The Copepod Acartia tonsa Dana in a Microtidal Mediterranean Lagoon: History of a Successful Invasion


Abstract
1. Introduction
2. Materials and Methods
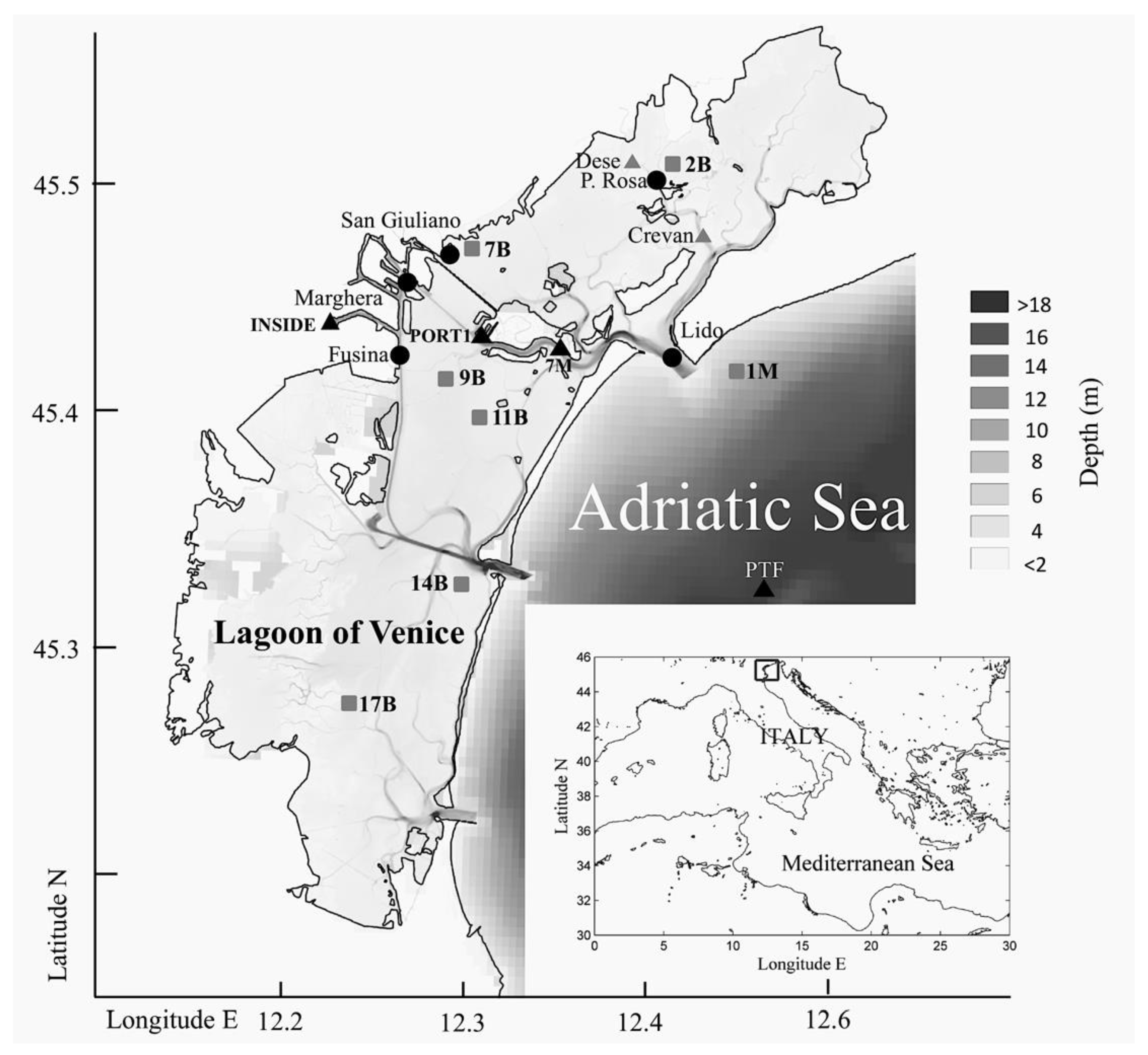
2.1. Study Area
2.2. Zooplankton and Environmental Variables Datasets
2.3. Statistical Analyses
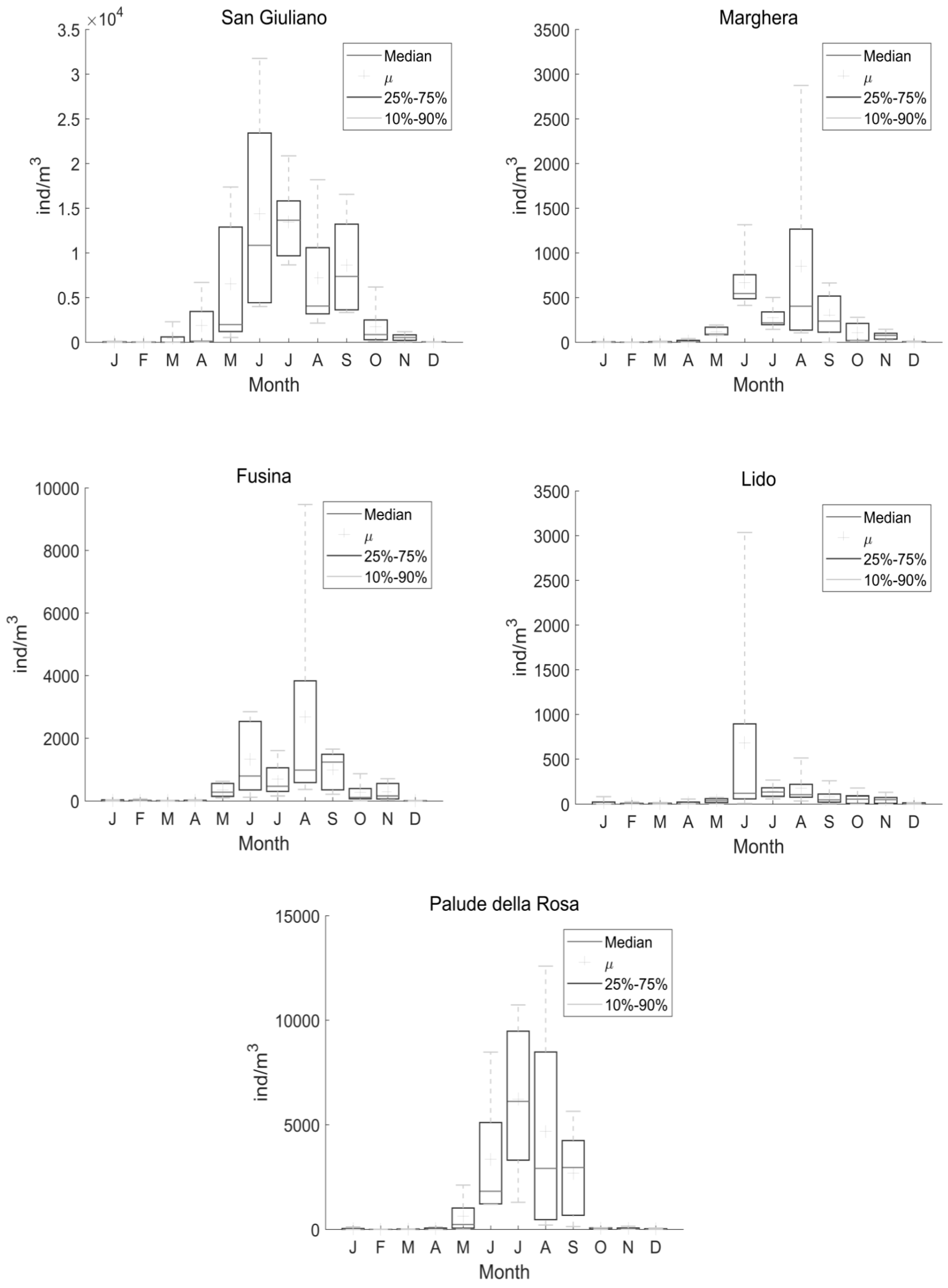
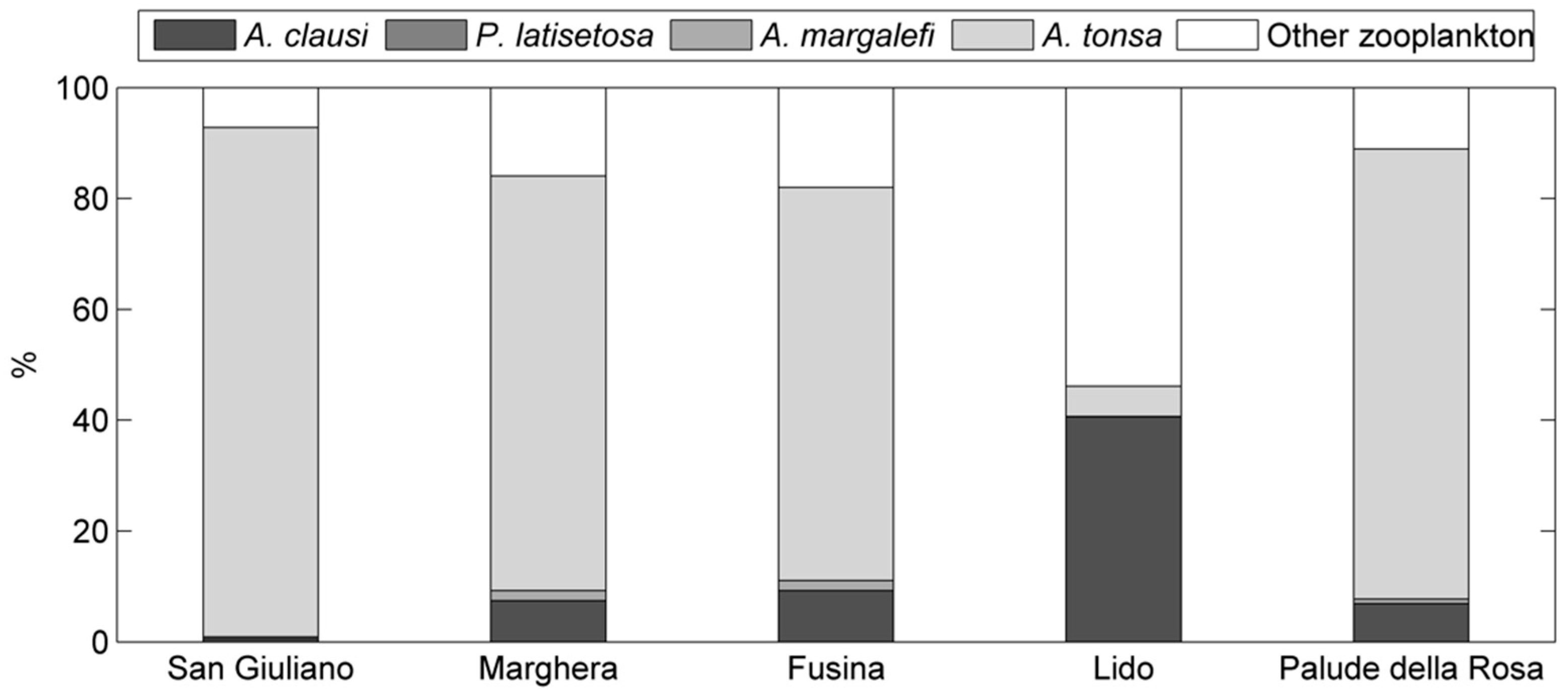
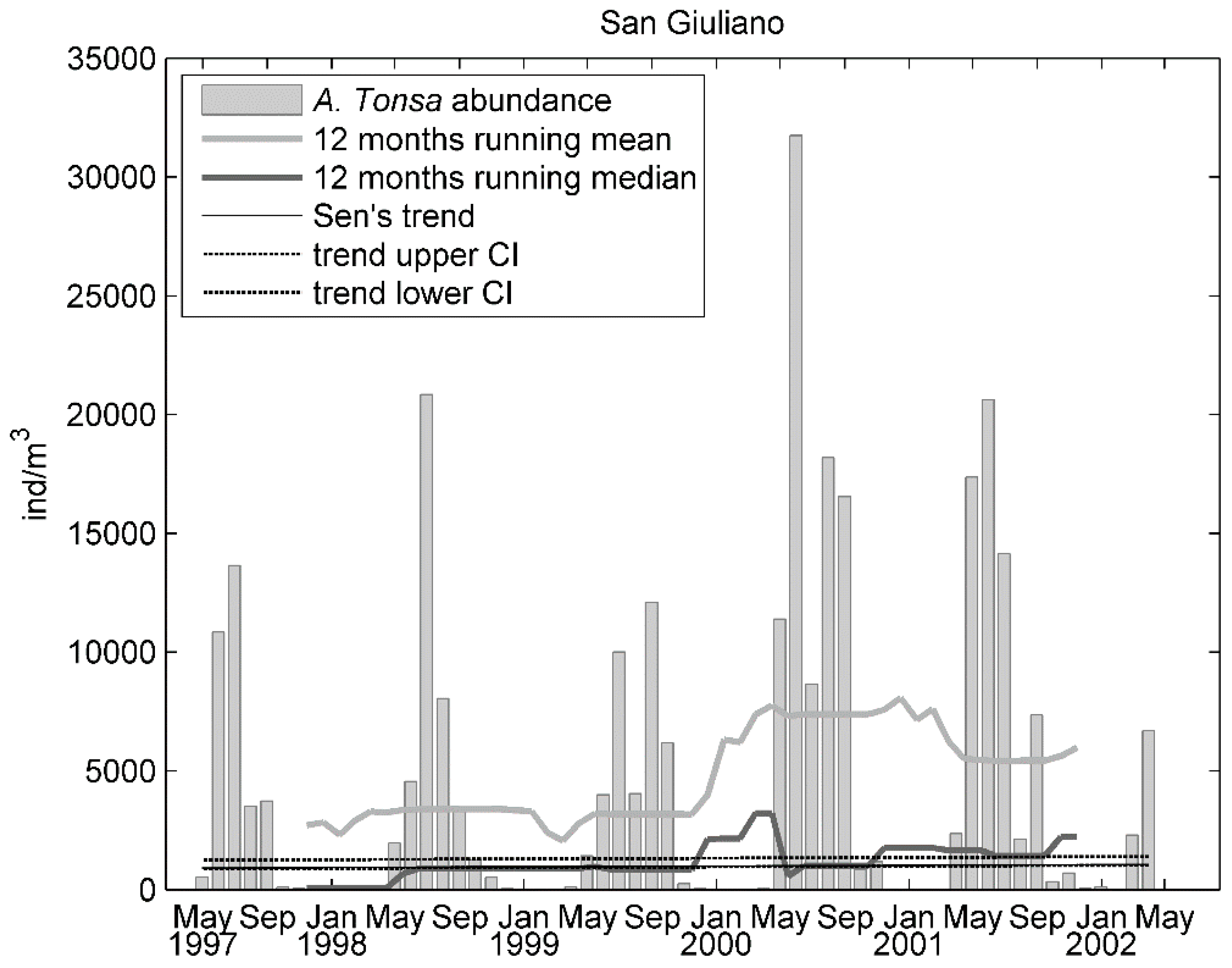
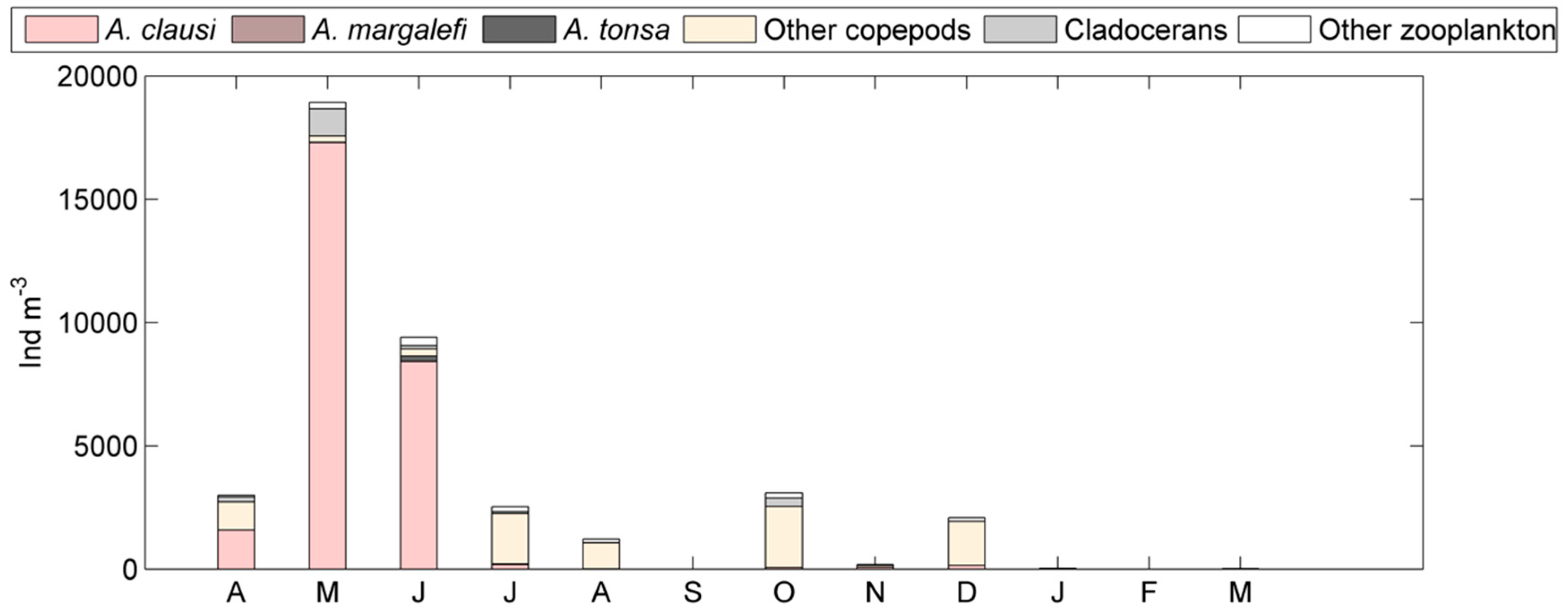
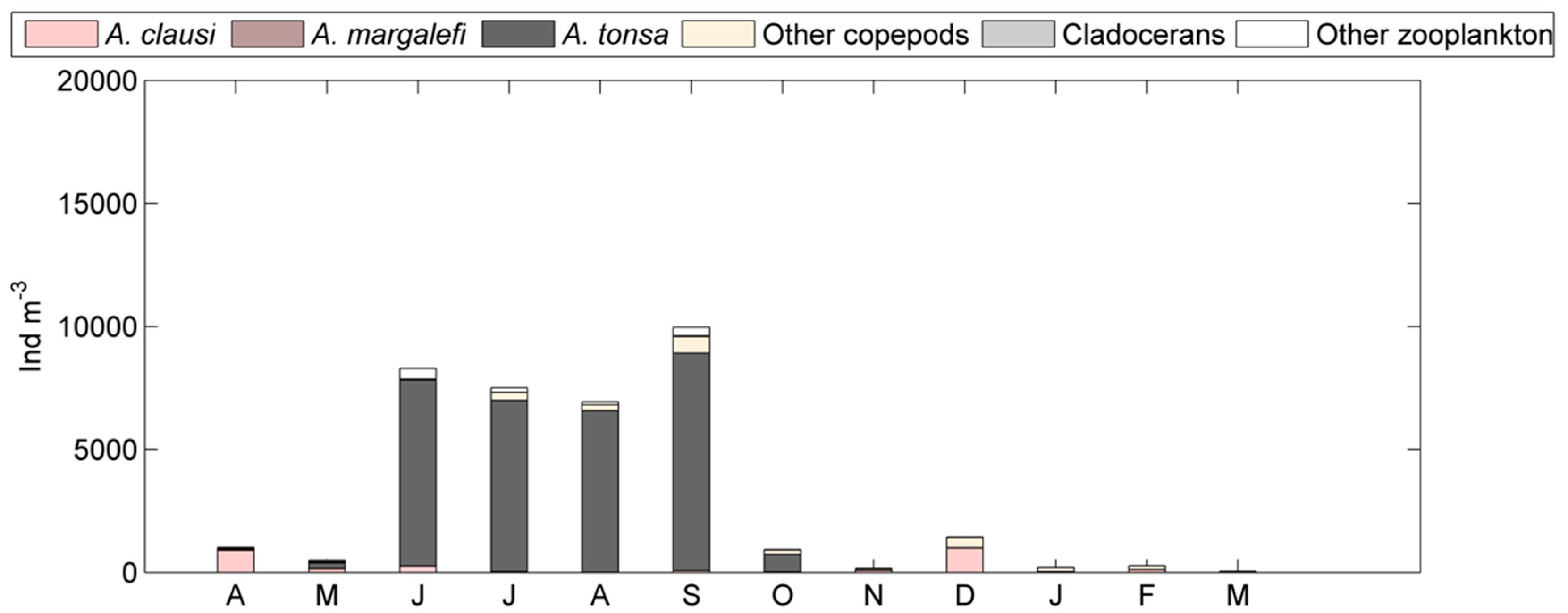
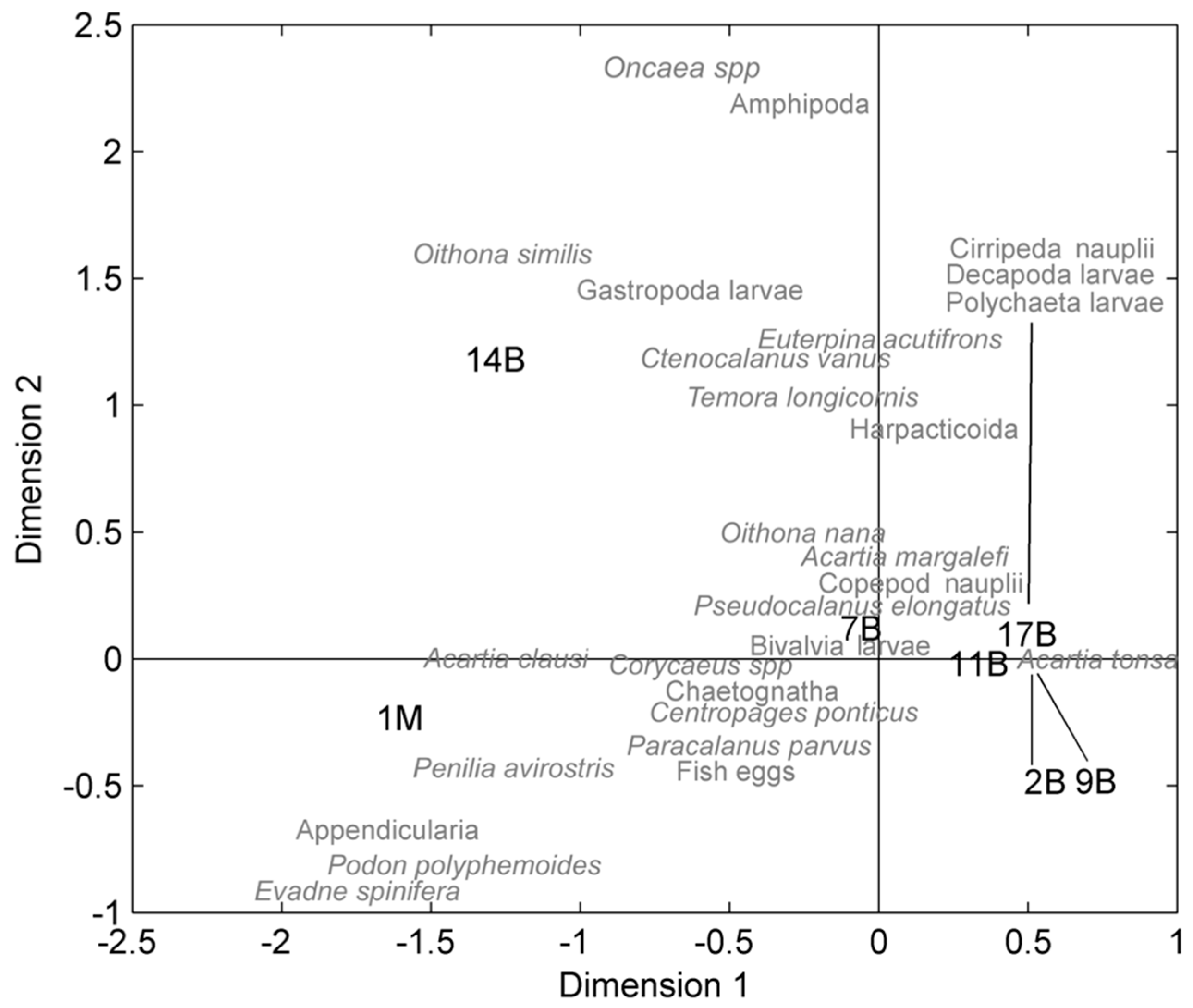
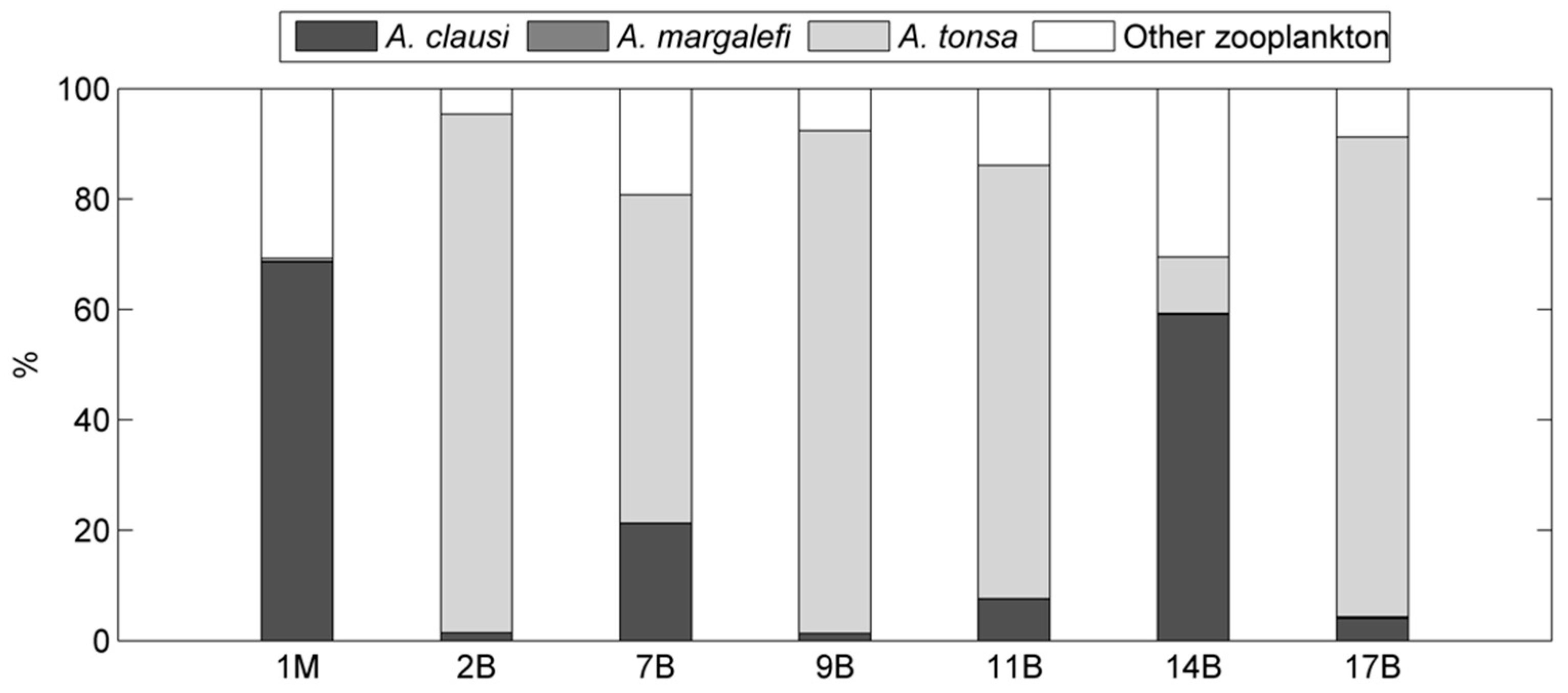
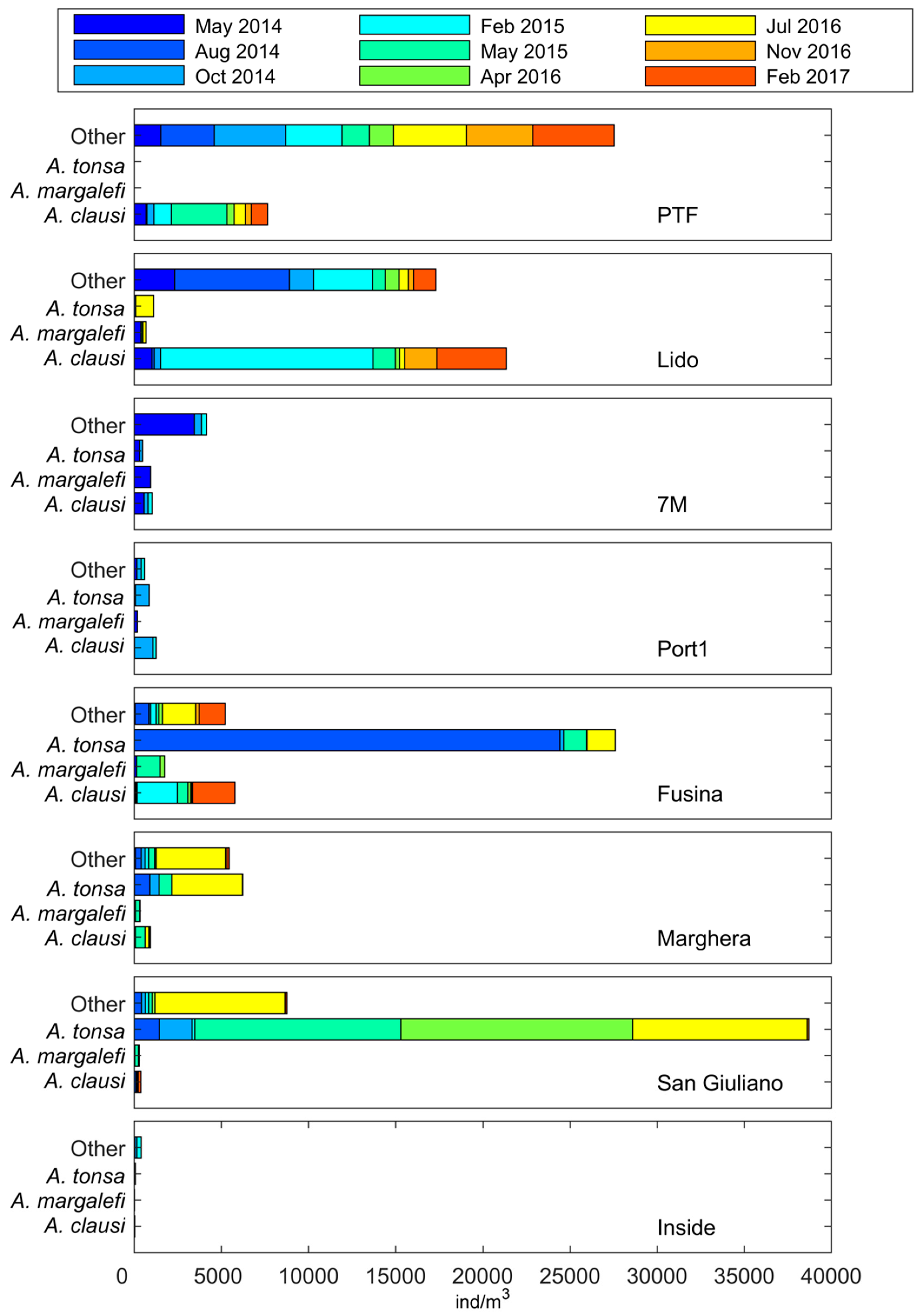
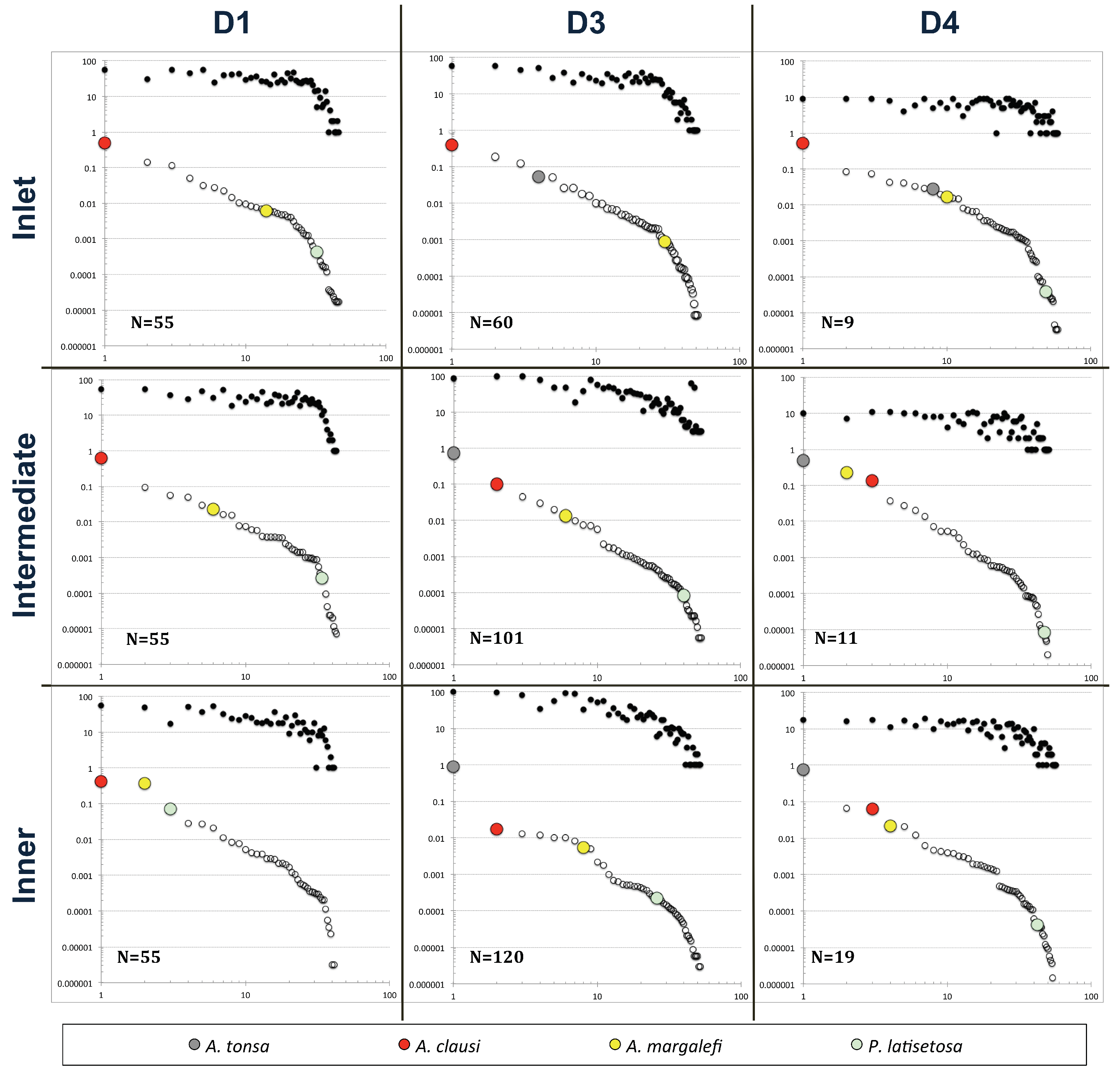
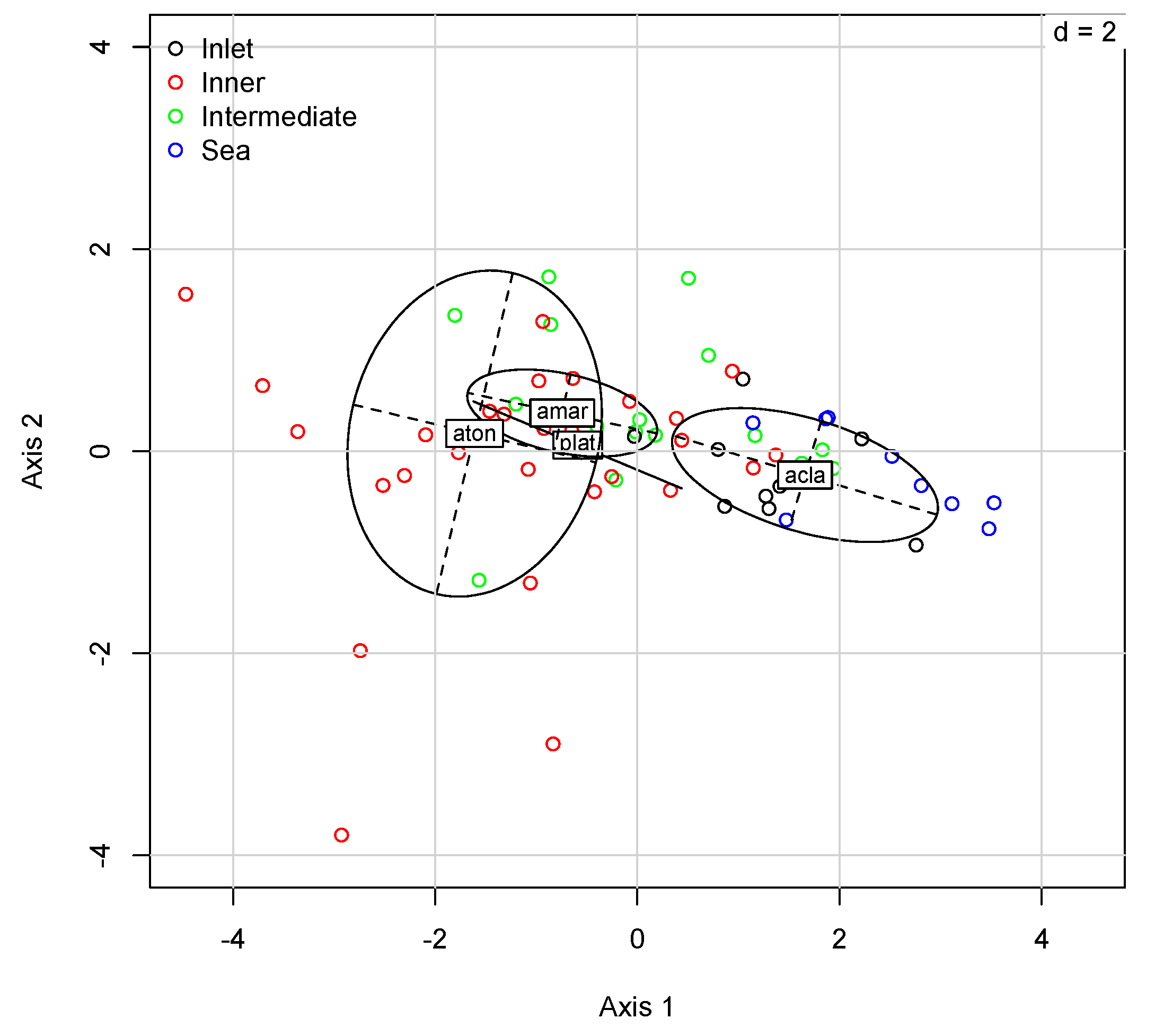
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- IMO (International Maritime Organization). Global Ballast Water Management Project: The Problem. Available online: http://globallast.imo.org/index. asp?page=problem.htm&menu=true (accessed on 7 May 2019).
- Gaudy, R.; Vinas, M. Premiere signalisation en Méditerrannée du copepode pélagique Acartia tonsa. Rapp. Comm. Int. Mer Mediterr. 1985, 219, 227–229. [Google Scholar]
- Seuront, L. First record of the calanoid copepod Acartia omorii (Copepoda: Calanoida: Acartiidae) in the southern bight of the North Sea. J. Plankton Res. 2005, 27, 1301–1306. [Google Scholar] [CrossRef]
- David, V.; Sautour, B.; Chardy, P. Successful colonization of the calanoid copepod Acartia tonsa in the oligo mesohaline area of the Gironde estuary (SW France) Natural or anthropogenic forcing? Estuar. Coast. Shelf Sci. 2007, 71, 429–442. [Google Scholar] [CrossRef]
- Belmonte, G.; Potenza, D. Biogeography of the family Acartiidae (Calanoida) in the Ponto Mediterranean Province. Hydrobiologia 2001, 453, 171–176. [Google Scholar] [CrossRef]
- Lakkis, S. Coexistence and competition within Acartia (Copepoda, Calanoida) congeners from Lebanese coastal water: Niche overlap measurements. In Ecology and Morphology of Copepods, Hydrobiologia; Ferrari, F.D., Bradley, B.P., Eds.; Kluwer Academic Publishers: Brussels, Belgium, 1994; Chapter 292–293; pp. 481–490. [Google Scholar]
- Hoffmeyer, M.S. Decadal change in zooplankton seasonal succession in the Bahía Blanca Estuary, Argentina, following introduction of two zooplankton species. J. Plankton Res. 1994, 26, 1–9. [Google Scholar] [CrossRef]
- Durbin, E.G.; Durbin, A.G.; Smayda, T.J.; Verity, P.G. Food limitation of production by adult Acartia tonsa in Narragansett bay, Rhode Island. Limnol. Oceanogr. 1983, 28, 1199–1213. [Google Scholar] [CrossRef]
- Brylinski, J.M. Report on the presence of Acartia tonsa Dana (Copepoda) in the harbour of Dunkirk (France) and its geographical distribution in Europe. J. Plankon Res. 1981, 3, 255–260. [Google Scholar] [CrossRef]
- Gruszka, P. The River Odra Estuary as a Gateway for Alien Species Immigration to the Baltic Sea Basin. Acta Hydrochim. Hydrobiol. 1999, 27, 374–382. [Google Scholar] [CrossRef]
- Farabegoli, A.; Ferrari, I.; Manzoni, C.; Pugnetti, A. Prima segnalazione nel Mare Adriatico del copepode calanoide Acartia tonsa. Nov. Thalass. 1989, 10, 207–208. [Google Scholar]
- Quarta, S.; Belmonte, G.; Caroppo, C.; Pacifico, P.; Petraroli, P. Zooplankton seasonal trend in the Lesina and Varano Lagoons (Apulian coast of Italy). Oebalia 1992, 17, 403–404. [Google Scholar]
- Comaschi, A.; Cavalloni, B. Variabilità stagionale dello zooplancton nella Palude della Rosa (laguna di Venezia bacino settentrionale). SITE 1995, 16, 75–78. [Google Scholar]
- Durbin, A.G.; Durbin, E.G. Standing stock and estimated production rates of phytoplankton and zooplankton in Narragansett Bay, Rhode Island. Estuaries 1981, 4, 24–41. [Google Scholar] [CrossRef]
- Roman, M.R. Utilization of detritus by the copepod Acartia tonsa. Limnol. Oceanogr. 1984, 5, 949–959. [Google Scholar] [CrossRef]
- Paffenöfer, G.A.; Stearn, D.E. Why is Acartia tonsa (Copepoda: Calanoida) restricted to nearshore environment? Mar. Ecol. Prog. Ser. 1988, 42, 33–38. [Google Scholar] [CrossRef]
- Zhang, J.; Ianora, A.; Wu, C.; Pellegrini, D.; Esposito, F.; Buttino, I. How to increase productivity of the copepod Acartia tonsa (Dana): Effects of population density and food concentration. Aquac. Res. 2015, 46, 2982–2990. [Google Scholar] [CrossRef]
- Tagliapietra, D.; Sigovini, M.; Volpi Ghirardini, A. A review of terms and definitions to categorise estuaries, lagoons and associated environments. Mar. Freshw. Res. 2009, 60, 497–509. [Google Scholar] [CrossRef]
- Tagliapietra, D.; Zanon, V.; Frangipane, G.; Umgiesser, G.; Sigovini, M. Physiographic zoning of the Venetian Lagoon. In Scientific Research and Safeguarding of Venice, 2010; CORILA: Venice, Italy, 2007; pp. 161–164. [Google Scholar]
- Occhipinti Ambrogi, A.; Marchini, A.; Cantone, G.; Castelli, A.; Chimenz, C.; Cormaci, M.; Froglia, C.; Furnari, G.; Gambi, M.C.; Giaccone, G.; et al. Alien species along the Italian coasts: An overview. Biol. Invasion 2011, 13, 215–237. [Google Scholar] [CrossRef]
- Keppel, E.; Sigovini, M.; Tagliapietra, D. A new geographical record of Polycera hedgpethi Er. Marcus, 1964 (Nudibranchia: Polyceridae) and evidence of its established presence in the Mediterranean Sea, with a review of its geographical distribution. Mar. Biol. Res. 2012, 8, 969–981. [Google Scholar] [CrossRef]
- Sfriso, A.; Campolin, M.; Sfriso, A.; Buosi, A.; Facca, C. Change of aquatic flora and vegetation in ecological gradients from the lagoon inlets to some fishing ponds of the Venice lagoon. Biol. Mar. Mediterr. 2012, 19, 53–56. [Google Scholar]
- Sfriso, A.; Facca, C. Annual growth and environmental relationships of the invasive species Sargassum muticum and Undaria pinnatifida in the lagoon of Venice. Estuar. Coast. Shelf Sci. 2013, 129, 162–172. [Google Scholar] [CrossRef]
- Tagliapietra, D.; Keppel, E.; Sigovini, M. First record of the colonial ascidian Didemnum vexillum Kott, 2002 in the Mediterranean: Lagoon of Venice (Italy). BioInvasions Rec. 2012, 1, 247–254. [Google Scholar] [CrossRef]
- Marchini, A.; Galil, B.S.; Occhipinti Ambrogi, A. Recommendations on standardizing lists of marine alien species: Lessons from the Mediterranean Sea. Mar. Pollut. Bull. 2015, 101, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Solidoro, C.; Pastres, R.; Cossarini, G.; Ciavatta, S. Seasonal and spatial variability of water quality parameters in the lagoon of Venice. J. Mar. Syst. 2004, 51, 7–18. [Google Scholar] [CrossRef]
- Gačić, M.; Kovačević, V.; Mancero Mosquera, I.; Mazzoldi, A.; Cosoli, S. Water fluxes between the Venice Lagoon and the Adriatic Sea. In Flooding and Environmental Challenges for Venice and Its Lagoon: State of Knowledge; Fletcher, C.A., Spencer, D.T., Eds.; Cambridge University: Cambridge, UK, 2005; pp. 431–444. [Google Scholar]
- Cucco, A.; Umgiesser, G. Modelling the Venice Lagoon residence time. Ecol. Model. 2006, 193, 34–51. [Google Scholar] [CrossRef]
- Ravera, O. The Lagoon of Venice: The result of both natural factors and human influence. J. Limnol. 2000, 59, 19–30. [Google Scholar] [CrossRef]
- Solidoro, C.; Bandelj, V.; Aubry Bernardi, F.; Camatti, E.; Ciavatta, S.; Cossarini, G.; Facca, C.; Franzoi, P.; Libralato, S.; Melaku Canu, D.; et al. Response of Venice Lagoon Ecosystem to Natural and Anthropogenic Pressures over the Last 50 Years. In Coastal Lagoons Critical Habitats of Environmental Change; Kennish, M., Paerl, H.W., Eds.; CRC Press: Boca Raton, FL, USA, 2010; pp. 483–511. [Google Scholar]
- Bianchi, F.; Acri, F.; Bernardi Aubry, F.; Berton, A.; Boldrin, A.; Camatti, E.; Cassin, D.; Comaschi, A. Can plankton communities be considered as bio indicators of water quality in the Lagoon of Venice? Mar. Pollut. Bull. 2003, 46, 964–971. [Google Scholar] [CrossRef]
- Bianchi, F.; Socal, G.; Alberighi, L.; Cioce, F. Cicli nictemerali dell’ossigeno disciolto nel bacino centrale della laguna di Venezia. Biol. Mar. Mediterr. 1996, 3, 628–630. [Google Scholar]
- Socal, G.; Bianchi, F.; Alberighi, L. Effects of thermal pollution and nutrient discharges on a spring phytoplankton bloom in the industrial area of the lagoon of Venice. Vie Milieu 1999, 49, 19–31. [Google Scholar]
- Harris, R.P.; Wiebe, P.H.; Lenz, J.; Skjoldal, H.R.; Huntley, M. ICES Zooplankton Methodology Manual; Academic Press: Cambridge, MA, USA, 2000; p. 684. [Google Scholar]
- Camatti, E.; Ferrari, I. Mesozooplankton. In Metodologie di Studio Del Plancton Marino; Socal, G., Buttino, I., Cabrini, M., Mangoni, O., Penna, A., Totti, C., Eds.; Manuali e Linee Guida: Ispra, Italy, 2010; Volume 56, pp. 489–506. [Google Scholar]
- Throndsen, J. Preservation and storage. In Phytoplankton Manual; Sournia, A., Ed.; UNESCO: Paris, France, 1978. [Google Scholar]
- Utermöhl, H. Zur Vervollkommnung der quantitativen Phytoplankton Methodik. Int. Ver. Theor. Angew. Limnol. 1958, 9, 1–38. [Google Scholar] [CrossRef]
- Zingone, A.; Honsell, G.; Marino, D.; Montresor, M.; Socal, G. Fitoplankton. Nov. Thalass. 1990, 11, 183–198. [Google Scholar]
- EPA (Environmental Protection Agency). Methods for Determination of Chemical Substances in Marine and Estuarine Environmental Matrices, 2nd ed.; EPA: Cincinnati, OH, USA, 1997.
- IRSA (Istituto di Ricerca sulle Acque). Metodi di Analisi Per Acque di Mare. Quad. Ist. Ric. Acque 1997, 59, 570–571. [Google Scholar]
- Hirsch, R.M.; Slack, J.R.; Smith, R.A. Techniques of trend analysis for monthly water quality data. Water Resour. Res. 1982, 18, 107–121. [Google Scholar] [CrossRef]
- Kendall, M.G. Rank Correlation Methods, 4th ed.; Charles Griffin: London, UK, 1975. [Google Scholar]
- Legendre, P.; Legendre, L.F. Numerical Ecology, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2012; Volume 24, p. 1006. [Google Scholar]
- Davis, J.C. Statistics and Data Analysis in Geology, 3rd ed.; Wiley, J.S., Ed.; Wiley: Hoboken, NJ, USA, 1986; p. 646. [Google Scholar]
- Dolédec, S.; Chessel, D.; Gimaret-Carpentier, C. Niche separation in community analysis: A new method. Ecology 2000, 81, 2914–2927. [Google Scholar] [CrossRef]
- Sen, P.K. Estimates of the regression coefficient based on Kendall’s Tau. J. Am. Stat. Assoc. 1968, 63, 1379–1389. [Google Scholar] [CrossRef]
- Zenetos, A.; Gofas, S.; Verlaque, M.; Cinar, M.E.; Garcia Raso, J.E.; Bianchi, C.N.; Morris, C.; Azzurro, E.; Bilecenoglu, M.; Froglia, C.; et al. Alien species in the Mediterranean Sea by 2010. A contribution to the application of European Union’s Marine Strategy Framework Directive (MSFD). Part I. Spatial distribution. Mediterr. Mar. Sci. 2010, 11, 381–493. [Google Scholar] [CrossRef]
- Carlton, J.T. Biological invasions and cryptogenic species. Ecology 1996, 77, 1653–1655. [Google Scholar] [CrossRef]
- Ruiz, G.M.; Fofonoff, P.; Hines, A. Non- indigenous species as stressors in estuarine and marine communities: Assessing invasion impacts and interactions. Limnol. Oceanogr. 1999, 44, 950–972. [Google Scholar] [CrossRef]
- Mack, R.N.; Simberloff, D.; Lonsdale, W.M.; Evans, H.; Clout, M.; Bazzaz, F.A. Biotic invasions: Causes, epidemiology, global consequences and control. Ecol. Appl. 2000, 10, 689–710. [Google Scholar] [CrossRef]
- Gubanova, A. Occurrence of Acartia tonsa Dana in the Black Sea. Was it introduced from the Mediterranean? Mediterr. Mar. Sci. 2000, 1, 105–109. [Google Scholar] [CrossRef]
- Gubanova, A.; Altukhova, D.; Stefanova, K.; Arashkevichc, E.; Kamburskad, L.; Prusovaa, I.; Svetlichnya, L.; Timoftee, F.; Uysalfand, Z. Species composition of Black Sea marine planktonic copepods. J. Mar. Syst. 2014, 135, 44–52. [Google Scholar] [CrossRef]
- Vidjak, O.; Bojanić, N.; de Olazabal, A.; Benzi, M.; Brautović, I.; Camatti, E.; Hure, M.; Lipej, L.; Lučić, D.; Pansera, M.; et al. Zooplankton in Adriatic port environments: Indigenous communities and non-indigenous species. Mar. Pollut. Bull. 2008. [Google Scholar] [CrossRef]
- Ruiz, G.M.; Carlton, J.T.; Grosholz, E.D.; Hines, A.H. Global invasions of marine and estuarine habitats by non-indigenous species: Mechanisms, extent and consequences. Am. Zool. 1997, 37, 621–632. [Google Scholar] [CrossRef]
- Comaschi, A.; Acri, F.; Alberghi, L.; Bastianini, M.; Bianchi, F.; Cavalloni, B.; Socal, G. Presenza di Acartia tonsa (Copepoda: Calanoida) nella laguna di Venezia. Biol. Mar. Mediterr. 1994, 1, 273–274. [Google Scholar]
- Bandelj, V.; Socal, G.; Park, Y.S.; Lek, S.; Coppola, J.; Camatti, E.; Capuzzo, E.; Milani, L.; Solidoro, C. Analysis of multitrophic plankton assemblages in the lagoon of Venice. Mar. Ecol. Prog. Ser. 2008, 368, 23–40. [Google Scholar] [CrossRef]
- Comaschi, A.; Acri, F.; Bianchi, F.; Bressan, M.; Camatti, E. Temporal changes of species belonging to Acartia genus (Copepoda: Calanoida) in the northern basin of the Venice lagoon. Boll. Mus. Civ. Stor. Nat. Venezia 2000, 50, 189–193. [Google Scholar]
- Camatti, E.; Comaschi, A.; Coppola, J.; Milani, L.; Minocci, M.; Socal, G. Analisi dei popolamenti zooplanctonici nella laguna di Venezia dal 1975 al 2004. Biol. Mar. Mediterr. 2006, 13, 46–53. [Google Scholar]
- Sei, S.; Ferrari, I. First report of the occurrence of Acartia tonsa (Copepoda, Calanoida) in the Lesina lagoon (South Adriatic Sea–Mediterranean Sea). Mar. Biod. Rec. 2008, 1, 37. [Google Scholar] [CrossRef]
- Sei, S.; Rossetti, G.; Villa, F.; Ferrari, I. Zooplankton variability related to environmental changes in a eutrophic coastal lagoon in the Po Delta. Hydrobiologia 1996, 329, 45–55. [Google Scholar] [CrossRef]
- Sfriso, A.; Buosi, A. Trophic status changes in the Venice lagoon during the last 40 years. In Proceedings of the 49th Congresso della Società Italiana di Biologia Marina Cesenatico (FC), Rome, Italy, 4–8 June 2018. [Google Scholar]
- Sfriso, A. Flora and vertical distribution of macroalgae in the lagoon of Venice: A comparison with previous studies. Plat Biosyst. 1987, 121, 69–85. [Google Scholar] [CrossRef]
- Sfriso, A.; Birkemeyer, T.; Ghetti, P.F. Benthic macrofauna changes in areas of Venice lagoon populated by seagrasses or seaweeds. Mar. Environ. Res. 2001, 52, 323–349. [Google Scholar] [CrossRef]
- Sfriso, A.; Facca, C.; Ghetti, P.F. Temporal and spatial changes of macroalgae and phytoplankton in a Mediterranean coastal area: The Venice lagoon as a case study. Mar. Environ. Res. 2003, 56, 617–636. [Google Scholar] [CrossRef]
- Sfriso, A.; Facca, C.; Ceoldo, S.; Silvestri, S.; Ghetti, P.F. Role of macroalgal biomass and clam fishing on spatial and temporal changes in N and P sedimentary pools in the central part of the Venice lagoon. Oceanol. Acta 2003, 26, 3–13. [Google Scholar] [CrossRef]
- Sfriso, A.; Marcomini, A. Decline of Ulva growth in the Lagoon of Venice. Bioresour. Technol. 1996, 58, 299–307. [Google Scholar] [CrossRef]
- Decker, M.B.; Breitburg, D.L.; Marcus, N.H. Geographical differences in behavioral responses to hypoxia: Local adaptation to an anthropogenic stressor? Ecol. Appl. 2003, 13, 1104–1109. [Google Scholar] [CrossRef]
- Kimmel, D.G.; Boicourt, W.C.; Pierson, J.J.; Roman, M.R.; Zhang, X. A comparison of the mesozooplankton response to hypoxia in Chesapeake Bay and the northern Gulf of Mexico using the biomass size spectrum. J. Exp. Mar. Biol. Ecol. 2009, 381, 65–73. [Google Scholar] [CrossRef]
- Mauchline, J. The biology of calanoid copepods. Adv. Mar. Biol. 1998, 33, 1–710. [Google Scholar]
- Castro-Longoria, E. Egg Production and Hatching Success of Four Acartia Species under Different Temperature and Salinity Regimes. J. Crustac. Biol. 2003, 23, 289–299. [Google Scholar] [CrossRef]
- El Shabrawy, G.M.; Belmonte, G. Abundance and affirmation of Paracartia latisetosa (copepoda, calanoida) in the inland lake Qarun (Egypt). Thalass. Sal. 2004, 27, 151–160. [Google Scholar]
- Kritchagin, N. Otchet o faunisticheskikh issledovaniyakh proizvedennikh v etom godu po porutcheniyu kievskogo obsh estva estestvoispitateley na beregakh Tchernogo morya. Zap. Kiev. Obs. Estestvoispitatalei 1873, 3, 346–430. [Google Scholar]
- Aravena, G.; Villate, F.; Uriarte, I.; Iriarte, A.; Ibáñez, B. Response of Acartia populations to environmental variability and effects of invasive congenerics in the estuary of Bilbao, Bay of Biscay. Estuar. Coast. Shelf Sci. 2009, 83, 621–628. [Google Scholar] [CrossRef]
- Gaudy, R.; Cervetto, G.; Pagano, M. Comparison of the metabolism of Acartia clausi and Acartia tonsa: Influence of temperature and salinity. J. Exp. Mar. Biol. Ecol. 2000, 247, 51–65. [Google Scholar] [CrossRef]
- Cervetto, G.; Gaudy, R.; Pagano, M. Influence of salinity on the distribution of Acartia tonsa (Copepoda, Calanoida). J. Exp. Mar. Biol. Ecol. 1999, 239, 33–45. [Google Scholar] [CrossRef]
- Biancalana, F.; Dutto, M.; Berasategui, A.; Kopprio, G.; Hoffmeyer, M. Mesozooplankton assemblages and their relationship with environmental variables: A study case in a disturbed bay (Beagle Channel, Argentina). Environ. Monit. Assess. 2014, 186, 8629–8647. [Google Scholar] [CrossRef] [PubMed]
- Dunbar, F.N.; Webber, M.K. The phytoplankton distribution in the eutrophic Kingston Harbour, Jamaica. Bull. Mar. Sci. 2003, 73, 343–359. [Google Scholar]
- Malej, A.; Tirelli, V.; Lučić, D.; Paliaga, P.; Vodopivec, M.; Goruppi, A.; Ancona, S.; Benzi, M.; Bettoso, N.; Camatti, E.; et al. Mnemiopsis leidyi in the northern Adriatic: Here to stay? J. Sea Res. 2017, 124, 10–16. [Google Scholar] [CrossRef]
- Irvine, K.; Moss, B.; Bales, M.; Snook, D. The changing ecosystem of a shallow, brackish lake, Hickling Broad, Norfolk, UK II. Trophic relationships with special reference to the role of Neomysis integer. Freshw. Biol. 1993, 29, 119–139. [Google Scholar] [CrossRef]
- Li, M.; Gargett, A.; Denman, K. What determines seasonal and interannual variability of phytoplankton and zooplankton in strongly estuarine systems? Application to the semi enclosed estuary of strait of Georgia and Juan de Fuca Strait. Estuar. Coast. Shelf Sci. 2000, 50, 467–488. [Google Scholar] [CrossRef]
- Poggiale, J.C.; Dauvin, J.C. Long term dynamics of three benthic Ampelisca (Crustacea Amphipoda) populations from the Bay of Morlaix (western English Channel) related to their disappearance after the ‘‘Amoco Cadiz’’ oil spill. Mar. Ecol. Prog. Ser. 2001, 214, 201–209. [Google Scholar] [CrossRef]
- Gucu, A.C. Can overfishing be responsible for the successful establishment of Mnemiopsis leidyi in the Black sea? Estuar. Coast. Shelf Sci. 2002, 54, 439–451. [Google Scholar] [CrossRef]
- Hays, G.C.; Richardson, A.J.; Robinson, C. Climate change and marine plankton. Trends Ecol. Evol. 2005, 20, 337–344. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Acartia clausi | Acartia margalefi | Acartia tonsa | Total Zooplankton | ||
|---|---|---|---|---|---|
| San Giuliano | Tau | −0.121 | −0.577 | 0.243 | 0.300 |
| p | 0.311 | 0.000 | 0.040 | 0.011 | |
| Fusina | Tau | −0.125 | −0.687 | −0.009 | −0.008 |
| p | 0.303 | 0.000 | 0.943 | 0.943 | |
| Marghera | Tau | −0.346 | −0.728 | −0.153 | −0.142 |
| p | 0.003 | 0.000 | 0.199 | 0.228 | |
| Lido | Tau | −0.144 | −0.540 | −0.094 | −0.083 |
| p | 0.225 | 0.003 | 0.430 | 0.480 | |
| Palude della Rosa | Tau | −0.059 | −0.688 | 0.078 | 0.067 |
| p | 0.620 | 0.000 | 0.514 | 0.572 |
| Stations | 1M | 2B | 7B | 9B | 11B | 14B | 17B | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Avg | SD | Avg | SD | Avg | SD | Avg | SD | Avg | SD | Avg | SD | Avg | SD | |
| Temperature (°C) | 16 | 8 | 16 | 9 | 17 | 9 | 18 | 8 | 16 | 10 | 16 | 10 | 16 | 9 |
| Salinity | 35 | 2 | 29 | 4 | 32 | 4 | 32 | 3 | 34 | 3 | 35 | 2 | 33 | 4 |
| Relative oxygen (%) | 96 | 5 | 100 | 9 | 102 | 15 | 90 | 7 | 83 | 19 | 92 | 7 | 95 | 5 |
| POC (µg L−1) | 308 | 73 | 750 | 630 | 747 | 415 | 668 | 386 | 485 | 211 | 345 | 141 | 947 | 886 |
| Chlorophyll a (µg L−1) | 1.78 | 0.92 | 4.40 | 4.32 | 7.02 | 6.21 | 7.81 | 12.69 | 2.09 | 1.46 | 1.54 | 0.55 | 3.84 | 4.52 |
| Total phytoplankton (cell L−1) | 2541 | 3176 | 10,831 | 16,019 | 4382 | 4592 | 5716 | 6874 | 2082 | 2406 | 2128 | 1993 | 8318 | 15,745 |
| Acartia tonsa(ind m−3) | 23 | 69 | 5998 | 10,333 | 976 | 1658 | 4456 | 11,209 | 2823 | 4986 | 75 | 192 | 1234 | 2744 |
| Other copepods (ind m−3) | 3702 | 5494 | 316 | 415 | 521 | 1015 | 257 | 312 | 602 | 588 | 588 | 1170 | 144 | 167 |
| Cladocerans (ind m−3) | 186 | 328 | 2 | 5 | 8 | 26 | 4 | 10 | 9 | 14 | 8 | 22 | 0 | 1 |
| Other zooplankton (ind m−3) | 142 | 115 | 69 | 110 | 136 | 290 | 176 | 244 | 161 | 272 | 56 | 54 | 42 | 53 |
| Total zooplankton (ind m−3) | 4053 | 5906 | 6386 | 10328 | 1642 | 2062 | 4893 | 11349 | 3595 | 5207 | 727 | 1233 | 1419 | 2781 |
| 1M | 2B | 7B | 9B | 11B | 14B | 17B | |
|---|---|---|---|---|---|---|---|
| Acartia clausi | 69 | 1 | 21 | 1 | 8 | 59 | 4 |
| Acartia tonsa | 1 | 94 | 59 | 91 | 79 | 10 | 87 |
| Centropages ponticus | 4 | 1 | 2 | 1 | 2 | 2 | 1 |
| Corycaeus spp. | 1 | ||||||
| Harpacticoid spp. | 3 | 2 | 1 | ||||
| Oithona similis | 1 | 6 | |||||
| Oncaea spp. | 2 | ||||||
| Paracalanus parvus | 14 | 2 | 4 | 2 | 5 | 5 | 2 |
| Pseudocalanus elongatus | 1 | 1 | 1 | 1 | |||
| Penilia avirostris | 1 | ||||||
| Podon polyphemoides | 3 | ||||||
| Chaetognaths | 1 | ||||||
| Cirriped larvae | 1 | 1 | |||||
| Decapod larvae | 6 | 2 | 3 | 1 | 1 | ||
| Fish eggs | 1 | 1 | |||||
| Gastropod larvae | 2 |
| Temperature | Salinity | Oxygen | Dissolved Inorganic Nitrogen (DIN) | Chlorophyll a | Particulate Organic Carbon (POC) | Phytoplankton | |
|---|---|---|---|---|---|---|---|
| (°C) | (%) | (µg L−1) | (µg L−1) | (µg L−1) | (cell L−1) | ||
| Acartia clausi | −0.15 | −0.09 | 0.02 | 0.09 | −0.03 | −0.03 | −0.10 |
| Acartia margalefi | 0.38 | −0.07 | 0.09 | −0.31 | 0.31 | 0.24 | 0.35 |
| Acartia tonsa | 0.49 | 0.12 | −0.17 | −0.48 | 0.54 | 0.45 | 0.62 |
| Centropages ponticus | 0.49 | 0.41 | −0.25 | −0.54 | 0.35 | 0.25 | 0.44 |
| Corycaeus spp. | −0.54 | −0.05 | 0.14 | 0.39 | −0.43 | −0.37 | −0.53 |
| Harpacticoid spp. | 0.17 | −0.07 | 0.01 | −0.10 | 0.20 | 0.23 | 0.12 |
| Oithona similis | −0.43 | −0.07 | 0.20 | 0.24 | −0.30 | −0.21 | −0.37 |
| Oncaea spp. | −0.39 | 0.05 | 0.02 | 0.23 | −0.28 | −0.31 | −0.36 |
| Paracalanus parvus | 0.08 | 0.31 | −0.11 | −0.19 | 0.09 | 0.04 | 0.07 |
| Pseudocalanus elongatus | −0.58 | −0.15 | 0.20 | 0.43 | −0.45 | −0.33 | −0.48 |
| Penilia avirostris | 0.19 | 0.30 | −0.25 | −0.24 | 0.14 | 0.01 | 0.16 |
| Podon polyphemoides | 0.17 | 0.06 | −0.09 | −0.22 | 0.15 | 0.01 | 0.24 |
| Chaetognats | 0.13 | 0.29 | −0.15 | −0.23 | 0.11 | −0.03 | 0.15 |
| Cirriped nauplii | 0.42 | 0.10 | −0.18 | −0.35 | 0.41 | 0.31 | 0.33 |
| Decapod larvae | 0.36 | 0.25 | −0.20 | −0.46 | 0.37 | 0.30 | 0.42 |
| Fish eggs | 0.44 | 0.24 | −0.17 | −0.41 | 0.39 | 0.19 | 0.42 |
| Gastropod larvae | 0.19 | 0.21 | −0.11 | −0.24 | 0.13 | 0.05 | 0.08 |
| Species | Inertia | OMI | Tol | RTol | p |
|---|---|---|---|---|---|
| Acartia tonsa | 12.76 | 2.80 | 1.35 | 8.61 | 0.497 |
| Acartia clausi | 9.37 | 3.13 | 2.08 | 4.15 | 0.012 |
| Paracalanus parvus | 11.78 | 3.91 | 2.58 | 5.28 | 0.034 |
| Decapoda larvae | 19.07 | 11.62 | 5.09 | 2.36 | 0.620 |
| Acartia margalefi | 6.37 | 1.66 | 0.90 | 3.80 | 0.880 |
| Podonidae | 7.09 | 2.36 | 0.30 | 4.43 | 0.818 |
| Penilia avirostris | 8.46 | 5.69 | 0.49 | 2.28 | 0.030 |
| Appendicularia | 9.84 | 3.35 | 2.38 | 4.09 | 0.567 |
| Centropages ponticus | 8.07 | 2.95 | 1.29 | 3.82 | 0.214 |
| Ascidiacea larvae | 14.44 | 10.33 | 1.55 | 2.56 | 0.728 |
| Echinodermata larvae | 10.35 | 6.66 | 0.79 | 2.91 | 0.015 |
| Oithona similis | 10.68 | 6.38 | 0.93 | 3.36 | 0.011 |
| Nauplii copepoda | 7.95 | 1.93 | 0.90 | 5.12 | 0.819 |
| Evadne nordmani | 7.08 | 5.52 | 0.21 | 1.35 | 0.849 |
| Harpacticoida | 13.32 | 1.63 | 2.48 | 9.22 | 0.802 |
| Clausocalanus spp. | 12.91 | 8.93 | 0.84 | 3.15 | 0.268 |
| Oncaeidae | 13.21 | 8.44 | 1.74 | 3.03 | 0.055 |
| Bivalvia larvae | 12.25 | 1.72 | 1.45 | 9.08 | 0.690 |
| Polychaeta larvae | 9.46 | 1.77 | 1.01 | 6.67 | 0.214 |
| Paracartia latisetosa1 | 9.80 | 4.41 | 3.11 | 2.29 | 0.910 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Camatti, E.; Pansera, M.; Bergamasco, A. The Copepod Acartia tonsa Dana in a Microtidal Mediterranean Lagoon: History of a Successful Invasion. Water 2019, 11, 1200. https://doi.org/10.3390/w11061200
Camatti E, Pansera M, Bergamasco A. The Copepod Acartia tonsa Dana in a Microtidal Mediterranean Lagoon: History of a Successful Invasion. Water. 2019; 11(6):1200. https://doi.org/10.3390/w11061200
Chicago/Turabian StyleCamatti, Elisa, Marco Pansera, and Alessandro Bergamasco. 2019. "The Copepod Acartia tonsa Dana in a Microtidal Mediterranean Lagoon: History of a Successful Invasion" Water 11, no. 6: 1200. https://doi.org/10.3390/w11061200
APA StyleCamatti, E., Pansera, M., & Bergamasco, A. (2019). The Copepod Acartia tonsa Dana in a Microtidal Mediterranean Lagoon: History of a Successful Invasion. Water, 11(6), 1200. https://doi.org/10.3390/w11061200