Spatially Explicit Modeling of Coupled Water and Carbon Processes Using a Distributed Ecohydrological Model in the Upper Heihe Watershed, China



 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
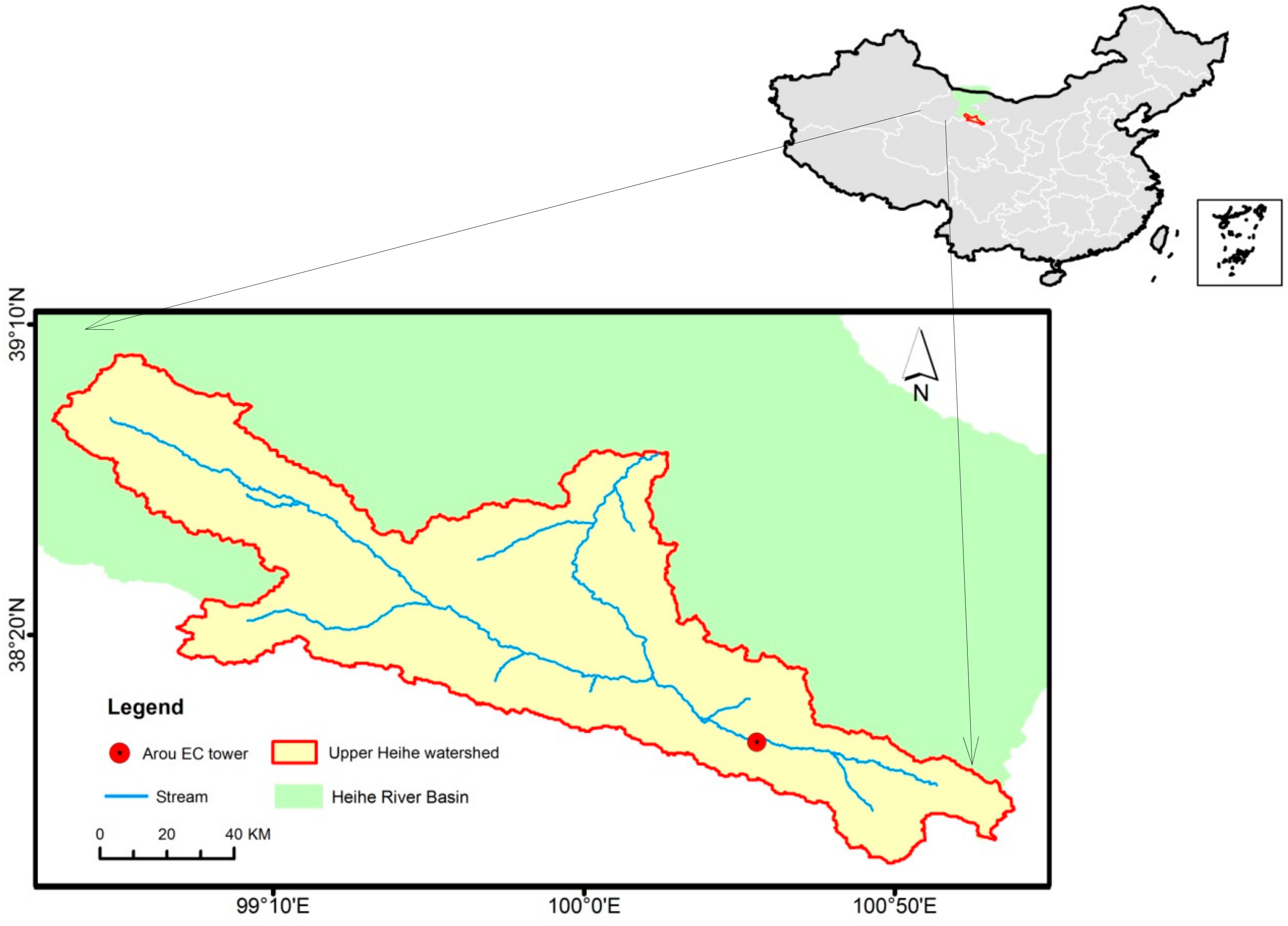
2.1. Study Area
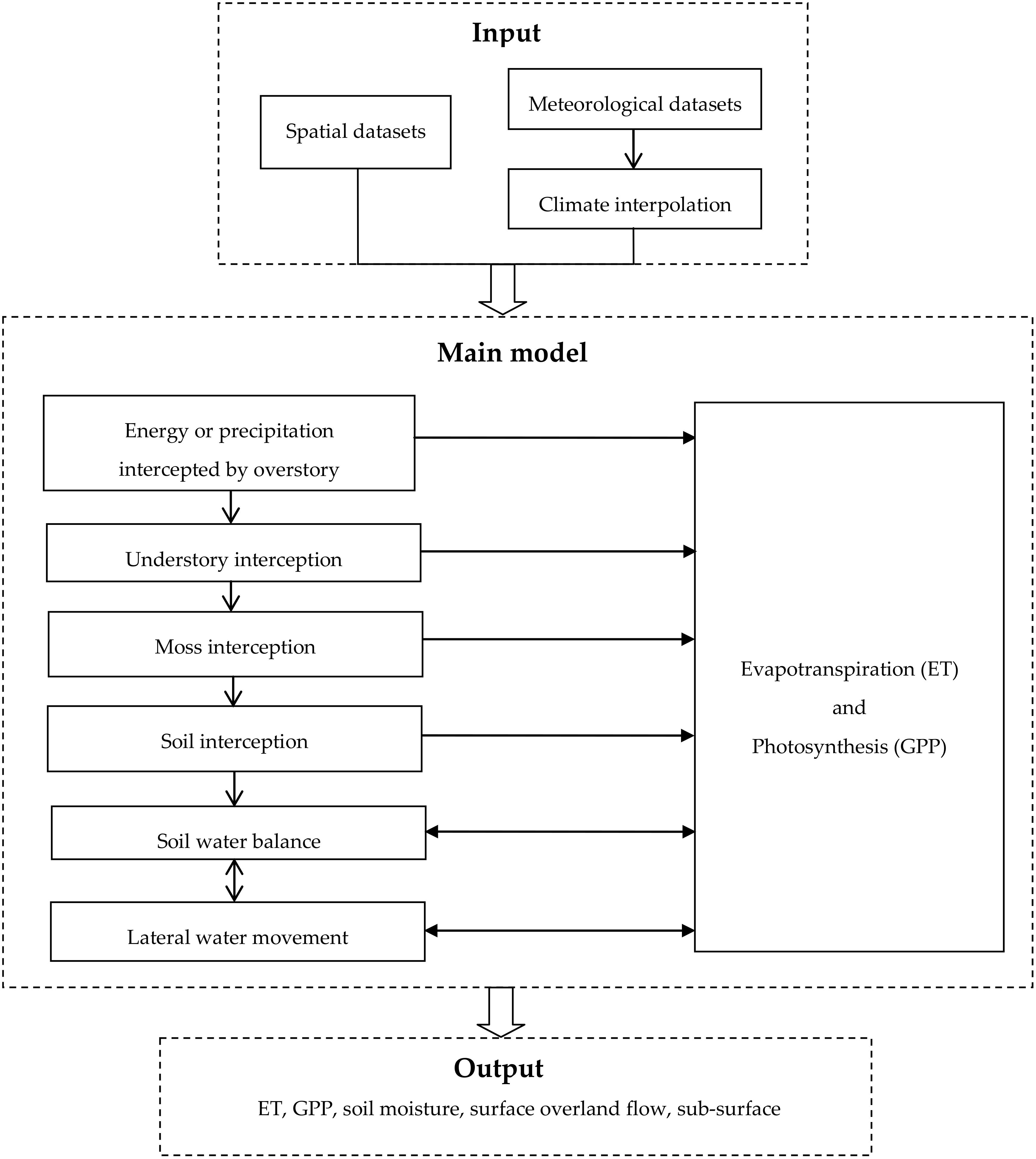
2.2. Model Discription
2.2.1. Ecophysiological Processes
2.2.2. Water Balance Calculation
2.3. Data Preparation
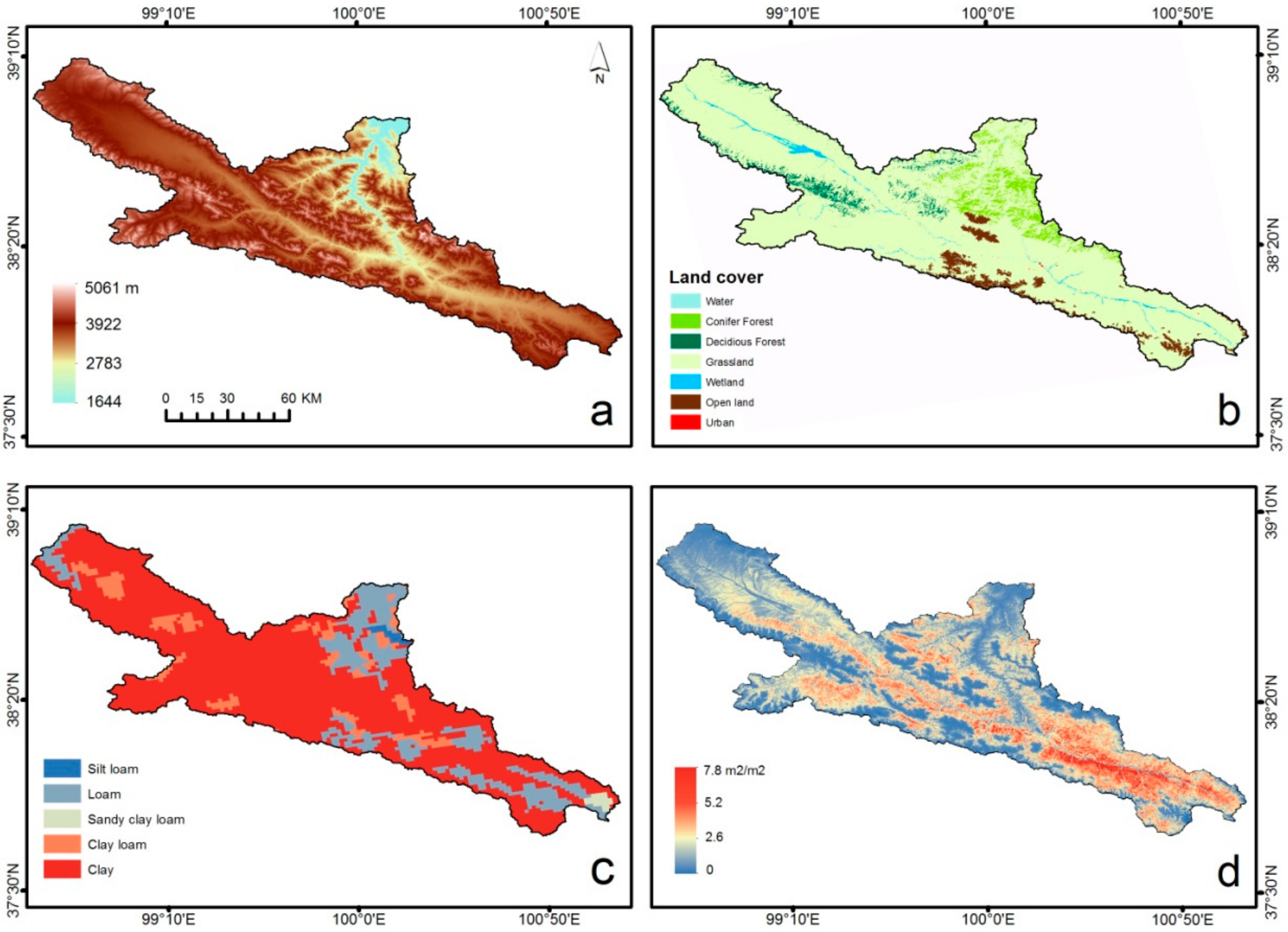
2.3.1. Spatial Datasets and Model Initialization
2.3.2. Meteorological Datasets and Flux Measurement
3. Results and Discussion
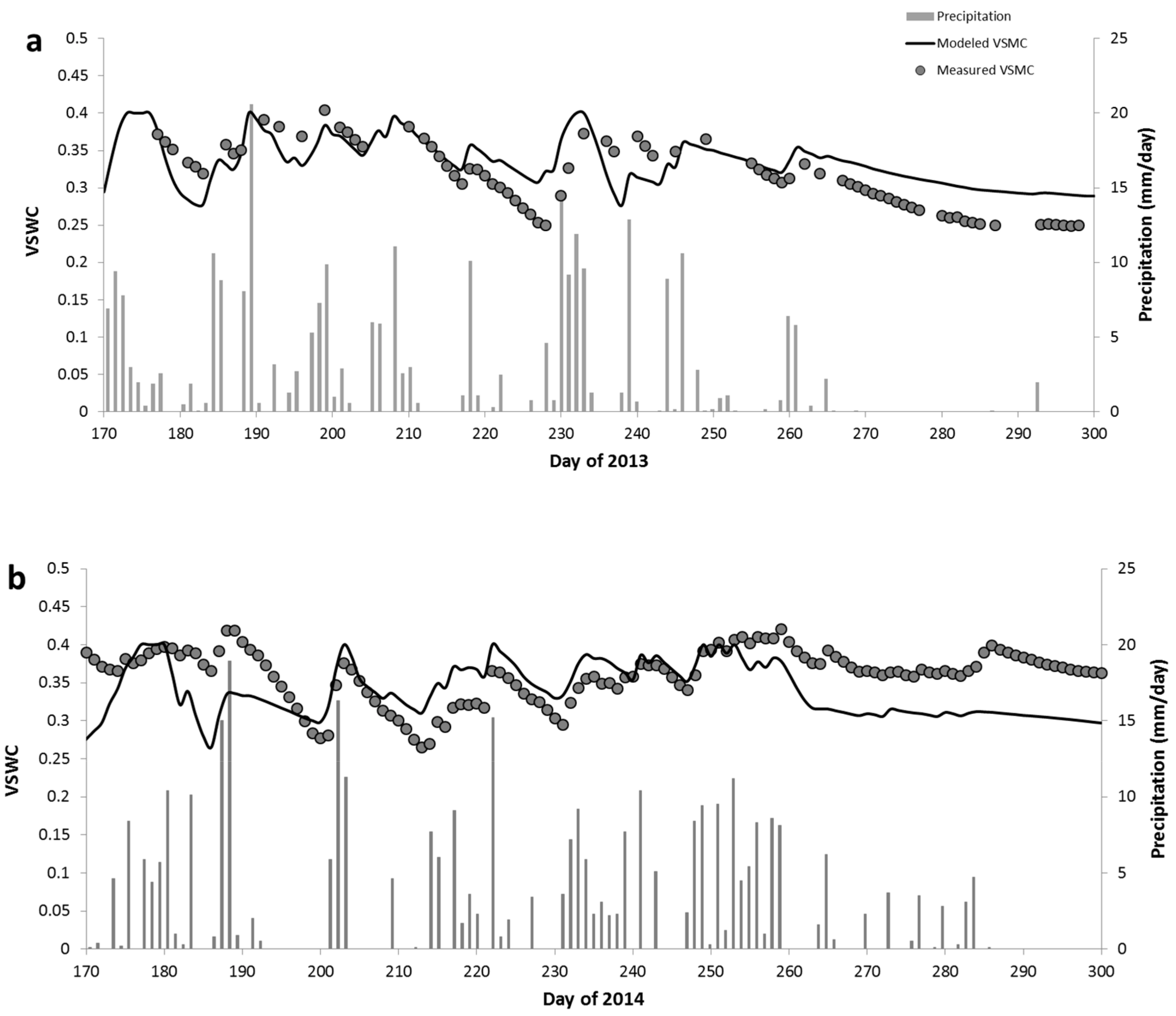
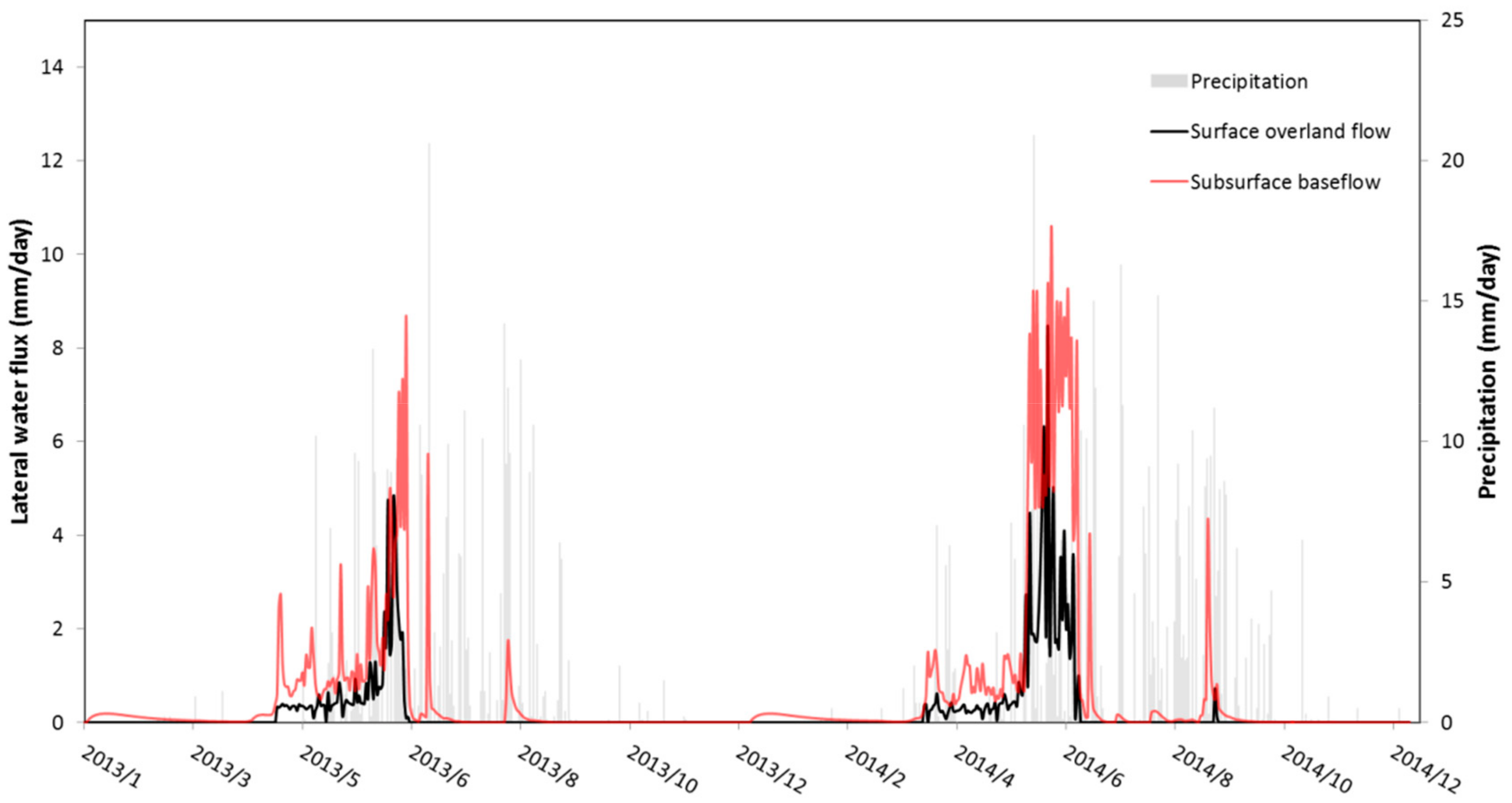
3.1. Temporal Patterns of Hydrological Indicators
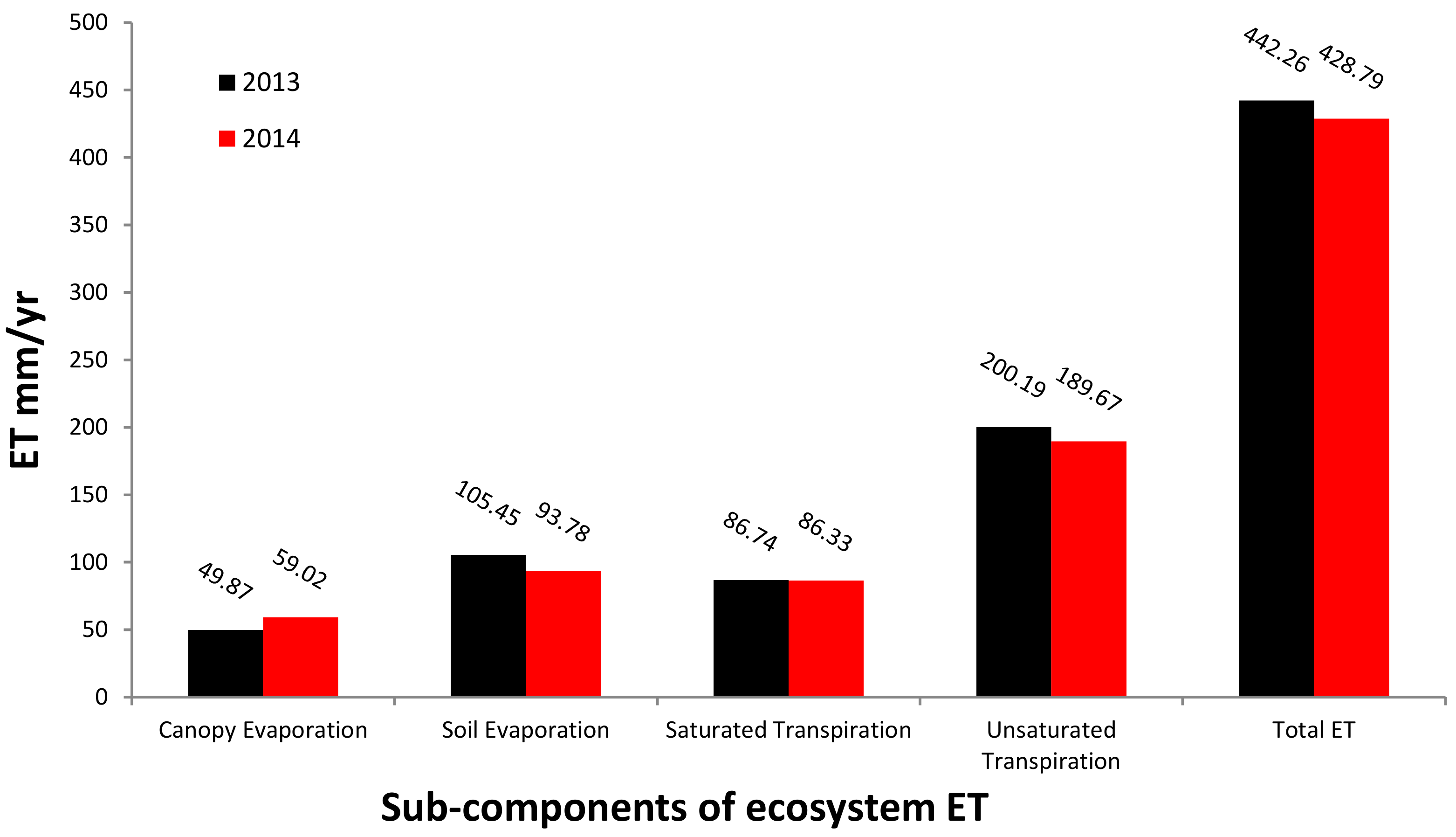
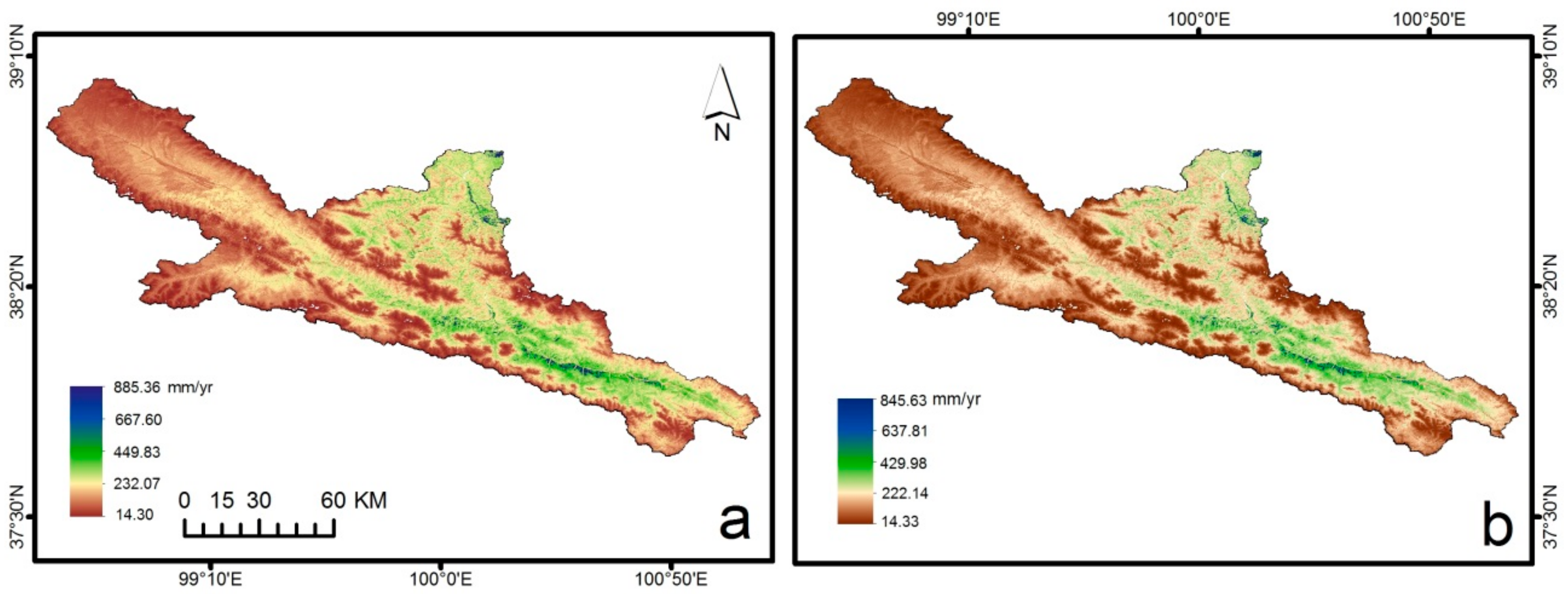
3.2. Spatial and Temporal Patterns of ET
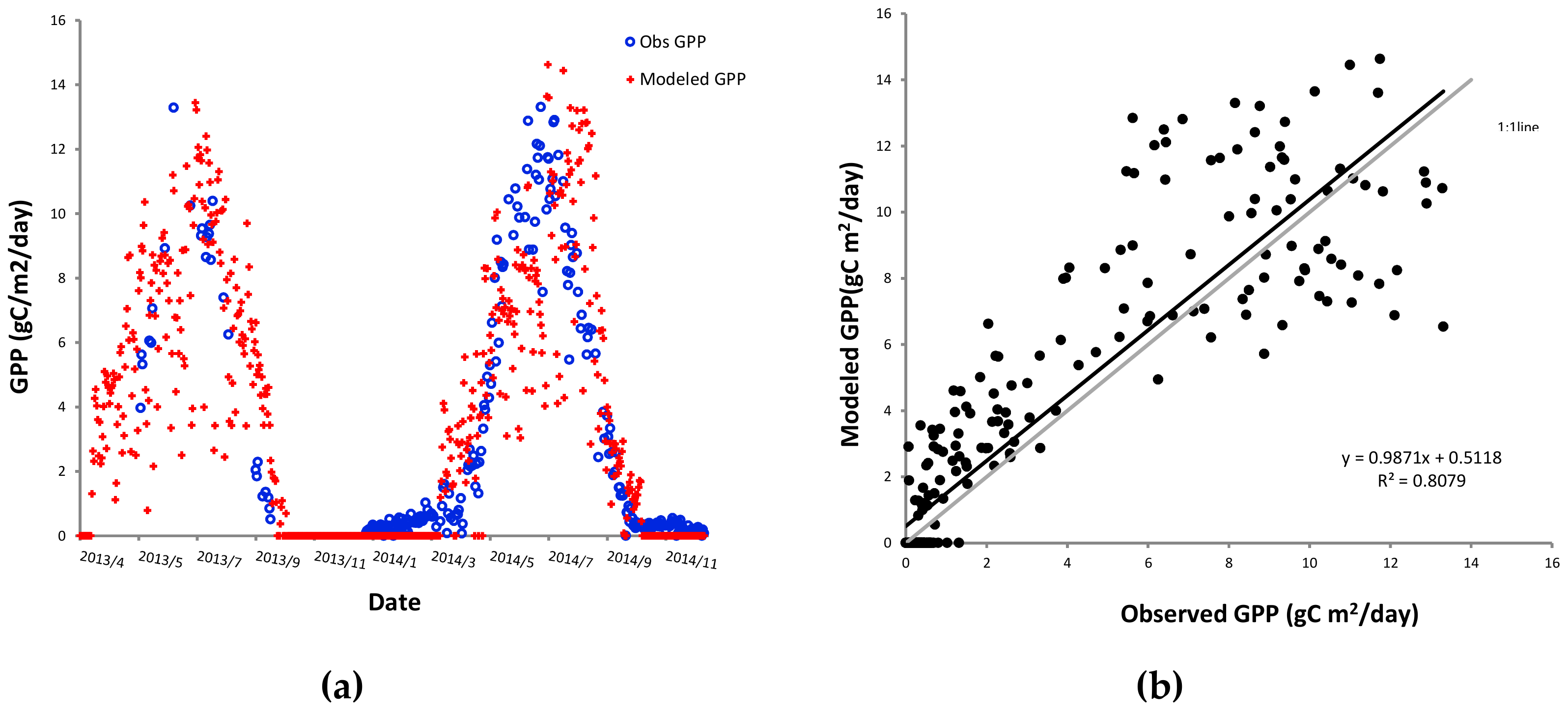
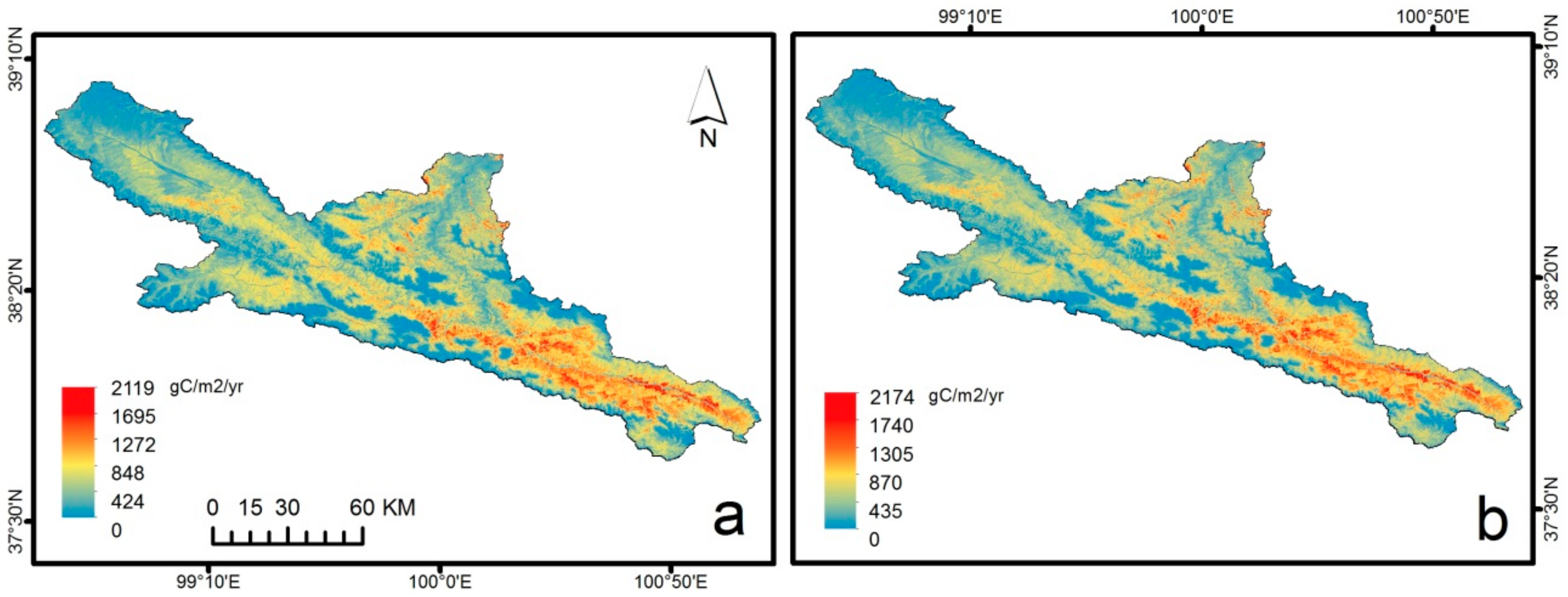
3.3. Spatial-Temporal Patterns of GPP
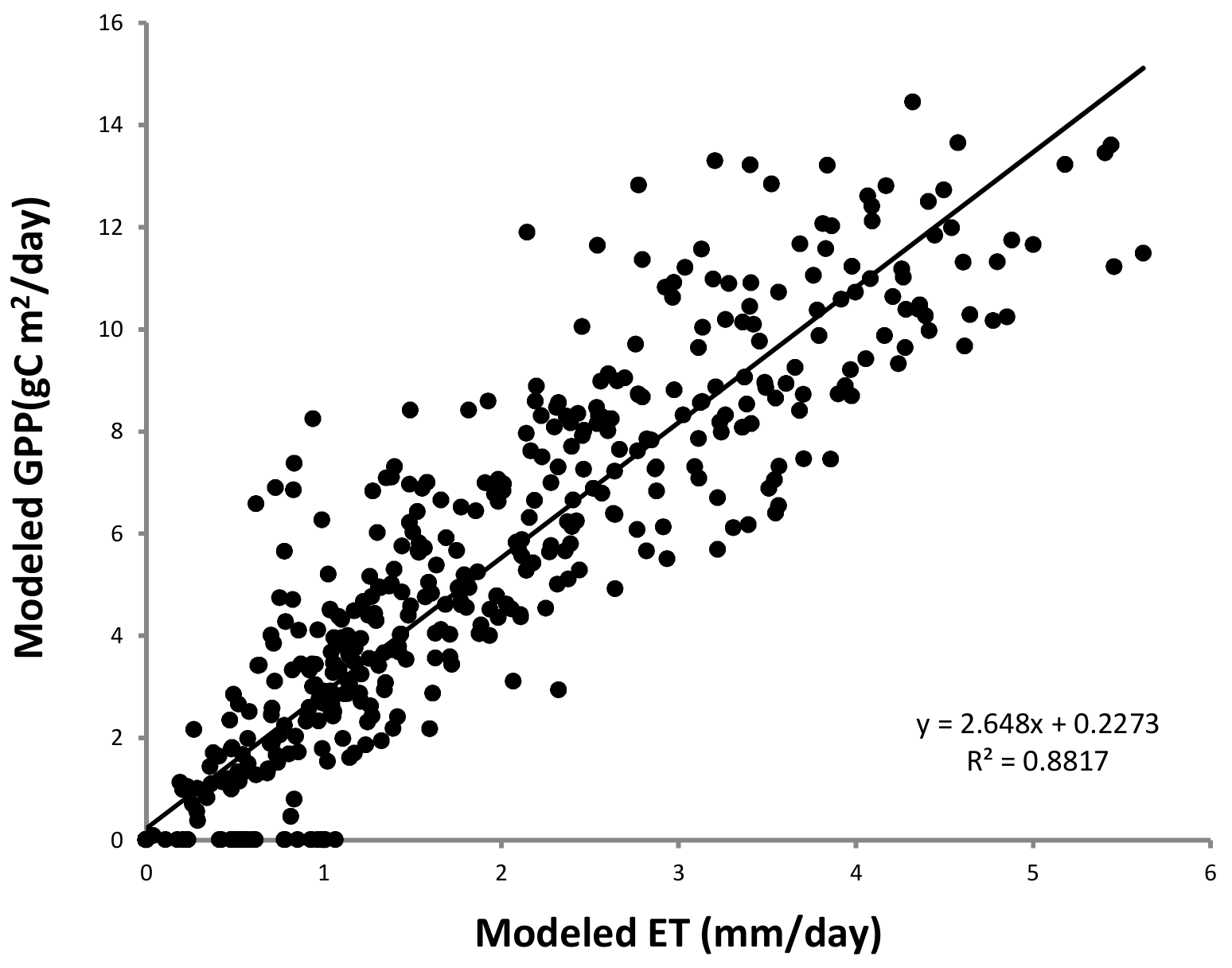
3.4. Co-Drive Mechanism of Ecosystem ET and GPP
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chen, B.; Chen, J.M.; Ju, W. Remote sensing-based ecosystem-atmosphere simulation scheme (EASS)—Model formulation and test with multiple-year data. Ecol. Model. 2007, 209, 277–300. [Google Scholar] [CrossRef]
- Govind, A.; Cowling, S.; Kumari, J.; Rajan, N.; Al-Yaari, A. Distributed modeling of ecohydrological processes at high spatial resolution over a landscape having patches of managed forest stands and crop fields in SW Europe. Ecol. Model. 2015, 297, 126–140. [Google Scholar] [CrossRef]
- Govind, A.; Kumari, J. Understanding the Terrestrial Carbon Cycle: An Ecohydrological Perspective. Int. J. Ecol. 2014, 2014, 712537. [Google Scholar] [CrossRef]
- Biederman, J.A.; Scott, R.L.; Goulden, M.L.; Vargas, R.; Litvak, M.E.; Kolb, T.E.; Yepez, E.A.; Oechel, W.C.; Blanken, P.D.; Bell, T.W.; et al. Terrestrial carbon balance in a drier world: The effects of water availability in southwestern North America. Glob. Chang. Biol. 2016, 22, 1867–1879. [Google Scholar] [CrossRef] [PubMed]
- Gea-Izquierdo, G.; Guibal, F.; Joffre, R.; Ourcival, J.M.; Simioni, G.; Guiot, J. Modelling the climatic drivers determining photosynthesis and carbon allocation in evergreen Mediterranean forests using multiproxy long time series. Biogeosciences 2015, 12, 3695–3712. [Google Scholar] [CrossRef]
- LU, L.; LI, X.; HUANG, C.; Frank, V. Analysis of the Spatio-Temporal Characteristics of Water Use Efficiency of Vegetation in West China. J. Glaciol. Geocryol. 2007, 29, 777–784. [Google Scholar]
- Wang, P.; Qiu, J.; Huo, Z.; Anderson, M.; Zhou, Y.; Bai, Y.; Liu, T.; Ren, S.; Feng, R.; Chen, P. Temporal Downscaling of Crop Coefficients for Winter Wheat in the North China Plain: A Case Study at the Gucheng Agro-Meteorological Experimental Station. Water 2017, 9, 155. [Google Scholar] [CrossRef]
- Richardson, A.D.; Anderson, R.S.; Arain, M.A.; Barr, A.G.; Bohrer, G.; Chen, G.; Chen, J.M.; Ciais, P.; Davis, K.J.; Desai, A.R.; et al. Terrestrial biosphere models need better representation of vegetation phenology: Results from the North American Carbon Program Site Synthesis. Glob. Chang. Biol. 2012, 18, 566–584. [Google Scholar] [CrossRef]
- Rodríguez, N.C.; Melgarejo, L.M.; Blair, M.W. Purple Passion Fruit, Passiflora edulis Sims f. edulis, Variability for Photosynthetic and Physiological Adaptation in Contrasting Environments. Agronomy 2019, 9, 231. [Google Scholar] [CrossRef]
- Gennaretti, F.; Gea-Izquierdo, G.; Boucher, E.; Berninger, F.; Arseneault, D.; Guiot, J. Ecophysiological modeling of photosynthesis and carbon allocation to the tree stem in the boreal forest. Biogeosciences 2017, 14, 4851–4866. [Google Scholar] [CrossRef]
- Nadal-Sala, D.; Keenan, T.F.; Sabaté, S.; Gracia, C. Forest Eco-Physiological Models: Water Use and Carbon Sequestration. In Managing Forest Ecosystems: The Challenge of Climate Change; Bravo, F., LeMay, V., Jandl, R., Eds.; Springer: Dordrecht, The Netherlands, 2017; pp. 81–102. [Google Scholar]
- Li, L.; Wang, Y.; Arora, V.K.; Eamus, D.; Shi, H.; Li, J.; Cheng, L.; Cleverly, J.; Hajima, T.; Ji, D.; et al. Evaluating Global Land Surface Models in CMIP5: Analysis of Ecosystem Water- and Light-Use Efficiencies and Rainfall Partitioning. J. Clim. 2018, 31, 2995–3008. [Google Scholar] [CrossRef]
- Baldocchi, D.D.; Wilson, K.B. Modeling CO2 and water vapor exchange of a temperate broadleaved forest across hourly to decadal time scales. Ecol. Model. 2001, 142, 155–184. [Google Scholar] [CrossRef]
- Emanuel, R.E.; Epstein, H.E.; McGlynn, B.L.; Welsch, D.L.; Muth, D.J.; D’Odorico, P. Spatial and temporal controls on watershed ecohydrology in the northern Rocky Mountains. Water Resour. Res. 2010, 46. [Google Scholar] [CrossRef]
- Govind, A.; Chen, J.M.; Margolis, H.; Ju, W.; Sonnentag, O.; Giasson, M. A spatially explicit hydro-ecological modeling framework (BEPS-TerrainLab V2.0): Model description and test in a boreal ecosystem in Eastern North America. J. Hydrol. 2009, 367, 200–216. [Google Scholar] [CrossRef]
- Sonnentag, O.; Talbot, J.; Chen, J.M.; Roulet, N.T. Using direct and indirect measurements of leaf area index to characterize the shrub canopy in an ombrotrophic peatland. Agric. For. Meteorol. 2007, 144, 200–212. [Google Scholar] [CrossRef]
- Zhengkun, Z.; Guangcan, Z.; Shunsheng, L.; Bin, P.; Zhiqiang, X.; Ping, X.; Xia, L. Effects of soil moisture on photosynthesis diurnal changes of Prunus sibirica L. Sci. Soil Water Conserv. 2012, 10, 99–104. [Google Scholar]
- Liu, Z.H.; McVicar, T.R.; Li, L.T.; Van Niel, T.G.; Yang, Q.K.; Li, R.; Mu, X.M. Modeling spatial distribution of pan evaporation based on quinat-variate thin plate spline function. Sci. Soil Water Conserv. 2006, 4, 23–30. [Google Scholar]
- Dang, S.Z.; Liu, C.M.; Wang, Z.G.; Wu, M.Y. Analyses on temporal variations of snowmelt runoff time in the upper reaches of Heihe River and its climate causes. J. Glaciol. Geocryol. 2012, 34, 920–926. [Google Scholar]
- Yan, M.; Li, Z.Y.; Tian, X.; Chen, E.X.; Gu, C.Y. Remote sensing estimation of gross primary productivity and its response to climate change in the upstream of Heihe River Basin. Chin. J. Plant Ecol. 2016, 40, 1–12. [Google Scholar]
- Shang, X.; Jiang, X.; Jia, R.; Wei, C. Land Use and Climate Change Effects on Surface Runoff Variations in the Upper Heihe River Basin. Water 2019, 11, 344. [Google Scholar] [CrossRef]
- Bi, L.M.; Hou, H.Y.; Yang, L.F. Analysis on change trend of runoff in Yingluoxia station of Heihe River. Yellow River 2013, 35, 23–25. [Google Scholar]
- Wang, R.; Cheng, Q.; Liu, L.; Yan, C.; Huang, G. Multi-Model Projections of Climate Change in Different RCP Scenarios in an Arid Inland Region, Northwest China. Water 2019, 11, 347. [Google Scholar] [CrossRef]
- Yang, Y.; Chen, R.; Han, C.; Qing, W. Measurement and estimation of the summertime daily evapotranspiration on alpine meadow in the Qilian Mountains, northwest China. Environ. Earth Sci. 2013, 68, 2253–2261. [Google Scholar] [CrossRef]
- Wang, Y. Remote Estimation of Carbon Fluxes and Carbon Sequestration Capacity over Cropland Based on Eddy Covariance Observation. Master’s Thesis, Henan Polytechnic University, Heinan Province, China, 2015. [Google Scholar]
- Sun, S.; Chen, B.; Ge, M.; Qu, J.; Che, T.; Zhang, H.; Lin, X.; Che, M.; Zhou, Z.; Guo, L.; et al. Improving soil organic carbon parameterization of land surface model for cold regions in the Northeastern Tibetan Plateau, China. Ecol. Model. 2016, 330, 1–15. [Google Scholar] [CrossRef]
- Wang, Y.H.; Yang, D.W.; Lei, H.M.; Yang, H. Impact of cryosphere hydrological processes on the river runoff in the upper reaches of Heihe River. J. Hydraul. Eng. 2015, 46, 1064–1071. [Google Scholar]
- Qin, J.; Ding, Y.; Wu, J.; Gao, M.; Yi, S.; Zhao, C.; Ye, B.; Li, M.; Wang, S. Understanding the impact of mountain landscapes on water balance in the upper Heihe River watershed in northwestern China. J. Arid Land 2013, 5, 366–383. [Google Scholar] [CrossRef]
- Lu, L.; Li, X.; Cheng, G.D. Landscape evolution in the middle Heihe River Basin of north-west China during the last decade. J. Arid Environ. 2003, 53, 395–408. [Google Scholar] [CrossRef]
- Chen, J.; Chen, B.; Black, T.A.; Innes, J.L.; Wang, G.; Kiely, G.; Hirano, T.; Wohlfahrt, G. Comparison of terrestrial evapotranspiration estimates using the mass transfer and Penman-Monteith equations in land surface models. J. Geophys. Res. Biogeosci. 2013, 118, 1715–1731. [Google Scholar] [CrossRef]
- Farquhar, G.D.; von Caemmerer, S.V.; Berry, J.A. A Biochemical Model of Photosynthetic CO2 Assimilation in Leaves of C3 Species. Planta 1980, 149, 78–90. [Google Scholar] [CrossRef]
- GALE, M.R.; GRIGAL, D.F. Vertical root distributions of northern tree species in relation to successional status. Can. J. For. Res. 1987, 17, 829–834. [Google Scholar] [CrossRef]
- Voinov, A.A.; Fitz, H.C.; Costanza, R. Surface water flow in landscape models: 1. Everglades case study. Ecol. Model. 1998, 108, 131–144. [Google Scholar] [CrossRef]
- Govind, A.; Chen, J.M.; McDonnell, J.; Kumari, J.; Sonnentag, O. Effects of lateral hydrological processes on photosynthesis and evapotranspiration in a boreal ecosystem. Ecohydrology 2011, 4, 394–410. [Google Scholar] [CrossRef]
- Oleson, K.W.; Lawrence, D.M.; Gordon, B.; Flanner, M.G.; Kluzek, E.; Peter, J.; Levis, S.; Swenson, S.C.; Thornton, E.; Feddema, J. Technical Description of Version 4.0 of the Community Land Model (CLM); NCAR Technical Note, NCAR/TN-478+STR; National Center for Atmospheric Research: Boulder, CO, USA, 2010. [Google Scholar]
- Li, X.; Cheng, G.; Liu, S.; Xiao, Q.; Ma, M.; Jin, R.; Che, T.; Liu, Q.; Wang, W.; Qi, Y.; et al. Heihe Watershed Allied Telemetry Experimental Research (HiWATER): Scientific Objectives and Experimental Design. Bull. Am. Meteorol. Soc. 2013, 94, 1145–1160. [Google Scholar] [CrossRef]
- Liu, S.M.; Xu, Z.W.; Wang, W.Z.; Jia, Z.Z.; Zhu, M.J.; Bai, J.; Wang, J.M. A comparison of eddy-covariance and large aperture scintillometer measurements with respect to the energy balance closure problem. Hydrol. Earth Syst. Sci. 2011, 15, 1291–1306. [Google Scholar] [CrossRef]
- Liu, S.; Li, X.; Xu, Z.; Che, T.; Xiao, Q.; Ma, M.; Liu, Q.; Jin, R.; Guo, J.; Wang, L.; et al. The Heihe Integrated Observatory Network: A Basin-Scale Land Surface Processes Observatory in China. Vadose Zone J. 2018, 17. [Google Scholar] [CrossRef]
- Lloyd, J.; Taylor, J.A. On the temperature dependence of soil respiration. Funct. Ecol. 1994, 8, 315–323. [Google Scholar] [CrossRef]
- Pan, X.; Li, X.; Cheng, G.; Chen, R.; Hsu, K. Impact Analysis of Climate Change on Snow over a Complex Mountainous Region Using Weather Research and Forecast Model (WRF) Simulation and Moderate Resolution Imaging Spectroradiometer Data (MODIS)-Terra Fractional Snow Cover Products. Remote Sens. 2017, 9, 774. [Google Scholar] [CrossRef]
- Molotch, N.P.; Blanken, P.D.; Williams, M.W.; Turnipseed, A.A.; Monson, R.K.; Margulis, S.A. Estimating sublimation of intercepted and sub-canopy snow using eddy covariance systems. Hydrol. Process. 2007, 21, 1567–1575. [Google Scholar] [CrossRef]
- Li, H.Y.; Wang, J. The snowmelt runoff model applied in the upper Heihe River Basin. J. Glaciol. Geocryol. 2008, 5, 769–775. [Google Scholar]
- Li, Q.; Chen, H.S. Variation of seasonal frozen soil in East China and their association with monsoon activity under the background of global warming. Clim. Chang. Res. Lett. 2013, 2, 47–53. [Google Scholar] [CrossRef]
- Schlesinger, W.H.; Jasechko, S. Transpiration in the global water cycle. Agric. For. Meteorol. 2014, 189, 115–117. [Google Scholar] [CrossRef]
- Wilson, K.B.; Baldocchi, D.D.; Hanson, P.J. Leaf age affects the seasonal pattern of photosynthetic capacityand net ecosystem exchange of carbon in a deciduous forest. Plant Cell Environ. 2001, 24, 571–583. [Google Scholar] [CrossRef]
- Garbulsky, M.F.; Penuelas, J.; Gamon, J.; Inoue, Y.; Filella, I. The photochemical reflectance index (PRI) and the remote sensing of leaf, canopy and ecosystem radiation use efficiencies: A review and meta-analysis. Remote Sens. Environ. 2011, 115, 281–297. [Google Scholar] [CrossRef]
- Savenije, H.H.G. HESS Opinions “Topography driven conceptual modelling (FLEX-Topo)”. Hydrol. Earth Syst. Sci. 2010, 14, 2681–2692. [Google Scholar] [CrossRef]
- Fischer, R.A.; Turner, N.C. Plant productivity in arid and semi-arid zones. Annu. Rev. Plant Physiol. 1978, 29, 277–317. [Google Scholar] [CrossRef]
- Chen, J.M.; Liu, J.; Cihlar, J.; Goulden, M.L. Daily canopy photosynthesis model through temporal and spatial scaling for remote sensing applications. Ecol. Model. 1999, 124, 99–119. [Google Scholar] [CrossRef]
- Jarvis, P.G. Interpretation of variations in leaf water potential and stomatal conductance found in canopies in field. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 1976, 273, 593–610. [Google Scholar] [CrossRef]
- Sonnentag, O.; Chen, J.M.; Roulet, N.T.; Ju, W.; Govind, A. Spatially explicit simulation of peatland hydrology and carbon dioxide exchange: Influence of mesoscale topography. J. Geophys. Res. Biogeosci. 2008, 113, G02005. [Google Scholar] [CrossRef]
- Tan, X.; Chang, S.X. Soil compaction and forest litter amendment affect carbon and net nitrogen mineralization in a boreal forest soil. Soil Tillage Res. 2007, 93, 77–86. [Google Scholar] [CrossRef]
- Traoré, S.; Thiombiano, L.; Millogo, J.R.; Guinko, S. Carbon and nitrogen enhancement in Cambisols and Vertisols by Acacia spp. in eastern Burkina Faso: Relation to soil respiration and microbial biomass. Appl. Soil Ecol. 2007, 35, 660–669. [Google Scholar]
- Govind, A.; Chen, J.M.; Ju, W. Spatially explicit simulation of hydrologically controlled carbon and nitrogen cycles and associated feedback mechanisms in a boreal ecosystem. J. Geophys. Res. Biogeosci. 2009, 114, G02006. [Google Scholar] [CrossRef]
- Arain, M.A.; Yuan, F.; Black, T.A. Soil–plant nitrogen cycling modulated carbon exchanges in a western temperate conifer forest in Canada. Agric. For. Meteorol. 2006, 140, 171–192. [Google Scholar] [CrossRef]
- Zhu, Z.; Sun, X.; Zhang, R.; Su, H.; Tang, X. Rapid measurements of CO2 flux density and water use efficiency of crop community. J. Appl. Ecol. 2004, 15, 1684–1686. [Google Scholar]
- Steduto, P.; Albrizio, R. Resource use efficiency of field-grown sunflower, sorghum, wheat and chickpea II. Water use efficiency and comparison with radiation use efficiency. Agric. For. Meteorol. 2005, 130, 269–281. [Google Scholar] [CrossRef]
- Yu, G.R.; Wang, Q.F.; Zhuang, J. Modeling the water use efficiency of soybean and maize plants under environmental stresses: Application of a synthetic model of photosynthesis-transpiration based on stomatal behavior. J. Plant Physiol. 2004, 161, 303–318. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter Names | Unit | Conifer | Deciduous | Grassland | Wetland |
|---|---|---|---|---|---|
| Canopy clumping index | - | 0.5 | 0.8 | 0.9 | 0.9 |
| Maximum stomatal conductance | m/s | 0.0016 | 0.005 | 0.0055 | 0.005 |
| Root decay rate | - | 0.94 | 0.96 | 0.95 | 0.93 |
| Maximum carboxylation rate at 25 °C | µmol/m2/s | 33 | 60 | 40 | 33 |
| Precipitation interception coefficient | mm/LAI/day | 0.2 | 0.3 | 0.25 | 0.1 |
| Max. leaf N content | % | 1.6 | 1.8 | 1.6 | 1.2 |
| Ration of gs,max of overstory to understory | - | 1.7 | 0.5 | 1.2 | 1 |
| Optimum temperature for photosynthesis | °C | 20 | 25 | 20 | 20 |
| Soil Hydraulic Parameter | Unit | Clay | Loam | Clay Loam | Sandy Loam |
|---|---|---|---|---|---|
| Field capacity | - | 0.4 | 0.27 | 0.3 | 0.23 |
| Wilting point | - | 0.27 | 0.12 | 0.2 | 0.1 |
| Porosity | % | 0.457 | 0.463 | 0.464 | 0.453 |
| Max. surface conductance | mm/s | 0.2 | 0.8 | 0.5 | 1 |
| Ksat,ver | m/day | 0.11 | 1.8 | 1.2 | 3.2 |
| Ksat,hor | m/day | 10 | 2.5 | 1.5 | 3.5 |
| Ks decay rate | m−1 | 0.02 | 0.04 | 0.029 | 0.042 |
| The exponent of the Brokes–Corey parameter | - | 11.4 | 5.39 | 5.5 | 4.9 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, H.; Chen, B.; Sun, S.; Zhang, H.; Measho, S.; Lin, X.; Guo, L. Spatially Explicit Modeling of Coupled Water and Carbon Processes Using a Distributed Ecohydrological Model in the Upper Heihe Watershed, China. Water 2019, 11, 1242. https://doi.org/10.3390/w11061242
Jin H, Chen B, Sun S, Zhang H, Measho S, Lin X, Guo L. Spatially Explicit Modeling of Coupled Water and Carbon Processes Using a Distributed Ecohydrological Model in the Upper Heihe Watershed, China. Water. 2019; 11(6):1242. https://doi.org/10.3390/w11061242
Chicago/Turabian StyleJin, Huiyu, Baozhang Chen, Shaobo Sun, Huifang Zhang, Simon Measho, Xiaofeng Lin, and Lifeng Guo. 2019. "Spatially Explicit Modeling of Coupled Water and Carbon Processes Using a Distributed Ecohydrological Model in the Upper Heihe Watershed, China" Water 11, no. 6: 1242. https://doi.org/10.3390/w11061242
APA StyleJin, H., Chen, B., Sun, S., Zhang, H., Measho, S., Lin, X., & Guo, L. (2019). Spatially Explicit Modeling of Coupled Water and Carbon Processes Using a Distributed Ecohydrological Model in the Upper Heihe Watershed, China. Water, 11(6), 1242. https://doi.org/10.3390/w11061242