Severe Water Deficiency during the Mid-Vegetative and Reproductive Phase has Little Effect on Proso Millet Performance



Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Environmental Conditions
2.3. Plant Analyses
2.3.1. Leaf Morphological Properties
2.3.2. Leaf Biochemical Properties
2.3.3. Leaf Physiological Properties
2.3.4. Leaf Optical Properties
2.3.5. Leaf Relative Water Content
2.3.6. Leaf Bulk Element Analysis
2.4. Soil Analysis
2.4.1. Analysis of Plant-Available Si in the Soil
2.4.2. Soil Bulk Element Analysis
2.5. Statistical Analysis
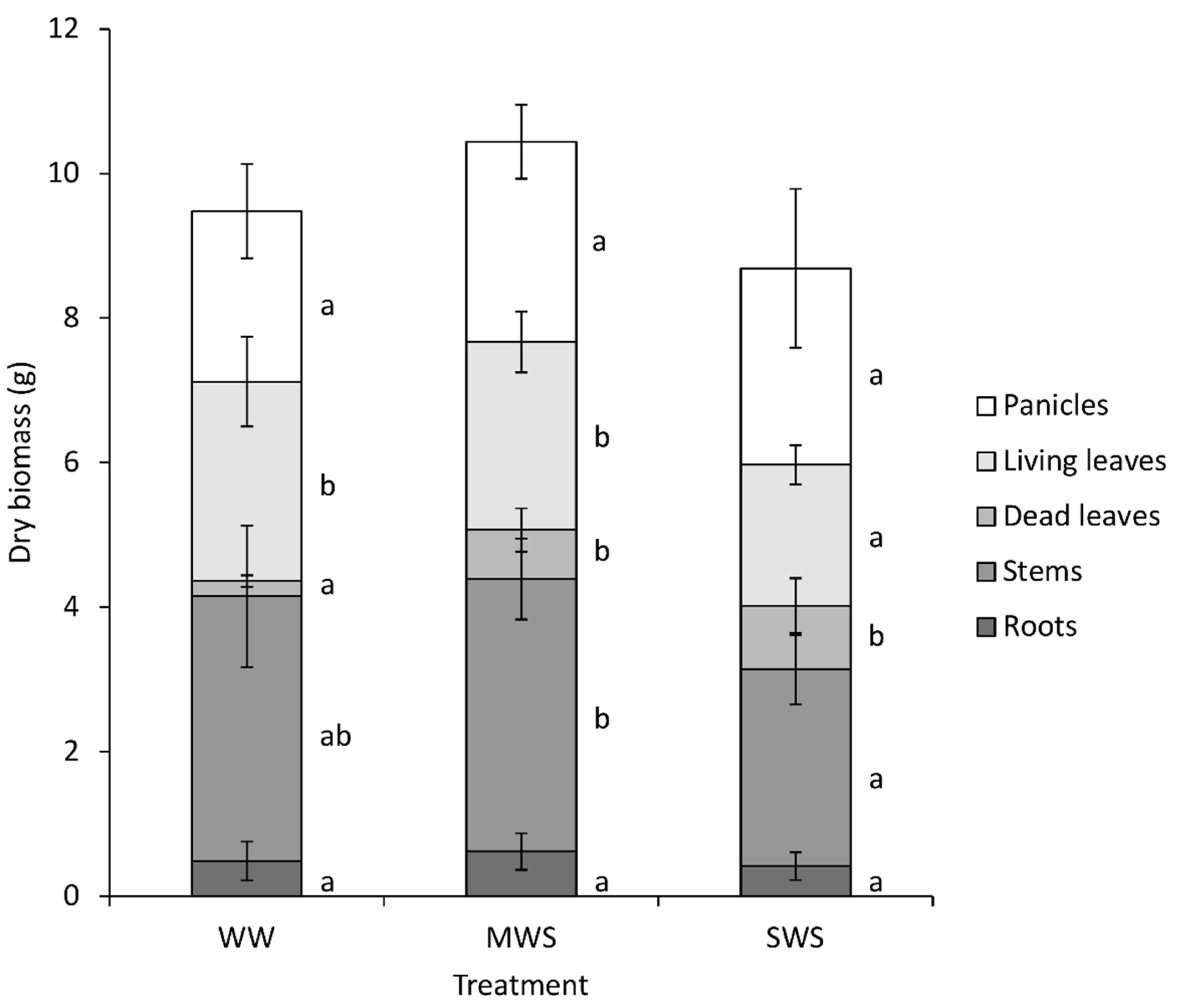
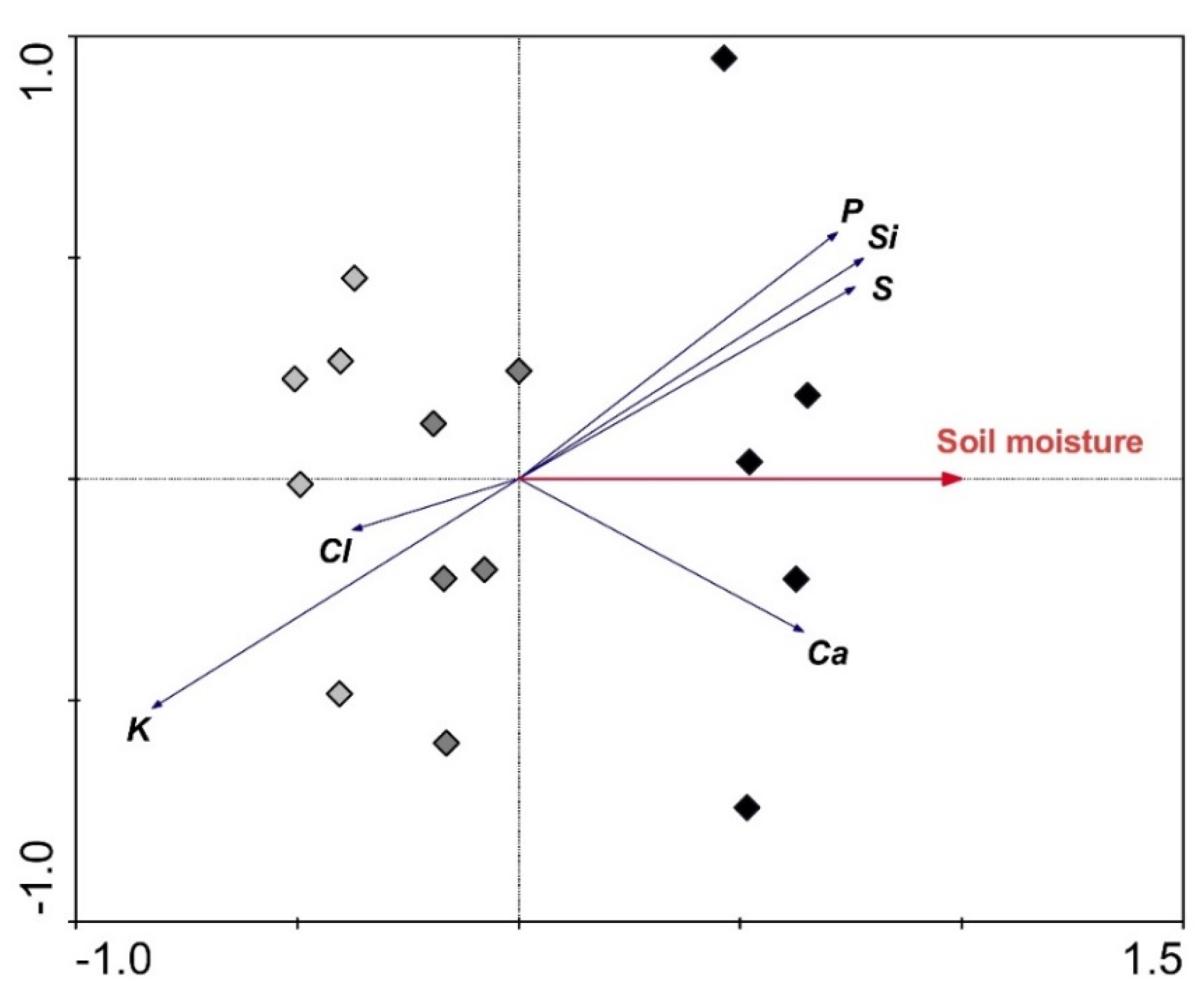
3. Results
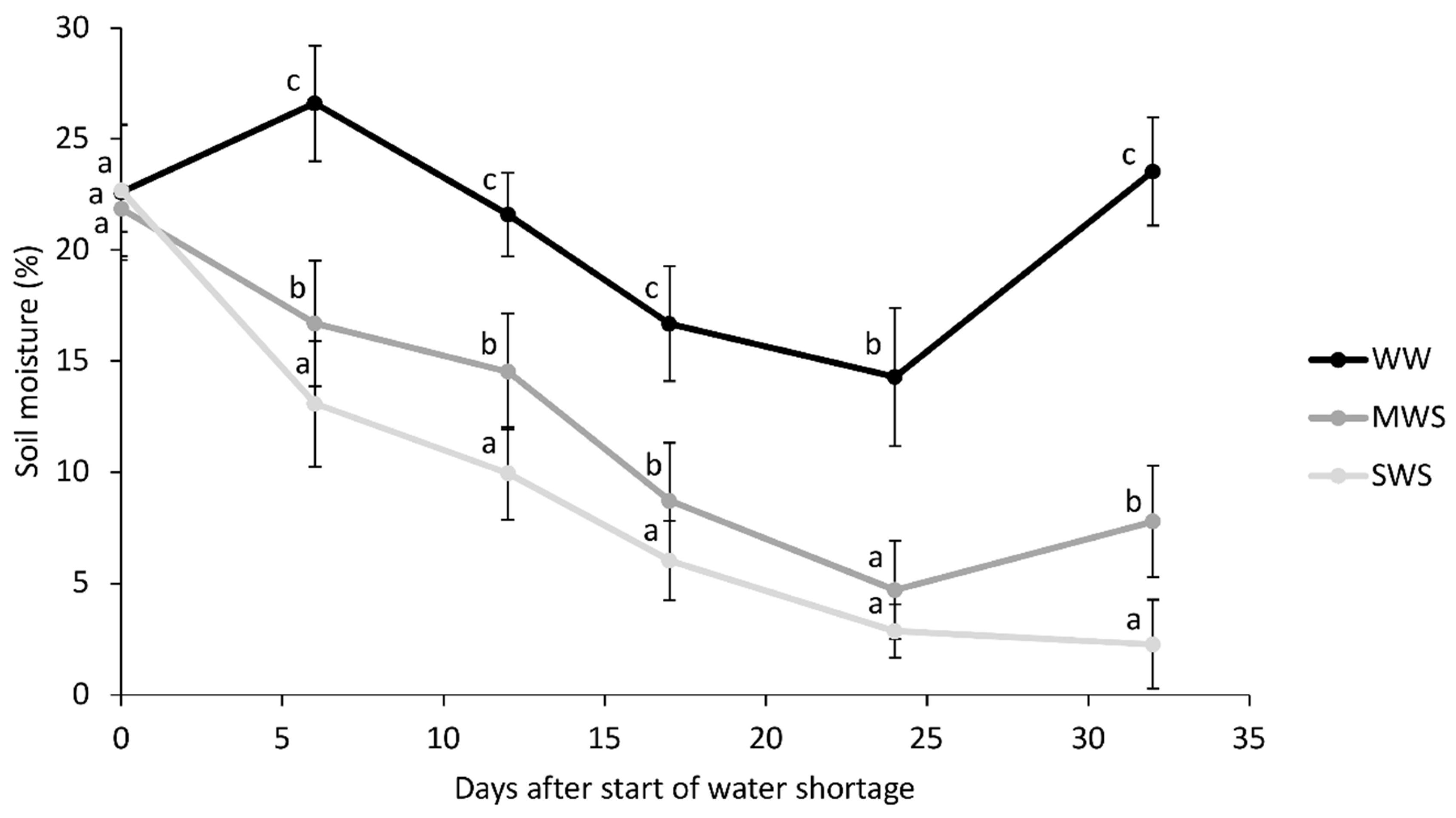
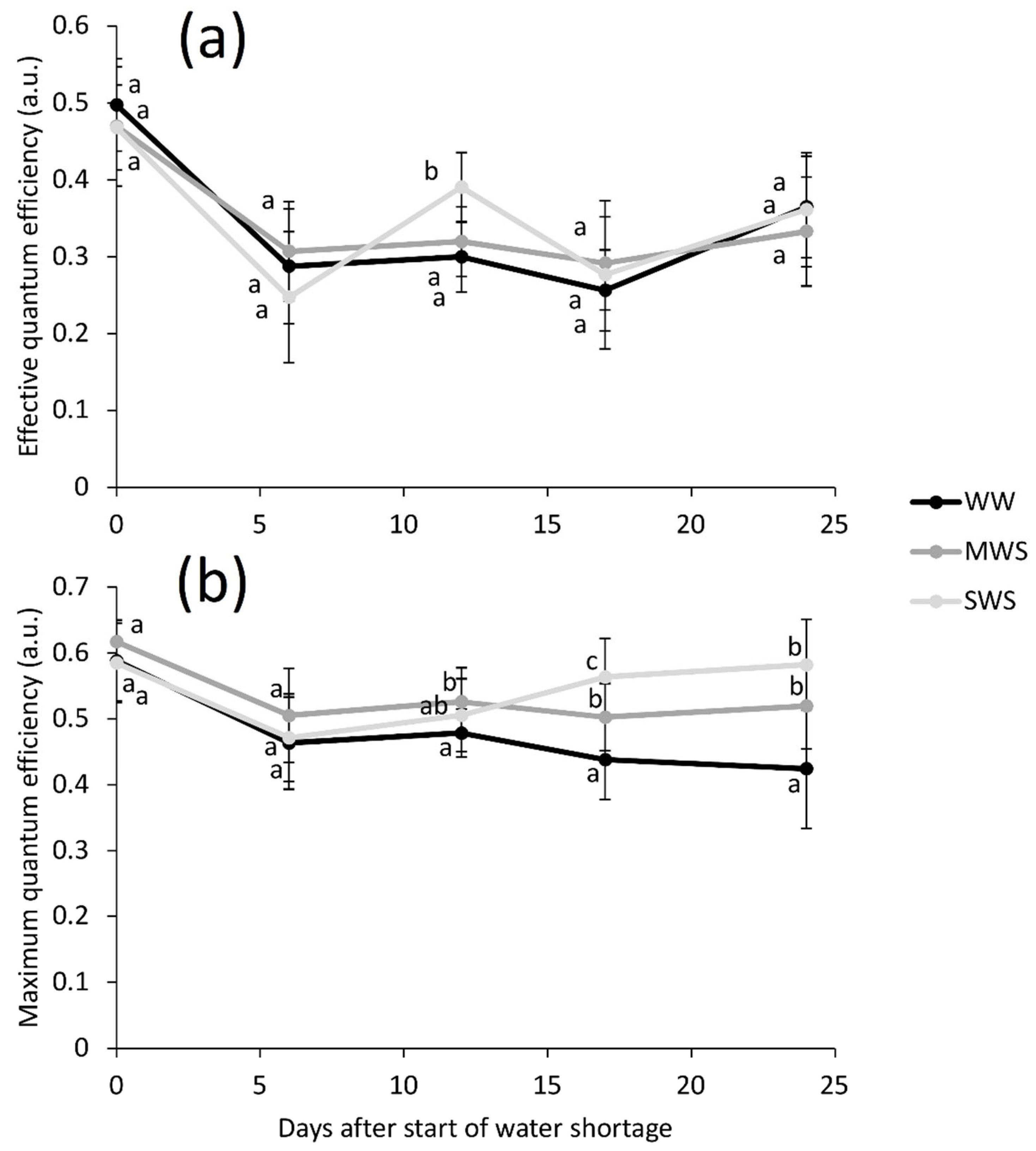
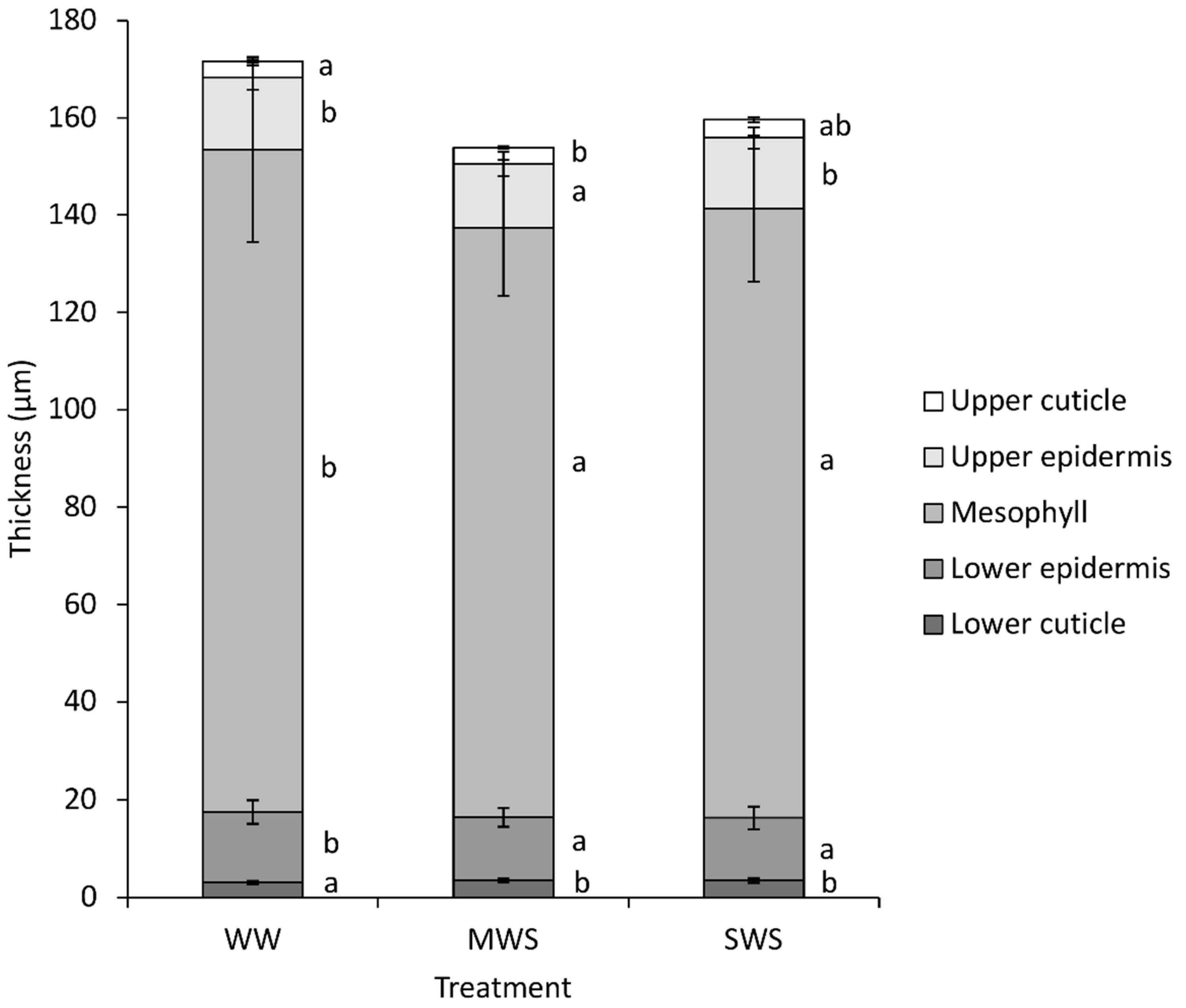
3.1. Environmental Conditions and Plant Parameters
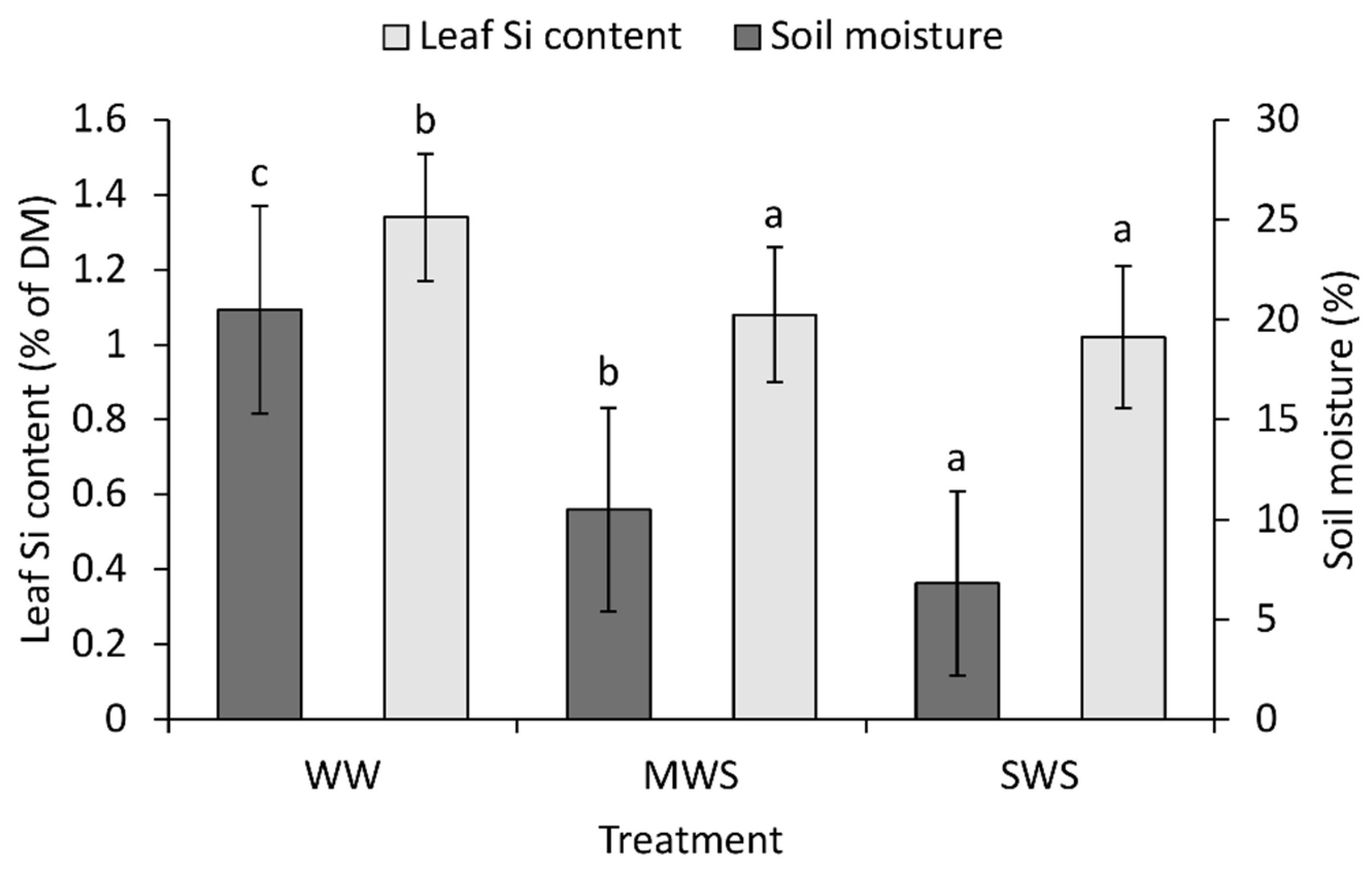
3.2. Soil Parameters
3.3. Relationships between Selected Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ray, R.L.; Fares, A.; Risch, E. Effects of drought on crop production and cropping areas in Texas. Agric. Environ. Lett. 2018, 3, 170037. [Google Scholar] [CrossRef]
- Water Scarcity and Drought in the European Union. Available online: http://ec.europa.eu/environment/pubs/pdf/factsheets/water_scarcity.pdf (accessed on 16 August 2019).
- Vanwindekens, F.M.; Gobin, A.; Curnel1, Y.; Planchon, V. New approach for mapping the vulnerability of agroecosystems based on expert knowledge. Math. Geosci. 2018, 50, 679–696. [Google Scholar] [CrossRef]
- Boyer, J.S. Plant productivity and environment. Science 1982, 218, 443–448. [Google Scholar] [CrossRef] [PubMed]
- Venuprasad, R.; Lafitte, H.; Atlin, G. Response to direct selection for grain yield under drought stress in rice. Crop Sci. 2007, 47, 285–293. [Google Scholar] [CrossRef]
- De Freitas, S.T.; Shackel, K.A.; Mitcham, E.J. Abscisic acid triggers whole-plant and fruit-specific mechanisms to increase fruit calcium uptake and prevent blossom end rot development in tomato fruit. J. Exp. Bot. 2011, 62, 2645–2656. [Google Scholar] [CrossRef]
- Li, Y.; Ye, W.; Wang, M.; Yan, X. Climate change and drought: A risk assessment of crop-yield impacts. Clim. Res. 2009, 39, 31–46. [Google Scholar] [CrossRef]
- Newton, A.C.; Flavell, A.J.; George, T.S.; Leat, P.; Mullholland, B.; Ramsay, L.; Revoredo-Giha, C.; Russell, J.; Steffenson, B.J.; Swanston, J.S.; et al. Crops that feed the world 4. Barley: A resilient crop? Strengths and weaknesses in the context of food security. Food Sec. 2011, 3, 141–178. [Google Scholar] [CrossRef]
- Saha, D.; Channabyre Gowda, M.V.; Arya, L.; Verma, M.; Bansal, K.C. Genetic and genomic resources of small millets. Crit. Rev. Plant Sci. 2016, 35, 56–79. [Google Scholar] [CrossRef]
- Wang, R.Y.; Wang, H.G.; Liu, X.Y.; Lian, S.; Chen, L.; Qiao, Z.J.; McInerney, C.E.; Wang, L. Drought-induced transcription of resistant and sensitive common millet varieties. J. Anim. Plant Sci. 2017, 27, 1303–1314. [Google Scholar]
- Kumar, A.; Tomer, V.; Kaur, A.; Kumar, V.; Gupta, K. Millets: A solution to agrarian and nutritional challenges. Agric. Food Sec. 2018, 7, 31. [Google Scholar] [CrossRef]
- Larcher, W. Physiological Plant Ecology: Ecophysiology and Stress Physiology of Functional Groups; Springer: Berlin, Germany, 2003. [Google Scholar]
- Petrov, P.; Petrova, A.; Dimitrov, I.; Tashev, T.; Olsovska, K.; Brestic, M.; Misheva, S. Relationships between leaf morpho-anatomy, water status and cell membrane stability in leaves of wheat seedlings subjected to severe soil drought. J. Agron. Crop Sci. 2018, 204, 219–227. [Google Scholar] [CrossRef]
- Kang, J.; Zhao, W.; Su, P.; Zhao, M.; Yang, Z. Sodium (Na+) and silicon (Si) coexistence promotes growth and enhances drought resistance of the succulent xerophyte Haloxylon ammodendron. Soil Sci. Plant Nutr. 2014, 60, 659–669. [Google Scholar] [CrossRef]
- Chen, D.; Wang, S.; Yin, L.; Deng, X. How does silicon mediate plant water uptake and loss under water deficiency? Front. Plant Sci. 2018, 9, 281. [Google Scholar] [CrossRef]
- Lux, A.; Luxová, M.; Hattori, T.; Inanaga, S.; Sugimoto, Y. Silicification in sorghum (Sorghum bicolor) cultivars with different drought tolerance. Physiol. Plant. 2002, 115, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Zou, C.; Wang, L.; Zhang, F. Silicon improves water use efficiency in maize plants. J. Plant Nutr. 2005, 27, 1457–1470. [Google Scholar] [CrossRef]
- Hattori, T.; Inanaga, S.; Tanimoto, E.; Lux, A.; Luxová, M.; Sugimoto, Y. Silicon-induced changes in viscoelastic properties of sorghum root cell walls. Plant Cell Physiol. 2003, 44, 743–749. [Google Scholar] [CrossRef] [PubMed]
- Saud, S.; Li, X.; Chen, Y.; Zhang, L.; Fahad, S.; Hussain, S.; Sadiq, A.; Chen, Y. Silicon application increases drought tolerance of Kentucky bluegrass by improving plant water relations and morphophysiological functions. Sci. World J. 2014, 2014, 368694. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.F.; Takahashi, E. Soil, Fertilizer, and Plant Silicon Research in Japan; Elsevier: Amsterdam, The Netherlands, 2002. [Google Scholar]
- Faisal, S.; Callis, K.L.; Slot, M.; Kitajima, K. Transpiration-dependent passive silica accumulation in cucumber (Cucumis sativus) under varying soil silicon availability. Botany 2012, 90, 1058–1064. [Google Scholar] [CrossRef]
- Grašič, M.; Malovrh, U.; Golob, A.; Vogel-Mikuš, K.; Gaberščik, A. Effects of water availability and UV radiation on silicon accumulation in the C4 crop proso millet. Photochem. Photobiol. Sci. 2019, 18, 375–386. [Google Scholar] [CrossRef]
- Grašič, M.; Dobravc, M.; Golob, A.; Vogel-Mikuš, K.; Gaberščik, A. Water shortage reduces silicon uptake in barley leaves. Agric. Water Manag. 2019, 217, 47–56. [Google Scholar] [CrossRef]
- Taylor, S.H.; Ripley, B.S.; Woodward, F.I.; Osborne, C.P. Drought limitation of photosynthesis differs between C3 and C4 grass species in a comparative experiment. Plant. Cell Environ. 2011, 34, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.F.; Miyake, Y.; Takahashi, E. Silicon as a beneficial element for crop plants. In Silicon in Agriculture; Datnoff, L.E., Snyder, G.H., Korndörfer, G.H., Eds.; Elsevier Science: Amsterdam, The Netherlands, 2001; pp. 17–39. [Google Scholar] [CrossRef]
- Kaufman, P.B.; Dayanandan, P.; Franklin, C.I.; Takeoka, Y. Structure and function of silica bodies in the epidermal system of grass shoots. Ann. Bot. 1985, 55, 487–507. [Google Scholar] [CrossRef]
- Lanning, F.C.; Eleuterius, L.N. Silica deposition in some C3 and C4 species of grasses, sedges and composites in the USA. Ann. Bot. 1989, 64, 395–410. [Google Scholar] [CrossRef]
- Klančnik, K.; Vogel-Mikuš, K.; Gaberščik, A. Silicified structures affect leaf optical properties in grasses and sedge. J. Photochem. Photobiol. B 2014, 130, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Klančnik, K.; Vogel-Mikuš, K.; Gaberščik, A. Leaf optical properties are affected by the location and type of deposited biominerals. J. Photochem. Photobiol. B 2014, 140, 276–285. [Google Scholar] [CrossRef]
- Ullah, S.; Schlerf, M.; Skidmore, A.K.; Hecker, C. Identifying plant species using mid-wave infrared (2.5–6 μm) and thermal infrared (8–14 μm) emissivity spectra. Remote Sens. Environ. 2012, 118, 95–102. [Google Scholar] [CrossRef]
- Noda, H.M.; Motohka, T.; Murakami, K.; Muraoka, H.; Nasahara, K.N. Accurate measurement of optical properties of narrow leaves and conifer needles with a typical integrating sphere and spectroradiometer. Plant Cell Environ. 2008, 36, 1903–1909. [Google Scholar] [CrossRef]
- Coops, N.C.; Stone, C. A comparison of field-based and modelled reflectance spectra from damaged Pinus radiata foliage. Aust. J. Bot. 2005, 53, 417–429. [Google Scholar] [CrossRef]
- Kovar, M.; Brestic, M.; Sytar, O.; Barek, V.; Hauptvogel, P.; Zivcak, M. Evaluation of Hyperspectral reflectance parameters to assess the leaf water content in soybean. Water 2019, 11, 443. [Google Scholar] [CrossRef]
- Castro-Esau, K.L.; Sanchez-Azofeifa, G.A.; Rivard, B.; Wright, S.J.; Quesada, M. Variability in leaf optical properties of Mesoamerican trees and the potential for species classification. Am. J. Bot. 2006, 93, 517–530. [Google Scholar] [CrossRef]
- Dim, J.R.; Kajiwara, K.; Honda, Y. Radiometric signature and spatial variability of the vegetation coverage of a boreal forest. Int. J. Remote Sens. 2008, 29, 6851–6871. [Google Scholar] [CrossRef]
- Havaux, M.; Lannoye, R. Chlorophyll fluorescence induction: A sensitive indicator of water stress in maize plants. Irrig. Sci. 1983, 4, 147–151. [Google Scholar] [CrossRef]
- Banks, J.M. Chlorophyll fluorescence as a tool to identify drought stress in Acer genotypes. Environ. Exp. Bot. 2018, 155, 118–127. [Google Scholar] [CrossRef]
- Guidi, L.; Lo Piccolo, E.; Mandi, M. Chlorophyll fluorescence, photoinhibition and abiotic stress: Does it make any difference the fact to be a C3 or C4 species? Front. Plant Sci. 2019, 10, 174. [Google Scholar] [CrossRef]
- Iqbal, N.; Hussain, S.; Raza, M.A.; Yang, C.Q.; Safdar, M.E.; Brestic, M.; Aziz, A.; Hayyat, M.S.; Asghar, M.A.; Wang, X.C.; et al. Drought tolerance of soybean (Glycine max L. Merr.) by improved photosynthetic characteristics and an efficient antioxidant enzyme activities under a split-root system. Front. Physiol. 2019, 10, 1–13. [Google Scholar] [CrossRef]
- Dąbrowski, P.; Baczewska-Dąbrowska, A.H.; Kalaji, H.M.; Goltsev, V.; Paunov, M.; Rapacz, M.; Wójcik-Jagła, M.; Pawluśkiewicz, B.; Bąba, W.; Brestic, M. Exploration of Chlorophyll a fluorescence and plant gas exchange parameters as indicators of drought tolerance in perennial ryegrass. Sensors 2019, 19, 2736. [Google Scholar] [CrossRef]
- Baltensperger, D.D. Progress with proso, pearl and other millets. In Trends in New Crops and New Uses; Janick, J., Whipkey, A., Eds.; ASHS Press: Alexandria, VA, USA, 2002; pp. 100–103. [Google Scholar]
- Shahidi, F.; Chandrasekara, A. Millet grain phenolics and their role in disease risk reduction and health promotion: A review. J. Funct. Foods 2013, 5, 570–581. [Google Scholar] [CrossRef]
- Kalinova, J.; Moudry, J. Content and quality of protein in proso millet (Panicum miliaceum L.) varieties. Plant Foods Hum. Nutr. 2006, 61, 45–49. [Google Scholar] [CrossRef]
- Upadhyaya, H.D.; Sharma, S.; Gowda, C.L.L.; Gopal Reddy, V.; Singh, S. Developing proso millet (Panicum miliaceum L.) core collection using geographic and morpho-agronomic data. Crop Pasture Sci. 2011, 62, 383–389. [Google Scholar] [CrossRef]
- Wang, R.; Hunt, H.V.; Qiao, Z.; Wang, L.; Han, Y. Diversity and cultivation of broomcorn millet (Panicum miliaceum L.) in China: A review. Econ. Bot. 2016, 70, 332–342. [Google Scholar] [CrossRef]
- Tadele, Z. Drought adaptation in millets. In Abiotic and Biotic Stress in Plants—Recent Advances and Future Perspectives; Shanker, A.K., Shanker, C., Eds.; IntechOpen: London, UK, 2016; pp. 639–662. [Google Scholar] [CrossRef]
- Gibson, D.J. Methods in Comparative Plant Population Ecology; Oxford University Press: Oxford, UK, 2015. [Google Scholar]
- Lichtenthaler, H.K.; Buschmann, C. Extraction of photosynthetic tissues: Chlorophylls and carotenoids. Curr. Protoc. Food Anal. Chem. 2001, 1, 165–170. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Buschmann, C. Chlorophylls and carotenoids: Measurement and characterization by UV-VIS spectroscopy. Curr. Protoc. Food Anal. Chem. 2001, 1, 171–178. [Google Scholar] [CrossRef]
- Drumm, H.; Mohr, H. The mode of interaction between blue (UV) light photoreceptor and phytochrome in anthocyanin formation of the Sorghum seedling. Photochem. Photobiol. 1978, 27, 241–248. [Google Scholar] [CrossRef]
- Caldwell, M.M. Solar ultraviolet radiation as an ecological factor for alpine plants. Ecol. Monogr. 1968, 38, 243–268. [Google Scholar] [CrossRef]
- Schreiber, U.; Kühl, M.; Klimant, I.; Reising, H. Measurement of chlorophyll fluorescence within leaves using a modified PAM fluorometer with a fiber-optic microprobe. Photosynth. Res. 1996, 47, 103–109. [Google Scholar] [CrossRef] [PubMed]
- González, L.; González-Vilar, M. Determination of relative water content. In Handbook of Plant Ecophysiology Techniques; Regiosa Roger, M.J., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2001; pp. 207–212. [Google Scholar]
- Nečemer, M.; Kump, P.; Ščančar, J.; Jaćimović, R.; Simčič, J.; Pelicon, P.; Budnar, M.; Jeran, Z.; Pongrac, P.; Regvar, M.; et al. Application of X-ray fluorescence analytical techniques in phytoremediation and plant biology studies. Spectrochim. Acta Part B At. Spectrosc. 2008, 63, 1240–1247. [Google Scholar] [CrossRef]
- Vekemans, B.; Janssens, K.; Vincze, L.; Adams, F.; Vanespen, P. Analysis of X-ray spectra by iterative least squares (AXIL): New developments. X-Ray Spectrom. 1994, 23, 278–285. [Google Scholar] [CrossRef]
- Kump, P.; Nečemer, M.; Rupnik, Z.; Pelicon, P.; Ponikvar, D.; Vogel-Mikuš, K.; Regvar, M.; Pongrac, P. Improvement of the XRF quantification and enhancement of the combined applications by EDXRF and micro-PIXE. In Integration of Nuclear Spectrometry Methods as a New Approach to Material Research; IAEA: Vienna, Austria, 2011; pp. 101–109. [Google Scholar]
- Korndörfer, G.H.; Coelho, M.N.; Snyder, G.H.; Mizutani, C.T. Evaluation of soil extractants for silicon availability in upland rice. R. Bras. Ci Solo 1999, 23, 101–106. [Google Scholar] [CrossRef]
- ter Braak, C.J.F.; Šmilauer, P. CANOCO Reference Manual and CanoDraw for Windows User’s Guide: Software for Canonical Community Ordination (Version 4.5); Microcomputer Power: Ithaca, NY, USA, 2002. [Google Scholar]
- Tao, G.-Q.; Letham, D.S.; Yong, J.W.H.; Zhang, K.; John, P.C.L.; Schwartz, O.; Chin Wong, S.; Farquhar, G.D. Promotion of shoot development and tuberisation in potato by expression of a chimaeric cytokinin synthesis gene at normal and elevated CO2 levels. Funct. Plant Biol. 2010, 37, 43–54. [Google Scholar] [CrossRef]
- Sharwood, R.E.; Sonawane, B.V.; Ghannoum, O. Photosynthetic flexibility in maize exposed to salinity and shade. J. Exp. Bot. 2014, 65, 3715–3724. [Google Scholar] [CrossRef]
- Lugojan, C.; Ciulca, S. Evaluation of relative water content in winter wheat. J. Hortic. Fores. Biotechnol. 2011, 15, 173–177. [Google Scholar]
- Wong, S.C.; Cowan, I.R.; Farquhar, G.D. Stomatal conductance correlates with photosynthetic capacity. Nature 1979, 282, 424–426. [Google Scholar] [CrossRef]
- Lawlor, D.W.; Cornic, G. Photosynthetic carbon assimilation and associated metabolism in relation to water deficits in higher plants. Plant Cell Environ. 2002, 25, 275–294. [Google Scholar] [CrossRef] [PubMed]
- Holaday, A.S.; Ritchie, S.W.; Nguyen, H.T. Effects of water deficit on gas-exchange parameters and ribulose 1,5-bisphosphate carboxylase activation in wheat. Environ. Exp. Bot. 1992, 32, 403–410. [Google Scholar] [CrossRef]
- Rivero, R.M.; Kojima, M.; Gepstein, A.; Sakakibara, H.; Mittler, R.; Gepstein, S.; Blumwald, E. Delayed leaf senescence induces extreme drought tolerance in a flowering plant. Proc. Natl. Acad. Sci. USA 2007, 104, 19631–19636. [Google Scholar] [CrossRef] [PubMed]
- Yong, J.W.H.; Letham, D.S.; Chin Wong, S.; Farquhar, G.D. Effects of root restriction on growth and associated cytokinin levels in cotton (Gossypium hirsutum). Funct. Plant Biol. 2010, 37, 974–984. [Google Scholar] [CrossRef]
- Björkman, O.; Demmig, B. Photon yield of O2 evolution and chlorophyll fluorescence characteristics at 77 K among vascular plants of diverse origins. Planta 1987, 170, 489–504. [Google Scholar] [CrossRef]
- Bi, H.; Kovalchuk, N.; Langridge, P.; Tricker, P.J.; Lopato, S.; Borisjuk, N. The impact of drought on wheat leaf cuticle properties. BMC Plant Biol. 2017, 17, 85. [Google Scholar] [CrossRef]
- Ma, J.F. Role of silicon in enhancing the resistance of plants to biotic and abiotic stresses. Soil Sci. Plant Nutr. 2004, 50, 11–18. [Google Scholar] [CrossRef]
- He, M.; Dijkstra, F.A. Drought effect on plant nitrogen and phosphorus: A meta-analysis. New Phytol. 2014, 204, 924–931. [Google Scholar] [CrossRef]
- Atkinson, R.R.L.; Mockford, E.J.; Bennett, C.; Christin, P.-A.; Spriggs, E.L.; Freckleton, R.P.; Thompson, K.; Rees, M.; Osborne, C.P. C4 photosynthesis boosts growth by altering physiology, allocation and size. Nat. Plants 2016, 2, 16038. [Google Scholar] [CrossRef] [PubMed]
- Angelo, C.L.; Pau, S. Root biomass and soil δ13C in C3 and C4 grasslands along a precipitation gradient. Plant Ecol. 2015, 216, 615–627. [Google Scholar] [CrossRef]
- Kostic, L.; Nikolic, N.; Bosnic, D.; Samardzic, J.; Nikolic, M. Silicon increases phosphorus (P) uptake by wheat under low P acid soil conditions. Plant Soil 2017, 419, 447–455. [Google Scholar] [CrossRef]
- Hepler, P.K. Calcium: A central regulator of plant growth and development. Plant Cell 2005, 17, 2142–2155. [Google Scholar] [CrossRef] [PubMed]
- Rouphael, Y.; Cardarelli, M.T.; Schwarz, D.; Franken, P.; Colla, G. Effects of drought on nutrient uptake and assimilation in vegetable crops. In Plant Responses to Drought Stress; Aroca, R., Ed.; Springer: Berlin, Germany, 2012; pp. 171–195. [Google Scholar] [CrossRef]
- Urbina, I.; Sardans, J.; Beierkuhnlein, C.; Jentsch, A.; Backhaus, S.; Grant, K.; Kreyling, J.; Peñuelasa, J. Shifts in the elemental composition of plants during a very severe drought. Env. Exp. Bot. 2015, 111, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Schilling, G.; Eißner, H.; Schmidt, L.; Peiter, E. Yield formation of five crop species under water shortage and differential potassium supply. J. Plant Nutr. Soil Sci. 2016, 179, 234–243. [Google Scholar] [CrossRef]
- Ceccato, P.; Flasse, S.; Tarantola, S.; Jacquemoud, S.; Gregoire, J.-M. Detecting vegetation leaf water content using reflectance in the optical domain. Remote Sens. Environ. 2001, 77, 22–33. [Google Scholar] [CrossRef]
- Kinzel, H. Calcium in the vacuoles and cell walls of plant tissue. Forms of deposition and their physiological and ecological significance. Flora 1989, 182, 99–125. [Google Scholar] [CrossRef]
- Gaffey, S.J. Spectral reflectance of carbonate minerals in the visible and near infrared (0.35–2.55 microns): Calcite, aragonite, and dolomite. Am. Mineral. 1986, 71, 151–162. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Sample Days | Parameter | Units | Treatment | ||
|---|---|---|---|---|---|---|
| WW | MWS | SWS– | ||||
| Morphological | 26 | Leaf thickness | µm | 171 ± 21 b | 155 ± 16 a | 159 ± 17 a |
| Specific leaf area | cm2 mg–1 | 0.306 ± 0.092 a | 0.305 ± 0.040 a | 0.302 ± 0.068 a | ||
| Upper surface stomata density | mm–2 | 105 ± 14 a | 115 ± 18 b | 110 ± 15 ab | ||
| Upper surface stomata length | µm | 31.1 ± 3.1 a | 31.2 ± 3.3 a | 31.3 ± 2.8 a | ||
| Lower surface stomata density | mm−2 | 87 ± 13 a | 91 ± 14 a | 89 ± 11 a | ||
| Lower surface stomata length | µm | 33.6 ± 3.3 a | 33.5 ± 2.7 a | 34.8 ± 3.2 a | ||
| Biochemical | 26 | Chlorophyll a | mg g–1 | 9.97 ± 3.23 a | 9.07 ± 1.81 a | 9.73 ± 1.88 a |
| Chlorophyll b | mg g–1 | 5.33 ± 1.96 a | 5.90 ± 2.27 a | 5.42 ± 1.75 a | ||
| Carotenoids | mg g–1 | 3.07 ± 1.05 a | 2.30 ± 0.53 a | 2.85 ± 0.50 a | ||
| Anthocyanins | a.u. g–1 | 179 ± 54 a | 239 ± 69 a | 188 ± 63 a | ||
| UV-B–absorbing compounds | a.u. g–1 | 493 ± 166 a | 479 ± 88 a | 534 ± 141 a | ||
| UV-A–absorbing compounds | a.u. g–1 | 738 ± 248 a | 702 ± 120 a | 770 ± 220 a | ||
| # Physiological | 6, 12, 17, 24/25 | Effective quantum efficiency | a.u. | 0.30 ± 0.07 a | 0.31 ± 0.06 a | 0.32 ± 0.10 a |
| Maximum quantum efficiency | a.u. | 0.45 ± 0.07 a | 0.51 ± 0.06 b | 0.53 ± 0.08 b | ||
| Stomatal conductance | mmol m–2 s–1 | 61 ± 28 c | 45 ± 21 b | 31 ± 15 a | ||
| Optical | 26 | UV-B | % | 10.45 ± 1.56 b | 8.33 ± 1.49 a | 8.39 ± 1.48 a |
| reflectance | UV-A | % | 7.85 ± 1.23 b | 6.88 ± 0.93 a | 6.80 ± 0.94 a | |
| Violet | % | 6.59 ± 0.45 ab | 6.34 ± 0.39 a | 6.80 ± 0.35 b | ||
| Blue | % | 6.53 ± 0.38 a | 6.54 ± 0.32 a | 7.02 ± 0.32 b | ||
| Green | % | 12.05 ± 2.82 a | 12.54 ± 2.77 a | 13.72 ± 3.25 a | ||
| Yellow | % | 10.42 ± 1.21 a | 10.93 ± 0.93 a,b | 11.99 ± 1.34 b | ||
| Red | % | 8.05 ± 1.31 a | 8.46 ± 1.31 a,b | 9.07 ± 1.62 b | ||
| Near infrared | % | 31.64 ± 4.94 a | 32.10 ± 4.77 a | 34.19 ± 5.74 a | ||
| Optical | 26 | UV-B | % | 0.21 ± 0.49 b | 0.03 ± 0.41 a | 0.15 ± 0.45 b |
| transmittance | UV-A | % | 0.63 ± 0.87 a | 0.58 ± 0.88 a | 0.81 ± 1.16 a | |
| Violet | % | 4.31 ± 0.77 a | 4.17 ± 0.73 a | 5.56 ± 1.07 b | ||
| Blue | % | 6.22 ± 0.82 a | 6.15 ± 0.82 a | 7.83 ± 1.24 b | ||
| Green | % | 22.05 ± 6.75 a | 21.97 ± 6.46 a | 22.72 ± 6.23 a | ||
| Yellow | % | 19.76 ± 2.33 a | 19.53 ± 1.87 a | 20.90 ± 2.59 a | ||
| Red | % | 12.40 ± 4.08 a | 12.58 ± 4.18 a | 14.09 ± 4.30 a | ||
| Near infrared | % | 65.31 ± 12.00 b | 63.92 ± 11.58 a,b | 60.45 ± 10.43 a | ||
| Element analysis | 32 | P | % of dry matter | 0.59 ± 0.14 b | 0.40 ± 0.08 a | 0.43 ± 0.07 a |
| S | % of dry matter | 0.24 ± 0.05 b | 0.17 ± 0.04 a | 0.17 ± 0.01 a | ||
| Cl | % of dry matter | 0.63 ± 0.06 a,b | 0.60 ± 0.06 a | 0.81 ± 0.22 b | ||
| K | % of dry matter | 3.70 ± 0.27 a | 3.76 ± 0.15 a | 4.18 ± 0.29 b | ||
| Ca | % of dry matter | 0.55 ± 0.08 b | 0.52 ± 0.06 a,b | 0.45 ± 0.04 a | ||
| Other | 27 | Leaf relative water content | % | 94.8 ± 2.9 b | 94.8 ± 0.9 b | 87.4 ± 8.4 a |
| 31 | Plant height | cm | 94.5 ± 8.2 c | 85.9 ± 9.1 b | 74.3 ± 5.4 a | |
| 31 | Lodged plants | % | 66.6 ± 12.0 b | 50.1 ± 2.9 a | 55.1 ± 14.3 a,b | |
| Element | Sample Days | Units | Before | After |
|---|---|---|---|---|
| Total Si | −1, 32 | % of DM | 6.65 ± 0.66 | 6.45 ± 0.78 |
| Total Ca | 3.70 ± 0.39 | 3.77 ± 0.23 |
| Element | Leaf Reflectance | |||||||
|---|---|---|---|---|---|---|---|---|
| UV-B | UV-A | Violet | Blue | Green | Yellow | Red | Near Infrared | |
| Si | 0.666 ** | 0.614 * | 0.030 | −0.093 | −0.031 | −0.032 | −0.089 | 0.016 |
| Ca | 0.300 | 0.214 | −0.715 ** | −0.848 ** | −0.612 * | −0.663 ** | −0.775 ** | −0.371 |
| Water-Related Parameters | Leaf Element Contents | |||||
|---|---|---|---|---|---|---|
| Si | P | S | Cl | K | Ca | |
| Soil moisture | 0.691 ** | 0.648 ** | 0.654 ** | −0.429 | −0.516 * | 0.580 * |
| Stomatal conductance | 0.549 * | 0.567 * | 0.484 | −0.487 | −0.715 ** | 0.341 |
| Leaf relative water content | 0.290 | 0.300 | 0.284 | −0.740 ** | −0.757 ** | 0.229 |
| Parameters | Plant Height | Living Leaf Biomass | Dead Leaf Biomass |
|---|---|---|---|
| Soil moisture | 0.834 ** | 0.603 * | −0.770 ** |
| Stomatal conductance | 0.660 ** | 0.429 | −0.612 * |
| Leaf relative water content | 0.623 * | 0.517 * | −0.627 * |
| Leaf Si content | 0.539 * | 0.743 ** | −0.449 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grašič, M.; Golob, A.; Vogel-Mikuš, K.; Gaberščik, A. Severe Water Deficiency during the Mid-Vegetative and Reproductive Phase has Little Effect on Proso Millet Performance. Water 2019, 11, 2155. https://doi.org/10.3390/w11102155
Grašič M, Golob A, Vogel-Mikuš K, Gaberščik A. Severe Water Deficiency during the Mid-Vegetative and Reproductive Phase has Little Effect on Proso Millet Performance. Water. 2019; 11(10):2155. https://doi.org/10.3390/w11102155
Chicago/Turabian StyleGrašič, Mateja, Aleksandra Golob, Katarina Vogel-Mikuš, and Alenka Gaberščik. 2019. "Severe Water Deficiency during the Mid-Vegetative and Reproductive Phase has Little Effect on Proso Millet Performance" Water 11, no. 10: 2155. https://doi.org/10.3390/w11102155
APA StyleGrašič, M., Golob, A., Vogel-Mikuš, K., & Gaberščik, A. (2019). Severe Water Deficiency during the Mid-Vegetative and Reproductive Phase has Little Effect on Proso Millet Performance. Water, 11(10), 2155. https://doi.org/10.3390/w11102155