016. [Google Scholar]
- Companhia Ambiental do Estado de São Paulo. Qualidade das Águas Interiores no Estado de São Paulo 2017; São Paulo State Inland Water Quality Report; CETESB: São Paulo, Brazil, 2018. [Google Scholar]
- Neilan, B.A.; Jacobs, D.; Goodman, A.E. Genetic diversity and phylogeny of toxic cyanobacteria determined by DNA polymorphisms within the phycocyanin locus. Appl. Environ. Microbiol. 1995, 61, 3875–3883. [Google Scholar] [CrossRef]
- Kellmann, R.; Mills, T.; Neilan, B.A. Functional modeling and phylogenetic distribution of putative cylindrospermopsin biosynthesis enzymes. J. Mol. Evol. 2006, 62, 267–280. [Google Scholar] [CrossRef]
- Mazmouz, R.; Chapuis-Hugon, F.; Mann, S.; Pichon, V.; Méjean, A.; Ploux, O. Biosynthesis of cylindrospermopsin and 7-epicylindrospermopsin in oscillatoria sp. strain PCC 6506: Identification of the cyr gene cluster and toxin analysis. Appl. Environ. Microbiol. 2010, 76, 4943–4949. [Google Scholar] [CrossRef]
- Rantala, A.; Fewer, D.P.; Hisbergues, M.; Rouhiainen, L.; Vaitomaa, J.; Borner, T.; Sivonen, K. Phylogenetic evidence for the early evolution of microcystin synthesis. Proc. Natl. Acad. Sci. USA 2004, 101, 568–573. [Google Scholar] [CrossRef] [PubMed]
- Laughinghouse, H.D.; Prá, D.; Silva-Stenico, M.E.; Rieger, A.; Frescura, V.D.S.; Fiore, M.F.; Tedesco, S.B. Biomonitoring genotoxicity and cytotoxicity of Microcystis aeruginosa (Chroococcales, Cyanobacteria) using the Allium cepa test. Sci. Total Environ. 2012, 432, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Bolch, C.J.S.; Blackburn, S.I. Isolation and purification of Australian isolates of the toxic cyanobacterium Microcystis aeruginosa Kütz. J. Appl. Phycol. 1996, 8, 5–13. [Google Scholar] [CrossRef]
- Rosini, E.F.; Tucci, A.; do Carmo, C.F.; Rojas, N.E.T.; de Barros, H.P.; Mallasen, M. Changes in phytoplankton spatial and temporal dynamics in a Brazilian tropical oligotrophic reservoir after net cage installation. Braz. J. Bot. 2016, 39, 569–581. [Google Scholar] [CrossRef]
- Utermöhl, H. Zur Vervollkommnung der quantitativen Phytoplankton-Methodik. SIL Commun. 1958, 9, 1–38. [Google Scholar] [CrossRef]
- Fonseca, B.M.; Ferragut, C.; Tucci, A.; Crossetti, L.O.; Ferrari, F.; de Bicudo, D.C.; Sant’Anna, C.L.; de Bicudo, C.E.M. Biovolume de cianobactérias e algas de reservatórios tropicais do Brasil com diferentes estados tróficos. Hoehnea 2014, 41, 9–30. [Google Scholar] [CrossRef]
- Hillebrand, H.; Dürselen, C.-D.; Kirschtel, D.; Pollingher, U.; Zohary, T. Biovolume calculation for pelagic and benthic microalgae. J. Phycol. 1999, 35, 403–424. [Google Scholar] [CrossRef]
- World Health Organization. Guidelines for Safe Recreational Water Environments; WHO Library Cats Data; World Health Organization: Geneva, Switzerland, 2003; Volume 1, pp. 3505–3518. [Google Scholar]
- Mihali, T.K.; Kellmann, R.; Muenchhoff, J.; Barrow, K.D.; Neilan, B.A. Characterization of the gene cluster responsible for cylindrospermopsin biosynthesis. Appl. Environ. Microbiol. 2008, 74, 716–722. [Google Scholar] [CrossRef]
- Mihali, T.K.; Kellmann, R.; Neilan, B.A. Characterisation of the paralytic shellfish toxin biosynthesis gene clusters in Anabaena circinalis AWQC131C and Aphanizomenon sp. NH-5. BMC Biochem. 2009, 10, 8. [Google Scholar] [CrossRef]
- Tillett, D.; Dittmann, E.; Erhard, M.; Von Döhren, H.; Börner, T.; Neilan, B.A. Structural organization of microcystin biosynthesis in Microcystis aeruginosa PCC7806: An integrated peptide-polyketide synthetase system. Chem. Biol. 2000, 7, 753–764. [Google Scholar] [CrossRef]
- Dittmann, E.; Fewer, D.P.; Neilan, B.A. Cyanobacterial toxins: Biosynthetic routes and evolutionary roots. FEMS Microbiol. Rev. 2013, 37, 23–43. [Google Scholar] [CrossRef] [PubMed]
- Mankiewicz-Boczek, J.; Izydorczyk, K.; Romanowska-Duda, Z.; Jurczak, T.; Stefaniak, K.; Kokocinski, M. Detection and monitoring toxigenicity of cyanobacteria by application of molecular methods. Environ. Toxicol. 2006, 21, 380–387. [Google Scholar] [CrossRef] [PubMed]
- Rantala, A.; Rajaniemi-Wacklin, P.; Lyra, C.; Lepistö, L.; Rintala, J.; Mankiewicz-Boczek, J.; Sivonen, K. Detection of microcystin-producing cyanobacteria in Finnish lakes with genus-specific microcystin synthetase gene E (mcyE) PCR and associations with environmental factors. Appl. Environ. Microbiol. 2006, 72, 6101–6110. [Google Scholar] [CrossRef] [PubMed]
- Kellmann, R.; Mihali, T.K.; Jeon, Y.J.; Pickford, R.; Pomati, F.; Neilan, B.A. Biosynthetic intermediate analysis and functional homology reveal a saxitoxin gene cluster in cyanobacteria. Appl. Environ. Microbiol. 2008, 74, 4044–4053. [Google Scholar] [CrossRef] [PubMed]
- Bober, B.; Bialczyk, J. Determination of the toxicity of the freshwater cyanobacterium Woronichinia naegeliana (Unger) Elenkin. J. Appl. Phycol. 2017, 29, 1355–1362. [Google Scholar] [CrossRef]
- Voloshko, L.; Kopecky, J.; Safronova, T.; Pljusch, A.; Titova, N.; Hrouzek, P.; Drabkova, V. Toxins and other bioactive compounds produced by cyanobacteria in Lake Ladoga. Est. J. Ecol. 2008, 57, 100–110. [Google Scholar] [CrossRef]
- Willame, R.; Jurczak, T.; Iffly, J.-F.; Kull, T.; Meriluoto, J.; Hoffmann, L. Distribution of Hepatotoxic Cyanobacterial Blooms in Belgium and Luxembourg. Hydrobiologia 2005, 551, 99–117. [Google Scholar] [CrossRef]
- Barboza, G.; Gorlach-Lira, K.; Sassi, C.; Sassi, R. Microcystins production and antibacterial activity of cyanobacterial strains of Synechocystis, Synechococcus and Romeria isolated from water and coral reef organisms of Brazilian coast. Rev. Biol. Trop. 2017, 65, 890–899. [Google Scholar] [CrossRef][Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Primer | Sequence (5′–3′) | AT (°C) | Fragment Size (bp) | Reference |
|---|---|---|---|---|---|
| cpc | PCβF | GGCTGCTTGTTTACGCGACA | 50 | 500–700 | [54] |
| PCαR | CCAGTACCACCAGCAACTAA | ||||
| cyrA | CatF1 | AGATGGTGCTTATTTTGAAC | 54 | 881 | [27] |
| CatR1 | TCTTCACAGATGACCTTCTT | ||||
| cyrB | CPS-F | AGTATATGTTGCGGGACTCG | 55 | 478 | [55] |
| CPS-R | CCCGCCAAGACAGAGGGTAG | ||||
| cyrC | CkcF3 | AATGATCGAAAACAGCAGTCGG | 54 | 325 | [27] |
| CkcR3 | TAGAACAATCATCCCACAACCT | ||||
| cyrJ | cyrJ-F | TTCTCTCCTTTCCCTATCTCTTTATCT | 53 | 536 | [26,56] * |
| cyrJ-R | GCTACGGTGCTGTACCAAGGGGC | ||||
| mcyE | mcyE-F2 | GAAATTTGTGTAGAAGGTGC | 56 | 809–812 | [57] |
| mcyE-R4 | AATTCTAAAGCCCAAAGACG | ||||
| sxtA | sxtA-F | GATGACGGAGTATTTGAAGC | 55 | 125 | [18] |
| sxtA-R | CTGCATCTTCTGGACGGTAA | ||||
| sxtB | SXTB-F | TTTGTAGGRCAGGCACTT | 53 | 400 | [26] |
| SXTB-R | ATCATCGGTATCATCGGTA | ||||
| sxtI | OCT-F | TGCCGTTTTGTGCTTAGATG | 61 | 923 | [26] |
| OCT-R | GGACGGAAGGACTCACGATA |
| Gene | DRY 1 | DRY 2 | RAINY 1 | RAINY 2 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SP1 | SP2 | SP3 | SP4 | SP1 | SP2 | SP3 | SP4 | SP1 | SP2 | SP3 | SP4 | SP1 | SP2 | SP3 | SP4 | |
| cpc | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| mcyE | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| cyrA | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| cyrB | + | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| cyrC | + | + | + | + | - | - | - | - | - | - | - | - | + | + | - | - |
| cyrJ | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| sxtA | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| sxtB | + | - | + | + | - | - | - | - | - | - | - | - | - | - | - | - |
| sxtI | - | - | - | - | - | - | + | - | - | - | - | - | - | - | + | + |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ribeiro, M.S.F.; Tucci, A.; Matarazzo, M.P.; Viana-Niero, C.; Nordi, C.S.F. Detection of Cyanotoxin-Producing Genes in a Eutrophic Reservoir (Billings Reservoir, São Paulo, Brazil). Water 2020, 12, 903. https://doi.org/10.3390/w12030903
Ribeiro MSF, Tucci A, Matarazzo MP, Viana-Niero C, Nordi CSF. Detection of Cyanotoxin-Producing Genes in a Eutrophic Reservoir (Billings Reservoir, São Paulo, Brazil). Water. 2020; 12(3):903. https://doi.org/10.3390/w12030903
Chicago/Turabian StyleRibeiro, Matheus S. F., Andrea Tucci, Matheus P. Matarazzo, Cristina Viana-Niero, and Cristina S. F. Nordi. 2020. "Detection of Cyanotoxin-Producing Genes in a Eutrophic Reservoir (Billings Reservoir, São Paulo, Brazil)" Water 12, no. 3: 903. https://doi.org/10.3390/w12030903
APA StyleRibeiro, M. S. F., Tucci, A., Matarazzo, M. P., Viana-Niero, C., & Nordi, C. S. F. (2020). Detection of Cyanotoxin-Producing Genes in a Eutrophic Reservoir (Billings Reservoir, São Paulo, Brazil). Water, 12(3), 903. https://doi.org/10.3390/w12030903