Bio-Ecological Traits of Western Tubenose Goby Proterorhinus semilunaris (Heckel, 1837): A Key to Understand Its Invasion Success

Abstract
1. Introduction
2. Materials and Methods
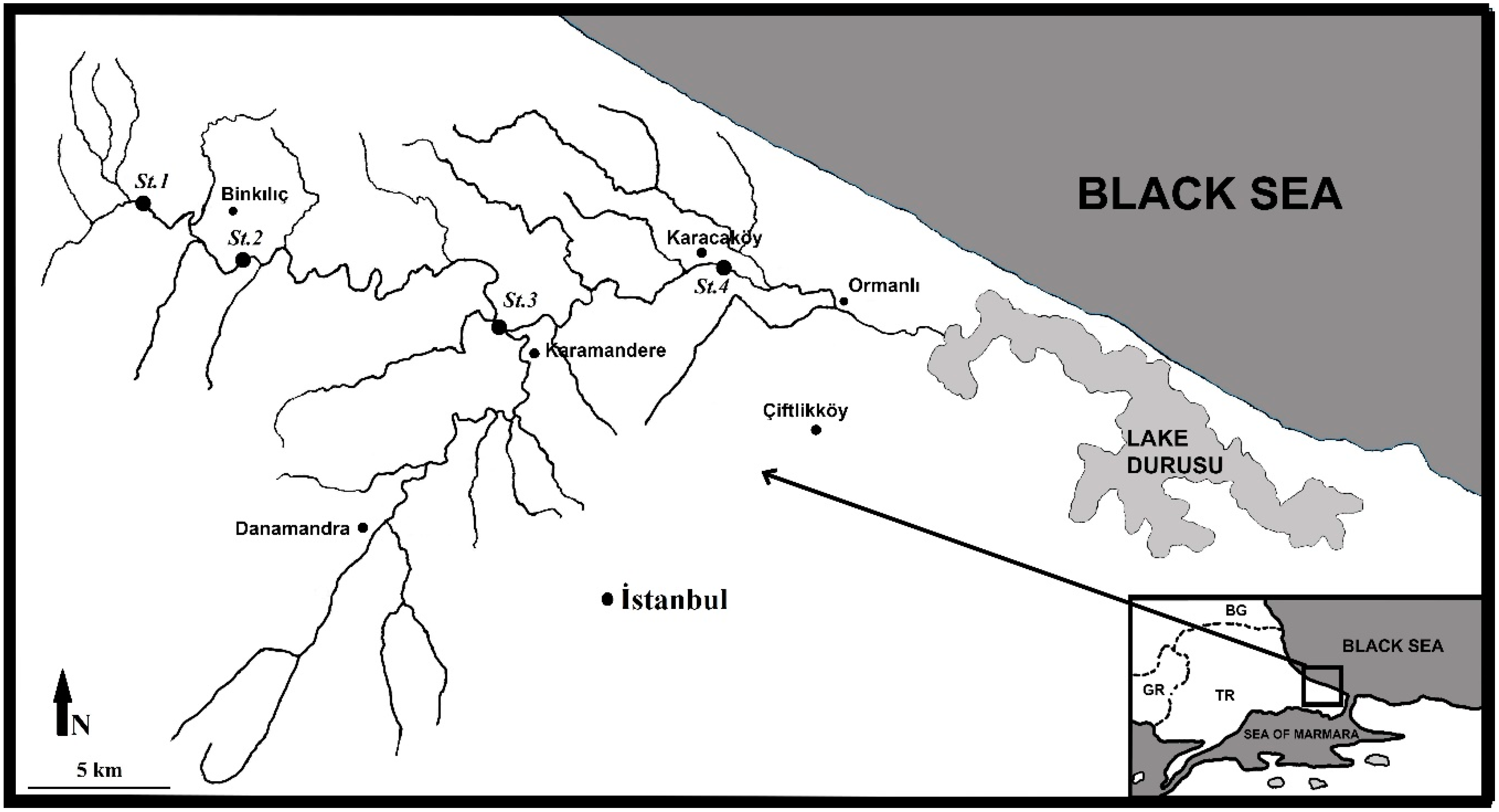
2.1. Study Area
- Şeytan Stream (St.1)-41°25′03.0000″, 28°08′18.4200″; a small (average 4 m width) and shallow (average 16 cm depth) stream with stony (cobbles) substrate.
- Binkılıç Stream (St.2)-41°23′56.4360″, 28°11′37.1760″; water depth varies depending on seasons (15–65 cm), and the substrate is mostly stony (cobbles and gravel).
- Karamandere Stream (St.3)-41°22′45.1200″, 28°17′45.9600″; the deepest (average 54 cm) station, and the substrate is mostly stony (boulders and cobbles) with leaf detritus.
- Karacaköy Stream (St.4)-41°23′58.0560″, 28°23′00.6720″; the stream bed and depth vary depending on seasons (3–16 m width and 11–45 cm depth), and the substrate is mostly sandy (rarely boulders and cobbles) (Figure 1).
2.2. Sampling
2.3. Growth Type
2.4. Reproduction
2.5. Feeding
3. Results
3.1. Biometrics
3.2. Growth
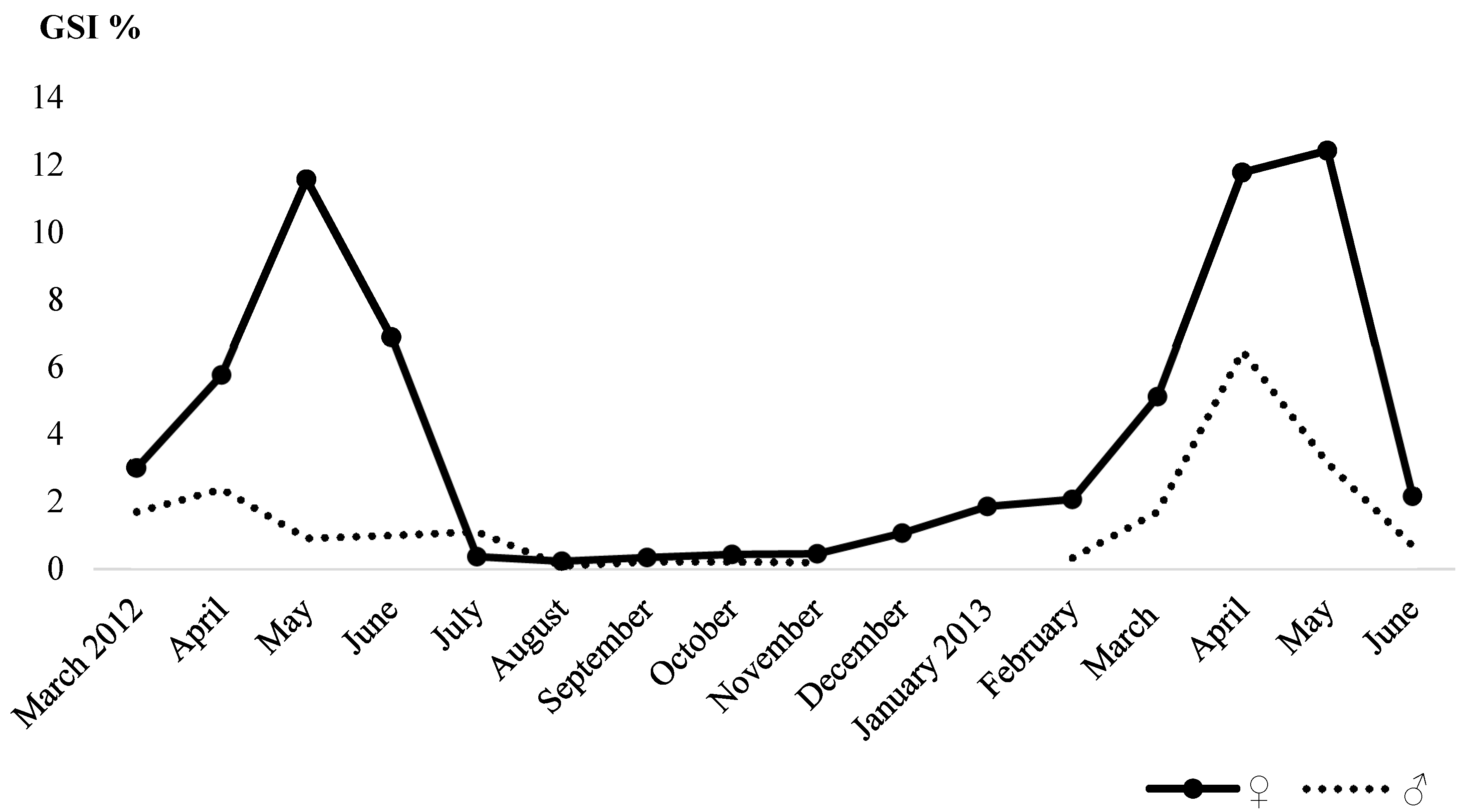
3.3. Reproduction
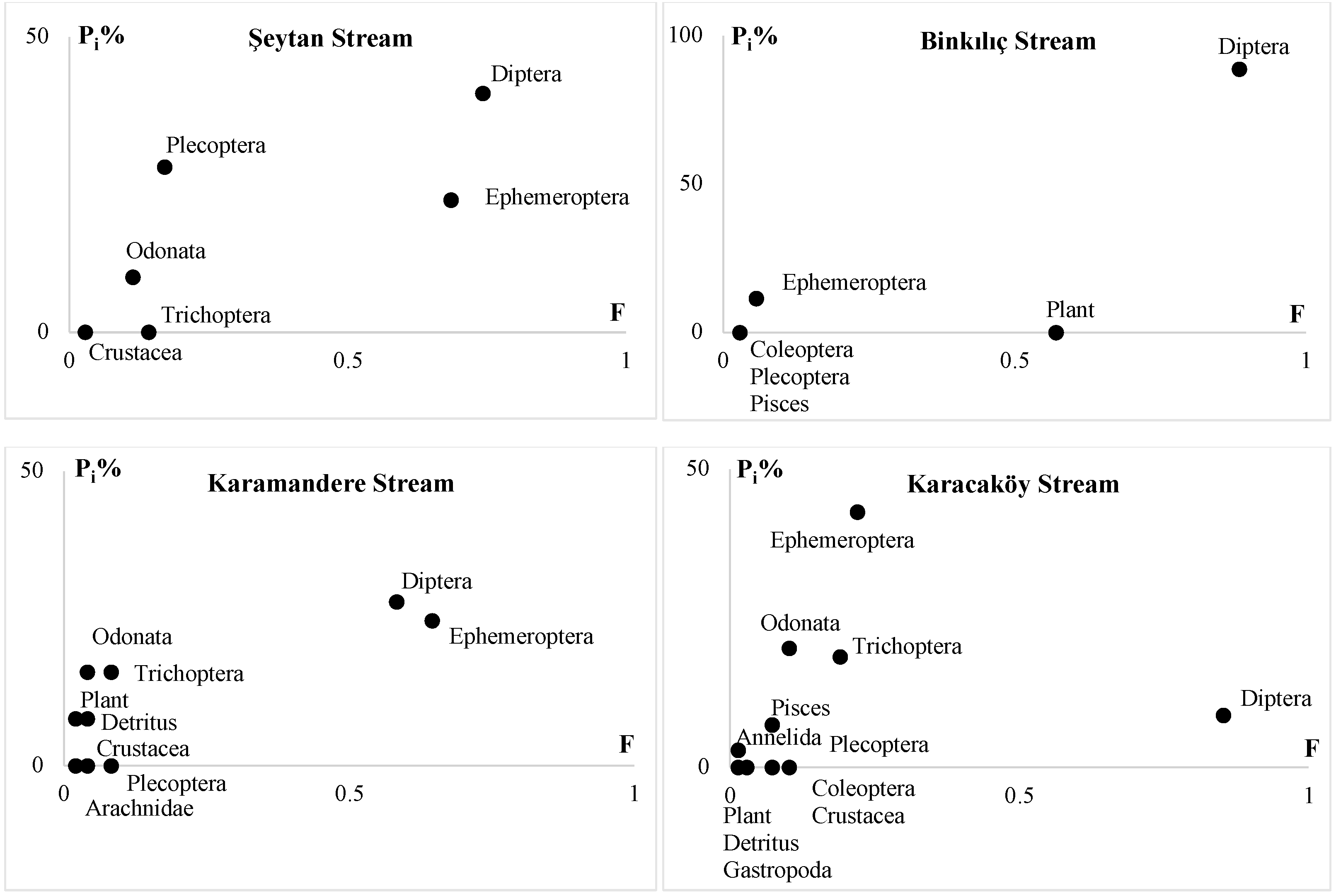
3.4. Feeding
4. Discussion
Funding
Acknowledgments
Conflicts of Interest
References
- Kottelat, M.; Freyhof, J. Handbook of European Freshwater Fishes; Publications Kottelat, Cornol and Freyhof: Berlin, Germany, 2007; p. 646. [Google Scholar]
- Von Landwüst, C. Expansion of Proterorhinus marmoratus (Teleostei, Gobiidae) into the River Moselle (Germany). Folia Zool. 2006, 55, 107–111. [Google Scholar]
- Jude, D.J.; Reider, R.H.; Smith, G.R. Establishment of Gobiidae in the Great Lakes Basin. Can. J. Fish. Aquat. Sci. 1992, 49, 416–421. [Google Scholar] [CrossRef]
- Kocovsky, P.M.; Tallman, J.A.; Jude, D.J.; Murphy, D.M.; Brown, J.E.; Stepien, C.A. Expansion of tubenose gobies Proterorhinus semilunaris into western Lake Erie and potential effects on native species. Biol. Invasions 2011, 13, 2775–2784. [Google Scholar] [CrossRef]
- Von Kessel, N.; Dorenbosch, M.; Kranenbarg, J.; Van der Velde, G.; Leuven, R.S.E.W. Invasive Ponto-Caspian gobies rapidly reduce the abundance of protected native bullhead. Aquat. Invasions 2016, 11, 179–188. [Google Scholar] [CrossRef]
- Manné, S.; Poulet, N. First record of the western tubenose goby Proterorhinus semilunaris (Heckel, 1837) in France. Knowl. Manag. Aquat. Ecosyst. 2008, 389, 3. [Google Scholar] [CrossRef]
- Adámek, Z.; Jurajda, P.; Prášek, V.; Sukop, I. Seasonal diet pattern of non-native tubenose goby (Proterorhinus semilunaris) in a lowland reservoir (Mušov, Czech Republic). Knowl. Manag. Aquat. Ecosyst. 2010, 397, 2. [Google Scholar] [CrossRef][Green Version]
- Tarkan, A.S.; Gaygusuz, Ö.; Özuluğ, M.; Gürsoy Gaygusuz, Ç.; Saç, G. Length–weight relationships of six freshwater fishes from the small streams flowing into Lake Sapanca, NW Turkey. J. Appl. Ichthyol. 2009, 25, 230–231. [Google Scholar] [CrossRef]
- Gürsoy Gaygusuz, Ç.; Tarkan, A.S.; Gaygusuz, Ö. The diel changes in feeding activity, microhabitat preferences and abundance of two freshwater fish species in small temperate streams (Ömerli, İstanbul). Ekoloji 2010, 19, 15–24. [Google Scholar] [CrossRef]
- Tarkan, A.S.; Karakuş, U.; Tepeköy, E.G.; Top, N.; Yalçın-Özdilek, Ş.; Partal, N.; Britton, J.R. Trophic interactions of two Ponto-Caspian Gobies in the Turkish part of their native range. Turk. J. Fish. Aquat. Sci. 2018, 18, 1279–1286. [Google Scholar] [CrossRef]
- Top, N.; Karakuş, U.; Tepeköy, E.G.; Britton, J.R.; Tarkan, A.S. Plasticity in life history traits of the native Proterorhinus semilunaris suggests high adaptive capacity in its invasive range. Knowl. Manag. Aquat. Ecosyst. 2018, 419, 48. [Google Scholar] [CrossRef]
- Zar, J.H. Biostatistical Analysis; Prentice Hall: Englewoods Cliffs, NJ, USA, 1999; p. 663. [Google Scholar]
- Le Cren, E.D. The length–weight relationship and seasonal cycle in gonad weight and condition in the perch (Perca fluviatilis). J. Anim. Ecol. 1951, 20, 201–218. [Google Scholar] [CrossRef]
- Froese, R. Cube law, condition factor, weight-length relationships: History, meta-analysis and recommendations. J. Appl. Ichthyol. 2006, 22, 241–253. [Google Scholar] [CrossRef]
- King, M. Fisheries Biology, Assessment and Management, 2nd ed.; Blackwell Scientific Publications: Oxford, UK, 2007; p. 381. [Google Scholar]
- Ricker, W.E. Computations and interpretation of biological statistics of fish populations. Fish. Res. Bd. Can. Bull. 1975, 191, 1–382. [Google Scholar]
- Sarkhanizadeh, A.; Valinassab, T.; Fatemi, M.R. Evaluation of some feeding indices of Atrobucca nibe in the Northern Oman Sea. J. Novel Appl. Sci. 2014, 3, 1268–1274. [Google Scholar]
- Hyslop, E.J. Stomach contents analyses—A review of methods and their application. J. Fish Biol. 1980, 17, 411–429. [Google Scholar] [CrossRef]
- Hayse, J.W. Feeding habits, age, growth, and reproduction of Atlantic spadefish Chaetodipterus faber (Pisces: Ephippidae) in South Carolina. Fish. Bull. 1990, 88, 67–83. [Google Scholar]
- Levins, R. Evolution in Changing Environments; Princeton University Press: Princeton, NJ, USA, 1968; p. 132. [Google Scholar]
- Hurlbert, S.H. The measurements of niche overlap and some relatives. Ecology 1978, 59, 67–77. [Google Scholar] [CrossRef]
- Krebs, C.J. Ecological Methodology; Addison-Wesley Educational Publishers: Menlo Park, CA, USA, 1998. [Google Scholar]
- Novakowski, G.G.; Hahn, N.S.; Fugi, R. Diet seasonality and food overlap of the fish assemblage in a pantanal pond. Neotrop. Ichthyol. 2008, 6, 567–576. [Google Scholar] [CrossRef]
- Costello, M.J. Predator feeding strategy and prey importance: A new graphical analysis. J. Fish Biol. 1990, 36, 261–263. [Google Scholar] [CrossRef]
- Amundsen, P.A.; Gabler, H.M.; Staldvik, F.J. A new approach to graphical analysis of feeding strategy from stomach contents data—Modification of the Costello (1990) method. J. Fish Biol. 1996, 48, 607–614. [Google Scholar]
- Tarkan, A.S.; Gaygusuz, Ö.; Acıpınar, H.; Gürsoy, Ç.; Özuluğ, M. Length–weight relationship of fishes from the Marmara region (NW-Turkey). J. Appl. Ichthyol. 2006, 22, 271–273. [Google Scholar] [CrossRef]
- Reichard, M.; Polačik, M.; Blažek, R.; Vrtílek, M. Female bias in the adult sex ratio of African annual fishes: Interspecific differences, seasonal trends and environmental predictors. Evolut. Ecol. 2014, 28, 1105–1120. [Google Scholar] [CrossRef]
- Székely, T.; Weissing, F.J.; Komdeur, J. Adult sex ratio variation: Implications for breeding systems. J. Evolut. Biol. 2014, 27, 1500–1512. [Google Scholar] [CrossRef] [PubMed]
- Field, D.L.; Pickup, M.; Barrett, S.C. Comparative analyses of sex-ratio variation in dioecious flowering plants. Evolution 2012, 67, 661–672. [Google Scholar] [CrossRef]
- Valová, Z.; Konečná, M.; Janáč, M.; Jurajda, P. Population and reproductive characteristics of a non-native western tubenose goby (Proterorhinus semilunaris) population unaffected by gobiid competitors. Aquat. Invasions 2015, 10, 57–68. [Google Scholar] [CrossRef]
- Hernaman, V.; Munday, P.L. Life-history characteristics of coral reef gobies. II. Mortality rate, mating system and timing of maturation. Mar. Ecol. Prog. Ser. 2005, 290, 223–237. [Google Scholar] [CrossRef]
- Grabowska, J.; Przybylski, M. Life-history traits of non-native freshwater fish invaders differentiate them from natives in the Central European bioregion. Rev. Fish Biol. Fish. 2015, 25, 165–178. [Google Scholar] [CrossRef]
- French, J.R.P.; Jude, D.J. Diets and diet overlap of nonindigenous Gobies and small benthic native fishes co-inhabiting the St. Clair River, Michigan. J. Great Lakes Res. 2001, 27, 300–311. [Google Scholar] [CrossRef]
- Všetičková, L.; Janáč, M.; Vašek, M.; Roche, K.; Jurajda, P. Non-native western tubenose gobies Proterorhinus semilunaris show distinct site, sex and age-related differences in diet. Knowl. Manag. Aquat. Ecosyst. 2014, 414, 10. [Google Scholar] [CrossRef]
- Shoryngin, A.A. Food and Feeding Relationships of Fish in the Caspian Sea; Pishchepromizdat: Moscow, Russia, 1952; p. 268. (In Russian) [Google Scholar]
- Ghedotti, M.J.; Smihula, J.C.; Smith, G.R. Zebra mussel predation by round gobies in the laboratory. J. Great Lakes Res. 1995, 21, 665–669. [Google Scholar] [CrossRef]
- Charlebois, P.M.; Marsden, J.E.; Goettel, R.G.; Wolfe, R.K.; Jude, D.J.; Rudnicka, S. The Round Goby, Neogobius melanostomus (PALLAS), a Review of European and North American Literature. Illinois-Indiana Sea Grant Program and Illinois Natural History Survey; INHS Special Pub.: Champaign, IL, USA, 1997; Volume 20, p. 61. [Google Scholar]
- Tarkan, A.S.; Tepeköy, E.G.; Karakuş, U.; Top, N.; Vilizzi, L. Plasticity in the feeding ecology of Native Ponto-Caspian gobies suggests establishment success in their non-native range. Int. Rev. Hydrobiol. 2019, in press. [Google Scholar] [CrossRef]
- Adámek, Z.; Andreji, J.; Gallardo, J.M. Food habits of four bottom-dwelling Gobiid species at the confluence of the Danube and Hron Rivers (South Slovakia). Int. Rev. Hydrobiol. 2007, 94, 554–563. [Google Scholar] [CrossRef]
- Keiper, J.B.; Walton, W.E.; Foote, B.A. Biology and ecology of higher Diptera from freshwater wetlands. Annu. Rev. Entomol. 2002, 47, 207–232. [Google Scholar] [CrossRef] [PubMed]
- Saç, G.; Özuluğ, M. Effects of environmental variables on the distribution of fish assemblages in an endorheic stream (İstanbul, Turkey). Fresenius Environ. Bull. 2017, 26, 7150–7159. [Google Scholar]
- Ab Hamid, S.; Md Rav, C.S. Application of aquatic insects (Ephemeroptera, Plecoptera and Trichoptera) in water quality assessment of Malaysian Headwater. Trop. Life Sci. Res. 2017, 28, 143–162. [Google Scholar] [CrossRef]
- Dökümcü, N. Determination of Benthic Macroinvertebrate Fauna of Istranca Stream (Durusu-İstanbul). Master’s Thesis, Graduate School of Science and Engineering Biology Department, İstanbul University, Istanbul, Turkey, 2013; p. 69. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Sex | n | Standard Length (SL, cm) | Body Weight (W, g) | Regression Parameters | 95% Cl of a | 95% Cl of b | r2 | |||
|---|---|---|---|---|---|---|---|---|---|---|
| Mean ± SD | Min.–Max. | Mean ± SD | Min.–Max. | a | b | |||||
| Female | 154 | 3.8 ± 0.6 | 2.4–5.4 | 1.65 ± 0.84 | 0.32–4.29 | 0.024 | 3.109 | 0.020–0.029 | 2.976–3.242 | 0.928 |
| Male | 90 | 3.9 ± 0.7 | 2.3–6.0 | 1.83 ± 1.22 | 0.33–7.09 | 0.025 | 3.037 | 0.021–0.030 | 2.915–3.158 | 0.953 |
| All Ind. | 310 | 3.6 ± 0.9 | 1.4–6.1 | 1.57 ± 1.05 | 0.08–7.09 | 0.027 | 3.011 | 0.025–0.029 | 2.956–3.065 | 0.975 |
| Indices | Şeytan Stream (St.1) | Binkılıç Stream (St.2) | Karamandere Stream (St.3) | Karacaköy Stream (St.4) |
|---|---|---|---|---|
| VI% | 29.63 | 36.21 | 31.75 | 13.33 |
| B | 3.28 | 2.29 | 3.14 | 3.66 |
| BA | 0.46 | 0.26 | 0.27 | 0.24 |
| Preys | Şeytan Stream (St.1) (n = 35) | Binkılıç Stream (St.2) (n = 35) | Karamandere Stream (St.3) (n = 48) | Karacaköy Stream (St.4) (n = 68) | ||||
|---|---|---|---|---|---|---|---|---|
| MI% | IRI% | MI% | IRI% | MI% | IRI% | MI% | IRI% | |
| Insecta | 100.00 | 99.88 | 99.87 | 99.28 | ||||
| Diptera | 86.57 | 75.53 | 53.28 | 95.71 | ||||
| Ephemeroptera | 12.75 | 24.45 | 45.86 | 2.00 | ||||
| Plecoptera | 0.35 | 0.01 | 0.14 | 0.19 | ||||
| Trichoptera | 0.20 | - | 0.45 | 1.50 | ||||
| Odonata | 0.13 | - | 0.28 | 0.50 | ||||
| Coleoptera | - | 0.01 | - | 0.11 | ||||
| Pisces | - | 0.08 | - | 0.42 | ||||
| Crustacea | <0.01 | - | 0.05 | 0.12 | ||||
| Annelida | - | - | - | 0.16 | ||||
| Arachnida | - | - | <0.01 | - | ||||
| Gastropoda | - | - | - | <0.01 | ||||
| Plant | - | 0.03 | 0.01 | 0.01 | ||||
| Detritus | - | - | 0.06 | 0.01 | ||||
| Preys and Indices | Sp., 2012 (n = 64) | Su., 2012 (n = 23) | Au., 2012 (n = 21) | Wi., 2012 (n = 10) | Sp., 2013 (n = 60) | Su., 2013 (n = 8) |
|---|---|---|---|---|---|---|
| IRI% | ||||||
| Diptera | 91.40 | 80.00 | 80.00 | 90.21 | 93.93 | 65.39 |
| Plecoptera | 0.20 | 0.04 | 0.04 | 2.96 | 0.05 | |
| Trichoptera | 0.19 | 18.93 | 18.93 | 1.61 | 0.07 | |
| Odonata | 0.16 | 0.69 | 0.69 | 0.01 | 24.23 | |
| Ephemeroptera | 7.97 | 0.34 | 0.34 | 4.99 | 5.92 | 10.38 |
| Coleoptera | 0.07 | 0.24 | 0.02 | |||
| MI% | ||||||
| Fish | 0.01 | 0.10 | 0.02 | 2.76 | ||
| Detritus | 0.01 | 0.58 | ||||
| Crustacea | 0.14 | 0.43 | 0.01 | 0.01 | ||
| Arachnida | <0.01 | 0.11 | ||||
| Annelida | 0.19 | |||||
| Plant | 0.10 | 0.14 | <0.01 | 0.25 | ||
| Gastropoda | 0.02 | |||||
© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saç, G. Bio-Ecological Traits of Western Tubenose Goby Proterorhinus semilunaris (Heckel, 1837): A Key to Understand Its Invasion Success. Water 2019, 11, 1247. https://doi.org/10.3390/w11061247
Saç G. Bio-Ecological Traits of Western Tubenose Goby Proterorhinus semilunaris (Heckel, 1837): A Key to Understand Its Invasion Success. Water. 2019; 11(6):1247. https://doi.org/10.3390/w11061247
Chicago/Turabian StyleSaç, Gülşah. 2019. "Bio-Ecological Traits of Western Tubenose Goby Proterorhinus semilunaris (Heckel, 1837): A Key to Understand Its Invasion Success" Water 11, no. 6: 1247. https://doi.org/10.3390/w11061247
APA StyleSaç, G. (2019). Bio-Ecological Traits of Western Tubenose Goby Proterorhinus semilunaris (Heckel, 1837): A Key to Understand Its Invasion Success. Water, 11(6), 1247. https://doi.org/10.3390/w11061247