Zooplankton Size as a Factor Determining the Food Selectivity of Roach (Rutilus Rutilus) in Water Basin Outlets


Abstract
1. Introduction
2. Methods
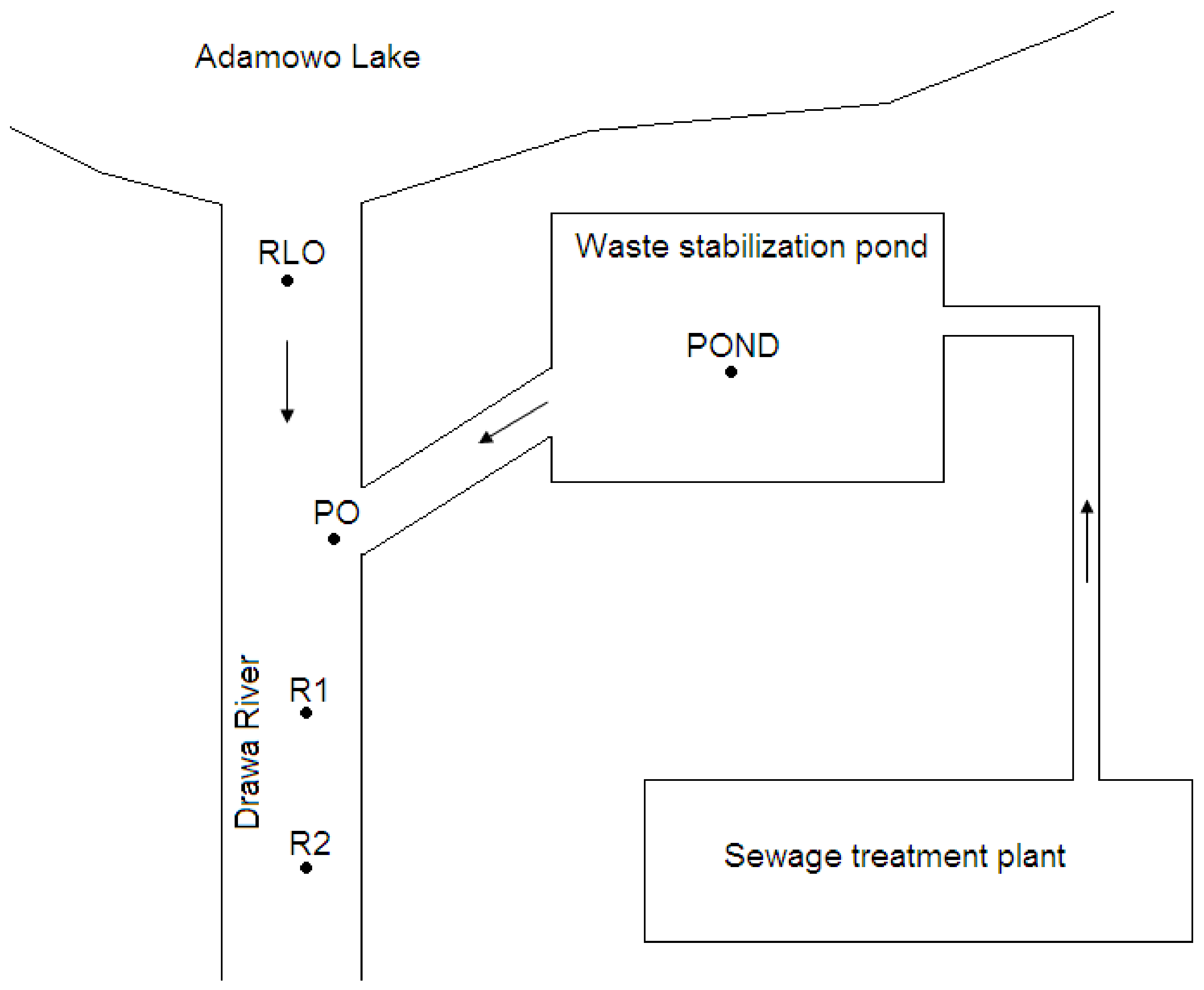
2.1. Study Area
2.2. Sampling Methods
2.3. Statistical Analyses
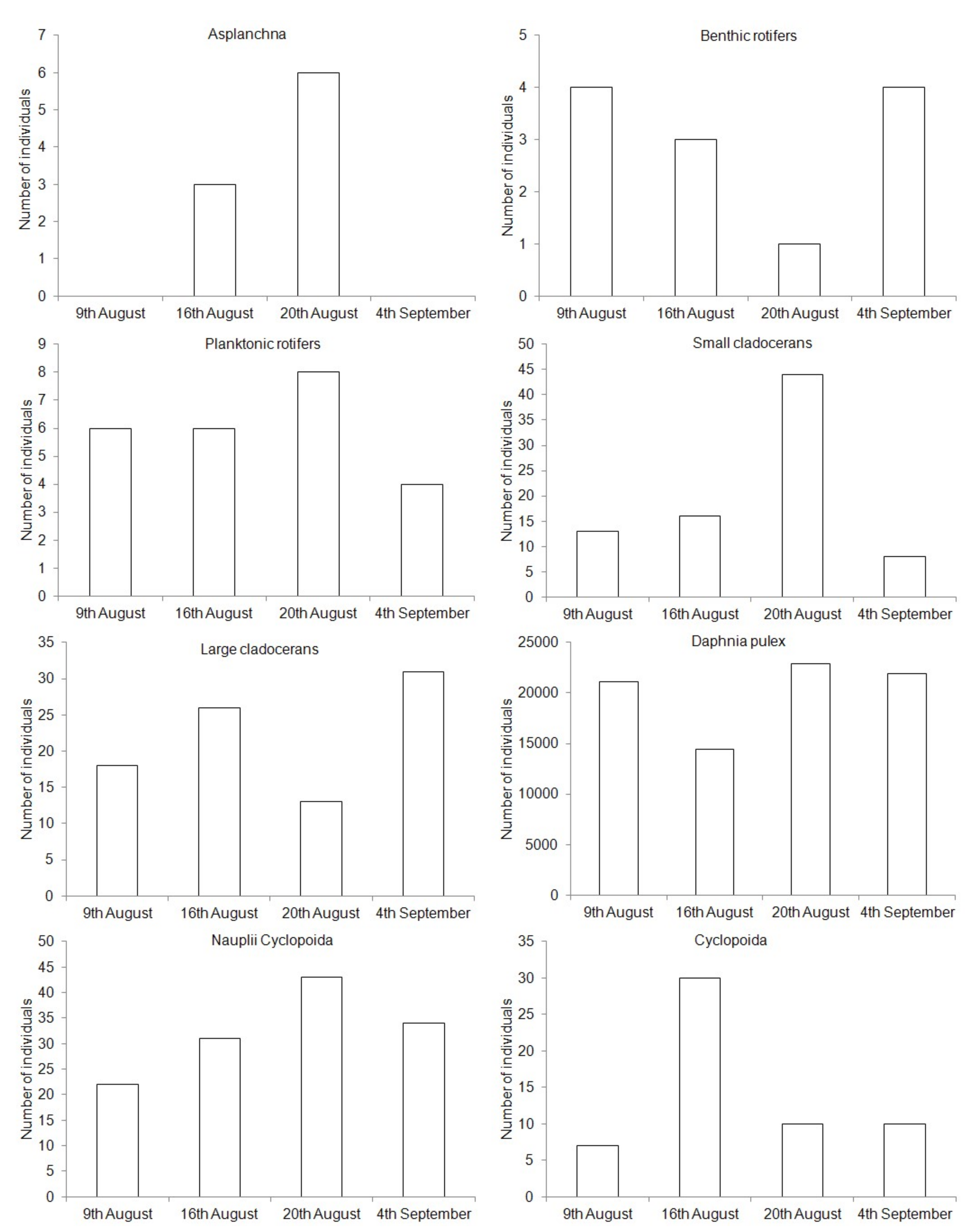
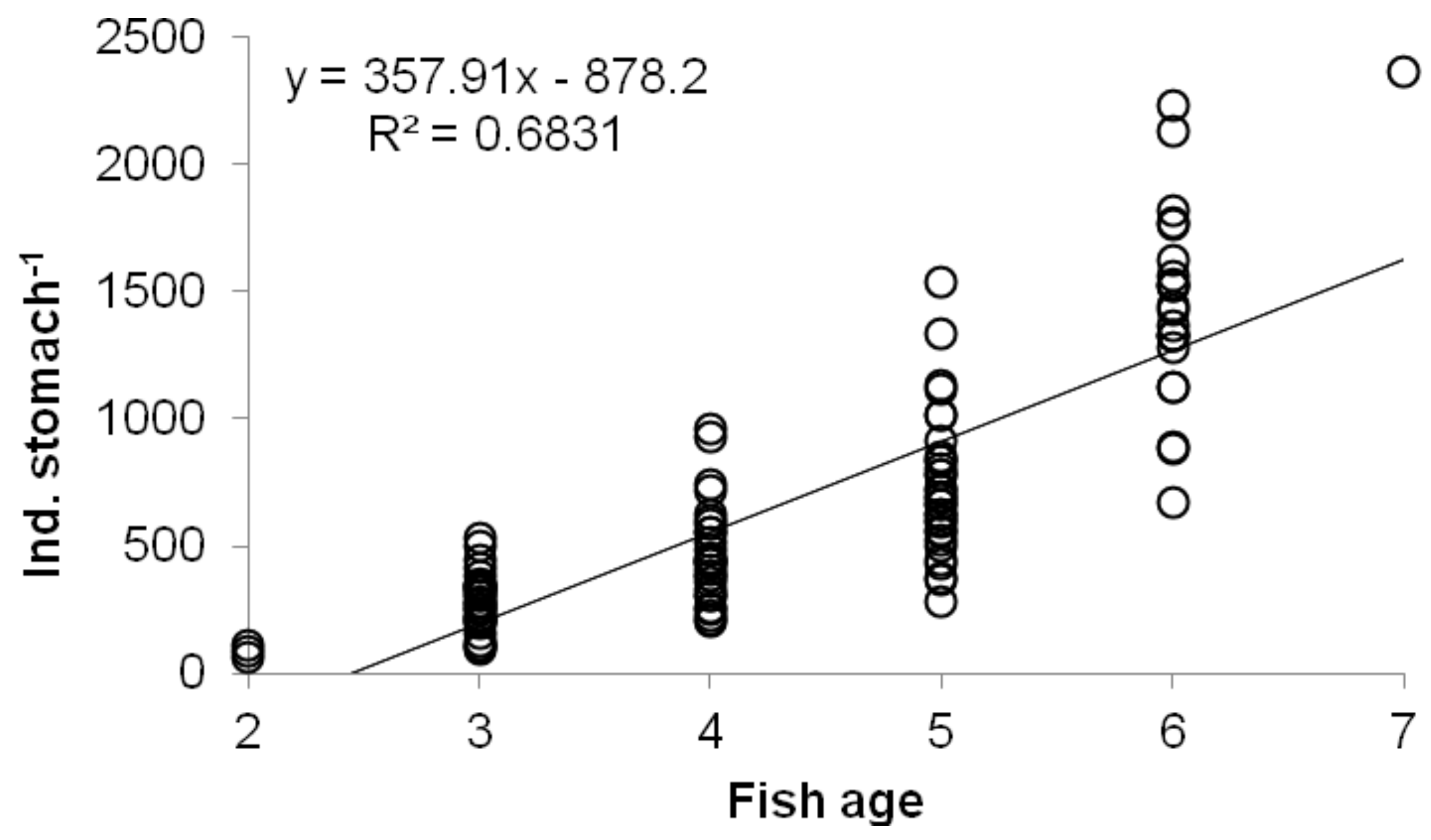
3. Results
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Gliwicz, Z.M. A lunar cycle in zooplankton. Ecology 1986, 67, 883–897. [Google Scholar] [CrossRef]
- Christoffersen, K.; Riemann, B.; Klysner, A.; Søndergaard, M. Potential role of fish predation and natural populations of zooplankton in structuring a plankton community in eutrophic lake water. Limnol. Oceanogr. 1993, 38, 561–573. [Google Scholar] [CrossRef]
- Wissel, B.; Boeing, W.J.; Ramcharan, C.W. Effects of water color on predation regimes and zooplankton assemblages in freshwater lakes. Limnol. Oceanogr. 1998, 48, 1965–1976. [Google Scholar] [CrossRef]
- Hoffman, J.C.; Smith, M.E.; Lehman, J.T. Perch or plankton: Top-down control of Daphnia by yellow perch (Perca flavescens) or Bythotrephes cederstroemi in an inland lake? Freshw. Biol. 2001, 46, 759–775. [Google Scholar] [CrossRef]
- Jack, J.D.; Thorp, J.H. Impacts of fish predation on an Ohio River zooplankton community. J. Plankton Res. 2002, 24, 119–127. [Google Scholar] [CrossRef]
- Czerniawski, R.; Domagała, J. Reduction of zooplankton communities in small lake outlets in relation to abiotic and biotic factors. Oceanol. Hydrobiol. Stud. 2013, 42, 123–131. [Google Scholar] [CrossRef]
- Czerniawski, R.; Sługocki, Ł.; Kowalska-Góralska, M. Diurnal changes of zooplankton community reduction rate at lake outlets and related environmental factors. PLoS ONE 2016, 11, e0158837. [Google Scholar] [CrossRef]
- Walks, D.J.; Cyr, H. Movement of plankton through lake-stream systems. Freshw. Biol. 2004, 49, 745–759. [Google Scholar] [CrossRef]
- Chang, K.H.; Doi, H.; Imai, H.; Gunji, F.; Nakano, S.I. Longitudinal changes in zooplankton distribution below a reservoir outfall with reference to river planktivory. Limnology 2008, 9, 125–133. [Google Scholar] [CrossRef]
- Mehner, T. Influence of spring warming on the predation rate of underyearling fish on daphnia—A deterministic simulation approach. Freshw. Biol. 2000, 45, 253–263. [Google Scholar] [CrossRef]
- Kuczyńska-Kippen, N.M.; Nagengast, B. The influence of the spatial structure of hydromacrophytes and differentiating habitat on the structure of rotifer and cladoceran communities. Hydrobiologia 2006, 559, 203–212. [Google Scholar] [CrossRef]
- Estlander, S.; Nurminen, L.; Olin, M.; Vinni, M.; Horppila, J. Seasonal fluctuations in macrophyte cover and water transparency of four brown-water lakes: Implications for crustacean zooplankton in littoral and pelagic habitats. Hydrobiologia 2009, 620, 109–120. [Google Scholar] [CrossRef]
- Kamarainen, A.M.; Rowland, F.E.; Biggs, R.; Carpenter, S.R. Zooplankton and the total phosphorus-chlorophyll a relationship: Hierarchical bayesian analysis of measurement error. Can. J. Fish. Aquat. Sci. 2008, 65, 2644–2655. [Google Scholar] [CrossRef]
- Gołdyn, R.; Kowalczewska-Madura, K. Interactions between phytoplankton and zooplankton in the hypertrophic swarzędzkie Lake in western Poland. J. Plankton Res. 2008, 30, 33–42. [Google Scholar]
- Pourriot, R.; Rougier, C.; Miquelis, A. Origin and development of river zooplankton: Example of the Marne. Hydrobiologia 1997, 345, 143–148. [Google Scholar] [CrossRef]
- Ejsmont-Karabin, J.; Kruk, M. Effects of contrasting land use on free-swimming rotifer communities of streams in Masurian Lake District, Poland. Hydrobiologia 1998, 387/388, 241–249. [Google Scholar] [CrossRef]
- Radwan, S. Wrotki (Rotifera); Wydawnictwo Uniwersytetu Łódzkiego: Łódź, Polska, 2004; pp. 1–447. [Google Scholar]
- Rybak, J.I.; Błędzki, L.A. Planktonic Crustaceans of Freshwaters; Wydawnictwo Uniwersytetu Warszawskiego: Warszawa, Poland, 2010; pp. 1–366. [Google Scholar]
- Czerniawski, R.; Domagała, J. Small dams profoundly alter the spatial and temporal composition of zooplankton communities in running waters. Int. Rev. Hydrobiol. 2014, 99, 300–311. [Google Scholar] [CrossRef]
- Venables, W.N.; Ripley, B.D. Modem Applied Statistics with S. Fourth Edition; Springer: New York, USA, 2002; pp. 1–487. [Google Scholar]
- R Core Team. A Language and Environment for Statistical Computing—R Foundation for Statistical Computing; Vienna, Austria. 2018. Available online: https://www.R-project.org (accessed on 20 December 2018).
- Czerniawski, R.; Domagała, J. Zooplankton communities of two lake outlets in relation to abiotic factors. Open Life Sci. 2010, 5, 240–255. [Google Scholar] [CrossRef]
- Czerniawski, R.; Domagała, J. Similarities in zooplankton community between river drawa and its two tributaries (Polish part of River Odra). Hydrobiologia 2010, 638, 137–149. [Google Scholar] [CrossRef]
- Bone, Q.; Moore, R. Biology of Fishes; Taylor and Francis: New York, NY, USA, 2008; pp. 1–497. [Google Scholar]
- Akopian, M.; Garnier, J.; Pourriot, R. A large reservoir as a source of zooplankton for the river: Structure of the populations and influence of fish predation. J. Plankton Res. 1999, 21, 285–297. [Google Scholar] [CrossRef]
- Meeren, T.; Olsen, E.R.; Hamre, K.; Fyhn, J.H. Biochemical composition of copepods for evaluation of feed production of juvenile marine fish. Aquaculture 2008, 274, 375–379. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Date | Fish Age | Length (cm) | Mass (g) | Condition Factor |
|---|---|---|---|---|
| 9th August | 6+ (n = 2) | 13.70 ± 0.57 | 35.05 ± 5.16 | 1.36 ± 0.03 |
| 5+ (n = 10) | 10.88 ±1.05 | 17.25 ± 4.92 | 1.31 ± 0.08 | |
| 4+ (n = 7) | 10.31 ± 0.39 | 13.80 ± 1.91 | 1.25 ± 0.11 | |
| 3+ (n = 11) | 8.69 ± 0.34 | 8.09 ± 1.58 | 1.22 ± 0.13 | |
| 16th August | 6+ (n = 4) | 12.58 ± 0.62 | 23.90 ± 3.63 | 1.20 ± 0.10 |
| 5+ (n = 6) | 10.57 ± 1.05 | 14.78 ± 3.55 | 1.24 ± 0.10 | |
| 4+ (n = 13) | 9.14 ± 0.82 | 9.30 ± 2.99 | 1.17 ± 0.11 | |
| 3+ (n = 5) | 7.42 ± 0.22 | 4.72 ± 0.69 | 1.15 ± 0.08 | |
| 2+ (n = 2) | 6.20 ± 0.14 | 2.55 ± 0.07 | 1.07 ± 0.10 | |
| 20th August | 7+ (n = 1) | 14.30 | 39.40 | 1.35 |
| 6+ (n = 7) | 12.37 ± 0.76 | 24.63 ± 3.93 | 1.29 ± 0.09 | |
| 5+ (n = 7) | 11.03 ± 0.73 | 17.30 ± 3.96 | 1.27 ± 0.07 | |
| 4+ (n = 5) | 9.40 ± 0.72 | 10.74 ± 2.44 | 1.28 ± 0.05 | |
| 3+ (n = 9) | 7.42 ± 0.36 | 5.19 ± 0.77 | 1.26 ± 0.07 | |
| 2+ (n = 1) | 5.60 | 2.20 | 1.25 | |
| 4th September | 6+ (n = 9) | 12.16 ± 0.30 | 23.14 ± 1.90 | 1.29 ± 0.04 |
| 5+ (n = 9) | 11.38 ± 0.31 | 18.88 ± 1.71 | 1.28 ± 0.05 | |
| 4+ (n = 3) | 8.47 ± 0.46 | 7.50 ± 1.68 | 1.22 ± 0.09 | |
| 3+ (n = 9) | 7.49 ± 0.33 | 5.31 ± 0.94 | 1.25 ± 0.10 |
| Zooplankton Group | RLO | POND | PO | R1 | R2 |
|---|---|---|---|---|---|
| Asplanchna | 17.4 ± 18.2 a | 4.7 ± 8.9 ab | 2.5 ± 5.0 b | 12.4 ± 11.2 ab | 9.6 ± 9.6 ab |
| Benthic rotifers | 18.9 ± 11.6 a | 4.3 ± 6.8 a | 2.4 ± 1.1 a | 23.3 ± 22.0 a | 24.5 ± 17.9 a |
| Planktonic rotifers | 351.5 ± 216.7 a | 37.7 ± 30.3 ab | 24.9 ± 21.2 b | 313.6 ± 178.8 ab | 325.4 ± 202.5 ab |
| Small cladocerans | 38.6 ± 16.8 ab | 54.2 ± 51.8 ab | 39.7 ± 45.4 ab | 46.8 ± 32.8 ab | 28.3 ± 15.8 ab |
| Large cladocerans | 6.0 ± 3.9 ab | 0.6 ± 0.7 ab | - | 2.9 ± 2.1 ab | 1.0 ± 0.5 ab |
| Daphnia pulex | - | 122.9 ± 62.5 a | 84.4 ± 57.5 a | 9.1 ± 9.6 b | 0.8 ± 1.0 b |
| Nauplii Cyclopoida | 57.5 ± 59.3 a | 7.7 ± 6.5 ab | 4.5 ± 3.1 b | 61.4 ± 73.8 ab | 50.5 ± 65.3 ab |
| Cyclopoida | 21.2 ± 10.2 a | 6.9 ± 2.9 b | 3.9 ± 2.4 b | 20.6 ± 9.3 a | 8.5 ± 4.6 ab |
| Fixed Effects | Estimate | Std. Error | z Value | p |
|---|---|---|---|---|
| (Intercept) | 0.803 | 0.426 | 1.883 | 0.060 |
| R1 vs PO | 1.492 | 0.492 | 3.035 | 0.002 |
| R2 vs PO | 1.248 | 0.500 | 2.496 | 0.013 |
| Benthic rotifers | −0.311 | 0.602 | −0.516 | 0.606 |
| Cyclopoida | 0.336 | 0.538 | 0.625 | 0.532 |
| Daphnia pulex | 1.825 | 0.581 | 3.142 | 0.002 |
| Large cladocerans | −21.717 | 42.375 | −0.512 | 0.608 |
| Nauplii Cyclopoida | 0.251 | 0.538 | 0.466 | 0.641 |
| Planktonic rotifers | 1.167 | 0.594 | 1.965 | 0.049 |
| Small cladocerans | 1.678 | 0.502 | 3.344 | 0.001 |
| RLO | 0.001 | 0.001 | 1.209 | 0.227 |
| POND | 0.013 | 0.003 | 4.384 | <0.001 |
| R1 vs PO: Benthic rotifers | 0.951 | 0.714 | 1.333 | 0.182 |
| R2 vs PO: Benthic rotifers | 1.273 | 0.719 | 1.769 | 0.077 |
| R1 vs PO: Cyclopoida | 0.240 | 0.663 | 0.362 | 0.717 |
| R2 vs PO: Cyclopoida | −0.400 | 0.682 | −0.586 | 0.558 |
| R1 vs PO: Daphnia pulex | −3.903 | 0.635 | −6.149 | <0.001 |
| R2 vs PO: Daphnia pulex | −6.017 | 0.850 | −7.075 | <0.001 |
| R1 vs PO: Large cladocerans | 20.295 | 42.376 | 0.479 | 0.632 |
| R2 vs PO: Large cladocerans | 18.908 | 42.377 | 0.446 | 0.655 |
| R1 vs PO: Nauplii Cyclopoida | 1.137 | 0.656 | 1.733 | 0.083 |
| R2 vs PO: Nauplii Cyclopoida | 1.148 | 0.663 | 1.730 | 0.084 |
| R1 vs PO: Planktonic rotifers | 1.282 | 0.614 | 2.087 | 0.037 |
| R2 vs PO: Planktonic rotifers | 1.527 | 0.621 | 2.458 | 0.014 |
| R1 vs PO: Small cladocerans | −0.993 | 0.614 | −1.619 | 0.106 |
| R2 vs PO: Small cladocerans | −1.173 | 0.624 | −1.879 | 0.060 |
| Fish age | 2+ | 3+ | 4+ | 5+ | 6+ | 7+ | |
| n | 3 | 34 | 28 | 32 | 22 | 1 | |
| Zooplankton group | Asplanchna | - | 0.03 ± 0.10 a | 0.02 ± 0.09 a | 0.01 ± 0.04 a | 0.01 ± 0.03 a | - |
| Benthic rotifers | 0.88 ± 1.52 a | 0.10 ± 0.31 a | 0.02 ± 0.06 a | - | - | - | |
| Pelagic rotifers | 1.02 ± 0.91 a | 0.23 ± 0.47 a | 0.03 ± 0.10 a | 0.02 ± 0.08 a | - | - | |
| Small cladocerans | 2.50 ± 0.22 a | 0.40 ± 0.83 ab | 0.16 ± 0.35 ab | 0.05 ± 0.12 b | 0.04 ± 0.07 b | 0.25 ab | |
| Large cladocerans | 0.88 ± 1.52 a | 0.26 ± 0.47 a | 0.15 ± 0.23 a | 0.10 ± 0.16 a | 0.09 ± 0.12 a | 0.08 a | |
| Daphnia pulex | 90.96 ± 7.01 a | 98.11 ± 2.13 a | 99.16 ± 1.13 ab | 99.65 ± 0.32 ab | 99.74 ± 0.27 b | 99.58 ab | |
| Nauplii | 3.32 ± 3.29 a | 0.39 ± 0.62 ab | 0.18 ± 0.40 ab | 0.02 ± 0.07 b | 0.01 ± 0.03 b | - | |
| Cyclopoida | 0.44 ± 0.76 a | 0.47 ± 0.80 a | 0.27 ± 0.52 a | 0.15 ± 0.20 a | 0.12 ± 0.20 a | - | |
| Site RLO | n | r | p |
|---|---|---|---|
| Ab. Asplanchna vs. Stom. Asplanchna | 4 | 0.74 | 0.262 |
| Ab. Benthic rotifers vs. Stom. Benthic rotifers | 4 | −0.74 | 0.262 |
| Ab. Planktonic rotifers vs. Stom. Pelagic rotifers | 4 | 0.32 | 0.683 |
| Ab. Small cladocerans vs. Stom. Small cladocerans | 4 | 0.80 | 0.200 |
| Ab. Large cladocerans vs. Stom. Large cladocerans | 4 | 0.99 | 0.001 |
| Ab. Daphnia pulex vs. Stom. Daphnia pulex | 1 | - | - |
| Ab. Nauplii Cyclopoida vs. Stom. Nauplii | 4 | 0.63 | 0.367 |
| Ab. Cyclopoida vs. Stom. Cyclopoida | 4 | 0.80 | 0.200 |
| Site PO | |||
| Ab. Asplanchna vs. Stom. Asplanchna | 1 | - | - |
| Ab. Benthic rotifers vs. Stom. Benthic rotifers | 4 | 0.94 | 0.051 |
| Ab. Planktonic rotifers vs. Stom. Pelagic rotifers | 4 | 0.32 | 0.684 |
| Ab. Small cladocerans vs. Stom. Small cladocerans | 4 | 0.80 | 0.200 |
| Ab. Large cladocerans vs. Stom. Large cladocerans | 0 | - | - |
| Ab. Daphnia pulex vs. Stom. Daphnia pulex | 4 | 0.80 | 0.200 |
| Ab. Nauplii Cyclopoida vs. Stom. Nauplii | 4 | 0.83 | 0.167 |
| Ab. Cyclopoida vs. Stom. Cyclopoida | 4 | 0.60 | 0.400 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Czerniawski, R.; Krepski, T. Zooplankton Size as a Factor Determining the Food Selectivity of Roach (Rutilus Rutilus) in Water Basin Outlets. Water 2019, 11, 1281. https://doi.org/10.3390/w11061281
Czerniawski R, Krepski T. Zooplankton Size as a Factor Determining the Food Selectivity of Roach (Rutilus Rutilus) in Water Basin Outlets. Water. 2019; 11(6):1281. https://doi.org/10.3390/w11061281
Chicago/Turabian StyleCzerniawski, Robert, and Tomasz Krepski. 2019. "Zooplankton Size as a Factor Determining the Food Selectivity of Roach (Rutilus Rutilus) in Water Basin Outlets" Water 11, no. 6: 1281. https://doi.org/10.3390/w11061281
APA StyleCzerniawski, R., & Krepski, T. (2019). Zooplankton Size as a Factor Determining the Food Selectivity of Roach (Rutilus Rutilus) in Water Basin Outlets. Water, 11(6), 1281. https://doi.org/10.3390/w11061281