Cover or Velocity: What Triggers Iberian Barbel (Luciobarbus Bocagei) Refuge Selection under Experimental Hydropeaking Conditions?


 , ,
, ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Fish Sampling and Handling
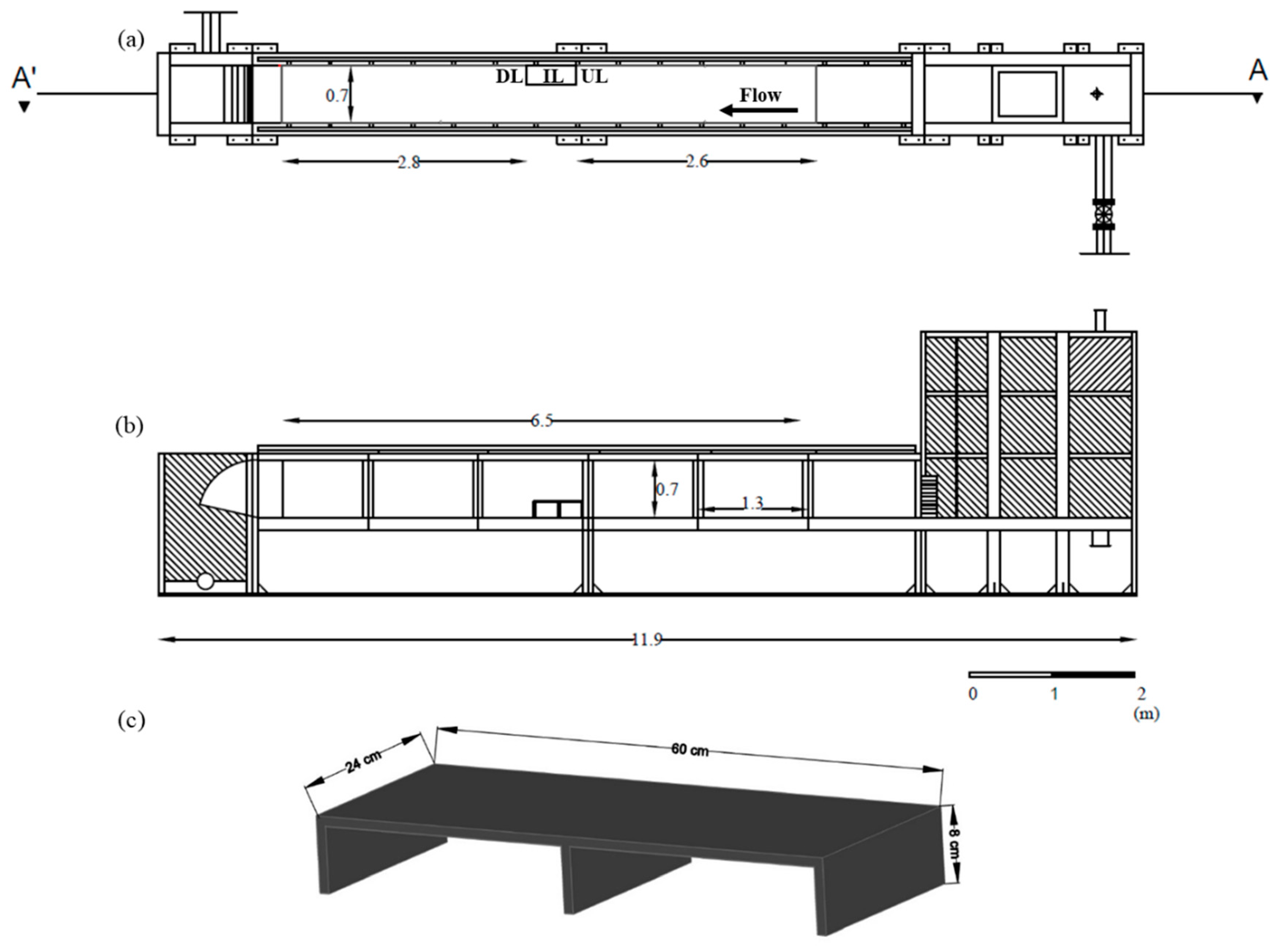
2.2. Experimental Facilities
2.3. Flow Events
2.4. Physiological Responses
2.5. Structure Use
3. Results
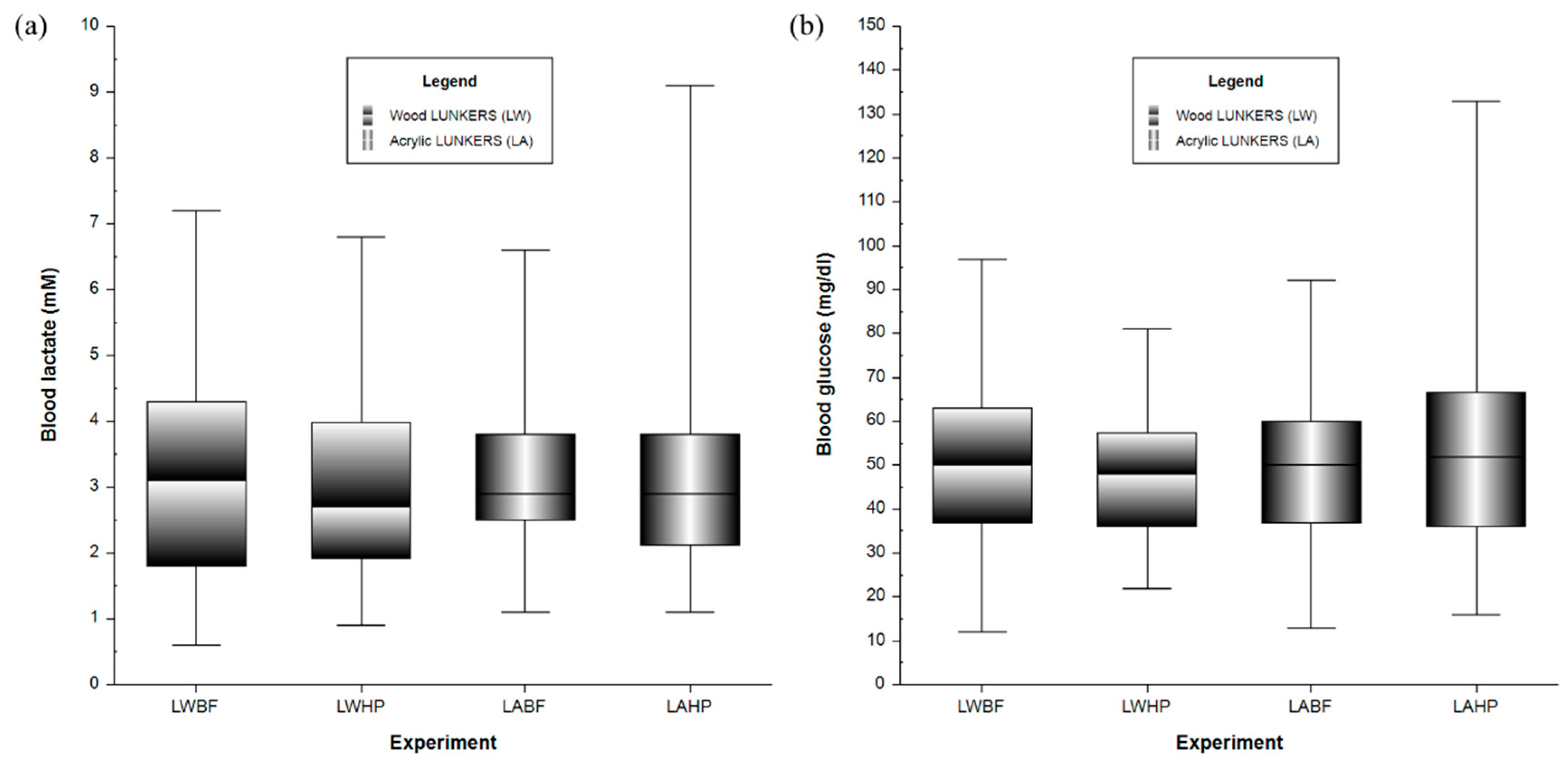
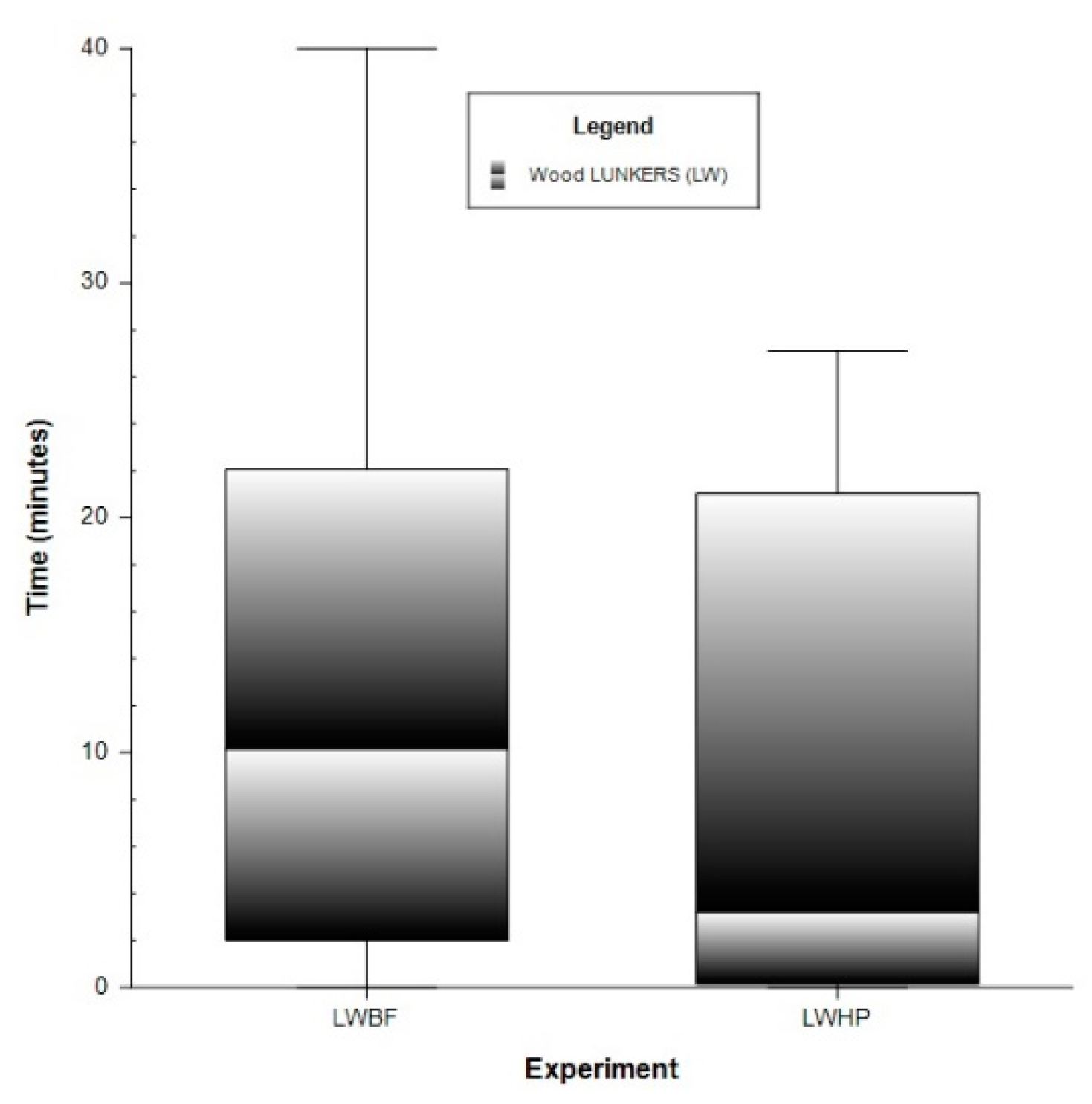
3.1. Physiological Responses
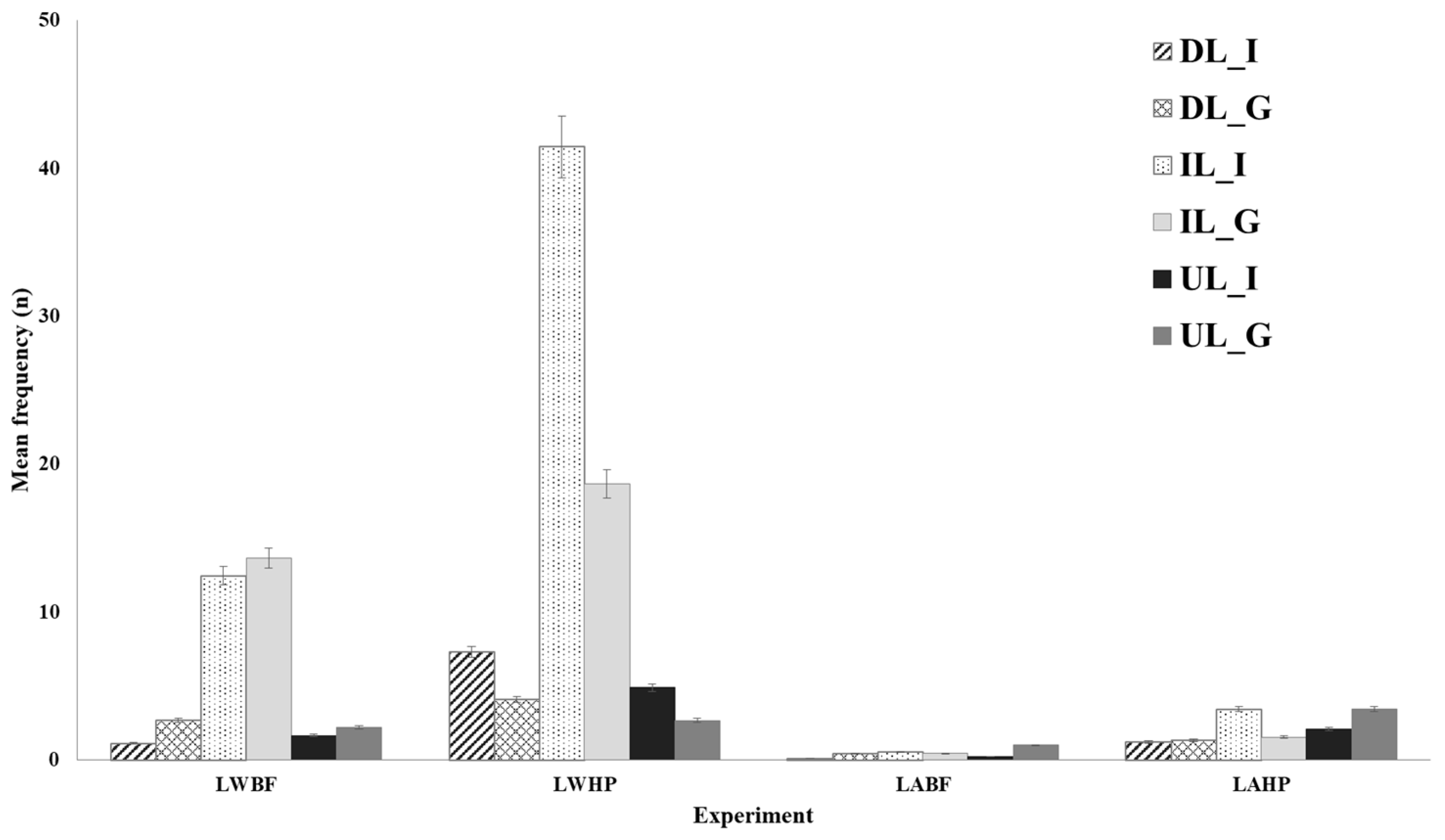
3.2. Structure Use
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Ethical Statement
References
- Moog, O. Quantification of daily peak hydropower effects on aquatic fauna and management to minimize environmental impacts. Regul. Rivers Res. Manag. 1993, 8, 5–14. [Google Scholar] [CrossRef]
- Shuster, W.D.; Zhang, Y.; Roy, A.H.; Daniel, F.B.; Troyer, M. Characterizing storm hydrograph rise and fall dynamics with stream stage data. J. Am. Water Res. Assoc. 2008, 44, 1431–1440. [Google Scholar] [CrossRef]
- Hauer, C.; Unfer, G.; Holzapfel, P.; Haimann, M.; Habersack, H. Impact of channel bar form and grain size variability on estimated stranding risk of juvenile brown trout during hydropeaking. Earth Surf. Process. Landf. 2014, 39. [Google Scholar] [CrossRef]
- Hauer, C.; Holzapfel, P.; Leitner, P.; Graf, W. Longitudinal assessment of hydropeaking impacts on various scales for an improved process understanding and the design of mitigation measures. Sci. Total Environ. 2017, 575, 1503–1514. [Google Scholar] [CrossRef]
- Vanzo, D.; Zolezzi, G.; Siviglia, A. Eco-hydraulic modelling of the interactions between hydropeaking and river morphology. Ecohydrology 2016, 9, 421–437. [Google Scholar] [CrossRef]
- Moreira, M.; Hayes, D.S.; Boavida, I.; Schletterer, M.; Schmutz, S.; Pinheiro, A. Ecologically-based criteria for hydropeaking mitigation: A review. Sci. Total Environ. 2019, 657, 1508–1522. [Google Scholar] [CrossRef]
- Parasiewicz, P.; Schmutz, S.; Moog, O. The effects of managed hydropower peaking on the physical habitat, benthos and fish fauna in the river Bregenzerach in Austria. Fish. Manag. Ecol. 1998, 5, 403–417. [Google Scholar] [CrossRef]
- Tuhtan, J.A.; Noack, M.; Wieprecht, S. Estimating stranding risk due to hydropeaking for juvenile European grayling considering river morphology. KSCE J. Civ. Eng. 2012, 16, 197–206. [Google Scholar] [CrossRef]
- Young, P.S.; Cech, J.J.; Thompson, L.C. Hydropower-related pulsed-flow impacts on stream fishes: A brief review, conceptual model, knowledge gaps, and research needs. Rev. Fish Biol. Fish. 2011, 21, 713–731. [Google Scholar] [CrossRef]
- Hayes, D.; Moreira, M.; Boavida, I.; Haslauer, M.; Unfer, G.; Zeiringer, B.; Greimel, F.; Auer, S.; Ferreira, T.; Schmutz, S. Life stage-specific hydropeaking flow rules. Sustainability 2019, 11, 1547. [Google Scholar] [CrossRef]
- Greimel, F.; Schülting, L.; Graf, W.; Bondar-Kunze, E.; Auer, S.; Zeiringer, B.; Hauer, C. Hydropeaking impacts and mitigation. In Riverine Ecosystem Management; Springer: Cham, Switzerland, 2018; pp. 91–110. [Google Scholar] [CrossRef]
- Bruder, A.; Tonolla, D.; Schweizer, S.P.; Vollenweider, S.; Langhans, S.D.; Wuest, A. A conceptual framework for hydropeaking mitigation. Sci. Total Environ. 2016, 568, 1204–1212. [Google Scholar] [CrossRef] [PubMed]
- Premstaller, G.; Cavedon, V.; Pisaturo, G.R.; Schweizer, S.; Adami, V.; Righetti, M. Hydropeaking mitigation project on a multi-purpose hydro-scheme on Valsura River in South Tyrol/Italy. Sci. Total Environ. 2017, 574, 642–653. [Google Scholar] [CrossRef] [PubMed]
- Ribi, J.M.; Boillat, J.L.; Peter, A.; Schleiss, A.J. Attractiveness of a lateral shelter in a channel as a refuge for juvenile brown trout during hydropeaking. Aquat. Sci. 2014, 76, 527–541. [Google Scholar] [CrossRef]
- Vehanen, T.; Bjerke, P.L.; Heggenes, J.; Huusko, A.; Maki-Petays, A. Effect of fluctuating flow and temperature on cover type selection and behaviour by juvenile brown trout in artificial flumes. J. Fish Biol. 2000, 56, 923–937. [Google Scholar] [CrossRef]
- Costa, M.J.; Boavida, I.; Almeida, V.; Cooke, S.J.; Pinheiro, A.N. Do artificial velocity refuges mitigate the physiological and behavioural consequences of hydropeaking on a freshwater Iberian cyprinid? Ecohydrology 2018, 11. [Google Scholar] [CrossRef]
- Chun, S.N.; Cocherell, S.A.; Cocherell, D.E.; Miranda, J.B.; Jones, G.J.; Graham, J.; Klimley, A.P.; Thompson, L.C.; Cech, J.J. Displacement, velocity preference, and substrate use of three native California stream fishes in simulated pulsed flows. Environ. Biol. Fish. 2010, 90, 43–52. [Google Scholar] [CrossRef]
- Scruton, D.A.; Pennell, C.; Ollerhead, L.M.N.; Alfredsen, K.; Stickler, M.; Harby, A.; Robertson, M.; Clarke, K.D.; LeDrew, L.J. A synopsis of ’hydropeaking’ studies on the response of juvenile Atlantic salmon to experimental flow alteration. Hydrobiologia 2008, 609, 263–275. [Google Scholar] [CrossRef]
- Korman, J.; Campana, S.E. Effects of hydropeaking on nearshore habitat use and growth of age-0 rainbow trout in a large regulated river. In Transactions of the American Fisheries Society; American Fisheries Society: Bethesda, MD, USA, 2011; Volume 138, pp. 76–87. [Google Scholar] [CrossRef]
- Noatch, M.R.; Suski, C.D. Non-physical barriers to deter fish movements. Environ. Rev. 2012, 20, 71–82. [Google Scholar] [CrossRef]
- Jesus, J.; Teixeira, A.; Natário, S.; Cortes, R. Repulsive effect of stroboscopic light barriers on native salmonid (Salmo trutta) and Cyprinid (Pseudochondrostoma duriense and Luciobarbus bocagei) species of Iberia. Sustainability 2019, 11, 1332. [Google Scholar] [CrossRef]
- Liao, J.C. A review of fish swimming mechanics and behaviour in altered flows. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2007, 362, 1973–1993. [Google Scholar] [CrossRef]
- Pankhurst, N.W. The endocrinology of stress in fish: An environmental perspective. Gen. Comp. Endocrinol. 2011, 170, 265–275. [Google Scholar] [CrossRef]
- Costa, M.J.; Lennox, R.J.; Katopodis, C.; Cooke, S.J. Is there evidence for flow variability as an organism-level stressor in fluvial fish? J. Ecohydraul. 2017, 2, 68–83. [Google Scholar] [CrossRef]
- Costa, M.J.; Fuentes-Perez, J.F.; Boavida, I.; Tuhtan, J.A.; Pinheiro, A.N. Fish under pressure: Examining behavioural responses of Iberian barbel under simulated hydropeaking with instream structures. PLoS ONE 2019, 14, e0211115. [Google Scholar] [CrossRef] [PubMed]
- Iowa Department of Natural Resources. River Restoration Toolbox Practice Guide 6—Aquatic Habitat/Cover Features; Iowa Department of Natural Resources: Des Moines, IA, USA, 2018. Available online: https://www.iowadnr.gov/Portals/idnr/uploads/RiverRestoration/toolbox/aquatic (accessed on 17 January 2020).
- United States Department of Agriculture. Stream Habitat Enhancement Using LUNKERS; Natural Resources Conservation Service: Washington, DC, USA, 2007; Volume 654. Available online: http://directives.sc.egov.usda.gov/OpenNonWebContent.aspx?content=17825.wba (accessed on 17 January 2020).
- Roseboom, D.; Hill, T.; Beardsley, J.; Rodsater, J.; Duong, L. Streambank Stabilization and In-Stream Habitat Enhancement with LUNKERS Bank Revetments; Illinois Department of Natural Resources, Nonpoint Pollution Control Program, Illinois State Water Survey; Conservation Technology Information Center: Indiana, IN, USA, 1998; Available online: https://www.ideals.illinois.edu/bitstream/handle/2142/77748/FMSR1ed_ch4OCRd.pdf?sequence=2 (accessed on 17 January 2020).
- Peake, S.J. Swimming Performance and Behavior of Fish Species Endemic to Newfoundland and Labrador: A Literature Review for The Purpose of Establishing Design and Water Velocity Criteria for Fishways and Culverts; Canadian Manuscript Report of Fisheries and Aquatic Sciences; Fisheries and Oceans Canada: Ottawa, OT, Canada, 2008. [Google Scholar]
- Boavida, I.; Jesus, J.B.; Pereira, V.; Santos, C.; Lopes, M.; Cortes, R.M.V. Fulfilling spawning flow requirements for potamodromous cyprinids in a restored river segment. Sci. Total Environ. 2018, 635, 567–575. [Google Scholar] [CrossRef] [PubMed]
- White, W.; Beardsley, J.; Tomkins, S. Waukegan River Illinois National Nonpoint Source Monitoring Program Project; Illinois State Water Survey, Institute of Natural Resource Sustainability, University of Illinois at Urbana-Champaign: Champaign, IL, USA, 2011; Available online: https://www.isws.illinois.edu/pubdoc/CR/ISWSCR2011-01.pdf (accessed on 17 January 2020).
- Costa, M.J.; Ferreira, M.T.; Pinheiro, A.N.; Boavida, I. The potential of lateral refuges for Iberian barbel under simulated hydropeaking conditions. Ecol. Eng. 2019, 127, 567–578. [Google Scholar] [CrossRef]
- Oliveira, J.M.; Ferreira, A.P.; Ferreira, M.T. Intrabasin variations in age and growth of Barbus bocagei populations. J. Appl. Ichthyol. 2002, 18, 134–139. [Google Scholar] [CrossRef]
- Martínez-Capel, F.; García de Jalón, D. Desarrollo de curvas de preferencia de microhábitat para Leuciscus pyrenaicus y Barbus bocagei por buceo en el río Jarama (Cuenca del Tajo). Limnetica 1999, 17, 71–83. [Google Scholar]
- Rodríguez-Ruiz, A.; Granado-Lorencio, C. Spawning period and migration of three species of cyprinids in a stream with Mediterranean regimen (SW Spain). J. Fish Biol. 1992, 41, 545–556. [Google Scholar] [CrossRef]
- European Committee for Standardization (CEN). Water quality: Sampling of fish with electricity. In European Standard EN 14022:2003; European Comission: Brussels, Belgium, 2003. [Google Scholar]
- INAG. Manual Para a Avaliação Biológica da Qualidade da Água em Sistemas Fluviais Segundo a Directiva Quadro da Água—Protocolo de Amostragem e Análise Para a Fauna Piscícola; Ministério do Ambiente, do Ordenamento do Território e do Desenvolvimento Regional, Instituto da Água: Lisbon, Portugal, 2008. [Google Scholar]
- Ministério do Ambiente, do Ordenamento do Território e do Desenvolvimento Regional. Decreto-Lei n.° 236/98. In Diário da República n.° 176/1998, Série I-A de 1998-08-01; Anexo, X., Portugal, G.D., Eds.; Ministério do Ambiente, do Ordenamento do Território e do Desenvolvimento Regional: Lisbon, Portugal, 1998; Available online: http://data.dre.pt/eli/dec-lei/236/1998/08/01/p/dre/pt/html (accessed on 1 May 2019).
- Harby, A.; Noack, M. Rapid flow fluctuations and impacts on fish and the aquatic ecosystem. In Ecohydraulics—An Integrated Approach; Maddock, I., Harby, A., Kemp, P., Wood, P., Eds.; Wiley-Blackwell: Chichester, UK, 2013; pp. 323–335. [Google Scholar]
- Santos, J.M.; Rivaes, R.; Boavida, I.; Branco, P. Structural microhabitat use by endemic cyprinids in a Mediterranean-type river: Implications for restoration practices. Aquat. Conserv. Mar. Freshw. Ecosyst. 2018, 28, 26–36. [Google Scholar] [CrossRef]
- Mateus, C.S.; Quintella, B.R.; Almeida, P.R. The critical swimming speed of Iberian barbel Barbus bocagei relation to size and sex. J. Fish Biol. 2008, 73, 1783–1789. [Google Scholar] [CrossRef]
- Amaral, S.D.; Branco, P.; da Silva, A.T.; Katopodis, C.; Viseu, T.; Ferreira, M.T.; Pinheiro, A.N.; Santos, J.M. Upstream passage of potamodromous cyprinids over small weirs: The influence of key-hydraulic parameters. J. Ecohydraul. 2016, 1, 79–89. [Google Scholar] [CrossRef]
- Branco, P.; Santos, J.M.; Katopodis, C.; Pinheiro, A.; Ferreira, M.T. Effect of flow regime hydraulics on passage performance of Iberian chub (Squalius pyrenaicus) (Günther, 1868) in an experimental pool-and-weir fishway. Hydrobiologia 2013, 714, 145–154. [Google Scholar] [CrossRef]
- Romão, F.; Santos, J.; Katopodis, C.; Pinheiro, A.; Branco, P. How does season affect passage performance and fatigue of potamodromous cyprinids? An experimental approach in a vertical slot fishway. Water 2018, 10, 395. [Google Scholar] [CrossRef]
- Amaral, S.; Branco, P.; Katopodis, C.; Ferreira, M.; Pinheiro, A.; Santos, J. Passage performance of potamodromous cyprinids over an experimental low-head ramped weir: The effect of ramp length and slope. Sustainability 2019, 11, 1456. [Google Scholar] [CrossRef]
- Weihs, D. Hydromechanics of fish schooling. Nature 1973, 241, 290–291. [Google Scholar] [CrossRef]
- Sumpter, J.P. The endocrinology of stress. In Fish Stress and Health in Aquaculture; Iwama, G.K., Pickering, A.D., Sumpter, J.P., Schreck, C.B., Eds.; Cambridge University Press: Cambridge, UK, 1997; pp. 95–118. [Google Scholar]
- Beecham, R.V.; Small, B.C.; Minchew, C.D. Using portable lactate and glucose meters for catfish research: Acceptable alternatives to established laboratory methods? N. Am. J. Aquac. 2006, 68, 291–295. [Google Scholar] [CrossRef]
- Stoot, L.J.; Cairns, N.A.; Cull, F.; Taylor, J.J.; Jeffrey, J.D.; Morin, F.; Mandelman, J.W.; Clark, T.D.; Cooke, S.J. Use of portable blood physiology point-of-care devices for basic and applied research on vertebrates: A review. Conserv. Physiol. 2014, 2. [Google Scholar] [CrossRef]
- Tanner, R.K.; Fuller, K.L.; Ross, M.L. Evaluation of three portable blood lactate analysers: Lactate pro, lactate scout and lactate plus. Eur. J. Appl. Physiol. 2010, 109, 551–559. [Google Scholar] [CrossRef]
- Zar, J.H. Biostatistical Analysis, 5th ed.; Pearson Prentice Hall: Upper Saddle River, NJ, USA, 2010. [Google Scholar]
- NNCSS 11 Statistical Software. NCSS, LLC: Kaysville, UT, USA, 2016. Available online: http://ncss.com/software/ncss (accessed on 1 May 2019).
- Walters, K.; Coen, L.D. A comparison of statistical approaches to analyzing community convergence between natural and constructed oyster reefs. J. Exp. Mar. Biol. Ecol. 2006, 330, 81–95. [Google Scholar] [CrossRef]
- Branco, P.; Santos, J.M.; Amaral, S.; Romao, F.; Pinheiro, A.N.; Ferreira, M.T. Potamodromous fish movements under multiple stressors: Connectivity reduction and oxygen depletion. Sci. Total Environ. 2016, 572, 520–525. [Google Scholar] [CrossRef]
- Anderson, M.J.; Robinson, J. Permutation tests for linear models. Aust. N. Z. J. Stat. 2001, 43, 75–88. [Google Scholar] [CrossRef]
- Anderson, M.; Gorley, R.N.; Clarke, R.K. Permanova+ for Primer: Guide to Software and Statistical Methods; PRIMER-E Ltd.: Plymouth, UK, 2008. [Google Scholar]
- Flodmark, L. Cortisol and glucose responses in juvenile brown trout subjected to a fluctuating flow regime in an artificial stream. J. Fish Biol. 2002, 60, 238–248. [Google Scholar] [CrossRef]
- Alexandre, C.M.; Almeida, P.R.; Neves, T.; Mateus, C.S.; Costa, J.L.; Quintella, B.R. Effects of flow regulation on the movement patterns and habitat use of a potamodromous cyprinid species. Ecohydrology 2016, 9, 326–340. [Google Scholar] [CrossRef]
- Auer, S.; Zeiringer, B.; Fuhrer, S.; Tonolla, D.; Schmutz, S. Effects of river bank heterogeneity and time of day on drift and stranding of juvenile European grayling (Thymallus thymallus L.) caused by hydropeaking. Sci. Total Environ. 2017, 575, 1515–1521. [Google Scholar] [CrossRef]
- Boavida, I.; Harby, A.; Clarke, K.D.; Heggenes, J. Move or stay: Habitat use and movements by Atlantic salmon parr (Salmo salar) during induced rapid flow variations. Hydrobiologia 2016, 785, 261–275. [Google Scholar] [CrossRef]
- Benitez, J.-P.; Ovidio, M. The influence of environmental factors on the upstream movements of rheophilic cyprinids according to their position in a river basin. Ecol. Freshw. Fish 2018, 27, 660–671. [Google Scholar] [CrossRef]
- Kelly, B.; Smokorowski, K.E.; Power, M. Impact of river regulation and hydropeaking on the growth, condition and field metabolism of Brook Trout (Salvelinus fontinalis). Ecol. Freshw. Fish 2017, 26, 666–675. [Google Scholar] [CrossRef]
- De Vocht, A.; Baras, E. Effect of hydropeaking on migrations and home range of adult Barbel (Barbus barbus) in the river Meuse. In Proceedings of the 5th Conference on Fish Telemetry held in Europe, Ustica, Italy, 9–13 June 2003; pp. 35–44. [Google Scholar]
- Dijkgraaf, S. The functioning and significance of the lateral-line organs. Biol. Rev. 1963, 38, 51–105. [Google Scholar] [CrossRef]
- Boavida, I.; Santos, J.M.; Ferreira, T.; Pinheiro, A. Barbel habitat alterations due to hydropeaking. J. Hydro Environ. Res. 2015, 9, 237–247. [Google Scholar] [CrossRef]
- Enders, E.C.; Boisclair, D.; Roy, A.G. A model of total swimming costs in turbulent flow for juvenile Atlantic salmon (Salmo salar). Can. J. Fish. Aquat. Sci. 2005, 62, 1079–1089. [Google Scholar] [CrossRef]
- Standen, E.M.; Hinch, S.G.; Healey, M.C.; Farrell, A.P. Energetic costs of migration through the Fraser River Canyon, British Columbia, in adult pink (Oncorhynchus gorbuscha) and sockeye (Oncorhynchus nerka) salmon as assessed by EMG telemetry. Can. J. Fish. Aquat. Sci. 2002, 59, 1809–1818. [Google Scholar] [CrossRef]
- Wang, H.; Chanson, H. Modelling upstream fish passage in standard box culverts: Interplay between turbulence, fish kinematics, and energetics. River Res. Appl. 2018, 34, 244–252. [Google Scholar] [CrossRef]
- Person, E.; Bieri, M.; Peter, A.; Schleiss, A.J. Mitigation measures for fish habitat improvement in Alpine rivers affected by hydropower operations. Ecohydrology 2014, 7, 580–599. [Google Scholar] [CrossRef]
- Gasith, A.; Resh, V.H. Streams in Mediterranean climate region: Abiotic influences and biotic responses to predictable seasonal events. Annu. Rev. Ecol. Syst. 1999, 30, 51–81. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Event | Number of Replicates | Discharge (L/s) | Time of Exposure 1 (min) | Water Depth (cm) | Maximum Registered Velocity 2 (m/s) |
|---|---|---|---|---|---|
| BF | 9 | 7 | 40 | 8.5 | 0.18 |
| HP | 9 | 60 | 40 | 25 | 0.71 |
| Factor | Pairwise Comparison | Results | |
|---|---|---|---|
| t | p | ||
| Structure | LWHP vs. LAHP | 1.70 | 0.007 |
| LWBF vs. LABF | 2.00 | 0.003 | |
| Tested Metrics | Kruskal–Wallis Multiple Comparison Tests | Pairwise Significant Differences | |
|---|---|---|---|
| χ2 | p | ||
| DL_I | 19.145 | 0.0002 | LWBF vs. LWHP LWHP vs. LABF LWHP vs. LAHP |
| DL_G | 9.901 | 0.01 | LWHP vs. LABF LWBF vs. LABF |
| IL_I | 21.217 | 0.00009 | LWBF vs. LABF LWHP vs. LABF LWHP vs. LAHP |
| IL_G | 23.114 | 0.00004 | LWBF vs. LABF LWBF vs. LAHP LWHP vs. LABF LWHP vs. LAHP |
| UL_I | 11.891 | 0.007 | LWHP vs. LABF |
| UL_G | 4.41 | 0.22 | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moreira, M.; Costa, M.J.; Valbuena-Castro, J.; Pinheiro, A.N.; Boavida, I. Cover or Velocity: What Triggers Iberian Barbel (Luciobarbus Bocagei) Refuge Selection under Experimental Hydropeaking Conditions? Water 2020, 12, 317. https://doi.org/10.3390/w12020317
Moreira M, Costa MJ, Valbuena-Castro J, Pinheiro AN, Boavida I. Cover or Velocity: What Triggers Iberian Barbel (Luciobarbus Bocagei) Refuge Selection under Experimental Hydropeaking Conditions? Water. 2020; 12(2):317. https://doi.org/10.3390/w12020317
Chicago/Turabian StyleMoreira, Miguel, Maria João Costa, Jorge Valbuena-Castro, António N. Pinheiro, and Isabel Boavida. 2020. "Cover or Velocity: What Triggers Iberian Barbel (Luciobarbus Bocagei) Refuge Selection under Experimental Hydropeaking Conditions?" Water 12, no. 2: 317. https://doi.org/10.3390/w12020317
APA StyleMoreira, M., Costa, M. J., Valbuena-Castro, J., Pinheiro, A. N., & Boavida, I. (2020). Cover or Velocity: What Triggers Iberian Barbel (Luciobarbus Bocagei) Refuge Selection under Experimental Hydropeaking Conditions? Water, 12(2), 317. https://doi.org/10.3390/w12020317