River-Wide Habitat Availability for Fish Habitat Guilds: Implications for In-Stream Flow Protection

Abstract
1. Introduction
2. Materials and Methods
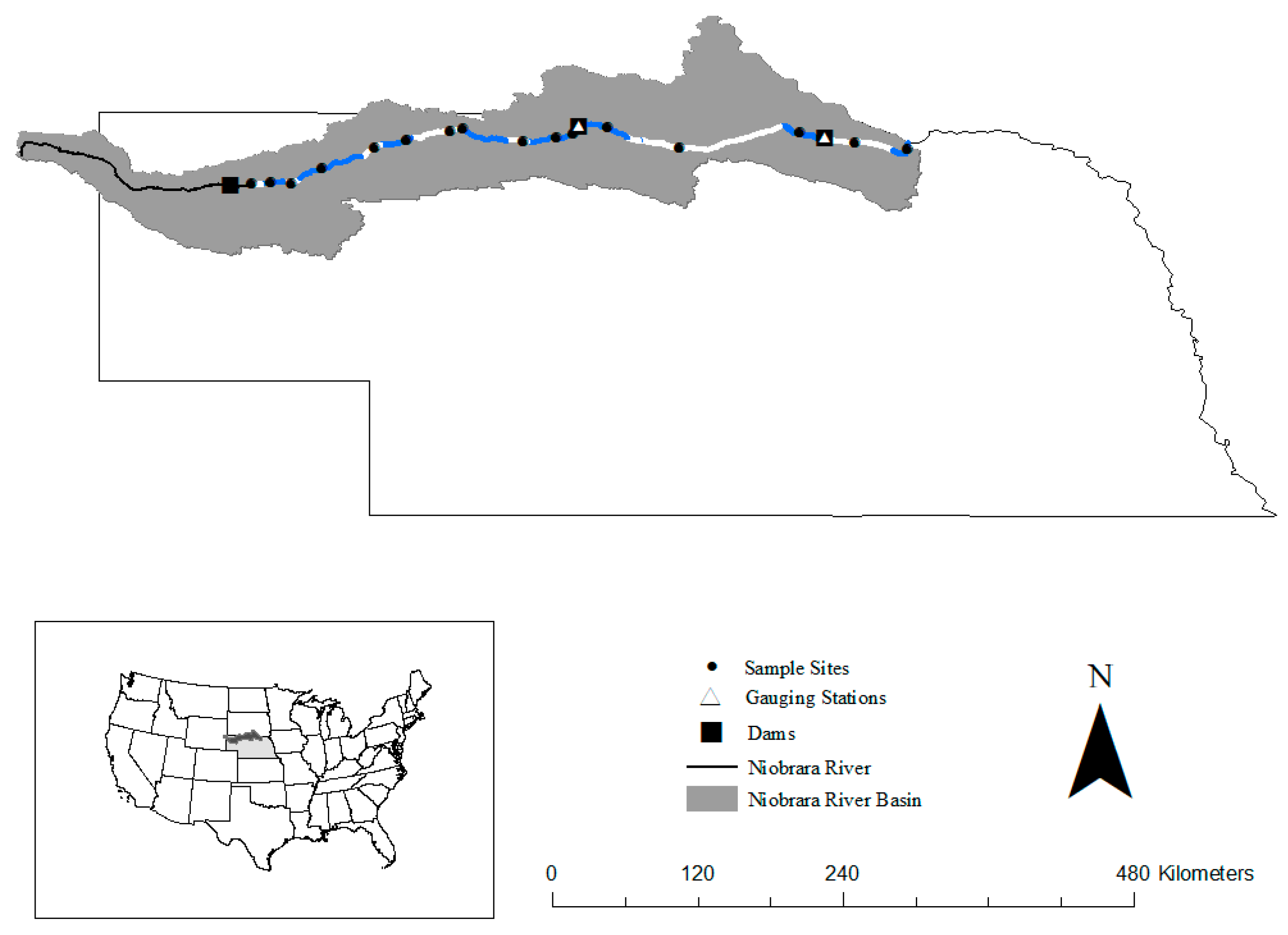
2.1. Niobrara River
2.2. Guild Specific Habitat Availability
2.2.1. Fish Species Surveys
2.2.2. Habitat Mapping Surveys
2.2.3. Development of Fish Habitat Guilds
2.3. Guild Specific Habitat Suitability
2.4. Time-Series Analysis for Variability in Discharge
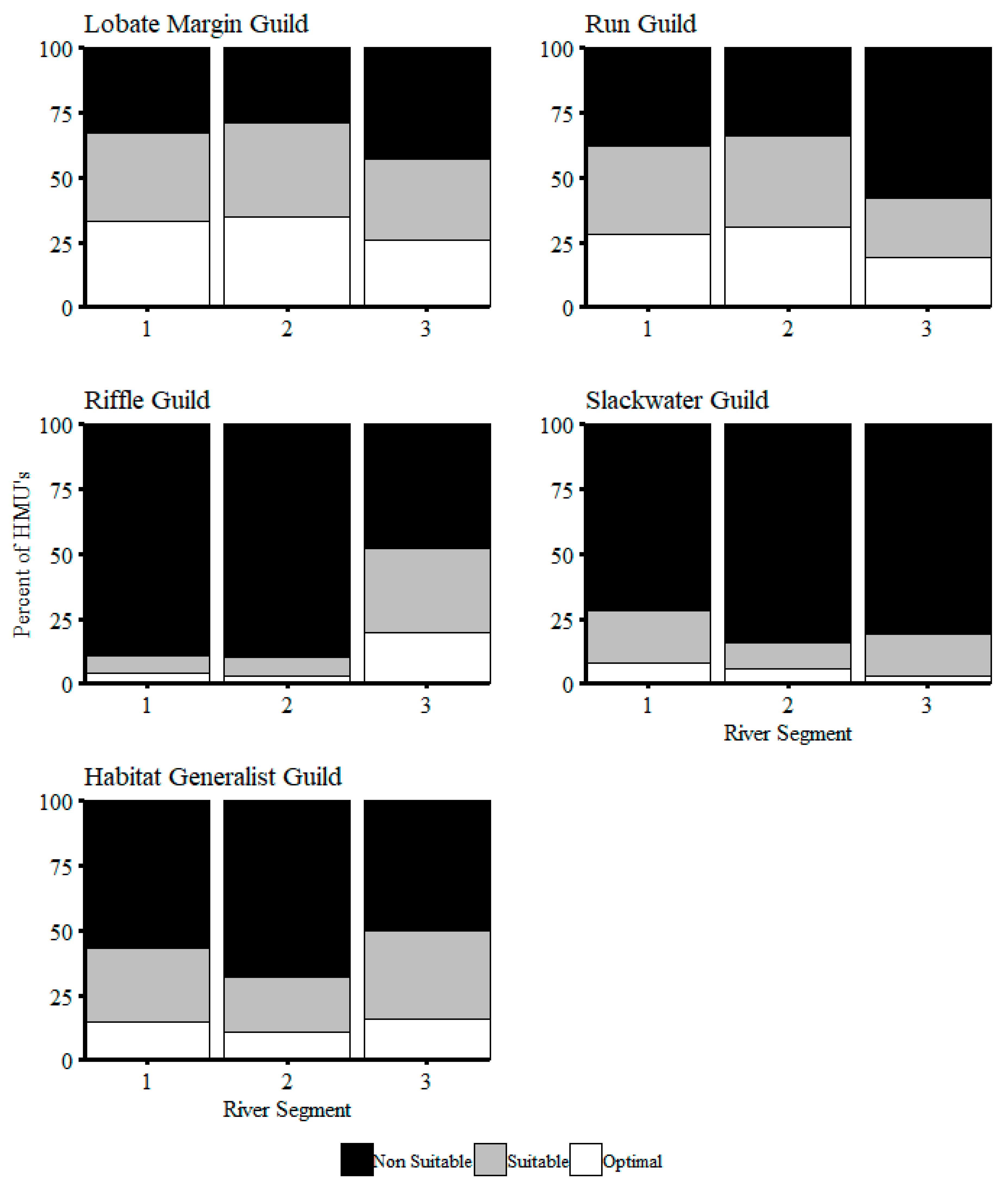
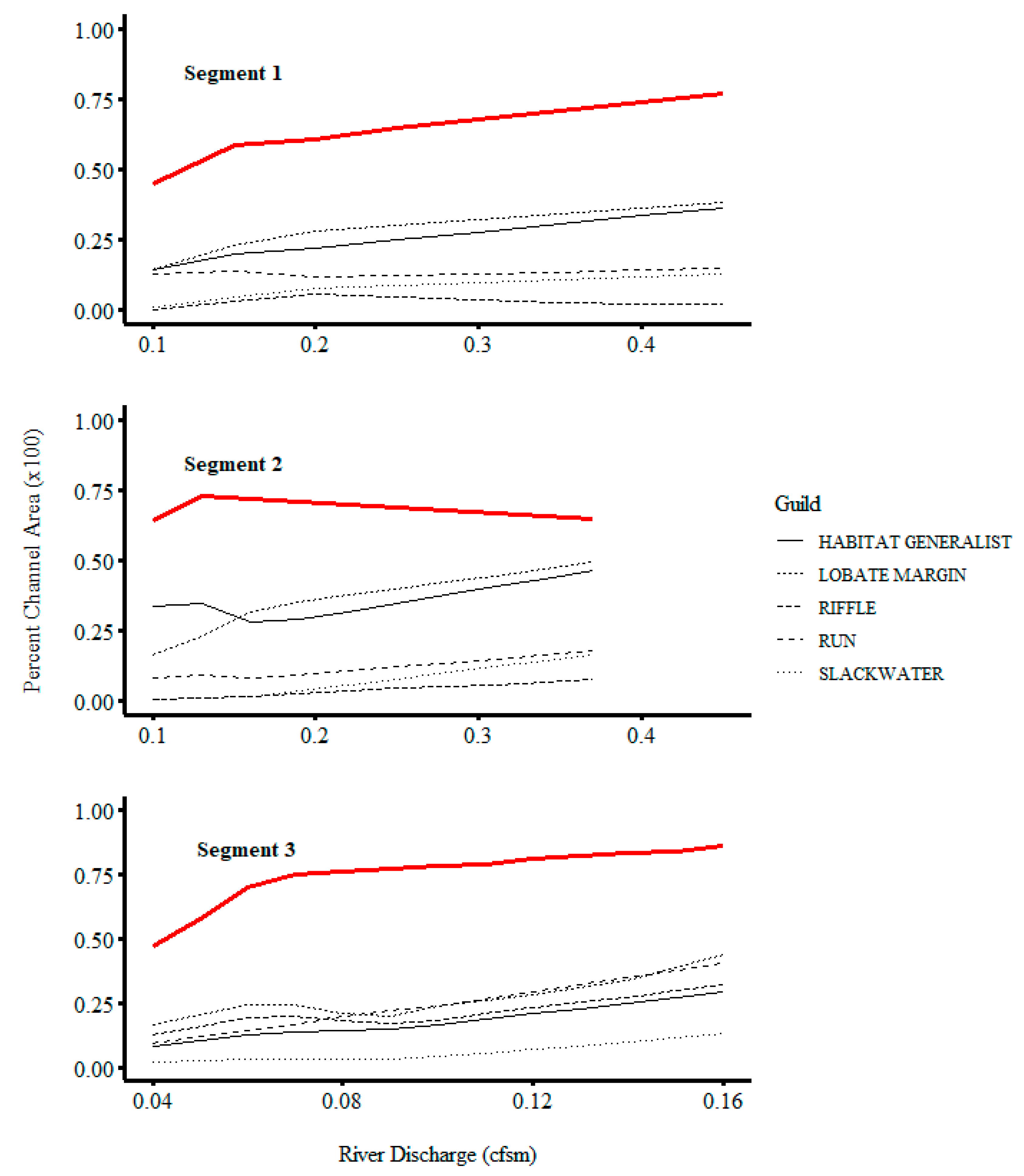
3. Results
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bunn, S.E.; Arthington, A.H. Basic principles and ecological consequences of altered flow regimes for aquatic biodiversity. Environ. Manag. 2002, 30, 492–507. [Google Scholar] [CrossRef]
- Humphries, P.; Neckeis, H.; Finlayson, B. The river wave concept: Integrating river ecosystem models. BioScience 2014, 64, 870–882. [Google Scholar] [CrossRef]
- Poff, N.L.; Allan, J.D.; Bain, M.B.; Karr, J.R.; Prestegaard, K.L.; Richter, B.D.; Sparks, R.E.; Stromberg, J.C. The natural flow regime: A new paradigm for riverine conservation and restoration. BioScience 1997, 47, 769–784. [Google Scholar] [CrossRef]
- Thoms, M.C.; Parsons, M. Identifying spatial and temporal patterns in the hydrological character of the Condamine-Balonne River, Australia, using multivariate statistics. River Res. Appl. 2003, 19, 443–457. [Google Scholar] [CrossRef]
- Jowett, I.G. In-stream flow methods: A comparison of approaches. River Res. Appl. 1996, 13, 115–127. [Google Scholar] [CrossRef]
- Lamouroux, N.; Capra, H.; Pouilly, C.; Souchon, Y. Fish habitat preferences in large streams of Southern France. Freshw. Biol. 1999, 42, 673–687. [Google Scholar] [CrossRef]
- Matthews, W.J.; Marsh-Matthews, E. Effects of drought on fish across axes of space, time and ecological complexity. Freshw. Biol. 2003, 48, 1232–1253. [Google Scholar] [CrossRef]
- Lamouroux, N.; Souchon, Y. Simple predictions of in-stream habitat model outputs for fish habitat guilds in large streams. Freshw. Biol. 2002, 47, 1531–1542. [Google Scholar] [CrossRef]
- Lamouroux, N.; Oliver, J. Testing predictions of changes in fish abundance and community structure after flow restoration in four reaches of a large river (French Rhone). Freshw. Biol. 2015, 60, 1118–1130. [Google Scholar] [CrossRef]
- Persinger, J.W.; Orth, D.J.; Averett, A.W. Using habitat guilds to develop habitat suitability criteria for a warmwater stream fish assemblage. River Res. Appl. 2011, 27, 956–966. [Google Scholar] [CrossRef]
- Aarts, B.G.W.; Nienhuis, P.H. Fish zonations and guilds as the basis for assessment of ecological integrity of large rivers. Hydrobiologia 2003, 500, 157–178. [Google Scholar] [CrossRef]
- Leonard, P.M.; Orth, D.J. Use of habitat guilds of fishes to determine in-stream flow requirements. N. Am. J. Fish. Manag. 1988, 8, 399–409. [Google Scholar] [CrossRef]
- Welcomme, R.L.; Winemiller, K.O.; Cowx, I.G. Fish environmental guilds as a tool for assessment of ecological condition of rivers. River Res. Appl. 2006, 22, 377–396. [Google Scholar] [CrossRef]
- Perkin, J.S.; Gido, K.B.; Falke, J.A.; Fausch, K.D.; Crockett, H.; Johnson, E.R.; Sanderson, J. Groundwater declines are linked to changes in Great Plains stream fish assemblages. Proc. Natl. Acad. Sci. USA 2017, 114, 7373–7378. [Google Scholar] [CrossRef]
- Parasiewicz, P. Developing a reference habitat template and ecological management scenarios using the MesoHABSIM model. River Res. Appl. 2007, 23, 924–932. [Google Scholar] [CrossRef]
- Vezza, P.; Parasiewicz, P.; Spatrani, M.; Comoglio, C. Habitat modeling in high-gradient streams: The mesoscale approach and application. Ecol. Appl. 2014, 24, 844–861. [Google Scholar] [CrossRef] [PubMed]
- Alexander, J.S.; Zelt, R.B.; Schaepe, N.J. Hydrogeomorphic and Hydraulic Habitats of the Niobrara River, Nebraska—with Special Emphasis on the Niobrara National Scenic River; U.S. Geological Survey Scientific Investigations Report: Denver, CO, USA, 2010.
- McManamay, R.A.; Bevelhimer, M.S.; Kao, S.C. Updating the US hydrologic classification: An approach to clustering and stratifying ecohydrologic data. Ecohydrology 2014, 7, 903–926. [Google Scholar] [CrossRef]
- Fischer, J.R.; Paukert, C.P. Habitat relationships with fish assemblages in minimally disturbed Great Plains regions. Ecol. Freshw. Fish. 2008, 17, 597–609. [Google Scholar] [CrossRef]
- Bain, M.B.; Finn, J.T.; Booke, H.E. A quantitative method for sampling riverine microhabitats by electrofishing. N. Am. J. Fish. Manag. 1985, 5, 489–493. [Google Scholar] [CrossRef]
- Austrian Standard ONORM 6232. Richtlinien fuer die oekologische Untersuchung und Bewertung von Fleissgewaessern; Oesterreichische Normungsinstitut: Vienna, Austria, 1995. [Google Scholar]
- Milligan, G.W.; Cooper, M.C. An examination of procedures for determining the number of clusters in a data set. Psychometrika 1985, 50, 159–179. [Google Scholar] [CrossRef]
- Pegg, M.A.; Behmer, A.T.; Parasaweicz, P.; Rogers, J.N. Application of mesohabitat fish use information to identify guilds for lotic systems. J. Appl. Ichthyol. 2014, 30, 1065–1068. [Google Scholar] [CrossRef]
- Pearce, J.; Ferrier, S. Evaluating the predictive performance of habitat models developed using logistic regression. Ecol. Model. 2000, 133, 225–245. [Google Scholar] [CrossRef]
- Capra, H.; Breil, P.; Souchon, Y. A new tool to interpret magnitude and duration of fish habitat variations. River Res. Appl. 1995, 10, 281–289. [Google Scholar] [CrossRef]
- Parasiewicz, P. Habitat time-series analysis to define flow-augmentation strategy for the Quinebaug River, Connecticut and Massachusetts, USA. River Res. Appl. 2008, 24, 439–452. [Google Scholar] [CrossRef]
- Foubert, A.; Lecomte, F.; Legendre, P.; Cusson, M. Spatial organisation of fish communities in the St. Lawrence River: A test for longitudinal gradients and spatial heterogeneities in a large river system. Hydrobiologia 2018, 809, 155–173. [Google Scholar] [CrossRef]
- Aadland, L.P. Stream habitat types: Their fish assemblages and relationship to flow. N. Am. J. Fish. Manag. 1993, 13, 790–806. [Google Scholar] [CrossRef]
- Spurgeon, J.J.; Pegg, M.A.; Parasiewicz, P.; Rogers, J. Diversity of river fishes influenced by habitat heterogeneity across hydrogeomophic divisions. River Res. Appl. 2018, 34, 797–806. [Google Scholar] [CrossRef]
- Schlosser, I.J.; Angermeier, P.L. Spatial variation in demographic processes of lotic fishes: Conceptual models, empirical evidence, and implications for conservation. Am. Fish. Soc. Symp. 1995, 17, 392–401. [Google Scholar]
- Jackson, D.A.; Peres-Neto, P.R.; Olden, J.D. What controls who is where in freshwater fish communities–the roles of biotic, abiotic, and spatial factors. Can. J. Fish. Aquat. Sci. 2001, 58, 157–179. [Google Scholar]
- Bowles, D.E.; Peitz, D.G.; Cribbs, J.T. Aquatic invertebrate community structure in the Niobrara River, Agate Fossil Beds National Monument, Nebraska, 1996–2009. Great Plains Res. 2013, 23, 1–10. [Google Scholar]
- Spurgeon, J.J.; Stasiak, R.H.; Cunningham, G.R.; Pope, K.L.; Pegg, M.A. Status of native stream fishes within selected protected areas of the Niobrara River in Western Nebraska. Great Plains Res. 2013, 24, 71–78. [Google Scholar] [CrossRef]
- Poff, N.L.; Richter, B.D.; Arthington, A.H.; Bunn, S.E.; Naiman, R.J.; Kendy, E.; Acreman, M.; Apse, C.; Bledsoe, B.P.; Freeman, M.C.; et al. The ecological limits of hydrologic alteration (ELOHA): A new framework for developing regional environmental flow standards. Freshw. Biol. 2010, 55, 147–170. [Google Scholar] [CrossRef]
- Poff, N.L.; Zimmerman, J.K.H. Ecological responses to altered flow regimes: A literature review to inform the science and management of environmental flows. Freshw. Biol. 2010, 55, 194–205. [Google Scholar] [CrossRef]
- Moyle, P.B.; Light, T. Fish Invasions in California: Do abiotic factors determine success? Ecology 1996, 77, 1666–1670. [Google Scholar] [CrossRef]
- Perkin, J.S.; Knorp, N.E.; Boersig, T.C.; Gebhard, A.E.; Hix, L.A.; Johnson, T.C. Life history theory predicts long-term fish assemblage response to stream impoundment. Can. J. Fish. Aquat. Sci. 2017, 74, 228–239. [Google Scholar] [CrossRef]
- Orth, D.J. Ecological considerations in the development and application of in-stream flow-habitat models. River Res. Appl. 1987, 1, 171–181. [Google Scholar] [CrossRef]
- Travnichek, V.H.; Bain, M.B.; Maceina, M.J. Recovery of warmwater fish assemblage after the initiation of a minimum-flow release downstream from a hydroelectric dam. Trans. Am. Fish. Soc. 1995, 124, 836–844. [Google Scholar] [CrossRef]
- Richter, B.; Baumgartner, J.; Wigington, D. How much water does a river need? Freshw. Biol. 1997, 37, 31–249. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Guild Name | Guild Members | Guild Description |
|---|---|---|
| Lobate Margin | Bigmouth shiner, Notropis dorsalis Red shiner, Cyprinella lutrensis Plains topminnow, Fundulus sciadicus Fathead minnow, Pimphales promelas Brook Stickleback, Culaea inconstans, Age 0 Channel catfish, Ictalurus punctatus | Inhabit areas of low velocity and shallow depths on the margin of channels. May include areas with lower dissolved oxygen and fluctuating temperatures. |
| Run | Sand shiner, Notropis stramineus Emerald shiner, Notropis atherinoides, | May include main channel areas with greater velocities and depths. |
| Riffle | Brassy minnow, Hybognathus hankinsoni, Central stoneroller, Campostoma anomalum Longnose dace, Rhinichthys cataractae, River shiner, Notropis blennius | Typically found in clearer water with lower velocity main channel habitats. Often areas include coarse substrate and aquatic vegetation. |
| Slackwater | Largemouth bass, Micropterus salmoides, Bluegill, Lepomis macrochirus, Green sunfish, Lepomis cyanellus, Yellow perch, Perca flavescens | Inhabits off-channel pools or backwaters near stream edges. Inhabited areas often lower in velocity and greater depths. |
| Habitat Generalist | White sucker, Catostomus commersonii, Creek chub, Semotilus atromaculatus, Shorthead redhorse, Moxostoma macrolepidotum, Age 0 River carpsucker, Carpiodes carpio | Inhabit a wide range of areas within river channel and backwaters. May include areas with lower dissolved oxygen and fluctuating temperatures. |
| Guild | Presence Model | Estimate | Abundance Model | ESTIMATE |
|---|---|---|---|---|
| Lobate Margin | Area under ROC | 77% | Area under ROC | 54% |
| Vegetation (overhanging) | −0.85 (0.32) | Velocity 75–90 cm s−1 | 0.51 (1.13) | |
| Woody Debris | 0.82 (0.43) | Velocity 90–105 cm s−1 | −0.96 (1.44) | |
| Shallow Margin | 0.79 (0.25) | |||
| Eroded | −0.56 (0.28) | |||
| Fast Run | 2.68 (1.51) | |||
| Run | 1.45 (0.46) | |||
| Depth >125 cm | 5.79 (5.10) | |||
| Velocity 15–30 cm s−1 | 4.35 (1.36) | |||
| Microlithal | −5.2 (2.21) | |||
| Run | Area under ROC | 68% | Area under ROC | 66% |
| Woody Debris | 0.88 (0.42) | Velocity 75–90 cm s−1 | 3.33 (1.78) | |
| Eroded | 0.52 (0.25) | Velocity 90–105 cm s−1 | −6.01 (2.68) | |
| Depth 75–100 cm | −2.9 (2.2) | Gigalithal | 6.01 (4.11) | |
| Depth >125 cm | −23.2 (13.10) | |||
| Velocity 15–30 cm s−1 | −1.77 (1.10) | |||
| Microlithal | −5.18 (1.90) | |||
| Riffle | Area under ROC | 81% | Area under ROC | 86% |
| Eroded | −0.63 (0.34) | Vegetation (submerged) | 4.72 (3.09) | |
| Stable | −0.43 (0.25) | Shading | −0.76 (0.85) | |
| Main Lobe | −1.21 (0.52) | Woody Debris | 2.74 (1.47) | |
| Depth 50–75 cm | 3.22 (1.05) | Stable | 1.03 (0.66) | |
| Velocity 90–105 cm s−1 | −4.22 (2.13) | Secondary Lobe | −4.21 (1.54) | |
| Akal | 6.91 (2.09) | Sidearm | −8.6 (2.89)) | |
| Mesolithal | 4.37 (1.46) | Velocity 30–45 cm s−1 | 15.03 (4.66) | |
| Slackwater | Area under ROC | 79% | Area under ROC | 74% |
| Depth 25–50 cm | 3.57 (1.32) | Shallow Margin | 1.64 (0.95) | |
| Depth 50–75 cm | 3.81 (1.18) | Run | −1.88 (1.11) | |
| Velocity 60–75 cm s−1 | −6.4 (1.90) | |||
| Akal | 2.27 (1.93) | |||
| Microlithal | −8.4 (3.37) | |||
| Habitat Generalist | Area under ROC | 78% | Area under ROC | 74% |
| Vegetation (submerged) | 1.68 (0.52) | Shallow Margins | −0.89 (0.38) | |
| Woody Debris | 0.74 (0.47) | Depth 50–75 cm s−1 | 2.69 (1.28) | |
| Eroded | 0.64 (0.28) | Velocity 60–75 cm s−1 | −2.42 (1.49) | |
| Stable | −0.55 (0.23) | Gigalithal | −6.08 (4.49) | |
| Glide | 2.24 (0.78) | |||
| Depth 50–75 cm | 2.97 (1.07) | |||
| Depth > 125 cm | −22.38 (13.51) | |||
| Velocity 15–30 cm s−1 | 3.46 (1.38) | |||
| Velocity 45–60 cm s−1 | −2.37 (1.10) | |||
| Macrolithal | 6.73 (3.36) |
| Segment | Guild | Community % | Habitat Availability (%CA) | ||
|---|---|---|---|---|---|
| 0.10 cfsm | 0.25 cfsm | 0.45 cfsm | |||
| 1 | Lobate Margin | 47 | 14 | 30 | 39 |
| Habitat Generalist | 33 | 14 | 25 | 36 | |
| Riffle | 2 | 0 | 5 | 2 | |
| Slackwater | 4 | 1 | 9 | 13 | |
| Run | 14 | 13 | 12 | 15 | |
| 0.10 cfsm | 0.22 cfsm | 0.37 cfsm | |||
| 2 | Lobate Margin | 22 | 16 | 38 | 49 |
| Habitat Generalist | 71 | 34 | 32 | 46 | |
| Riffle | 0 | 4 | 3 | 8 | |
| Slackwater | 1 | 1 | 6 | 17 | |
| Run | 6 | 8 | 11 | 18 | |
| 0.04 cfsm | 0.10 cfsm | 0.16 cfsm | |||
| 3 | Lobate Margin | 39 | 17 | 24 | 44 |
| Habitat Generalist | 12 | 8 | 17 | 29 | |
| Riffle | 23 | 13 | 19 | 32 | |
| Slackwater | 2 | 2 | 5 | 13 | |
| Run | 24 | 10 | 24 | 40 | |
| Segment | Discharge Condition | Bioperiod | ||||
|---|---|---|---|---|---|---|
| RG | OE | OL | ES | LS | ||
| 1 and 2 | Base (Common) discharge (cfsm) | 0.14 | 0.16 | 0.17 | 0.19 | 0.19 |
| Duration under persistent (days) | 32 | 45 | 21 | 18 | 20 | |
| Catastrophic duration (days) | 92 | 92 | 59 | 55 | 47 | |
| Trigger Discharge (cfsm) | 0.06 | 0.10 | 0.08 | 0.12 | 0.10 | |
| Duration under persistent (days) | 8 | 9 | 7 | 7 | 7 | |
| Catastrophic duration (days) | 16 | 18 | 8 | 11 | 11 | |
| Subsistence (Rare) Discharge (cfsm) | 0.05 | 0.08 | 0.06 | 0.11 | 0.09 | |
| Duration under persistent (days) | 5 | 6 | 4 | 3 | 6 | |
| Catastrophic duration (days) | 8 | 10 | 5 | 8 | 9 | |
| 3 | Base (Common) Discharge (cfsm) | 0.09 | 0.12 | 0.13 | 0.10 | 0.10 |
| Duration under persistent (days) | 47 | 46 | 22 | 39 | 12 | |
| Catastrophic duration (days) | 92 | 92 | 59 | 54 | 29 | |
| Trigger Discharge (cfsm) | 0.06 | 0.07 | 0.08 | 0.07 | 0.07 | |
| Duration under persistent (days) | 11 | 8 | 7 | 6 | 4 | |
| Catastrophic duration (days) | 30 | 25 | 12 | 9 | 6 | |
| Subsistence (Rare) Discharge (cfsm) | 0.06 | 0.06 | 0.07 | 0.07 | 0.07 | |
| Duration under persistent (days) | 10 | 4 | 4 | 3 | 3 | |
| Catastrophic duration (days) | 21 | 6 | 7 | 6 | 5 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Spurgeon, J.; Pegg, M.; Parasiewicz, P.; Rogers, J. River-Wide Habitat Availability for Fish Habitat Guilds: Implications for In-Stream Flow Protection. Water 2019, 11, 1132. https://doi.org/10.3390/w11061132
Spurgeon J, Pegg M, Parasiewicz P, Rogers J. River-Wide Habitat Availability for Fish Habitat Guilds: Implications for In-Stream Flow Protection. Water. 2019; 11(6):1132. https://doi.org/10.3390/w11061132
Chicago/Turabian StyleSpurgeon, Jonathan, Mark Pegg, Piotr Parasiewicz, and Joe Rogers. 2019. "River-Wide Habitat Availability for Fish Habitat Guilds: Implications for In-Stream Flow Protection" Water 11, no. 6: 1132. https://doi.org/10.3390/w11061132
APA StyleSpurgeon, J., Pegg, M., Parasiewicz, P., & Rogers, J. (2019). River-Wide Habitat Availability for Fish Habitat Guilds: Implications for In-Stream Flow Protection. Water, 11(6), 1132. https://doi.org/10.3390/w11061132