Composition and Dynamics of Phytoplankton in the Coastal Bays of Maryland, USA, Revealed by Microscopic Counts and Diagnostic Pigments Analyses

Abstract
1. Introduction
2. Materials and Methods
2.1. Study Location and Environmental Parameters
Sample Collection
2.2. Sample Analyses
2.2.1. Nutrient Analysis
2.2.2. HPLC Pigment Analysis
2.2.3. Phytoplankton Microscopic Identification
2.2.4. Freshwater Flow
2.2.5. Statistical Analyses
3. Results
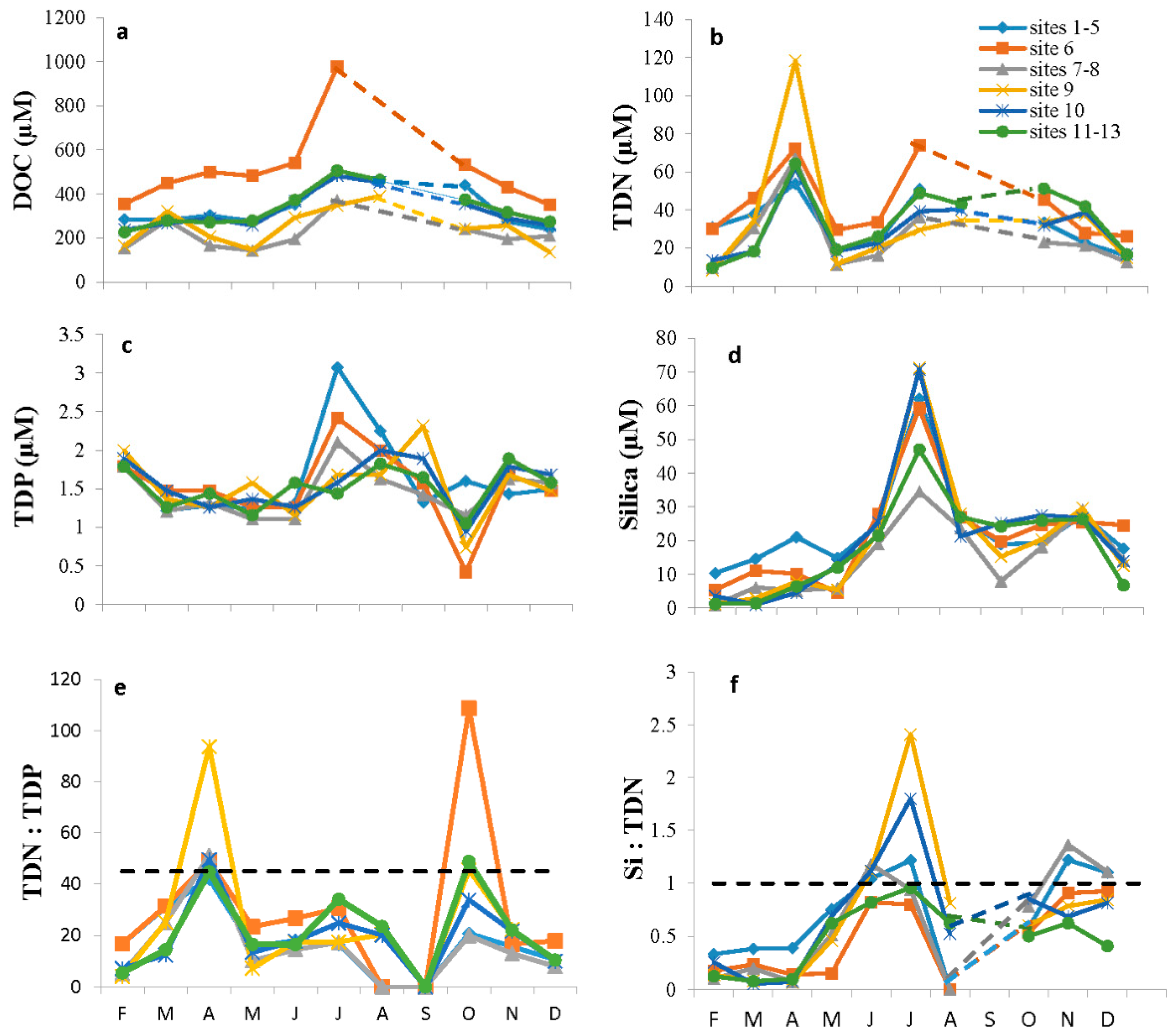
3.1. The Physical Environment
The Chemical Environment
3.2. Phytoplankton Species Composition
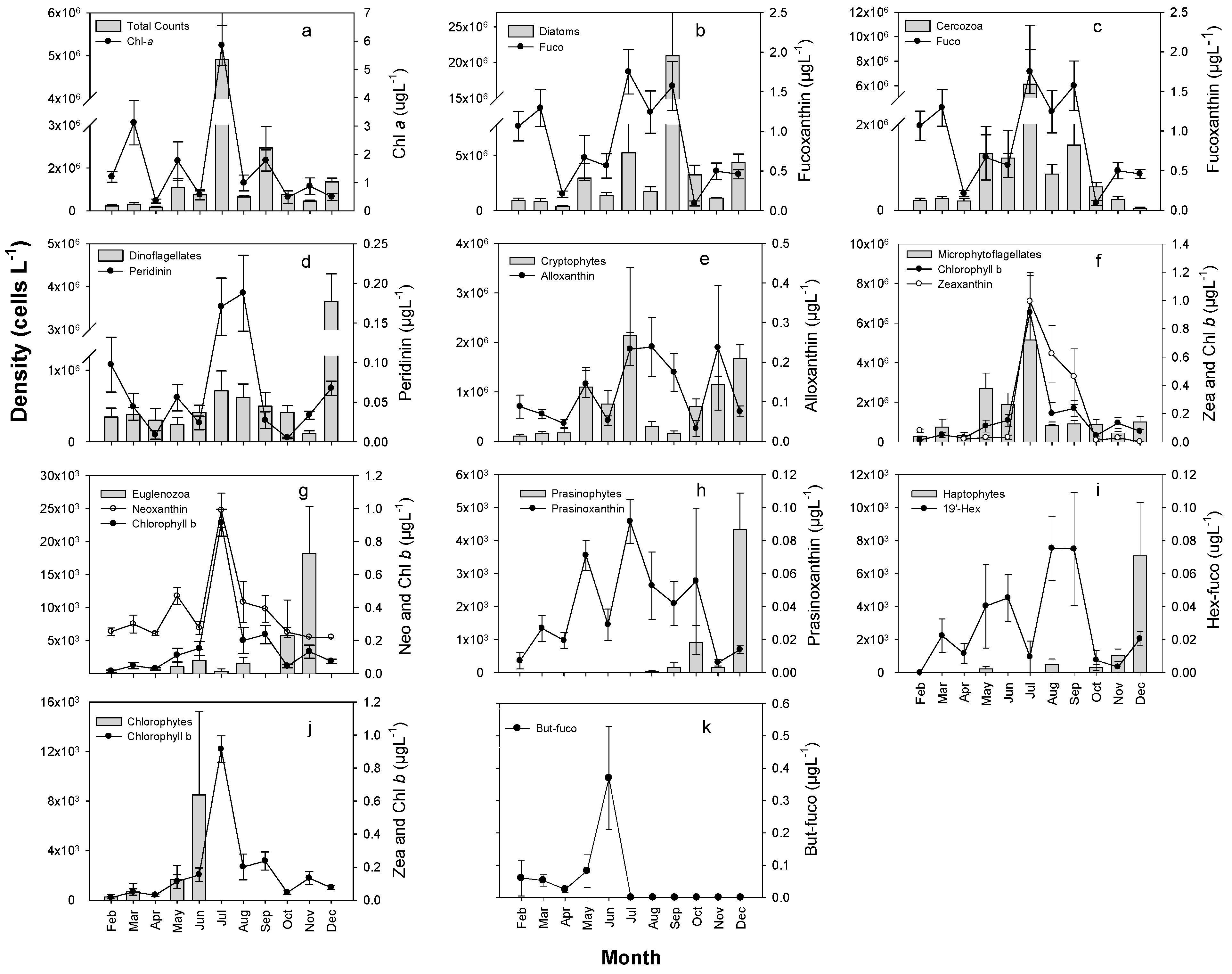
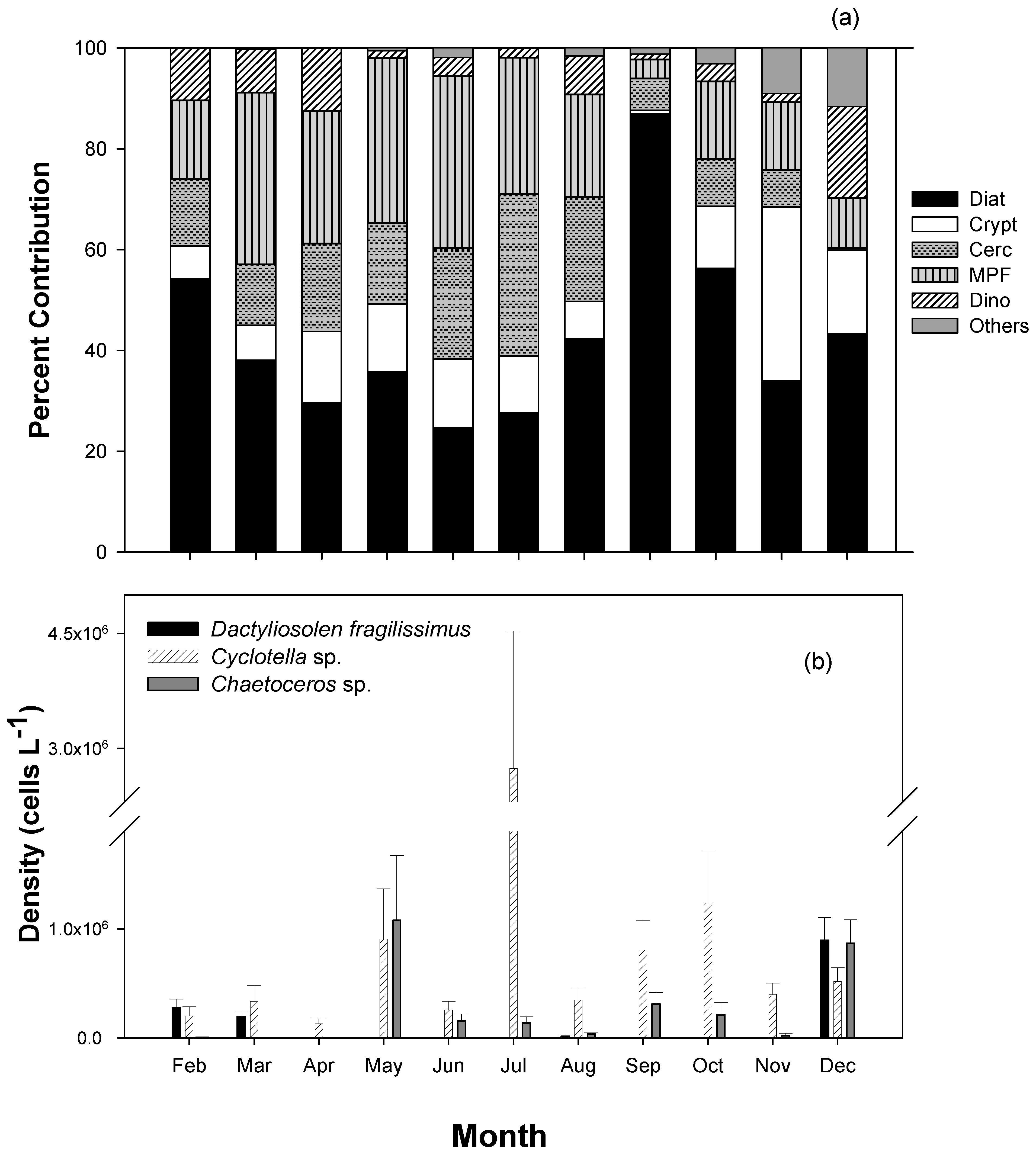
3.3. Seasonal Composition and Variation of Major Phytoplankton Taxonomic Groups Based on Photosynthetic Pigments and Microscopic Counts
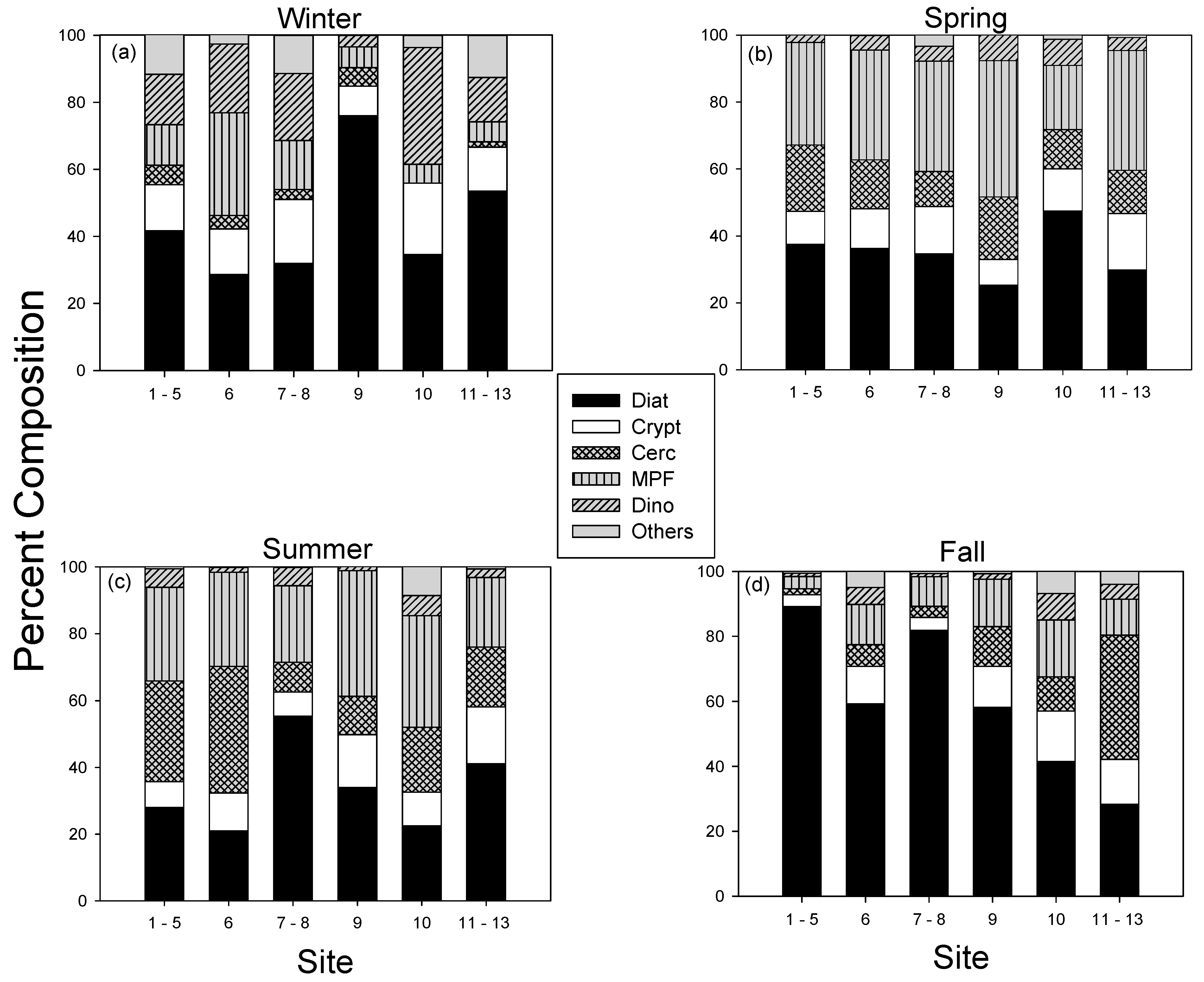
3.4. Spatial Variations in the Abundance of Major Taxa and Dominant Species in the MCBs
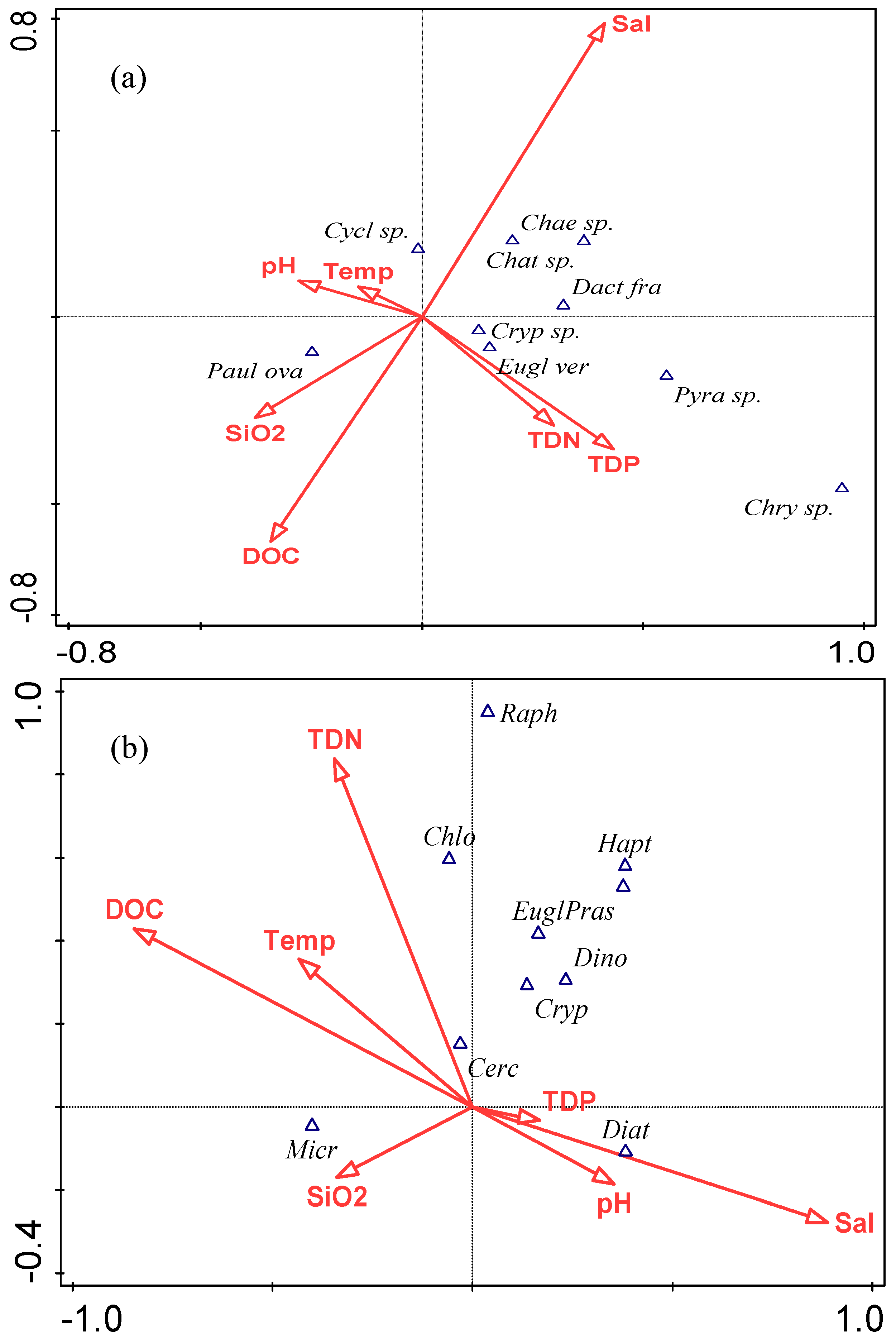
3.5. Relationships between Phytoplankton Assemblage based on Microscopic Counts and Physico-Chemical Variables
4. Discussion
4.1. Phytoplankton Taxa Composition and Variation
4.2. Seasonal Patterns of Succession of Major Phytoplankton Taxa
4.3. Spatial Variation of Phytoplankton Abundance
4.4. Dominance of Diatoms and Phosphorus Limitation
5. Conclusions
- Diatoms were the most abundant taxonomic group and showed strong correlations with its diagnostic pigment, fucoxanthin. There were also significant contributions from the nanophytoplankton especially in summer when they dominated the community, attributed largely to changes in nutrient composition and concentrations.
- Similar to other nutrient enriched estuaries along the US east coast, the MCBs displayed a distinct seasonal composition and variability in phytoplankton. Diatoms dominated the winter community while the relative abundance of the MPF and cercozoans increased, with diatoms and cryptophytes/cercozoans co-dominating in fall.
- Spatial variations in phytoplankton abundance and composition were likely due to environmental factors like watershed characteristics, nutrient levels and salinity. Sites receiving freshwater inputs directly from tributaries displayed high phytoplankton biomass.
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Silva, A.; Mendes, C.R.; Palma, S.; Brotas, V. Short-time scale variation of phytoplankton succession in Lisbon bay (Portugal) as revealed by microscopy cell counts and HPLC pigment analysis. Estuar. Coast. Shelf Sci. 2008, 79, 230–238. [Google Scholar]
- Garmendia, M.; Borja, A.; Franco, J.; Revilla, M. Phytoplankton composition indicators for the assessment of eutrophication in marine waters: Present state and challenges within the European directives. Mar. Pollut. Bull. 2013, 66, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Marshall, H.G. Phytoplankton of the York River. J. Coast. Res. 2009, 57, 59–65. [Google Scholar] [CrossRef]
- Glibert, P.M.; Wazniak, C.; Hall, M.; Sturgis, B. Seasonal and interannual trends in nitrogen in Maryland’s Coastal Bays and relationships with brown tide. Ecol. Appl. 2007, 17, S79–S87. [Google Scholar] [CrossRef]
- Gobler, C.J.; Sunda, W.G. Ecosystem disruptive algal blooms of the brown tide species Aureococcus anophagefferens and Aureoumbra lagunensis. Harmful Algae 2012, 14, 36–45. [Google Scholar] [CrossRef]
- Wilhelm, C.; Rudolf, I.; Renner, W. A quantitative method based on HPLC–aided pigment analysis to monitor structure and dynamics of the phytoplankton assemblage—A study from Lake Meerelder Mar (Eifel, Germany). Arch. Fur Hydrobiol. 1991, 123, 21–35. [Google Scholar]
- Domingues, R.B.; Barbosas, A.; Galvao, H. Constraints on the use of phytoplankton as a biological quality element within the Water Framework Directive in Portuguese waters. Mar. Pollut. Bull. 2008, 59, 1389–1395. [Google Scholar] [CrossRef] [PubMed]
- Harvey, H.W. Measurement of Phytoplankton Population. J Mar. Biol. Assoc. UK 1934, 19, 761–773. [Google Scholar] [CrossRef]
- Wright, S.W.; Jeffrey, S.W.; Mantoura, R.F.C.; Llewellyn, C.A.; Bjornland, T.; Repeta, D.; Welschmeyer, N. Improved HPLC method for the analysis of chlorophylls and carotenoids from marine phytoplankton. Mar. Ecol. Prog. Ser. 1991, 77, 183–196. [Google Scholar]
- Jeffrey, S.W.; Vesk, M. Introduction to marine phytoplankton and their pigment signatures. In Phytoplankton Pigments in Oceanography; Jeffrey, S.W., Mantoura, R.F.C., Wright, S.W., Eds.; UNESCO Publishing: Paris, France, 1997; pp. 37–84. [Google Scholar]
- Barlow, R.G.; Sessions, H.; Balarin, M.; Weeks, S.; Whittle, C.; Hutchings, L. Seasonal variation in phytoplankton in the southern Benguela: Pigment indices and ocean colour. Afr. J. Mar. Sci. 2005, 27, 275–288. [Google Scholar]
- Roy, R.; Pratihary, A.; Mangesh, G.; Naqvi, S.W.A. Spatial variation of phytoplankton pigments along the southwest coast of India. Estuar. Coast. Shelf Sci. 2006, 69, 189–195. [Google Scholar]
- Seoane, S.; Garmendia, M.; Revilla, M.; Borja, A.; Franco, J.; Orive, E.; Valencia, V. Phytoplankton pigments and epifluorescence microscopy as tools for ecological status assessment in coastal and estuarine waters, within the Water framework Directive. Mar. Pollut. Bull. 2011, 62, 1484–1497. [Google Scholar] [PubMed]
- Gieskes, W.W.C.; Kraay, G.W. Dominance of Cryptophyceae during the phytoplankton spring bloom in the central North Sea detected by HPLC analysis of pigments. Mar. Biol. 1983, 75, 179–185. [Google Scholar] [CrossRef]
- Wright, S.W.; Thomas, D.P.; Marchant, H.J.; Higgins, H.W.; Mackey, M.D.; Mackey, D.J. Analysis of phytoplankton of the Australian sector of the Southern Ocean: Comparisons of the microscopy and size frequency data with interpretations of pigment HPLC data using the ‘Chemtax’ matrix factorization program. Mar. Ecol. Prog. Ser. 1996, 144, 285–298. [Google Scholar] [CrossRef]
- Moreno, V.; Marrero, P.; Morales, J.; Llerandi García, C.; Villagarcía Úbed, M.D.; Rueda, M.J.; Llinás, O. Phytoplankton functional community structure in Argentinian continental shelf determined by HPLC pigment signatures. Estuar. Coast. Shelf Sci. 2012, 100, 72–81. [Google Scholar]
- Millie, D.F.; Schofield, O.M.; Kirkpatrick, G.J.; Johnsen, G.; Tester, P.A.; Vinyard, B.T. Detection of harmful algal blooms using photopigments and absorption signatures: A case study of the Florida red tide dinoflagellate, Gymnodinium breve. Limnol. Oceanogr. 1997, 42, 1240–1251. [Google Scholar] [CrossRef]
- Tango, P.; Butler, W.; Wazniak, C. Assessment of Harmful algal bloom species in the Maryland Coastal Bays. In Maryland’s Coastal Bays: Ecosystem Health Assessment; Chapter 7.2; Maryland Department of Natural Resources, Tidewater Ecosystem Assessment: Annapolis, MD, USA, 2004; pp. 11–27. [Google Scholar]
- Tango, P.; Butler, W.; Wazniak, C. Analysis of phytoplankton populations in the Maryland Coastal Bays. In Maryland’s Coastal Bays: Ecosystem Health Assessment; Chapter 8.1; Maryland Department of Natural Resources, Tidewater Ecosystem Assessment: Annapolis, MD, USA, 2004; pp. 2–34. [Google Scholar]
- Trice, T.M.; Gilbert, P.M.; Van Heukelem, L. HPLC pigment records provide evidence of past blooms of Aureococcus anophagefferens in the coastal bays of Maryland and Virginia, USA. Harmful Algae 2004, 3, 295–304. [Google Scholar]
- Wazniak, C.E.; Hall, M.R.; Carruthers, T.J.; Sturgis, B.; Dennison, W.C.; Orth, R.J. Linking water quality to living resources in a Mid-Atlantic lagoon system, USA. Ecol. Appl. 2007, 17, S64–S78. [Google Scholar] [CrossRef]
- Fertig, B.; O’Neil, J.M.; Beckert, K.A.; Cain, C.J.; Needham, D.M.; Carruthers, T.J.B.; Dennison, W. Elucidating terrestrial nutrient sources to a coastal lagoon, Chincoteague Bay, Maryland USA. Estuar. Coast. Shelf Sci. 2013, 116, 1–10. [Google Scholar] [CrossRef]
- Glibert, P.M.; Hinkle, D.C.; Sturgis, B.; Jesien, R.V. Eutrophication of a Maryland/Virginia Coastal Lagoon: A tipping point, ecosystem changes, and potential causes. Estuar. Coasts 2014, 37, 128–146. [Google Scholar]
- Oseji, O.F.; Chigbu, P.; Oghenekaro, E.; Waguespack, Y.; Chen, N. Spatial and temporal patterns of phytoplankton composition and abundance in the Maryland Coastal Bays: The influence of freshwater discharge and anthropogenic activities. Estuar. Coast. Shelf Sci. 2018, 207, 119–131. [Google Scholar] [CrossRef]
- Duan, S.; Chen, N.; Kaushal, S.; Chigbu, P.; Ishaque, A.; May, E.B.; Oseji, O.F. Dynamics of dissolved organic carbon and nitrogen in the Maryland Coastal Bays. Estuar. Coast. Shelf Sci. 2015, 164, 451–462. [Google Scholar] [CrossRef]
- Mulholland, M.R.; Boneillo, G.E.; Bernhardt, P.W.; Minor, E.C. Comparison of nutrient and microbial dynamics over a seasonal cycle in a Mid-Atlantic Coastal lagoon prone to Aureococcus anophagefferens (Brown Tide) blooms. Estuar. Coasts 2009, 32, 1176–1194. [Google Scholar] [CrossRef]
- Boynton, W.R.; Hagy, J.D.; Murray, L.; Stokes, C. A comparative analysis of eutrophication patterns in a temperate coastal Lagoon. Estuaries 1996, 19, 408–421. [Google Scholar] [CrossRef]
- Wazniak, C.; Hall, M.; Cain, C.; Wilson, D.; Jesien, R.; Thomas, J.; Carruthers, T.; Dennison, W. State of the Maryland Coastal Bays; A Report; Maryland Coastal Bays Program: Berlin, MD, USA, 2004; 44p. [Google Scholar]
- Pritchard, D.W. Salt balance and exchange rate for Chincoteague Bay. Chesap. Sci. 1960, 1, 48–57. [Google Scholar] [CrossRef]
- Bratton, J.F.; Bohlke, J.K.; Krantz, D.E.; Tobias, C.R. Flow and geochemistry of groundwater beneath a back-barrier lagoon: The subterranean estuary at Chincoteague Bay Maryland, USA. Mar. Chem. 2009, 113, 78–92. [Google Scholar] [CrossRef]
- Kennish, M.J.; Paerl, H.W. Coastal Lagoons: Critical Habitats of Environmental Change; CRC Press: Boca Raton, FL, USA, 2010; 568p. [Google Scholar]
- Jeffrey, S.W.; Wright, S.W. Qualitative and Quantitative Analysis of SCOR Reference Algal Cultures. In Phytoplankton Pigments in Oceanography: Guidelines to Modern Methods; Jeffrey, S.W., Mantoura, R.F.C., Wright, S.W., Eds.; UNESCO: Paris, France, 1997; pp. 343–360. ISBN 92-3-103275-5. [Google Scholar]
- Chen, J.; Oseji, O.; Mitra, M.; Waguespack, Y.; Chen, N. Phytoplankton pigments in Maryland Coastal Bay sediments as biomarkers of sources of organic matter to benthic Community. J. Coast. Res. 2015, 32, 768–775. [Google Scholar] [CrossRef]
- Zapata, M.; Rodríguez, F.; Garrido, J.L. Separation of chlorophylls and carotenoids from marine phytoplankton: A new HPLC method using a reversed phase C8 column and pyridine containing mobile phases. Mar. Ecol. Prog. Ser. 2000, 195, 29–45. [Google Scholar] [CrossRef]
- Ter Braak, C.J.F.; Smilauer, P. CANOCO Reference Manual and CanoDraw for Windows User’s Guide: Software for Canonical Community Ordination (Version 4.5); Biometris Wageningen: Wageningen, The Netherlands, 2002. [Google Scholar]
- Marshall, H.G. The composition of phytoplankton within the Chesapeake Bay plume and adjacent waters off the Virginia coast, U.S.A. Estuar. Coast. Shelf Sci. 1982, 5, 29–43. [Google Scholar] [CrossRef]
- Marshall, H.G.; Cohn, M.S. Distribution and composition of phytoplankton in northeastern coastal waters of the United States. Estuar. Coast. Shelf Sci. 1983, 17, 119–131. [Google Scholar] [CrossRef]
- Marshall, H.G.; Lacouture, R. Seasonal patterns of growth and composition of phytoplankton in the lower Chesapeake Bay and vicinity. Estuar. Coast. Shelf Sci. 1986, 23, 115–130. [Google Scholar]
- Hitchcock, G.; Phlips, E.; Brand, L.; Morrison, D. Plankton Blooms. In Florida Bay Science Program: A Synthesis of Research on Florida Bay; Fish and Wildlife Research Institute Technical Report TR-11; Hunt, J.H., Nuttle, W., Eds.; Florida Fish and Wildlife Research Institute: St Petersburg, FL, USA, 2007; pp. 77–91. [Google Scholar]
- Trigueros, J.M.; Ansotegui, A.; Orive, E.; Nó, M.L. Morphology and distribution of two brackish diatoms (Bacillariophyceae): Cyclotella atomus Hustedt and Thalassiosira guillardii Hasle in the estuary of Urdaibai (northern Spain). Nova Hedwig. 2000, 70, 431–450. [Google Scholar]
- Lohrenz, S.E.; Carroll, C.L.; Weidemann, A.D.; Tuel, M. Variations in phytoplankton pigments, size structure and community composition related to wind forcing and water mass properties on the North Carolina inner shelf. Cont. Shelf Res. 2003, 23, 1447–1464. [Google Scholar]
- Rodrı’guez, F.; Pazos, Y.; Maneiro, J.; Zapata, M. Temporal variation in phytoplankton assemblages and pigment composition at a fixed station of the Ria of Pontevedra (NW Spain). Estuar. Coast. Shelf Sci. 2003, 58, 499–515. [Google Scholar] [CrossRef]
- Litchman, E.; Klausmeier, C.A.; Yoshiyama, K. Contrasting size evolution in marine and freshwater diatoms. Proc. Natl. Acad. Sci. USA 2009, 106, 2665–2670. [Google Scholar] [CrossRef] [PubMed]
- Tamm, M.; Freiberg, R.; Tõnno, I.; Nõges, P.; Nõges, T. Pigment-Based Chemotaxonomy—A quick alternative to determine algal assemblages in large shallow eutrophic Lake? PLoS ONE 2015, 10, e0122526. [Google Scholar] [CrossRef]
- Seoane, S.; Laza-Martinez, A.; Orive, E. Monitoring phytoplankton assemblages in estuarine waters: The application of pigment analysis and microscopy to size-fractionated samples. Estuar. Coast. Shelf Sci. 2006, 67, 343–354. [Google Scholar] [CrossRef]
- Mallin, M. Phytoplankton ecology of North Carolina estuaries. Estuaries 1994, 17, 561–574. [Google Scholar] [CrossRef]
- Cohen, R.R.H. Physical processes and the ecology of a winter dinoflagellate bloom of Katodinium rotundatum. Mar. Ecol. Prog. Ser. 1985, 26, 135–144. [Google Scholar]
- Johnsen, G.; Sakshaug, E.; Vernet, M. Pigment composition, spectral characterization and photosynthetic parameters in Chrysochromulina polylepis. Mar. Ecol. Prog. Ser. 1992, 83, 241–249. [Google Scholar]
- Roy, S.; Llewellyn, C.A.; Egeland, E.S.; Johnsen, G. Phytoplankton Pigments: Characterization, Chemotaxonomy and Applications in Oceanography; Cambridge University Press: Cambridge, UK, 2012; 845p. [Google Scholar]
- Schluter, L.; Havskum, H. Phytoplankton pigments in relation to carbon content in phytoplankton communities. Mar. Ecol. Prog. Ser. 1997, 155, 55–65. [Google Scholar] [CrossRef]
- Latasa, M.; Scharek, R.; Le Gall, F.; Guillou, L. Pigment suites and taxonomic groups in Prasinophyceae. J. Phycol. 2004, 40, 1149–1155. [Google Scholar] [CrossRef]
- Phlips, E.J.; Badylak, S.; Christman, M.C.; Lasi, M.A. Climatic trends and temporal patterns of phytoplankton composition, abundance and succession in the Indian River Lagoon, Florida, USA. Estuar. Coasts 2010, 33, 498–512. [Google Scholar] [CrossRef]
- Deonarine, S.N.; Gobler, C.J.; Lonsdale, D.J.; Caron, D.A. Role of zooplankton in the onset and demise of harmful brown tide blooms (Aureococcus anophagefferens) in US mid-Atlantic estuaries. Aquat. Microb. Ecol. 2006, 4, 181–195. [Google Scholar] [CrossRef]
- Badylak, S.; Phlips, E.J. Spatial and temporal patterns of phytoplankton composition in a subtropical coastal lagoon, the Indian River Lagoon, Florida, USA. J. Plankton Res. 2004, 26, 1229–1247. [Google Scholar] [CrossRef]
- Phlips, E.J.; Badylak, S.; Grosskopf, T. Factors affecting the abundance of phytoplankton in a restricted sub-tropical lagoon, the Indian River Lagoon, Florida, USA. Estuar. Coast. Shelf Sci. 2002, 55, 385–402. [Google Scholar] [CrossRef]
- Bellinger, E.G.; Sigee, D.C. Freshwater Algae: Identification and Use as Bioindicators; John Wiley and Sons: Oxford, UK, 2010; 265p. [Google Scholar]
- Abirhire, O.; North, R.L.; Hunter, K.; Vandergutch, D.; Sereda, J.; Hudson, J. Environmental factors influencing phytoplankton communities in Lake Diefenbaker, Saskatchewan, Canada. J. Great Lakes Res. 2015, 41, 118–128. [Google Scholar] [CrossRef]
- Örnólfsdóttir, E.B.; Lumsden, S.E.; Pinckney, J.L. Nutrient pulsing as a regulator of phytoplankton abundance and community composition in Galveston Bay, Texas. J. Exp. Mar. Biol. Ecol. 2004, 303, 197–220. [Google Scholar] [CrossRef]
- Dorado, S.; Booe, J.; Steichen, J.; McInnes, A.; Windham, R.; Shepard, A.; Lucchese, A.; Preischel, H.; Pinckney, J.L.; Davis, S.; et al. Towards an understanding of the interactions between freshwater inflows and phytoplankton communities in a subtropical estuary in the Gulf of Mexico. Public Libr. Sci. One 2015, 10, e0130931. [Google Scholar] [CrossRef] [PubMed]
- Gin, K.Y.H.; Zhang, S.; Lee, Y.K. Phytoplankton community structure in Singapore’s coastal waters using HPLC pigment analysis and flow cytometry. J. Plankton Res. 2003, 25, 1507–1519. [Google Scholar] [CrossRef]
- Ondrusek, M.E.; Bidigare, R.R.; Sweet, S.T.; Defreitas, D.A.; Brooks, J.M. Distribution of phytoplankton pigments in the North Pacific Ocean in relation to physical and optical variability. Deep Sea Res. 1991, 38, 243–266. [Google Scholar] [CrossRef]
- Donald, K.M.; Scanlan, D.J.; Carr, N.G.; Mann, N.H.; Joint, I. Comparative phosphorus nutrition of the marine cyanobacterium Synechococcus WH7803 and the marine diatom Thalassiosira weissflogii. J. Plankton Res. 1997, 19, 1793–1813. [Google Scholar] [CrossRef]
- Cloern, J.E.; Dufford, R. Phytoplankton community ecology: Principles applied in San Francisco Bay. Mar. Ecol. Prog. Ser. 2005, 285, 11–28. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pigment | Abr. | Marker for | Size (µM) | |
|---|---|---|---|---|
| 1 | Peridinin | Peri | Dinoflagellates | >20 |
| 2 | Fucoxanthin | Fuco | Diatoms | >20 |
| 3 | 19’-Butanoyloxy-Fucoxanthin | But-fuco | Pelagophytes | 2–20 |
| 4 | Neoxanthin | Neo | Chlorophytes | 2–20 |
| 5 | Prasinoxanthin | Pras | Prasinophytes | 2–20 |
| 6 | Violaxanthin | Viola | Chlorophytes | 2–20 |
| 7 | 19’-Hexanoyloxy-fucoxanthin | Hex-fuco | Prymnesiophytes | 2–20 |
| 8 | Alloxanthin | Allo | Cryptophytes | 2–20 |
| 9 | Lutein | Lut | Chlorophytes | 2–20 |
| 10 | Zeaxanthin | Zea | Cyanobacteria | <2 |
| 11 | Chlorophyll b | Chl b | Chlorophytes | <2 |
| 12 | Chlorophyll a | Chl a | NS |
| DOC (µM) | TDN (µM) | TDP (µM) | Silica (µM) | TDN/TDP | |||||
|---|---|---|---|---|---|---|---|---|---|
| Site | Mean | SE | Mean | SE | Mean | SE | Mean | SE | |
| 1 | 317.59 | 21.65 | 26.12 | 2.26 | 1.45 | 0.10 | 20.70 | 5.24 | 18.01 |
| 2 | 321.25 | 21.61 | 35.99 | 4.92 | 1.80 | 0.21 | 33.46 | 5.11 | 19.99 |
| 3 | 332.8 | 30.18 | 29.19 | 3.70 | 1.54 | 0.13 | 18.59 | 5.43 | 18.95 |
| 4 | 339.11 | 28.14 | 31.61 | 3.37 | 1.85 | 0.44 | 23.70 | 4.94 | 17.09 |
| 5 | 327.85 | 24.23 | 37.53 | 7.74 | 1.57 | 0.15 | 20.83 | 3.36 | 23.90 |
| 6 | 513.67 | 51.91 | 42.98 | 5.17 | 1.53 | 0.15 | 21.77 | 4.6 | 28.09 |
| 7 | 250.45 | 31.62 | 28.65 | 5.34 | 1.45 | 0.12 | 20.65 | 4.98 | 19.76 |
| 8 | 287.4 | 86.05 | 36.6 | 11.59 | 1.46 | 0.13 | 11.12 | 2.69 | 25.07 |
| 9 | 344.01 | 77.17 | 48.8 | 13.16 | 1.54 | 0.13 | 19.70 | 5.94 | 31.69 |
| 10 | 462.01 | 113.19 | 53.94 | 19.48 | 1.56 | 0.10 | 21.11 | 5.79 | 34.58 |
| 11 | 431.03 | 129.13 | 49.94 | 15.81 | 1.50 | 0.15 | 18.92 | 4.48 | 33.29 |
| 12 | 468.87 | 86.92 | 49.42 | 10.77 | 1.42 | 0.14 | 18.27 | 4.16 | 34.80 |
| 13 | 502.14 | 104.73 | 53.49 | 14.31 | 1.63 | 0.10 | 17.27 | 4.15 | 32.82 |
| Kingdom Chromista | Species | Kingdom Chromista | Species |
|---|---|---|---|
| Phylum Bacillariophyta | *Achnanthes sp. Bory | Phylum Bacillariophyta | *Hemiaulus sp. Ehrenberg |
| *Actinoptychus sp. C.G. Ehrenberg | *Leptocylindrus danicus Cleve | ||
| *Amphiprora sp. Ehrenberg | Leptocylindrus minimus Gran | ||
| *Asterionella glacialis Castracane | *Licmophora sp. C.A. Agardh | ||
| *Aulacoseira granulata (Ehrenberg) Simonsen | Melosira moniliformis (O.F.Müller) C. Agardh | ||
| *Biddulphia mobiliensis (J.W.Bailey) Grunow | Melosira nummuloides C.A. Agardh | ||
| *Cerataulina bergonii Ostenfeld | Melosira sp. C.A. Agardh | ||
| *Chaetoceros debilis Cleve | *Minutocellus polymorphus (Hargraves & Guillard) | ||
| Chaetoceros eibenii Grunow | *Navicula sp. J.B.M. Bory de Saint-Vincent | ||
| Chaetoceros neogracilis S.L.VanLand. | Nitzschia pungens Grunow ex Cleve | ||
| Chaetoceros sp. C.G. Ehrenberg | Nitzschia sp. A.H. Hassall | ||
| Chaetoceros subtilis Cleve | *Odontella aurita (Lyngbye) C.Agardh | ||
| *Cocconeis sp. C.G. Ehrenberg | Odontella mobilensis (Bailey) Grunow | ||
| *Corethron criophilum Castracane | *Paralia sulcata (Ehrenberg) Cleve | ||
| Corethron sp. Castracane | *Pleurosigma sp. W. Smith | ||
| *Coscinodiscus sp. C.G. Ehrenberg | Pseudonitzschia sp. H. Peragallo | ||
| Cyclotella sp. (F.T. Kützing) A. de Brébisson | *Proboscia alata (Brightwell) Sundström | ||
| Cylindrotheca closterium (Ehrenberg) Reimann & | Rhizosolenia setigera Brightwell | ||
| *Cymbella sp. C.A. Agardh | Rhizosolenia stolterfothii H.Peragallo | ||
| Dactyliosolen fragilissimus (Bergon) Hasle | Rhizosolenia styliformis T.Brightwell | ||
| Dactyliosolen sp. A.F. Castracane | Rhizosolenia hebetata F. Semipina (Hensen) Gran | ||
| *Ditylum brightwellii (T.West) Grunow | *Schroederella delicatula (H.Peragallo) Pavillard | ||
| *Eucampia zodiacus Ehrenberg | Skeletonema costatum (Greville) Cleve | ||
| *Flagilaria sp. Lyngbye | *Striatella sp. | ||
| *Guinardia flaccida (Castracane) H.Peragallo | *Tabellaria sp. Kutzing | ||
| Guinardia delicatula (Cleve) Hasle | *Thalassionema sp. | ||
| Gyrosigma sp. A.H. Hassall | Thalassiosira sp. Cleve | ||
| Phylum Miozoa | Alexandrium sp. Halim | Phylum Chlorophyta (Kingdom Plantae) | *Ankistrodesmus sp. Corda |
| *Amphidinium crassum Lohmann | *Pediastrum duplex Meyen | ||
| Amphidinium sphenoides Wulff | *Chlorella sp. M.Beijerinck | ||
| *Tripos furca (Ehrenberg) F.Gómez | Pyramimonas sp. Schmarda | ||
| Ceratium sp. Shrank | Unid. Chlorophycean sphere | ||
| Ceratium tripos (O.F.Müller) Nitzsch | *Scenedesmus quadricauda (Turpin) Brébisson | ||
| Dinophysis acuminata Claparède & Lachmann | Scenedesmus sp. Meyen | ||
| Dinophysis sp. Ehrenberg | Phylum Cryptophyta (Kingdom Chromista) | Cryptomonas sp. Ehrenberg | |
| Diplopsalis sp. Bergh | Phylum Haptophyta (Kingdom Chromista) | Chrysochromulina sp. Lackey | |
| *Gonyaulax sp. Diesing | Phylum Cercozoa (Kingdom Chromista) | Paulinella ovalis (A.Wulff) P.W. Johnson, P.E. Hargraves & J.M. Sieburth | |
| Gonyaulax spinifera (Claparède & Lachmann) Diesing | Phylum Euglenozoa (Kingdom Protozoa) | Euglena sp. Ehrenberg | |
| Akashiwo sanguinea (K.Hirasaka) G. Hansen & O. Moestrup | Eutreptia sp. Perty | ||
| *Gyrodinium lachryma Meunier | Phylum Ochrophyta (Kingdom Chromista) | *Apedinella radians (Lohmann) P.H.Campbell Chattonella sp. B.Biecheler Ochromonas sp. Vysotskii | |
| Gyrodinium sp. Kofoid & Swezy | |||
| Gymnodinium sp. Stein | |||
| Heterocapsa triquetra (Ehrenberg) Stein | |||
| Heterocapsa rotundata (Lohmann) G. Hansen | |||
| Karlodinium veneficum (D. Ballantine) J.Larsen | |||
| Prorocentrum micans Ehrenberg | |||
| Prorocentrum cordatum (Ostenfeld) J.D. Dodge | |||
| Prorocentrum sp. Ehrenberg | |||
| Prorocentrum triestinum J.Schiller | |||
| *Protoperidinium brevipes Paulsen | |||
| Protoperidinium depressum Bailey | |||
| Protoperidinium sp. Bergh | |||
| *Scrippsiella trochoidea (Stein) Loeblich III |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oseji, O.F.; Fan, C.; Chigbu, P. Composition and Dynamics of Phytoplankton in the Coastal Bays of Maryland, USA, Revealed by Microscopic Counts and Diagnostic Pigments Analyses. Water 2019, 11, 368. https://doi.org/10.3390/w11020368
Oseji OF, Fan C, Chigbu P. Composition and Dynamics of Phytoplankton in the Coastal Bays of Maryland, USA, Revealed by Microscopic Counts and Diagnostic Pigments Analyses. Water. 2019; 11(2):368. https://doi.org/10.3390/w11020368
Chicago/Turabian StyleOseji, Ozuem F., Chunlei Fan, and Paulinus Chigbu. 2019. "Composition and Dynamics of Phytoplankton in the Coastal Bays of Maryland, USA, Revealed by Microscopic Counts and Diagnostic Pigments Analyses" Water 11, no. 2: 368. https://doi.org/10.3390/w11020368
APA StyleOseji, O. F., Fan, C., & Chigbu, P. (2019). Composition and Dynamics of Phytoplankton in the Coastal Bays of Maryland, USA, Revealed by Microscopic Counts and Diagnostic Pigments Analyses. Water, 11(2), 368. https://doi.org/10.3390/w11020368