Abstract
The effects of fish cannery discharge (FCD) on bacteria in marine coastal sediments were investigated. Redox potentials were measured, and granulometry was determined by wet ASTM sieving, and with the Sedigraph method. Prokaryotic abundance (PA) was determined by epifluorescence microscopy (DAPI staining), and faecal indicator bacteria (FIB) enumerated with the multiple test tube and most probable number method. Total lipids were determined gravimetrically, and sterols analysed by GC/MSD. Bacterial community composition was determined after total DNA isolation, Illumina MiSeq amplification, and SILVAngs processing pipeline. The FCD was rich in lipids, heterotrophic prokaryotes and FIB. The bacterial community of the FCD was dominated by Firmicutes and Gammaproteobacteria and many potentially pathogenic bacteria. Highly porosusgravelly sands clogged with fish remains transitioned to less permeable sandy muds away from the FCD. All sediments were anoxic with extremely negative potentials around the outfall. High surface PA and FIB spread 300 m from the outfall. Gammaproteobacteria and Deltaproteobacteria appeared in all sediments. Sulfurovum and Anaerolineaceae characterized the most polluted locations where gammaproteobacterial Woeseiaceae/JTB255 marine benthic group declined. Gammaproteobacteria and Bacteroidetes characterized surface sediments, while Chloroflexi and Deltaproteobacteria prevailed in deeper layers. The FCD enriched sediments in lipids and allochthonous bacteria degrading sanitary quality, lowering the permeability, redox potential, and bacterial diversity.
1. Introduction
Food processing industries, such as fish canneries are a significant source of pollution in the Mediterranean Sea [1]. Their effluents contain large amounts of fats, oils, fish remains, ammonia, phosphate, sulphur and often have a high biochemical oxygen demand. The bacterial community of such effluents can be enriched with phyla such as Bacteroidetes, Chloroflexi, Thermotogae, Synergystetes, and Firmicutes (in a case study of a tuna cannery) which are often characteristic of anaerobic environments [2]. Fish cannery discharges can also contain specific bacterial groups able to produce extracellular lipase enzymes that hydrolyze triglycerides and cholesterol [3,4,5,6,7,8]. Moreover, faecal indicator bacteria and various potentially pathogenic genera [9] are commonly very abundant in such effluents, indicating their negative impact on the sanitary quality of the receiving environment. In particular, fish and molluscs, when exposed to foreign potentially pathogenic microbes, can display various negative immune responses proteins [10,11,12].
Marine coastal sediments represent a preferential sink for various organic and inorganic contaminants which can affect their biological communities. Since bacteria are the most numerous organisms in sediments and have a crucial role in biogeochemical cycles, is very important to assess the potential impact of certain types of human contamination on their diversity, distribution and abundance. In general, sediment bacteria are prevalently attached to sediment grain surfaces, a vast matrix of inorganic and organic solid surfaces with heterogeneous and complex organic polymers that provide the substrates for their growth [13]. Recent studies on bacterial communities in intertidal and sublittoral surfaces by next-generation sequencing of the 16 S rRNA have identified some of the dominant groups within the bacterial assemblages of coastal marine sediments. Those bacterial communities typically include cosmopolitan taxa of Gamma- and Deltaproteobacteria such as the Woeseiaceae/JTB255 marine benthic group and the uncultured deltaproteobacteria Sva0081 sediment group, respectively, which have a multifaceted role in carbon and sulphur cycling [14,15]. Other benthic taxa that appear in marine coastal and intertidal sediments belong to Epsilonproteobacteria, Chloroflexi, Bacteroidetes, Planctomycetes, Acidobacteria and Firmicutes [16,17,18,19]. The distribution and abundance of those dominant members of the bacterial community are correlated to the median grain size of the sediment, to the amount of dissolved oxygen [19] and the permeability at the specific site [20]. Some researches [21,22] have also reported that the size and diversity of sediment bacterial communities can be influenced negatively by the presence of wastewaters coming from urban areas, petroleum refineries and shipyards.
In this study, the main aim was to investigate the influence of industrial organic contamination and changes in sediment conditions on bacterial community structure and abundance in surface (0 cm) and subsurface (5 cm) sediments. For this reason, the sediments of a semi-enclosed inlet, (Valdibora bay in the NE-Adriatic), exposed to a fish cannery wastewater were analysed. Besides, since the cannery wastewaters did not receive any treatment, the presence of FIB and PPB was quantified in the effluent and the sediments to assess potential sanitary risk and evaluate the input of allochthonous bacteria to marine sediments.
2. Materials and Methods
2.1. Study Location and Sampling
Valdibora bay is located on the northern side of Rovinj occupying an area of 1.3 km2 (Figure 1). The bay has an average depth of 16 m that slightly increases towards the west to a maximum of 22 m. In this area, there are various minor sources of pollution such as drainage of storm waters, occasional leakages from a few septic tanks, and a significant input of wastewaters represented by a fish cannery discharge.
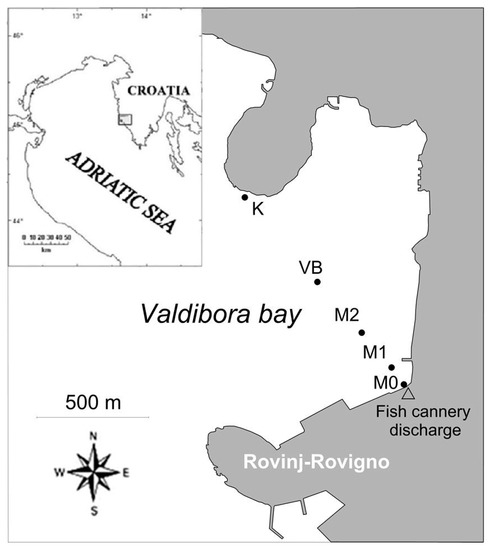
Figure 1.
Sampling stations in Valdibora bay.
The cannery has been operating more-less continuously since 1877 and has provided a considerable and long-term input of wastewaters to the bay of Valdibora. Every month, the cannery releases between 15,000 and 20,000 m3 of untreated effluents while other minor sewage outfalls and leaching from old septic tanks provide ~1000 m3 of sewage waters. Those waters can be considered hypoxic since, at maximum, oxygen reaches a saturation of 40% (municipal service of Rovinj, pers. comm.).
During a single sampling in February 2016, sediment cores (10.7 cm2 in section and 10 cm in length) were collected by scuba divers at five stations located along a transect stretching from the fish cannery outfall to the opposite side of the bay. The station M0 was situated in the proximity of the outfall (5 m distance) while other stations were located at increasing distances from the outfall: at 100 m (M1, 5 m depth), at 300 m (M2, 10 m depth) and at 600 m (VB, 19 m depth). At 1100 m from the outfall, station K (6 m depth) was selected as a control station as it was outside the reach of the primary pollution sources. Wastewater samples from the FCD were collected in triplicate from the discharge pipe just before the outfall.
2.2. Redox Potential (Eh), Granulometric Composition and Permeability
Duplicate sediment cores were collected at each station to determine the redox potential (Eh). The sediment samples were sealed with rubber caps and analysed in the laboratory. Redox potential (Eh) was measured in situ by a vertical penetration (every cm) of a Pt electrode connected to an Ag/AgCl reference electrode. The standardization of the electrode system was performed with quinhydrone buffer solutions of pH = 4 and pH = 7 prepared according to Metrohm, Switzerland. For the granulometric composition analysis, a single composite sample (down to 5 cm) was taken from each station. Each sample was wet sieved through a set of 7 standard ASTM sieves (4, 2, 1, 0.5, 0.25, 0.125, 0.063 mm mesh) in one ϕ-interval. The fraction that passed through the 0.063 mm sieve was collected and analysed following the standard sedigraph procedure [23]. The material retained on the sieves was dried and weighed. The data obtained by both techniques were merged to determine a continuous grain size range and analysed with the statistic package Gradistat v 6.0. Sediments were classified according to the Folk’s scheme based on gravel-sand-mud ratio [24].
The sediment permeability was calculated based on median grain size (dg) following the empirical relation by [25]: k = Dar · 735·106·dg2, where k is the permeability in m2, and Dar is the conversion factor for unit Darcy into m2 (=9.869 × 10−13). Based on median grain size and permeability, the sediment could be classified as fine-grained and low permeable to impermeable (dg < 165 µm, k < 2 × 10−11 m2), medium-grained and medium permeable (165 µm < dg < 370 µm, 2 × 10−11 m2 < k < 1 × 10−10 m2), and coarse-grained and highly permeable (370 µm < dg, 1 × 10−10 m2 < k).
2.3. Sanitary Quality of Sediments and Wastewaters
Faecal coliforms and streptococci were determined in the surface layer and from the depth of 5 cm in the sediments using the multiple test tube most probable number (MPN) method [26]. Sediment samples (5 g) were diluted by ten times their weight in phosphate buffer (44.75 mL) amended with 0.25 mL of Tween 80. Once diluted, the samples were homogenized three times for 3 min in an ultrasonic bath. As follows, 10-fold dilutions were prepared using phosphate buffer without Tween 80.
The quantification of FC was performed by testing five replicates from each dilution (10, 1 and 0.1 mL). Each replicate was inoculated into individual tubes containing lactose broth and incubated at 35 °C for 24 h. The positive cultures (showed gas formation) were submitted to a confirmation test by inoculation in MacConkey broth and incubation at 44.5 °C for 24 h. Positive cultures showed a yellow colour development from the violet-like colour of the medium. To determine FS five replicates from each multiple test tube dilution (10, 1 and 0.1 mL) were inoculated into individual tubes containing azide dextrose broth and incubated at 35 °C for 48 h. All the cultures showing turbidity after the incubation were subjected to the confirmation test in ethyl violet azide broth and incubated at 35 °C for 24 h. The presence of FS was indicated by the formation of a purple precipitate at the bottom of the tube, or occasionally by dense turbidity, all according to manufacturer guideline [27]. The positive readings from the confirmation tests (MacConkey and ethyl violet azide broth) were recorded and compared with the MPN index to get the MPN of FC and FS. Since the 95% confidence intervals for these data are relatively large, the acquired results were used to evaluate a general trend for faecal pollution in sediments.
Faecal coliforms and streptococci in wastewaters from the FCD were quantified by the membrane filtration method [26]. Sample aliquots of 0.1 mL, 0.01 mL and 0.001 mL were diluted with phosphate buffer and filtered through 0.45 µm pore size membrane filters (47 mm diameter). For FC counts membrane filters were incubated on the surface of mFC agar at 44.5 °C for 24 h. The colonies with a characteristic blue colour were counted. FS counts were determined by incubating membrane filters on the surface of Slanetz-Bartley agar at 36 °C for 48 h. Filters that developed red centred colonies were subjected to a confirmation test by transferring them and incubating on the surface of bile aesculin agar at 44.5 °C for two hours. The colonies characterized by a brown ring were faecal streptococci. The incubations for all samples were performed in dry-air bacteriological incubators.
2.4. Prokaryotic Abundance (PA)
A sediment aliquot (20 cm3) from the surface layer and the depth of 5 cm from each station was immediately preserved in formaldehyde (4% final concentration) and stored at 4 °C) until further processing. Subsamples from the sediment aliquot were diluted in filter-sterilized seawater amended with Tween 80, shook and sonicated with a probe. An aliquot (2 mL) was stained with 4,6-diamidino-2-phenylindole (DAPI; 1 μg mL−1 final conc.) for 10 min and filtered onto 0.2 μm black polycarbonate filters (Nuclepore, Whatman, UK). PA was determined by epifluorescence microscopy (Leitz Laborlux D). The counted number of signals was expressed as the number of prokaryotic cells per g of wet sediment or per mL of effluent [28].
2.5. Total Lipid and Sterol Analysis
A sediment aliquot from the surface layer and the depth of 5 cm from each station was lyophilized (Labconco FreeZone) and 6–7 g was taken for analysis. Particulate matter from the wastewater of the FCD was collected onto 0.7 µm glass fibre filters (pre-heated at 450 °C) by filtering 100 mL of sewage. Total lipids were determined after extraction following [29].
The extraction and purification procedure for the sterol determination was performed according to [30]. After derivatization with BSTFA-TMCS (at 60 °C for 1 h [31] sterols were analysed by Agilent gas_liquid chromatography (GLC) 6890 N GC System equipped with a 5973 Network Mass Selective Detector, Zebron ZB-5 MSi capillary column (30 m × 0.25 mm × 0.25 μm; 5% Phenyl—95% Dimethylpolysiloxane) and ultra-high purity helium as the carrier gas. The GLC settings were programmed to get a column temperature rise from 150 °C (1 min) by 20 °C/min up to 310 °C (5 min), at a constant column pressure of 2.17 kPa. Retention times, peak areas and mass spectra were recorded with Chemstation software. Data were acquired in the full scan mode between ions of m/z 50 and 550. Sterol standards including coprostanol (COP), cholesterol (CHL), β–sitosterol (b-SIT), 5 α cholestane, perylene (IS) and N, O-bis(trimethylsilyl)trifluoroacetamide (BSTFA) with 1% trimethylchlorosilane (TMCS) were provided by Sigma-Aldrich Chemical Company (Steinheim, Germany).
2.6. Extraction and Sequencing of DNA
A sample of 100 mL of wastewater from the FCD was filtered onto 0.2 µm Nucleopore polycarbonate membrane filters (Whatman, UK) with a vacuum pump. Filters were stored in 1 mL sucrose buffer (40 mM EDTA, 50 mM Tris-HCl and 0.75 M sucrose), frozen in liquid nitrogen and afterwards stored at −80 °C. The DNA was extracted according to [32] 1 g of sediment aliquots from the surface layer and the depth of 5 cm of each station was weighted, and the DNA was extracted using the UltraClean Soil DNA Isolation Kit (MO BIO, USA) according to the manufacturer’s instructions. The bacterial V3-V4 16 S rRNA region was amplified using bacterial primers S-D-Bact-0341-b-S-17 (5’-CCTACGGGNGGCWGCAG-3’) and S-D-Bact-0785-a-A-21 (5’-GACTACHVGGGTATCTAATCC-3’) [33] in four parallel reactions. Each 25 µL PCR reaction contained: 1× DreamTaq Green PCR Master Mix (Thermo Fisher Scientific, Waltham, MA, USA), 0.5 µM of forward and reverse primers and 10 ng of DNA template. The PCR amplification conditions were: 5 min initial denaturation at 95 °C, 30 cycles of 40 s denaturation at 95 °C, 2 min annealing at 55 °C and 1 min elongation at 72 °C, finalized by 10 min at 72 °C. After pooling of the replicate reactions, PCR products were purified using the Wizard SV Gel and PCR Clean-Up System (Promega, Madison, WI, USA) and sent for sequencing on the Illumina MiSeq platform (2 × 250 bp paired-end) at IMGM Laboratories (Martinsried, Germany).
The forward and reverse sequences contained in fastq files were assembled using mothur’s command and split into sample-specific fasta files using mothur’s command split groups [34]. Multifasta files were processed by the SILVAngs 1.3 pipeline [35] as described in [36]. Briefly, sequences were aligned against the SILVA SSU rRNA SEED using the SILVA Incremental Aligner (SINA) [37]. Sequences shorter than 50 aligned nucleotides, with more than 2% of ambiguities or 2% of homopolymers were removed. Putative contaminations and artefacts reads with a low alignment quality (50 alignment identity, 40 alignment score reported by SINA), were excluded from downstream analysis. Identical sequences were identified (dereplication) and the unique sequences were clustered (Operational Taxonomic Units [OTU]) at 97% sequence identity using cd-hit-est [38] running in accurate mode and ignoring overhangs. The representative OTU sequence was classified against the SILVA SSU Ref dataset (release 123.1) using blastn (version 2.2.30 +) with standard settings [39].
Statistical data regarding the SILVAngs pipeline analysis are given in the supplementary materials (Table S1). The sequencing effort applied was insufficient to determine the whole bacterial richness, as could be observed in the rarefaction curves that did not level off even for the samples with the highestnumber of sequences (Figure S1).
The dataset of maximum successful sequencing depth by SILVAngs was used for the analysis of principal coordinates (PCoA) to asses and illustrate the differences in bacterial community composition in the sediments. PCoA was based on Bray-Curtis dissimilarity matrix (Archaea, chloroplast related sequences and No Relative taxa were excluded) and done by R software environment.
3. Results
3.1. Granulometric Composition, Permeability and Redox Potential (Eh)
According to the granulometric composition, median grain sizes (dg) and permeability (k), the sediments from coastal stations M0 and K were classified as gravelly sandy (gS), coarse-grained and highly permeable. Due to the gradual increase of silt and clay fractions toward the central part of the bay the type of sediment changed to slightly gravelly muddy sand (g)mS at M1, and slightly gravelly sandy mud (g)sM at M2 and VB. The sediment at those stations was classified as fine-grained, low permeable to impermeable (Table 1).

Table 1.
Type, median grain size (dg) and permeability (k) of sediments in the research area gS-gravelly sand, (g)mS-slightly gravelly muddy sand and (g)sM-slightly gravelly sandy mud.
At the station M0 the Eh values were very high in the negative in the whole sediment core and its overlying water, indicating anoxia and stable reductive conditions (Eh down to −405 mV). At M1 and M2, the overlying waters displayed a positive Eh while both layers of the sediment cores had negative Eh. At VB, the overlying water and the surface sediment had positive Eh, which became negative in the deeper layer while at station K the overlying water and both sediment layers had positive Eh (Figure 2).
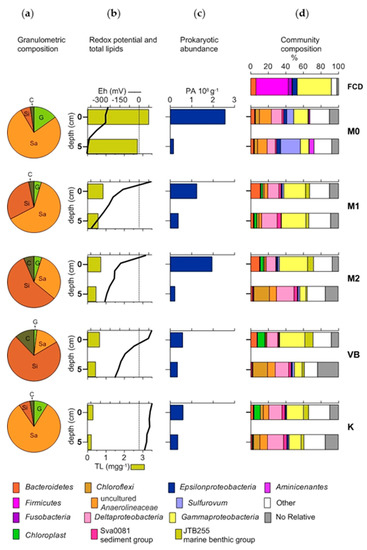
Figure 2.
(a) Granulometric composition (G-gravel, Sa-sand, Si-silt, C-clay), (b) redox potential (Eh, mV) and total lipid concentration (TL, mg g−1-dry sediment), (c) abundance of prokaryotes (PA, cells 108 g−1-wet sediment), (d) relative abundances of bacterial phyla in effluent (FCD) and sediment layers based on sequencing of 16 S rRNA genes (V3-V4 region). Only taxa that made up > 10% of total sequences in any given sample are presented.
3.2. Granulometric Composition, Permeability and Redox Potential (Eh)
The FCD effluent contained elevated concentrations of total lipids (TL) (387.9 mg L−1), cholesterol (CHL) and β-sitosterol (β-SIT) (Table 2). At the station M0 the concentrations of TL, CHL and β-SIT were the highest among the sediments and were similar in both layers. At this station, the relative abundance of coprostanol (COP) was higher by around two orders of magnitude with respect to the relative abundance of COP in the effluent. At the other stations, the concentration of total lipids showed a steady decline with increasing distance from the FCD. A common characteristic for all sediments was a mild enrichment of the surface layer with lipids and selected sterols with respect to the deeper layer (Figure 2).
Table 2.
Concentrations of the main sterols: coprostanol (COP), cholesterol (CHL) and β-sitosterol (β-SITin the FCD effluent (E) and sediment in Valdibora bay. The concentrations of sterols for effluent and sediment are expressed in μg L−1 and μg g−1, respectively.
3.3. Prokaryotic Abundance and Faecal Indicator Bacteria
The FCD effluent was characterized by very high PA (1.1 × 107 cells mL−1), FC (3 × 103 CFU mL−1) and FS (3 × 103 CFUmL−1), being each 0.03% out of the total PA. Both layers of sediment at M0, showed high concentrations of FIB while at M1 and M2, only the surface layer was enriched with FIB (Figure 3). At the remaining stations, FIB were still higher in the surface layer. Nevertheless, their overall concentration was relatively low, especially at K.
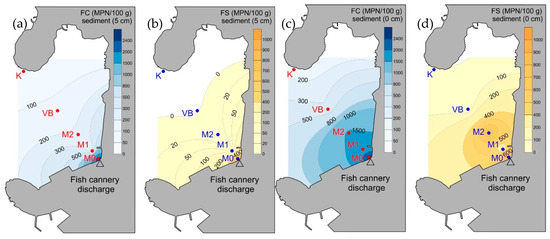
Figure 3.
Spatial distribution of faecal. indicator bacteria in sediments: (a) faecal coliforms at 0 cm, (b) faecal streptococci at 0 cm, (c) faecal coliforms at 5 cm, and (d) faecal streptococci at 5 cm.
In general, the levels of FS in sediments were at least twice as low as FC in the whole study area. PA showed a definite increase in the surface layer of M0, M1 and M2, with respect to the other stations but also to the corresponding deeper layers of sediment. At VB and K, the differences in PA between the layers were much lower than at M0, M1 and M2 (Figure 2).
3.4. The Structure of Bacterial Community
The alpha diversity (OTU richness) of bacterial communities in the FCD effluent was lower compared to most sediment samples. Within the surface sediment, alpha diversity continually increased from M0 towards VB and then decreased at station K. Generally, the alpha diversity was higher in the surface sediment with respect to the deeper one except for station M1 where the diversity was slightly increased in the lower layer (Table S2). The highest differences in diversity between the surface and the deeper layer were found at M0 (the most polluted) and VB (with the lowest permeability and smallest grain size).
The distribution of bacterial taxa in the FCD effluent and the sediment samples are presented in Figure 3. The bacterial community of the FCD effluent consisted prevalently of Gammaproteobacteria and Firmicutes. Gammaproteobacteria were characterized by the dominance of Acinetobacter, while among Firmicutes, the dominant genera were Vagococcus and Peptostreptococcus.
In the surface sediment at M0, the main representatives of the bacterial community were the phylum Chloroflexi and classes Gammaproteobacteria, Deltaproteobacteria and Epsilonproteobacteria. The shares of most of these taxa decreased (by about 4%) in the deeper sediment, while Epsilonproteobacteria Sulfuvorum considerably increased.
Some specific phyla such as Aminicenantes and Parcubacteria were detected in a noticeable proportion only at this station especially in the lower layer.
The surface sediment at M1 was characterized by Gammaproteobacteria (JTB255 marine benthic group, Marinicella, and Ectothiorhodospiraceae, Figure S2), Deltaproteobacteria (Sva0081 sediment group and uncultured Desulfobacteraceae, Sandaracinaceae and Syntrophobacteraceae, Figure S3) and Bacteroidetes. In the deeper sediment, Gammaproteobacteria kept a similar share while Deltaproteobacteria noticeably increased in comparison to the surface sediment.
The community structure of the surface sediment at M2 displayed a similar dominance of taxa (Gammaproteobacteria, Deltaproteobacteria, and Bacteroidetes) as at the station M1. The deeper sediment of M2 was characterized by a substantial increase of Deltaproteobacteria and a decrease of Gammaproteobacteria and Bacteroidetes. The phylum Chloroflexi also acquired increased importance in the deeper sediment where, besides uncultured Anaerolineaceae, different bacterial groups belonging to the class Dehalococcoidia were present.
At the station VB, the dominant taxa of the surface sediment consisted of Gammaproteobacteria, Deltaproteobacteria, chloroplast-related sequences and Bacteroidetes. The respective changes in their shares in deeper sediments were similar as observed in deeper sediment at the station M2.
At the control station K, apart from the dominance of Gammaproteobacteria, Deltaproteobacteria and chloroplast related sequences, Chloroflexi was also a relevant component of the surface sediment community. In the deeper sediment, Chloroflexi and Deltaproteobacteria became more abundant, while Gammaproteobacteria decreased in comparison to the surface layer.
The difference between bacterial community compositions of sediments was illustrated by the PCoA biplot (Figure 4), which explained 73.55% of the observed variations. By PCoA I, the sediments were mainly separated according to their depth and probably substrate quality (degradability), while PCoA II showed the separation according to the level of pollution.
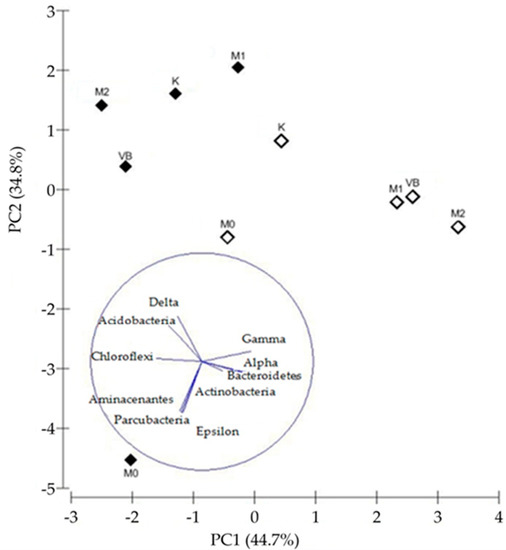
Figure 4.
Principal coordinates analysis (PCoA) ordination of a Bray-Curtis dissimilarity matrix based on the overall dataset of maximum successful sequencing depth by SILVAngs (15 most abundant taxa were presented as vectors) showing differences in bacterial community structures of surface (open diamonds) and deeper (black diamonds) sediments in Valdibora bay.
On PCoA I (44.95%) the relative abundance of gammaproteobacterial taxa separated the surface sediments (M1, M2, VB, K) and deeper sediment of M1 from all the other deeper sediments and surface sediment of M0, which had a higher relative abundance of taxa belonging to Chloroflexi, Delta- and Epsilonproteobacteria. The separation along PCoA II (28.60%) was governed by the abundance of “Candidatus Thiobios” and Marinicella (M1, M2), uncultured Desulfobacteraceae, R103-B63 and Sulfurovum (M0) in the polluted sediments and by the abundance of Sva0081 sediment group and MSBL5 (K, VB, M2), JTB255 marine benthic group and Ectothiorhodospiraceae (K, VB) in the less polluted ones.
The diversity and abundance of PPB were analysed at the genus level, according to [40]. The results showed that in the FCD effluent, the relative abundance of PPB was 24.18%, while in the sediment there was a substantial decrease with shares ranging between 0.41% and 0.98%. The most abundant genera of PPB in the FCD effluent were Pseudomonas, Arcobacter, Escherichia/Shigella, Aeromonas and Clostridium, while in the sediment Pseudomonas and Escherichia/Shigella were most noticeable. Some PPB genera (Campylobacter, Chlamydia and Mycobacterium) were detected in the sediment while being absent in the effluent. On the other hand, the genera Neisseria and Yersinia were identified only in the FCD effluent (Figure 5).
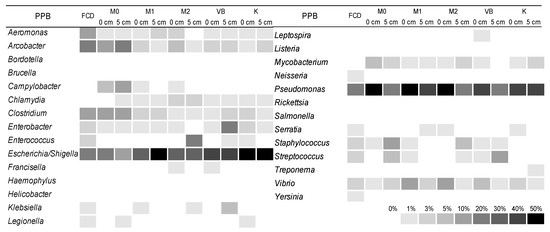
Figure 5.
Relative abundances (percentage of a specific pathogenic genus in total identified pathogenic bacteria) of potentially pathogenic bacterial genera in the effluent (FCD) and sediment layers.
4. Discussion
The untreated FCD effluent was substantially enriched with lipids with a significant contribution of CHL and β-SIT (from vegetable oil used in fish preservation) among sterols and deprived with oxygen as it usually occurs in similar industrial discharges. A high abundance of prokaryotes and faecal indicator bacteria was present in these waters. Such concentrations were more than twenty times higher than at an unpolluted referent station [40,41]. The ratio of FC/FS ~1 indicated the prevalence of a non-human source for these bacteria [42] since the cannery effluent contained exclusively fish remains and waste from fish processing. However, a relevant proportion of the effluent community was recognized as potentially pathogenic. Within the PPB community, the most abundant genera such as Aeromonas, Clostridium, Escherichia/Shigella, Pseudomonas, Staphylococcus, Streptococcus and Vibrio, have been previously found on fish and fish products [9]. On the other hand, Arcobacter, which was also equally represented in the FCD effluent, is considered a typical genus present in sewer systems [43].
The analysis of sterols suggested that COP in the FCD effluent did not derive from human faeces and probably resulted from bacterial reduction of CHL within the cannery pipe system where anoxic conditions persisted [44]. The FCD effluent contained a relevant proportion of anaerobic bacteria such as Lactobacillus and Bacteroides, which include genera and strains capable of reducing CHL to COP outside the human intestine [45,46]. Therefore, it can be assumed that COP production might be a result of their activity.
The phyla Firmicutes and Gammaproteobacteria dominated the bacterial community of the FCD effluent. Acinetobacter, the most abundant gammaproteobacterial genus in the FCD, has been shown to produce extracellular lipase enzymes that hydrolyze triglycerides (the main component of oils and fats) to fatty acids and glycerol [3,5]. Also, the genera Pseudomonas and Bacillus too abundant in the FCD have been found to have a potential role in the process of partial or complete cholesterol degradation [4].
The production of sulphides and ammonia due to sulphate reduction and protein breakdown during the decomposition of fish remains is typical for wastewaters from seafood processing industries [47]. Fish, including sardines treated in our cannery, can contain up to 260 mg S/100 g [48], providing a consistent source of sulphur-containing compounds to the pipe waters of the cannery discharge system. Within the bacterial community of the FCD effluent, several epsilonproteobacterial genera (Arcobacter, Sulfurimonas and Sulfurospirillum) containing strains capable of oxidizing sulphide to elemental sulphur [49,50] were detected suggesting their important role in the cycling of sulphur in the wastewater system.
The analysis of the bacterial community in the sediments has highlighted the ubiquitous presence of cosmopolitan bacteria from the JTB255 marine benthic group of the class Gammaproteobacteria in the surface sediments while Sva0081 sediment group of the Deltaproteobacteria were common and more abundant in the deeper layer. Both clades have been recognized to be critical players in the carbon and sulphur cycling in organic-rich sediments during its remineralization. JTB255 marine benthic group is one of the most significant dark carbon fixators fuelled by sulphur oxidation [11]. Its broad range of energy-yielding metabolisms possibly explains the general ubiquity and high abundance of this clade in marine sediments [15]. On the other hand, the Sva0081 sediment group, referring to sulphate-reducing bacteria, can use acetate, as a source of energy and carbon.
The bacterial community at station M0 exposed to chronic organic contamination differed substantially from all the other sampling sites in both layers. The main difference was due to the higher share of epsilonproteobacterial genera Sulfurovum/Sulfurimonas. These genera are involved in the oxidation of S0 [18], which regularly accumulate in pelagic oxygen minimum zones, hydrothermal vents and sediment surfaces [50]. Isolates of these genera are facultative or strict anaerobes which oxidize reduced sulphur species with nitrate or oxygen as electron acceptors [51]. Their dominance at impermeable sites in suboxic and anoxic niches has been previously reported in sublittoral surface sediments of the North Sea [20]. The station K, which had very similar granulometric composition and sediment permeability as M0 but without the enrichment of organic matter and clogging with fish remains showed some significant differences with respect to M0. At K, there was a much lower difference in bacterial abundance between the sediment layers and the overall presence of FIB was negligible. Other important differences emerged in the structure of the bacterial community, which was characterized by the absence of Epsilonproteobacteria and the prevalence of different subgroups inside the Gammaproteobacteria. The JTB255 marine benthic group represented the largest share within Gammaproteobacteria at K while it was very limited at M0, indicating its possible susceptibility to pollution.
Another relevant component of the bacterial community, which was detected mainly in deeper sediments was the phylum Chloroflexi, mostly represented by the class Dehalococcoidia (DEH). Chloroflexi phylotypes have been found as abundant bacterial groups in organic-rich deep marine sediments [13]. In our sediments, Chloroflexi, dominated by uncultured Anaerolineaceae, represented the third-largest share within the community of M0. This family is associated with the anaerobic degradation of long-chain n-alkanes [52], which have been previously found in high concentrations in the investigated area. Within the deeper layer of M0, an increasing share of the phylum Aminicentanes was also observed. This phylum might have been associated in a consortium with Anaerolineaceae and Actinobacteria and implicated in the further degradation of alkanes [52].
Apart from affecting the structure of the bacterial community the abundance of bacteria and FIB, the overall impact of the FCD could be seen as a general reduction in bacterial diversity which occurred in the proximity of the pollution source, similar to a previously reported case for northern Adriatic sediments influenced by hydrocarbon contamination [22].
A second important factor influencing bacterial diversity and community structure could be ascribed to the sediment characteristics such as granulometry and permeability. An increasing proportion of silt and clay toward the central part of the bay (M1, M2 and VB) has been accompanied by an increase in bacterial diversity in the surface layer with respect to both M0 (polluted) and K (unpolluted) which had bigger grain sizes. Such a response of bacterial diversity to grain size and permeability has also been shown for North Sea sediments [20]. Alpha diversity also displayed a decrease from the surface to the deeper layer of sediments, most probably due to a lower permeability and substrate availability. Regarding this trend, an exception was detected at the station M1, where bacterial diversity was a bit higher in the lower level, which might have been caused by wave resuspension or bioturbation by benthic fauna [53]. At stations M2 and VB in the central part of the bay, a significantly higher content of silt and clay, facilitated the accumulation of lipids which might have reduced the already low permeability of surface sediments and reduced the flow of relatively new organic matter into the deeper layers.
5. Conclusions
The FCD effluent introduced massive amounts of fatty fish remains, sulphur and allochthonous bacteria to the waters of Valdibora bay. The bacterial community of the effluent was characterized by the presence of Firmicutes and Gammaproteobacteria that include strains capable of degrading lipids and oxidize sulphide. The presence of high concentrations of FIB and a considerable PPB share in the effluent indicated a potential sanitary threat even though the profiles of faecal sterols and the ratios of FIB excluded a human faecal contribution. The influence of the FCD effluent on the sediment in the proximity of the outfall manifested as an increased concentration of FIB and lipid content by 1,2 orders of magnitude with respect to the unpolluted station K. The addition of lipids in the area lowered sediment permeability and redox potential. Genera belonging to Gamma- and Deltaproteobacteria, typically inhabiting sulfidic sediments, were the dominant part of the bacterial community in the sediments of Valdibora bay. The sediment alterations induced by the FCD affected the structure of the bacterial community by increasing the number of sequences related to Chloroflexi and Sulfurovum whose members are often involved in the degradation and oxidation of recalcitrant sulphur enriched organic matter. In the most polluted sediments, the decrease of the abundant and cosmopolitan member of marine sediments, the JTB255 marine benthic group, suggested its susceptibility to pollution. Apart from organic contamination, the other main factor which appeared to influence the bacterial community was the grain size of sediments. At the stations located in the middle part of the bay, an increase in alpha diversity in surface sediments occurred parallel to a decrease in grain size. However, this change in granulometric composition coupled with the lipid enrichment of the surface sediment reduced sediment permeability and pore water transport to the deeper layers of sediment. This condition was reflected by the bacterial community of the deeper layer as a reduction in bacterial diversity accompanied by a decrease in the relative abundance of sequences related to Gammaproteobacteria and Bacteroidetes and a relative increase of sequences related to Chloroflexi.
Supplementary Materials
The following are available online at https://www.mdpi.com/2073-4441/11/12/2566/s1, Figure S1: Rarefaction curves for the effluent (FCD) and sediment samples, Figure S2: Relative abundances of bacterial families, groups and genera of the phylum Gammaproteobacteria in effluent (FCD) and sediments, Figure S3: Relative abundances of bacterial families, groups and genera of the phylum Deltaproteobacteria in effluent (FCD) and sediments, Table S1: Sequence statistics obtained by SILVAngs pipeline, Table S2: Number of OTUs, richness estimates (Chao1 and Abundance-based Coverage Estimator [ACE] and Shannon’s diversity index following the normalisation step.
Author Contributions
Conceptualization, P.P., I.I. and A.B.; methodology, P.P., I.I. and I.F.; software, P.P., I.F. and A.B.; validation, I.I.; formal analysis I.I.; investigation, P.P. and I.F.; resources, P.P. and I.F.; data curation, I.I.; writing-original draft preparation, P.P. and A.B.; writing-review and editing, P.P. and A.B.; visualization, P.P., I.F. and A.B.; supervision I.I.; project administration, P.P.; funding acquisition, P.P.
Funding
This research received no external funding
Acknowledgments
We sincerely thank Mirjana Najdek Dragić for the general support and for the lipid and sterol analysis, and Marino Korlević for the molecular analysis.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- El-Kholy, R.; El Saadi, A.; Abdin, A.E.D. Pollution and Measures towards De-Pollution of the Mediterranean Sea. In Integrated Water Resources Management in the Mediterranean Region: Dialogue towards New Strategy; Springer: Dordrecht, The Netherlands, 2012; pp. 175–194. [Google Scholar]
- Militon, C.; Hamdi, O.; Michotey, V.; Fardeau, M.L.; Ollivier, B.; Bouallagui, H.; Hamdi, M.; Bonin, P. Ecological significance of Synergistetes in the biological treatment of tuna cooking wastewater by an anaerobic sequencing batch reactor. Environ. Sci. Pollut. Res. 2015, 22, 18230–18238. [Google Scholar] [CrossRef] [PubMed]
- El-Bestawy, E.; El-Masry, M.H.; El-Adl, N.E. The potentiality of free Gram-negative bacteria for removing oil and grease from contaminated industrial effluents. World J. Microbiol. Biotechnol. 2005, 21, 815–822. [Google Scholar] [CrossRef]
- Garcia, J.L.; Uhia, I.; Galán, B. Catabolism and biotechnological applications of cholesterol degrading bacteria. Microb. Biotechnol. 2012, 5, 679–699. [Google Scholar] [CrossRef] [PubMed]
- Phong, N.T.; Duyen, N.T.; Diep, C.N. Isolation and characterization of lipid-degrading bacteria in wastewater of food processing plants and restaurants in Can Tho city, Vietnam. Am. J. Life Sci. 2014, 2, 382–388. [Google Scholar] [CrossRef]
- Hassan, S.W.M.; Latif, H.H.A.E.; Ali, S.M. Production of Cold-Active Lipase by Free and Immobilized Marine Bacillus cereus HSS: Application in Wastewater Treatment. Front. Microbiol. 2018, 9, 2377. [Google Scholar] [CrossRef]
- Sandaruwani, A.; Kumarasinghe, C.; Samarakoon, D.; Ariyadasa, T.U.; Gunawardena, S.H.P. Investigation of the Efficiency of Dairy Wastewater Treatment Using Lipid-Degrading Bacterial Strains. In Proceedings of the Moratuwa Engineering Research Conference (MERCon), Moratuwa, Sri Lanka, 30 May–1 June 2018; pp. 362–366. [Google Scholar] [CrossRef]
- Kurhekar, J.V. Isolation and Characterization of Lipase Producing Bacteria from Restaurant Waste Water. World J. Pharm. Res. 2018, 6, 685–693. [Google Scholar]
- Novotny, L.; Dvorska, L.; Lorencova, A.; Beran, V.; Pavlik, I. Fish: A potential source of bacterial pathogens for human beings. Vet. Med. Czech 2004, 49, 343–358. [Google Scholar] [CrossRef]
- Fazio, F.; Saoca, C.; Costa, G.; Zumbo, A.; Piccione, G.; Parrino, V. Flow cytometry and automatic blood cell analysis in striped bass Morone saxatilis (Walbaum, 1792): A new hematological approach. Aquaculture 2019, 513, 734398. [Google Scholar] [CrossRef]
- Parrino, V.; Cappello, T.; Costa, G.; Cannavà, C.; Sanfilippo, M.; Fazio, F.; Fasulo, S. Comparative study of haematology of two teleost fish (Mugil cephalus and Carassius auratus) from different environments and feeding habits. Eur. Zool. J. 2018, 85, 194–200. [Google Scholar] [CrossRef]
- Parrino, V.; Costa, G.; Cannavà, C.; Fazio, E.; Bonsignore, M.; Saoca, C.; Piccione, G.; Fazio, F. Flow cytometry and micro-Raman spectroscopy: Identification of hemocyte populations in the mussel Mytilus galloprovincialis (Bivalvia: Mytilidae) from Faro Lake and Tyrrhenian Sea (Sicily, Italy). Fish Shellfish Immunol. 2018, 87, 1–8. [Google Scholar] [CrossRef]
- Inagaki, F.; Nunoura, T.; Nakagawa, S.; Teske, A.; Lever, M.; Lauer, A.; Suzuki, M.; Takai, K.; Delwiche, M.; Colwell, F.S.; et al. Biogeographical distribution and diversity of microbes in methane hydrate-bearing deep marine sediments on the Pacific Ocean Margin. Proc. Natl. Acad. Sci. USA 2006, 103, 2815–2820. [Google Scholar] [CrossRef]
- Dyksma, S.; Bischof, K.; Fuchs, B.M.; Hoffmann, K.; Meier, D.; Meyerdierks, A.; Pjevac, P.; Probandt, D.; Richter, M.; Stepanauskas, R.; et al. Ubiquitous Gammaproteobacteria dominate dark carbon fixation in coastal sediments. ISME J. 2016, 10, 1939–1953. [Google Scholar] [CrossRef]
- Mussmann, M.; Pjevac, P.; Krüger, K.; Dyksma, S. Genomic repertoire of the Woeseiaceae/JTB255, cosmopolitan and abundant core members of microbial communities in marine sediments. ISME J. 2017, 11, 1276. [Google Scholar] [CrossRef]
- Zinger, L.; Amaral-Zettler, L.A.; Fuhrman, J.A.; Horner-Devine, M.C.; Huse, S.M.; Welch, D.B.M.; Martiny, J.B.H.; Sogin, M.; Boetius, A.; Ramette, A. Global Patterns of Bacterial Beta-Diversity in Seafloor and Seawater Ecosystems. PLoS ONE 2011, 6, e24570. [Google Scholar] [CrossRef]
- Gobet, A.; Boer, S.I.; Huse, S.M.; Van Beusekom, J.E.E.; Quince, C.; Sogin, M.L.; Boetius, A.; Ramette, A. Diversity and dynamics of rare and of resident bacterial populations in coastal sands. ISME J. 2012, 6, 542–553. [Google Scholar] [CrossRef]
- Pjevac, P.; Kamyshny, A., Jr.; Dyksma, S.; Mussmann, M. Microbial consumption of zero-valence sulfur in marine benthic habitats. Environ. Microbiol. 2014, 16, 3416–3430. [Google Scholar] [CrossRef]
- Zheng, B.; Wang, L.; Liu, L. Bacterial community structure and its regulating factors in the intertidal sediment along the Liaodong Bay of Bohai Sea, China. Microbiol. Res. 2014, 169, 585–592. [Google Scholar] [CrossRef]
- Probandt, D.; Knittel, K.; Tegetmeyer, H.E.; Ahmerkamp, S.; Holtappels, M.; Amann, R. Permeability shapes bacterial communities in sublittoral surface sediments. Environ. Microbiol. 2017, 19, 1584–1599. [Google Scholar] [CrossRef]
- Drury, B.; Rosy-Marshall, E.; Kelly, J.J. Wastewater Treatment Effluent Reduces the Abundance and Diversity of Benthic Bacterial Communities in Urban and Suburban Rivers. Appl. Environ. Microbiol. 2013, 79, 1897–1905. [Google Scholar] [CrossRef]
- Korlević, M.; Žućko, J.; Najdek, M.; Blažina, M.; Pustijanac, E.; Zeljko, T.V.; Gačeša, R.; Baranašić, D.; Starčević, A.; Diminić, J.; et al. Bacterial diversity of polluted surface sediments in the northern Adriatic Sea. Syst. Appl. Microbiol. 2015, 38, 189–197. [Google Scholar] [CrossRef]
- Micromeritics SediGraph 5100 Particle Size Analysis System Operator’ Manual; Micromeritics Instrument Corporation: Norcross, GA, USA, 2002; Available online: https://www.micromeritics.com/pdf/products/sedigraph_5100_brochure.pdf (accessed on 4 December 2019).
- Folk, R.L. The distinction between grain size and mineral composition in sedimentary rock nomenclature. J. Geol. 1954, 62, 344–356. [Google Scholar] [CrossRef]
- Gangi, A.F. Permeability of unconsolidated sands and porous rocks. J. Geophys. Res. Solid 1985, 90, 3099–3104. [Google Scholar] [CrossRef]
- WHO. Guidelines for Health Related Monitoring of Coastal Recreational and Shellfish Areas. Part I: General Guidelines. In Long-term Programme for Pollution Monitoring and Research in the Mediterranean Sea (MED/POL Phase II); WHO: Geneva, Switzerland, 1994. [Google Scholar]
- Condalab, EVA Broth (Ethyl, Violet, Azide, Litsky) Cat. 1230, Conda Manual. Available online: https://www.chemie-brunschwig.ch/ (accessed on 11 November 2019).
- Epstein, S.S.; Rossel, J. Enumeration of sandy sediment bacteria: Search for optimal protocol. Mar. Ecol. Prog. Ser. 1995, 117, 289–298. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 910–917. [Google Scholar] [CrossRef]
- Isobe, K.O.; Tarao, M.T.; Zakaria, M.; Chiem, N.H.; Minh, L.Y.; Takada, H. Quantitative application of fecal sterols using gas chromatography-mass spectrometry to investigate fecal pollution in tropical waters: Western Malaysia and Mekong Delta, Vietnam. Environ. Sci. Technol. 2002, 36, 4497–4507. [Google Scholar] [CrossRef]
- Szűcs, S.; Sárváry, A.; Cain, T.; Ádány, R. Method validation for the simultaneous determination of fecal sterols in surface waters by gas chromatography—Mass spectrometry. J. Chromatogr. Sci. 2006, 44, 70–76. [Google Scholar] [CrossRef][Green Version]
- Massana, R.; Murray, A.E.; Preston, C.M.; Delong, E.F. Vertical distribution and phylogenetic characterization of marine planktonic archaea in the Santa Barbara Channel. Appl. Environ. Microbiol. 1997, 63, 50–56. [Google Scholar]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microb. 2009, 75, 7537–7541. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, 590–596. [Google Scholar] [CrossRef]
- Ionescu, D.; Christian, S.; Polerecky, L.; Munwes, Y.Y.; Lott, C.; Häusler, S.; Bižić-Ionescu, M.; Quast, C.; Peplies, J.; Glöckner, F.O.; et al. Microbial and Chemical Characterization of Underwater Fresh Water Springs in the Dead Sea. PLoS ONE 2012, 7, e38319. [Google Scholar] [CrossRef] [PubMed]
- Pruesse, E.; Peplies, J.; Glöckner, F.O. SINA: Accurate high-throughput multiple sequence alignment of ribosomal RNA genes. Bioinformatics 2012, 28, 1823–1829. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Godzik, A. Cd-hit: A fast program for clustering and comparing large sets of protein or nucleotide sequences. Bioinformatics 2006, 22, 1658–1659. [Google Scholar] [CrossRef] [PubMed]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [PubMed]
- Ivančić, I.; Paliaga, P.; Pfannkuchen, M.; Đakovac, T.; Najdek, M.; Steiner, P.; Korlević, M.; Markovski, M.; Baričević, A.; Smodlaka Tanković, M.; et al. Seasonal variations in extracellular enzymatic activity in marine snow-associated microbial communities and their impact on the surrounding water. FEMS Microbiol. Ecol. 2018, 94, 12. [Google Scholar] [CrossRef]
- Fuks, D.; Ivančić, I.; Najdek, M.; Njire, J.; Godrijan, J.; Maric Pfannkuchen, D.; Silovic, T.; Paliaga, P.; Blazina, M.; Precali, R.; et al. Changes in the planktonic community structure related to trophic conditions: The case study of the northern Adriatic Sea. J. Mar. Syst. 2012, 96, 95–102. [Google Scholar] [CrossRef][Green Version]
- Coyne, M.S.; Howell, J.M. The Fecal Coliform/Fecal Streptococci Ratio (FC/FS) And Water Quality in the Bluegrass Region of Kentucky. Soil Sci. 1994, 15, 1–4. [Google Scholar]
- Collado, L.; Figueras, M.J. Taxonomy, epidemiology and clinical relevance of the genus Arcobacter. Clin. Microbiol. Rev. 2011, 24, 174–192. [Google Scholar] [CrossRef]
- Paliaga, P.; Felja, I.; Ušić, U.; Ivančić, I.; Najdek, M. Accumulation and persistence of sewage and fish cannery pollution in coastal sediments (northern Adriatic Sea). J. Soils Sediments 2017. [Google Scholar] [CrossRef]
- Freier, T.A.; Beitz, D.C.; Li, L.; Hartman, P.A. Characterization of Eubacterium coprostanoligenes sp. nov., a cholesterol-reducing anaerobe. Int. J. Syst. Bacteriol. 1994, 44, 137–142. [Google Scholar] [CrossRef]
- Lye, H.S.; Rahmat-Ali, G.R.; Liong, M.T. Mechanisms of cholesterol removal by lactobacilli under conditions that mimic the human gastrointestinal tract. Int. Dairy J. 2010, 20, 169–175. [Google Scholar] [CrossRef]
- Omil, F.; Mendez, R.; Lema, J.M. Anaerobic treatment of saline wastewaters under high sulphide and ammonia content. Bioresour. Technol. 1995, 54, 269–278. [Google Scholar] [CrossRef]
- Murray, J.; Burt, J.R. The Composition of Fish; FAO Repository; FAO in Partnership with Support Unit for International Fisheries and Aquatic Research, SIFAR, Ministry of Technology, Torry Research Station: Rome, Italy, 2001. [Google Scholar]
- Gevertz, D.; Telang, A.J.; Voordouw, G.; Jenneman, G.E. Isolation and Characterization of Strains CVO and FWKO B, Two Novel Nitrate-Reducing, Sulfide-Oxidizing Bacteria Isolated from Oil Field Brine. Appl. Environ. Microbiol. 2000, 66, 2491–2501. [Google Scholar] [CrossRef] [PubMed]
- Lavik, G.; Stührmann, T.; Brüchert, V.; Van der Plas, A.; Mohrholz, V.; Lam, P.; Mussmann, M.; Fuchs, B.M.; Amann, R.; Lass, U.; et al. Detoxification of sulphidic African shelf waters by blooming chemolithotrophs. Nature 2009, 457, 581–585. [Google Scholar] [CrossRef]
- Inagaki, F.; Takai, K.; Kobayashi, H.; Nealson, K.H.; Horikoshi, K. Sulfurimonas autotrophica gen.nov., sp.nov., a novel sulfur-oxidizing epsilonproteobacterium isolated from hydrothermal sediments in the Mid-Okinawa Trough. Int. J. Syst. Evol. Microbiol. 2003, 53, 1801–1805. [Google Scholar] [CrossRef]
- Liang, B.; Wang, L.Y.; Mbadinga, S.M.; Liu, J.F.; Yang, S.Z.; Gu, J.D.; Mu, B.Z. Anaerolineaceae and Methanosaeta turned to be the dominant microorganisms in alkanes-dependent methanogenic culture after long-term of incubation. AMB Express 2015, 5, 37. [Google Scholar] [CrossRef]
- Böer, S.I.; Arnosti, C.; van Beusekom, J.E.E.; Boetius, A. Temporal variations in microbial activities and carbon turnover in subtidal sandy sediments. Biogeosciences 2009, 6, 1149–1165. [Google Scholar] [CrossRef]
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).