Direct and Indirect Impacts of Fish on Crustacean Zooplankton in Experimental Mesocosms


 ,
,  ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
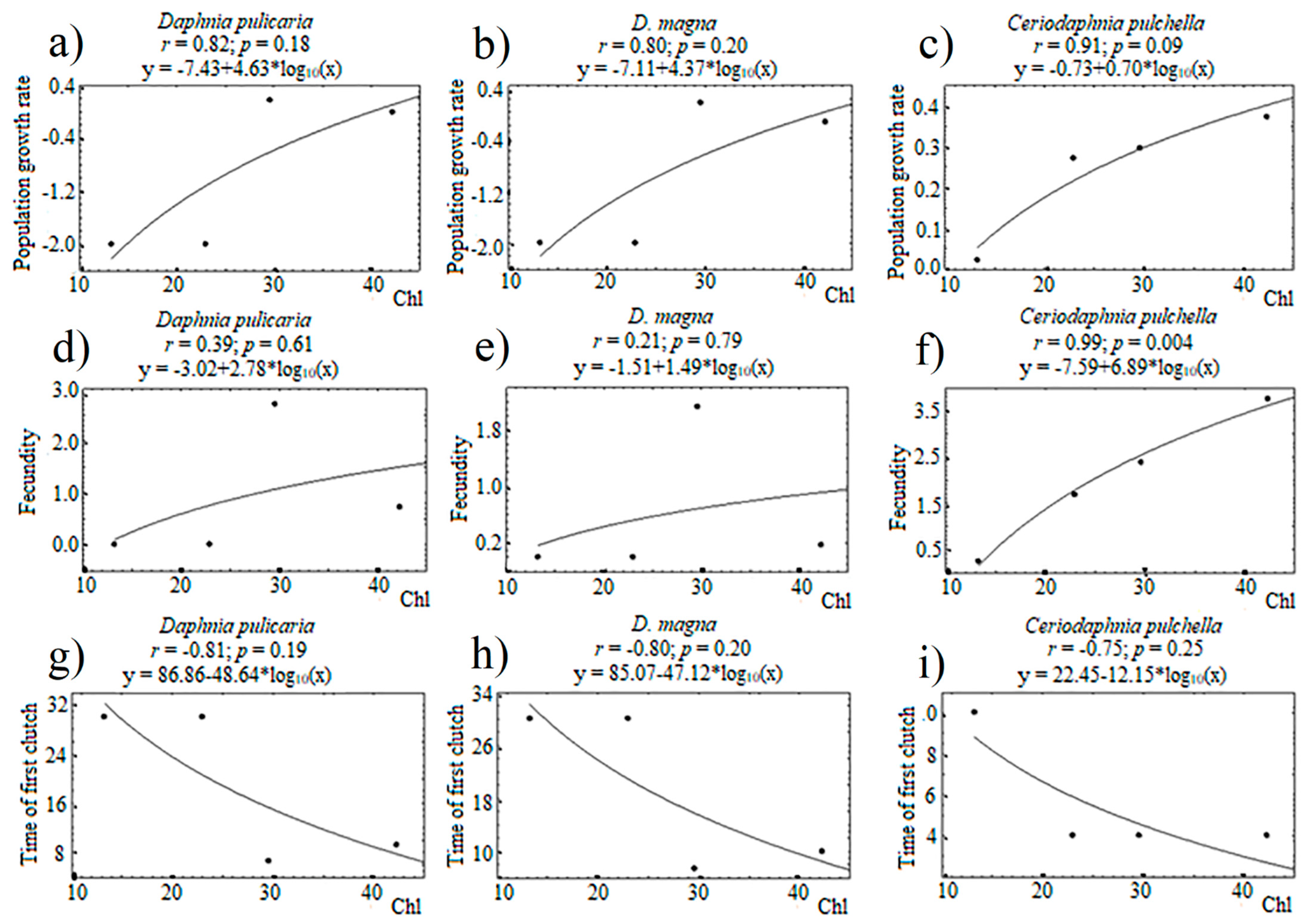
3. Results
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Prater, C.; Wagner, N.D.; Frost, P.C. Seasonal effects of food quality and temperature on body stoichiometry, biochemistry, and biomass production in Daphnia populations. Limnol. Oceanogr. 2018, 63, 1727–1740. [Google Scholar] [CrossRef]
- Sterner, R.W. Daphnia growth on varying quality of Scenedesmus: Mineral limitation of zooplankton. Ecology 1993, 74, 2351–2360. [Google Scholar] [CrossRef]
- Sterner, R.W.; Elser, J.J. Ecological Stoichiometry: The Biology of Elements from Molecules to the Biosphere, 1st ed.; Princeton University Press: Princeton, NJ, USA, 2002; ISSN 0-691-07491-7. [Google Scholar]
- Vanderploeg, H.A.; Nalepa, T.F.; Jude, D.J.; Mills, E.L.; Holeck, K.T.; Liebig, J.R.; Grigorovich, I.A.; Ojaveer, H. Dispersal and emerging ecological impacts of Ponto-Caspian species in the Laurentian Great Lakes. Can. J. Fish. Aquat. Sci. 2002, 59, 1209–1228. [Google Scholar] [CrossRef]
- Sundbom, M.; Vrede, T. Effects of fatty acids and phosphorus content of food on the growth, survival and reproduction of Daphnia. Freshw. Biol. 1997, 38, 665–674. [Google Scholar] [CrossRef]
- Gulati, R.D.; DeMott, W.R. The role of food quality for zooplankton: Remarks on the state-of-the-art, perspectives and priorities. Freshw. Biol. 1997, 38, 753–768. [Google Scholar] [CrossRef]
- Elser, J.J.; Urabe, J. The stoichiometry of consumer-driven nutrient recycling: Theory, observations, and consequences. Ecology 1999, 80, 735–751. [Google Scholar] [CrossRef]
- Ravet, J.L.; Persson, J.; Brett, M.T. Threshold dietary polyunsaturated fatty acid concentrations for Daphnia pulex growth and reproduction. Inland Waters 2012, 2, 199–209. [Google Scholar] [CrossRef]
- Makhutova, O.N.; Gladyshev, M.I.; Sushchik, N.N.; Dubovskaya, O.P.; Buseva, Z.F.; Fefilova, E.B.; Semenchenko, V.P.; Kalachova, G.S.; Kononova, O.N.; Baturina, M.A. Comparison of fatty acid composition of cladocerans and copepods from lakes of different climatic zones. Contemp. Probl. Ecol. 2014, 7, 474–483. [Google Scholar] [CrossRef]
- Gladyshev, M.I.; Sushchik, N.N.; Dubovskaya, O.P.; Buseva, Z.F.; Makhutova, O.N.; Fefilova, E.B.; Feniova, I.Y.; Semenchenko, V.P.; Kolmakova, A.A.; Kalachova, G.S.; et al. Fatty acid composition of Cladocera and Copepoda from lakes of contrasting temperature. Freshw. Biol. 2015, 60, 373–386. [Google Scholar] [CrossRef]
- Hart, R.C.; Bychek, E.A. Body size in freshwater planktonic crustaceans: An overview of extrinsic determinants and modifying influences of biotic interactions. Hydrobiologia 2011, 668, 61–108. [Google Scholar] [CrossRef]
- Achenbach, L.; Lampert, W. Effects of elevated temperatures on threshold food concentrations and possible competitive abilities of differently sized cladoceran species. Oikos 1997, 79, 469–476. [Google Scholar] [CrossRef]
- Gliwicz, Z.M. Food thresholds and body size in cladocerans. Nature 1990, 343, 638–640. [Google Scholar] [CrossRef]
- Gliwicz, Z.M. Between Hazards of Starvation and Risk of Predation: The Ecology of Off-Shore Animals Excellence in Ecology, Book 12; International Ecology Institute: Oldendorf/Luhe, Germany, 2003; p. 379. ISSN 0932-2205. [Google Scholar]
- Gliwicz, Z.M.; Lampert, W. Food thresholds in Daphnia species in the absence and presence of blue-green filaments. Ecology 1990, 71, 691–702. [Google Scholar] [CrossRef]
- Sterner, R.W.; Schulz, K.L. Zooplankton nutrition: recent progress and a reality check. Aquat. Ecol. 1998, 32, 261–279. [Google Scholar] [CrossRef]
- Sikora, A.; Dawidowicz, P.; von Elert, E. Daphnia fed algal food grown at elevated temperature have reduced fitness. J. Limnol. 2014, 73, 421–427. [Google Scholar] [CrossRef]
- Sikora, A.; Petzoldt, T.; Dawidowicz, P.; von Elert, E. Demands of eicosapentaenoic acid (EPA) in Daphnia: are they dependent on body size? Oecologia 2016, 182, 405–417. [Google Scholar] [CrossRef] [PubMed]
- Müller-Navarra, D.C.; Brett, M.T.; Liston, A.M.; Goldman, C.R. A highly unsaturated fatty acid predicts carbon transfer between primary producers and consumers. Nature 2000, 403, 74–77. [Google Scholar] [CrossRef]
- Wilson, A.E. Effects of zebra mussels on phytoplankton and ciliates: A field mesocosm experiment. J. Plankton Res. 2003, 25, 905–915. [Google Scholar] [CrossRef]
- Sommer, U.; Sommer, F. Cladocerans versus copepods: the cause of contrasting top-down controls on freshwater and marine phytoplankton. Oecologia 2006, 147, 183–194. [Google Scholar] [CrossRef]
- Maia-Barbosa, P.M.; Matsumura-Tundisi, T.M. Consumption of zooplanktonic organisms by Astyanax fasciatus Cuvier, 1819 (Osteichthyes, Characidae) in Lobo (Broa) reservoir, São Carlos, SP, Brazil. Hydrobiologia 1984, 113, 171–181. [Google Scholar] [CrossRef]
- Sommer, U.; Stibor, H. Copepoda–Cladocera–Tunicata: The role of three major mesozooplankton groups in pelagic food webs. Ecol. Res. 2002, 17, 161–174. [Google Scholar] [CrossRef]
- Güntzel, A.M.; Morita Melo, I.K.; Roche, K.F.; da Silva, V.F.B.; Pompiani, P.G. Cladocerans from gut contents of fishes associated to macrophytes from Taquari River Basin, MS, Brazil. Acta Limnol. Bras. 2012, 24, 97–102. [Google Scholar] [CrossRef]
- Bohl, E. Food supply and prey selection in planktivorous Cyprinidae. Oecologia 1982, 35, 134–138. [Google Scholar] [CrossRef] [PubMed]
- Okun, N.; Mehner, T. Distribution and feeding of juvenile fish on invertebrates in littoral reed (Phragmites) stands. Ecol. Freshw. Fish. 2005, 14, 139–149. [Google Scholar] [CrossRef]
- Brooks, J.L.; Dodson, S.I. Predation, body size and composition of plankton. Science 1965, 150, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Semenchenko, V.P.; Razlutskij, V.I.; Feniova, I.Y.; Aibulatov, D.N. Biotic relations affecting species structure in zooplankton communities. Hydrobiologia 2007, 579, 219–231. [Google Scholar] [CrossRef]
- Brabrand, A.; Bjorn, F.; Torsten, K.; Nilssen, P.J. Can iron defecation from fish influence phytoplankton production and biomass in eutrophic lakes? Limnol. Oceanogr. 1984, 29, 1330–1334. [Google Scholar] [CrossRef]
- Kolmakov, V.I.; Gladyshev, M.I. Growth and potential photosynthesis of cyanobacteria are stimulated by viable gut passage in crucian carp. Aquat. Ecol. 2003, 37, 237–242. [Google Scholar] [CrossRef]
- Kolmakov, V.I. Role of Microcystis aeruginosa passing through the digestive tracts of filter-feeding animals in eutrophic water reservoirs (review). Contemp. Probl. Ecol. 2014, 7, 455–464. [Google Scholar] [CrossRef]
- Gliwicz, Z.M.; Ghilarov, A.; Pijanowska, J. Food and predation as major factors limiting two natural populations of Daphnia cucullata Sars. Hydrobiologia 1981, 80, 205–218. [Google Scholar] [CrossRef]
- Feniova, I.; Dawidowicz, P.; Gladyshev, M.I.; Kostrzewska-Szlakowska, I.; Rzepecki, M.; Razlutskij, V.; Sushchik, N.N.; Majsak, N.; Dzialowski, A.R. Experimental effects of large-bodied Daphnia, fish and zebra mussels on cladoceran community and size structure. J. Plankton Res. 2015, 37, 611–625. [Google Scholar] [CrossRef]
- Błędzki, L.A.; Rybak, J.I. Freshwater Crustacean Zooplankton of Europe; Springer: Basel, Switzerland, 2016; p. 918. [Google Scholar] [CrossRef]
- Kuzmin, G.V. Phytoplankton. Methods of Ecosystem Studies in Inland Waters; Nauka: Moscow, USSR, 1975; pp. 73–87. [Google Scholar]
- Vinberg, G.G.; Lavrenteva, G.M. Guidelines for the Collection and Processing of Materials for Hydrobiological Studies in Freshwater Bodies. Phytoplankton and Its Products; GOSNIORKH: Leningrad, USSR, 1982; pp. 1–33. [Google Scholar]
- Mikheeva, T.M. Methods of quantitative estimates of nanophytoplankton (review). Hydrobiol. J. 1989, 25, 3–21. [Google Scholar]
- APHA. Standard Methods. Standard Methods for the Examination of Water and Wastewater, 21st ed.; American Public Health Association: Washington, DC, USA, 2005. [Google Scholar]
- Gladyshev, M.I.; Sushchik, N.N.; Kolmakova, A.A.; Kalachova, G.S.; Kravchuk, E.S.; Ivanova, E.A.; Makhutova, O.N. Seasonal correlations of elemental and ω3 PUFA composition of seston and dominant phytoplankton species in a eutrophic Siberian Reservoir. Aquat. Ecol. 2007, 41, 9–23. [Google Scholar] [CrossRef]
- Murphy, J.; Riley, J.P. A modified single solution method for the determination of phosphate in natural waters. Anal. Chim. Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Sushchik, N.N.; Gladyshev, M.I.; Kalachova, G.S.; Makhutova, O.N.; Ageev, A.V. Comparison of seasonal dynamics of the essential PUFA contents in benthic invertebrates and grayling Thymallus arcticus in the Yenisei river. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2006, 145, 278–287. [Google Scholar] [CrossRef] [PubMed]
- Gladyshev, M.I.; Semenchenko, V.P.; Dubovskaya, O.P.; Fefilova, E.B.; Makhutova, O.N.; Buseva, Z.F. Effect of temperature on contents of essential highly unsaturated fatty acids in freshwater zooplankton. Limnologica 2011, 41, 339–347. [Google Scholar] [CrossRef]
- Porter, K.G.; Feig, Y.S.; Vetter, E.F. Morphology, flow regimes, and filtering rates of Daphnia, Ceriodaphnia, and Bosmina fed natural bacteria. Oecologia 1983, 58, 156–163. [Google Scholar] [CrossRef]
- Pijanowska, J.; Dawidowicz, P.; Howe, A.; Weider, L.J. Predator-induced shifts in Daphnia life-histories under different food regimes. Arch. Hydrobiol. 2006, 167, 37–54. [Google Scholar] [CrossRef]
- DeMott, W.R.; Gulati, R.D.; Siewertsen, K. Effects of phosphorus-deficient diets on the carbon and phosphorus balance of Daphnia magna. Limnol. Oceanogr. 1998, 43, 1147–1161. [Google Scholar] [CrossRef]
- Brett, M.T.; Müller-Navarra, D.C.; Park, S.K. Empirical analysis of the effect of phosphorus limitation on algal food quality for freshwater zooplankton Limnol. Oceanogr. 2000, 45, 1564–1575. [Google Scholar] [CrossRef]
- Hall, S.R.; Leibold, M.A.; Lytle, D.A.; Smith, V.H. Stoichiometry and planktonic grazer composition over gradients of light, nutrients, and predation risk. Ecology 2004, 85, 2291–2301. [Google Scholar] [CrossRef][Green Version]
- Tilman, D. Niche tradeoffs, neutrality, and community structure: a stochastic theory of resource competition, invasion, and community assembly. PNAS 2004, 101, 10854–10861. [Google Scholar] [CrossRef] [PubMed]
- Ahlgren, G.; Goedkoop, W.; Markensten, H.; Sonesten, L.; Boberg, M. Seasonal variations in food quality for pelagic and benthic invertebrates in Lake Erken—The role of fatty acids. Freshw. Biol. 1997, 38, 555–570. [Google Scholar] [CrossRef]
- DeMott, W.R. The role of taste in food selection by freshwater zooplankton. Oecologia 1986, 69, 334–340. [Google Scholar] [CrossRef] [PubMed]
- Polishchuk, L.V.; Vijverberg, J. Contribution analysis of body mass dynamics in Daphnia. Oecologia 2005, 144, 268–277. [Google Scholar] [CrossRef]
- Gladyshev, M.I.; Sushchik, N.N.; Dubovskaya, O.P.; Makhutova, O.N.; Kalachova, G.S. Growth rate of Daphnia feeding on seston in a Siberian reservoir: the role of essential fatty acid. Aquat. Ecol. 2008, 42, 617–627. [Google Scholar] [CrossRef]
- Taipale, S.J.; Kainz, M.J.; Brett, M.T. Diet-switching experiments show rapid accumulation and preferential retention of highly unsaturated fatty acids in Daphnia. Oikos 2011, 120, 1674–1682. [Google Scholar] [CrossRef]
- Masclaux, H.; Bec, A.; Kainz, M.J.; Perriere, F.; Desvilettes, C.; Bourdier, G. Accumulation of polyunsaturated fatty acids by cladocerans: Effects of taxonomy, temperature and food. Freshw. Biol. 2012, 57, 696–703. [Google Scholar] [CrossRef]
- Hartwich, M.; Martin-Creuzburg, D.; Wacker, A. Seasonal changes in the accumulation of polyunsaturated fatty acids in zooplankton. J. Plankton Res. 2013, 35, 121–134. [Google Scholar] [CrossRef]
- Koussoroplis, A.M.; Kainz, M.J.; Striebel, M. Fatty acid retention under temporally heterogeneous dietary intake in a cladoceran. Oikos 2013, 122, 1017–1026. [Google Scholar] [CrossRef]
- Taipale, S.J.; Vuorio, K.; Brett, M.T.; Peltomaa, E.; Hiltunen, M.; Kankaala, P. Lake zooplankton δ13C values are strongly correlated with the δ13C values of distinct phytoplankton taxa. Ecosphere 2016, 7, e01392. [Google Scholar] [CrossRef]
- Gladyshev, M.I.; Kolmakov, V.I.; Dubovskaya, O.P.; Ivanova, E.A. Studying of algae food composition of Daphnia longispina during blue-green bloom of eutrophic pond. Dokl. Akad. Nauk. 2000, 371, 556–558. [Google Scholar]
- Weidman, P.R.; Schindler, D.W.; Thompson, P.L.; Vinebrooke, R.D. Interactive effects of higher temperature and dissolved organic carbon on planktonic communities in fishless mountain lakes. Freshw. Biol. 2014, 59, 889–904. [Google Scholar] [CrossRef]
- Ramenski, L.G. Introduction to the Complex Soil-Geobotanical Investigation of Lands; Selkhozgiz: Moscow, USSR, 1938; p. 620. (In Russian) [Google Scholar]
- Grime, J.P. Plant Strategis, Vegetation Processes, and Ecosystem Properties; John Wiley & Sons Ltd.: Chichester, UK, 2001; p. 456. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mi | MFi | Ei | EFi | Mf | MFf | Ef | EFf | F(H) | P | |
|---|---|---|---|---|---|---|---|---|---|---|
| C:N (mg/mg) | 7.16 ± 0.25 | 6.71 ± 0.28 | 8.18 ± 0.33 | 7.54 ± 0.19 | 7.39 ± 2.12 | 6.96 ± 0.16 | 8.29 ± 0.52 | 6.79 ± 0.16 | 12.4 | 0.09 |
| C:P (mg/mg) | 136.6 ± 15.2 | 129.0 ± 5.2 | 182.5 ± 10.6 | 150.1 ± 12.9 | 173.9 ± 103.1 | 156.7 ± 36.1 | 68.5 ± 3.5 | 287.6 ± 67.3 | 12 | 0.10 |
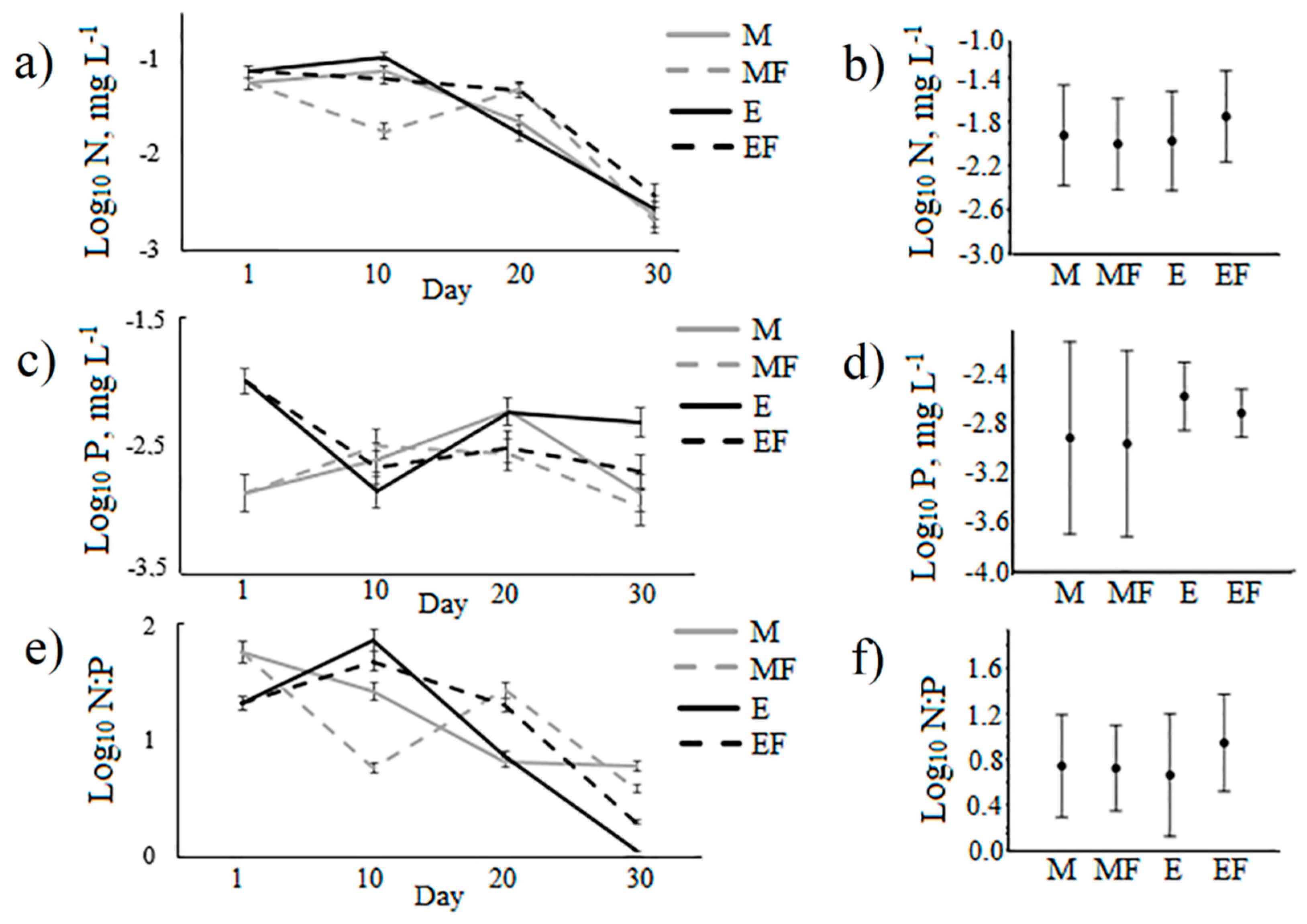
| N:P (mg/mg) | 19.0 ± 1.5ab | 19.2 ± 0.5ab | 22.8 ± 1.9b | 19.9 ± 1.3ab | 21.6 ± 7.0ab | 22.4 ± 4.9ab | 8.3 ± 0.8a | 42.0 ± 9.2c | 4.0 | 0.01 |
| Treatment | EPA, µg/L | DHA, µg/L | Total FA, µg/L |
|---|---|---|---|
| Mi | 3.78 ± 0.34A | 4.02 ± 0.49A | 54.86 ± 6.94AC |
| MFi | 4.47 ± 0.51A | 3.03 ± 0.90A | 54.55 ± 6.04AC |
| Ei | 4.61 ± 0.24A | 4.12 ± 0.11A | 101.07 ± 11.74BD |
| EFi | 4.93 ± 0.48A | 4.10 ± 0.26A | 113.06 ± 9.89B |
| Mf | 0.71 ± 0.24B | 0.08 ± 0.05B | 15.45 ± 3.12A |
| MFf | 1.56 ± 0.48B | 0.52 ± 0.39B | 29.51 ± 4.02AC |
| Ef | 0.52 ± 0.05B | 0.05 ± 0.05B | 16.76 ± 0.94A |
| EFf | 1.60 ± 0.42B | 0.89 ± 0.65B | 68.85 ± 13.92CD |
| Treatment | EPA:C, mg/g | DHA:C, mg/g | Total FA:C, mg/g |
|---|---|---|---|
| Mi | 7.7 ± 0.6A | 8.2 ± 1.0A | 113.0 ± 19.0 |
| MFi | 7.6 ± 0.6A | 5.2 ± 1.5AC | 93.0 ± 9.5 |
| Ei | 5.6 ± 0.2A | 5.0 ± 0.4AB | 122.1 ± 10.1 |
| EFi | 5.7 ± 0.3A | 4.8 ± 0.3AB | 130.6 ± 6.5 |
| Mf | 5.1 ± 1.1AB | 1.0 ± 0.6BD | 129.9 ± 41.1 |
| MFf | 6.1 ± 0.6A | 1.5 ± 1.0BCD | 130.4 ± 32.0 |
| Ef | 5.0 ± 0.9AB | 0.4 ± 0.4D | 161.5 ± 15.2 |
| EFf | 2.0 ± 0.5B | 1.1 ± 0.7BCD | 85.5 ± 16.5 |
| Source | Sum of Sqrs | df | Mean Square | F | P |
|---|---|---|---|---|---|
| Mesotrophic conditions (M, MF) | |||||
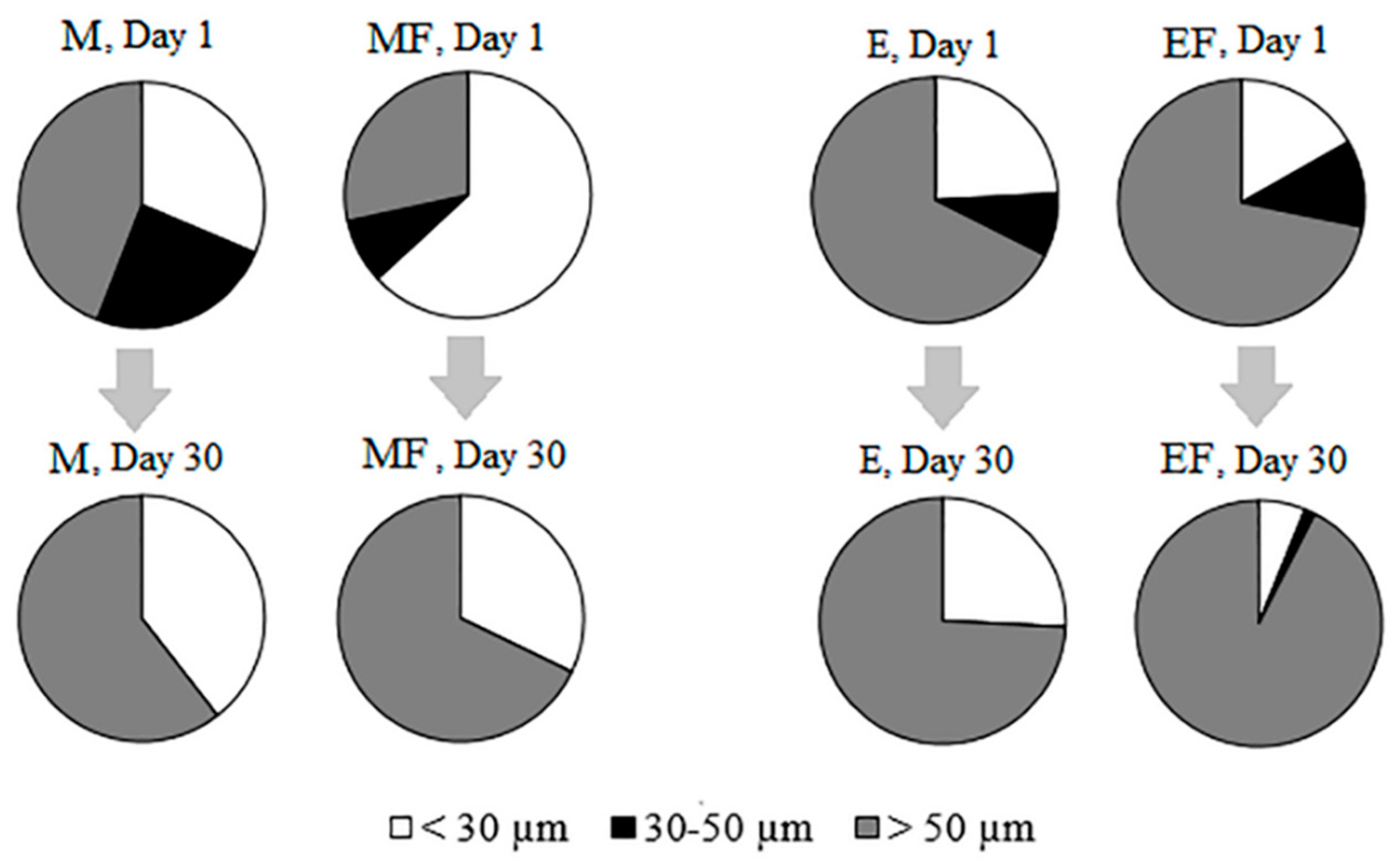
| Phytoplankton biomass <30 µm | |||||
| Fish (presence/absence) | 0.20 | 1 | 0.20 | 5.70 | 0.14 |
| Time, days (1, 10, 20, 30) | 1.85 | 3 | 0.62 | 7.26 | 0.02 |
| Time × Fish | 0.14 | 3 | 0.05 | 0.97 | 0.47 |
| Phytoplankton biomass 30–50 µm | |||||
| Fish (presence/absence) | 0.005 | 1 | 0.005 | 1.39 | 0.36 |
| Time, days (1, 10, 20, 30) | 4.26 | 3 | 1.42 | 28.26 | 0.001 |
| Time × Fish | 0.69 | 3 | 0.23 | 9.38 | 0.01 |
| Phytoplankton biomass >50 µm | |||||
| Fish (presence/absence) | 0.10 | 1 | 0.10 | 0.47 | 0.57 |
| Time, days (1, 10, 20, 30) | 0.77 | 3 | 0.26 | 4.25 | 0.06 |
| Time × Fish | 0.83 | 3 | 0.28 | 4.33 | 0.06 |
| Eutrophic conditions (E, EF) | |||||
| Phytoplankton biomass <30 µm | |||||
| Fish (presence/absence) | 0.37 | 1 | 0.37 | 20.8 | 0.04 |
| Time, days (1, 10, 20, 30) | 2.98 | 3 | 0.99 | 24.5 | 0.001 |
| Time× Fish | 0.38 | 3 | 0.12 | 2.13 | 0.20 |
| Phytoplankton biomass 30–50 µm | |||||
| Fish (presence/absence) | 0.02 | 1 | 0.02 | 0.16 | 0.73 |
| Time, days (1, 10, 20, 30) | 8.47 | 3 | 2.82 | 423.1 | << 0.001 |
| Time× Fish | 0.45 | 3 | 0.15 | 0.95 | 0.47 |
| Phytoplankton biomass >50 µm | |||||
| Fish (presence/absence) | 3.53 | 1 | 3.53 | 43.1 | 0.02 |
| Time, days (1, 10, 20, 30) | 44.97 | 3 | 14.99 | 291.1 | << 0.001 |
| Time× Fish | 4.28 | 3 | 1.43 | 13.89 | 0.004 |
| Mi | MFi | Ei | EFi | Mf | MFf | Ef | EFf | F(H) | P-Value | |
|---|---|---|---|---|---|---|---|---|---|---|
| C:N (mg/mg) | 4.65 ± 0.52 | 4.09 ± 0.07 | 4.46 ± 0.26 | 3.82 ± 0.42 | 4.50 ± 0.09 | 5.18 ± 0.28 | 4.26 ± 0.39 | 4.10 ± 0.12 | 1.8 | 0.15 |
| C:P (mg/mg) | 37.0 ± 0.6 | 59.7 ± 0.6 | 50.0 ± 10.5 | 45.1 ± 5.1 | 42.6 ± 0.4 | 35.6 ± 4.0 | 44.3 ± 1.0 | 44.2 ± 2.4 | 13.7 | 0.06 |
| Treatment | C:P (mg/mg) | P-Value | C:N (mg/mg) | P-Value | ||
|---|---|---|---|---|---|---|
| Zooplankton | Seston | Zooplankton | Seston | |||
| Mi | 37.0 ± 0.6A | 136.6 ± 15.2B | P = 0.03 | 4.65 ± 0.52A | 7.16 ± 0.25B | P = 0.03 |
| MFi | 59.7 ± 0.6A | 129.0 ± 5.2B | P = 0.03 | 4.09 ± 0.07A | 6.71 ± 0.28B | P = 0.03 |
| Ei | 50.0 ± 10.5A | 182.5 ± 10.6B | P = 0.03 | 4.46 ± 0.26A | 8.18 ± 0.33B | P = 0.03 |
| EFi | 45.1 ± 5.1A | 150.1 ± 12.9B | P = 0.03 | 3.82 ± 0.42A | 7.54 ± 0.19B | P = 0.03 |
| Mf | 42.6 ± 0.4A | 173.9 ± 103.1B | P = 0.03 | 4.50 ± 0.09A | 7.39 ± 2.12A | P = 0.31 |
| MFf | 35.6 ± 4.0A | 156.7 ± 36.1B | P = 0.03 | 5.18 ± 0.28A | 6.96 ± 0.16B | P = 0.03 |
| Ef | 44.3 ± 1.0A | 68.5 ± 3.5B | P = 0.03 | 4.26 ± 0.39A | 8.29 ± 0.52B | P = 0.03 |
| EFf | 44.2 ± 2.4A | 287.6 ± 67.3B | P = 0.03 | 4.10 ± 0.12A | 6.79 ± 0.16B | P = 0.03 |
| Treatment | EPA:C, mg/g | DHA:C, mg/g | Total FA:C, mg/g |
|---|---|---|---|
| Mi | 1.05 ± 0.04A | 0.75 ± 0.10ABD | 12.88 ± 0.73A |
| MFi | 0.97 ± 0.06A | 0.56 ± 0.09AC | 10.28 ± 0.68AC |
| Ei | 0.91 ± 0.03A | 0.99 ± 0.04B | 11.57 ± 0.25AD |
| EFi | 0.85 ± 0.02AC | 1.06 ± 0.01B | 10.55 ± 0.08ADC |
| Mf | 0.47 ± 0.09B | 0.25 ± 0.03C | 3.66 ± 0.34E |
| MFf | 0.91 ± 0.13A | 0.67 ± 0.14ABC | 7.60 ± 1.18BC |
| Ef | 0.54 ± 0.03BC | 0.36 ± 0.09CD | 5.2 ± 0.38BE |
| EFf | 0.82 ± 0.05A | 0.89 ± 0.11AB | 9.27 ± 0.78CD |
| Treatment | EPA:C, mg/g | P-Value | DHA:C, mg/g | P-Value | Total FA:C, mg/g | P-Value | |||
|---|---|---|---|---|---|---|---|---|---|
| Zooplankton | Seston | Zooplankton | Seston | Zooplankton | Seston | ||||
| Mi | 1.07 ± 0.03A | 7.7 ± 0.6B | P = 0.03 | 0.77 ± 0.07A | 8.2 ± 1.0B | P = 0.03 | 12.87 ± 0.73A | 113 ± 19.0B | P = 0.03 |
| MFi | 1.00 ± 0.06A | 7.6 ± 0.6B | P = 0.03 | 0.57 ± 0.09A | 5.2 ± 1.5B | P = 0.03 | 10.30 ± 0.71A | 93.0 ± 9.5B | P = 0.03 |
| Ei | 0.93 ± 0.03A | 5.6 ± 0.2B | P = 0.03 | 0.97 ± 0.03A | 5.0 ± 0.4B | P = 0.03 | 11.60 ± 0.25A | 122.1 ± 10.1B | P = 0.03 |
| EFi | 0.87 ± 0.03A | 5.7 ± 0.3B | P = 0.03 | 1.10 ± 0.00A | 4.8 ± 0.3B | P = 0.02 | 10.53 ± 0.09A | 130.6 ± 6.5B | P = 0.03 |
| Mf | 0.47 ± 0.09A | 5.1 ± 1.1B | P = 0.03 | 0.23 ± 0.03A | 1.0 ± 0.6A | P = 0.30 | 3.67 ± 0.33A | 129.9 ± 41.1B | P = 0.03 |
| MFf | 0.93 ± 0.13A | 6.1 ± 0.6B | P = 0.03 | 0.63 ± 0.15A | 1.5 ± 1.0A | P = 0.31 | 7.60 ± 1.15A | 130.4 ± 32.0B | P = 0.03 |
| Ef | 0.53 ± 0.03A | 5.0 ± 0.9B | P = 0.03 | 0.37 ± 0.09A | 0.4 ± 0.4A | P = 0.66 | 5.30 ± 0.36A | 161.5 ± 15.2B | P = 0.03 |
| EFf | 0.83 ± 0.07A | 2.0 ± 0.5B | P = 0.03 | 0.90 ± 0.12A | 1.1 ± 0.7A | P = 1 | 9.27 ± 0.79A | 85.5 ± 16.5B | P = 0.03 |
| Chlorophyll | M | MF | E | EF | F (H) | P |
|---|---|---|---|---|---|---|
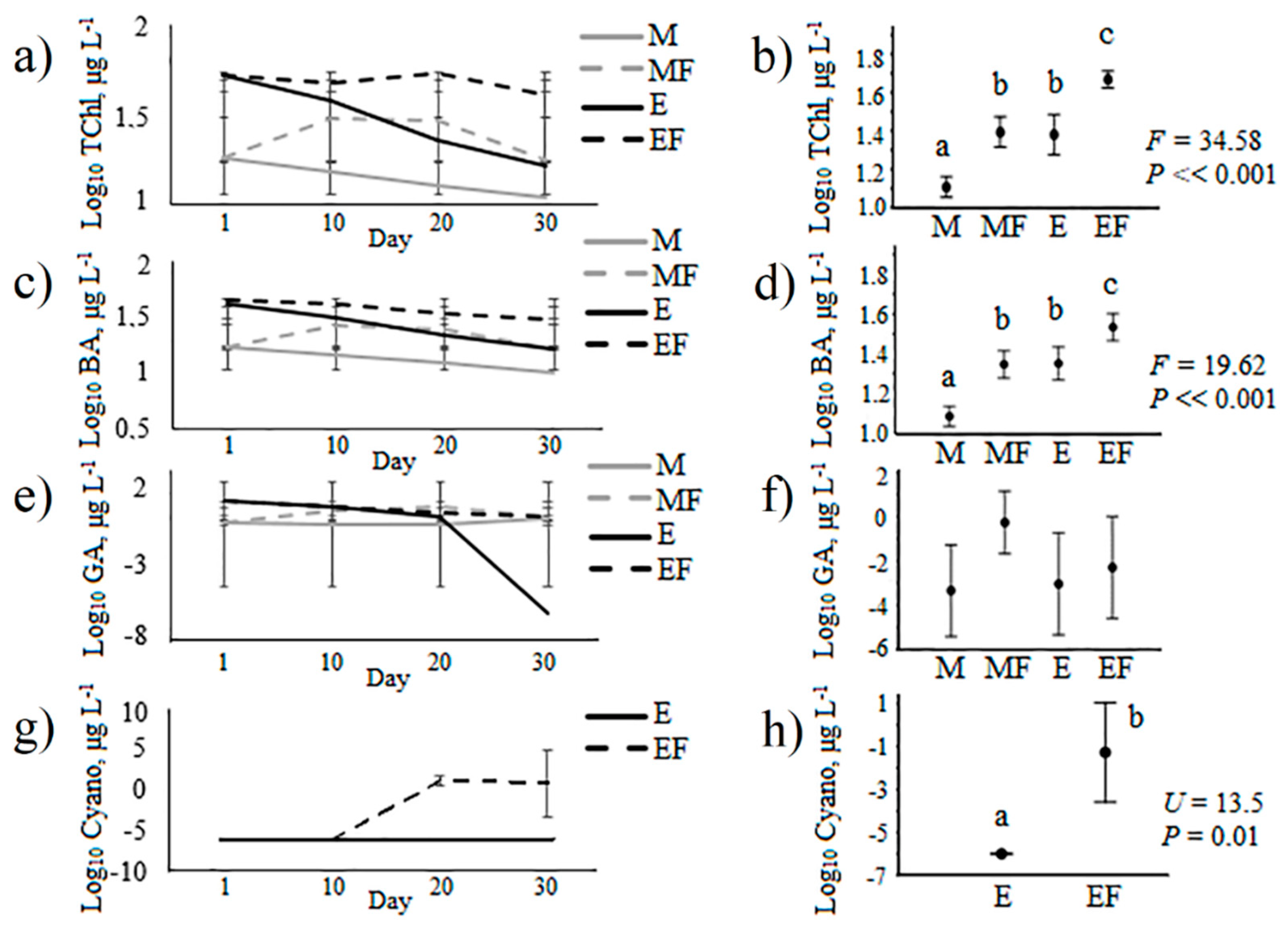
| Total, µg L−1 | 13.26 ± 0.48a | 29.60 ± 1.21b | 22.95 ± 1.21c | 42.28 ± 1.26d | 125.4 | <<0.01 |
| Diatoms&Dinoflagellates, µg L−1 | 12.61 ± 0.37a | 25.23 ± 0.95b | 20.71 ± 0.66c | 28.98 ± 0.69d | 102.3 | <<0.01 |
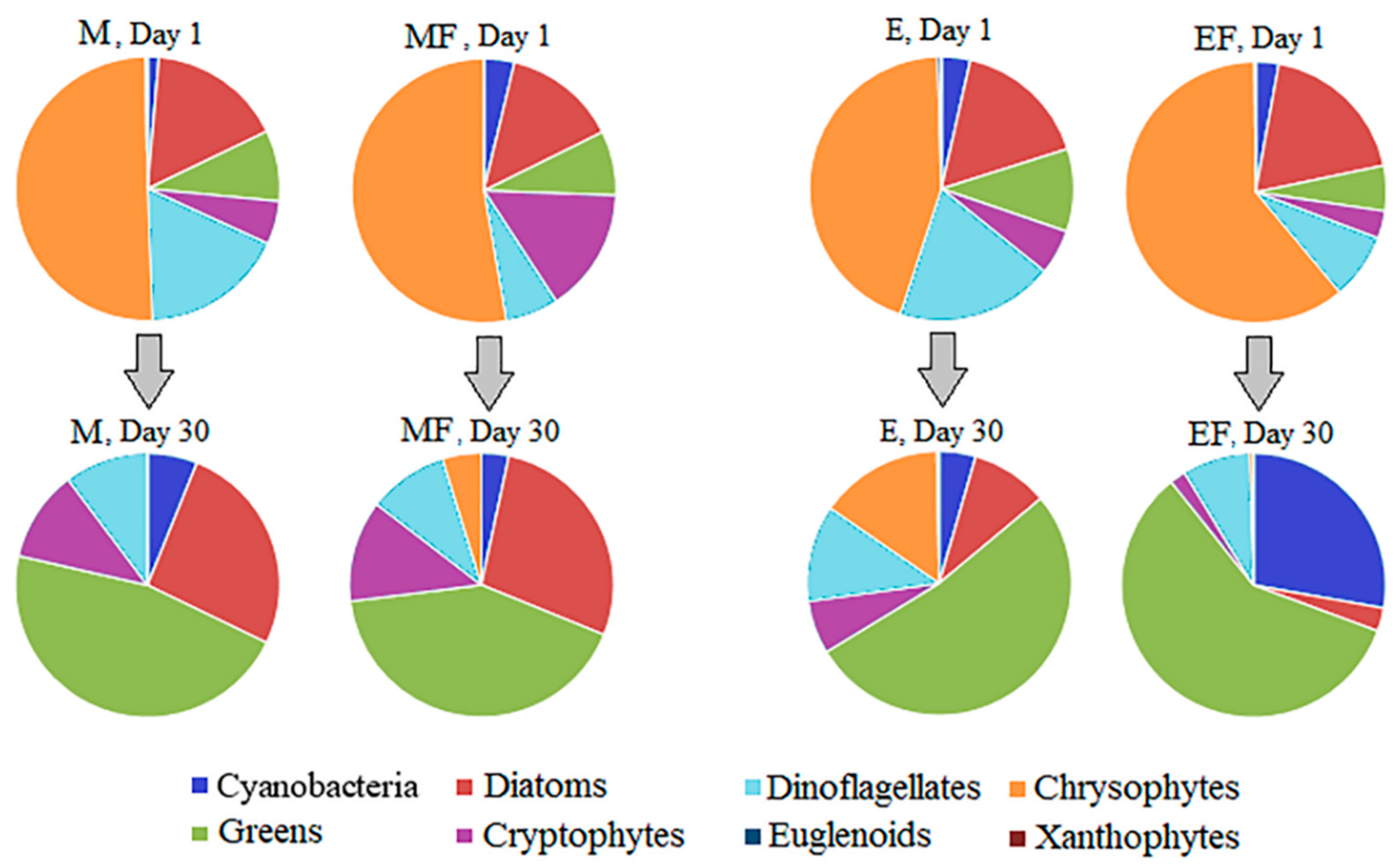
| Greens algae, µg L−1 | 0.62 ± 0.14a | 4.33 ± 0.46b | 2.23 ± 0.60c | 5.74 ± 0.79b | 16.66 | <<0.01 |
| Cyanobacteria, µg L−1 | 0a | 0.04 ± 0.04a | 0a | 7.56 ± 1.13b | 77.63 | <<0.01 |
| Treatments | Mesocosms | D. pulicaria | D. magna | C. pulchella | F (H) | P |
|---|---|---|---|---|---|---|
| M | 13.3 ± 0.5 | 12.4 ± 0.5 | 14.2 ± 1.6 | 15.4 ± 1.4 | 4.1 | 0.25 |
| MF | 29.6 ± 1.2A | 20.9 ± 0.8B | 19.8 ± 0.8B | 31.5 ± 1.5A | 53.8 | <<0.01 |
| E | 22.9 ± 1.2 | 25.7 ± 1.5 | 23.8 ± 1.3 | 27.0 ± 1.5 | 1.2 | 0.33 |
| EF | 42.3 ± 1.3 | 41.7 ± 1.9 | 46.4 ± 4.5 | 45.9 ± 1.3 | 4.3 | 0.23 |
| Treatments | D. pulicaria | D. magna | C. pulchella | |
|---|---|---|---|---|
| Population growth rate | ||||
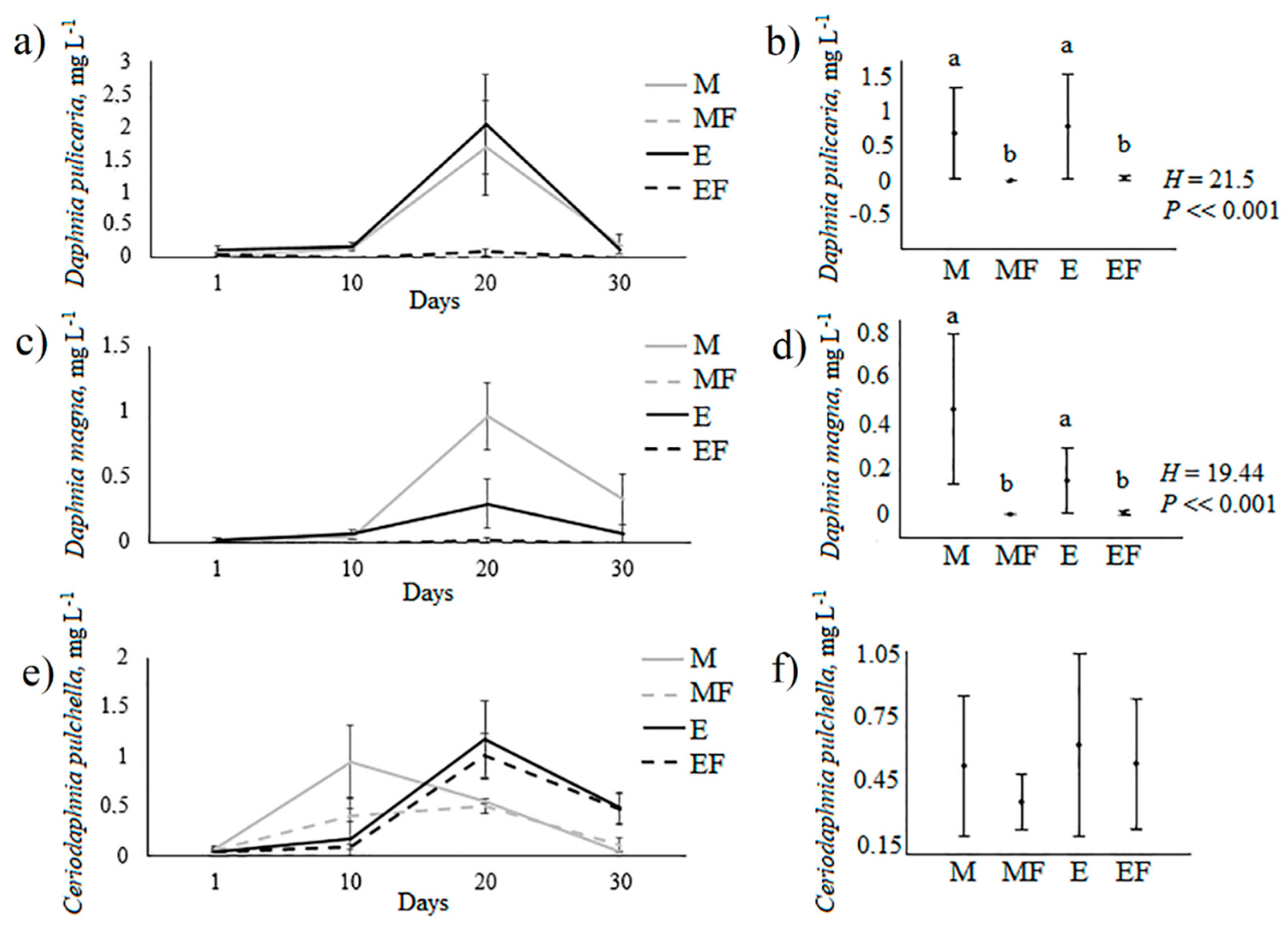
| M | −2 ± 0a, * | −2 ± 0a, * | 0.02 ± 0.01a, ** | H* = 7.6, P* = 0.02 |
| MF | 0.17 ± 0.03b, * | 0.14 ± 0.03b, * | 0.29 ± 0.02bc, ** | H* = 6.0, P* = 0.05 |
| E | −2 ± 0a, * | −2 ± 0a, * | 0.27 ± 0.04b, ** | H* = 7.6, P* = 0.02 |
| EF | −0.013 ± 0.04c, * | −0.15 ± 0.10c, * | 0.37 ± 0.01c, ** | H* = 6.0, P* = 0.05 |
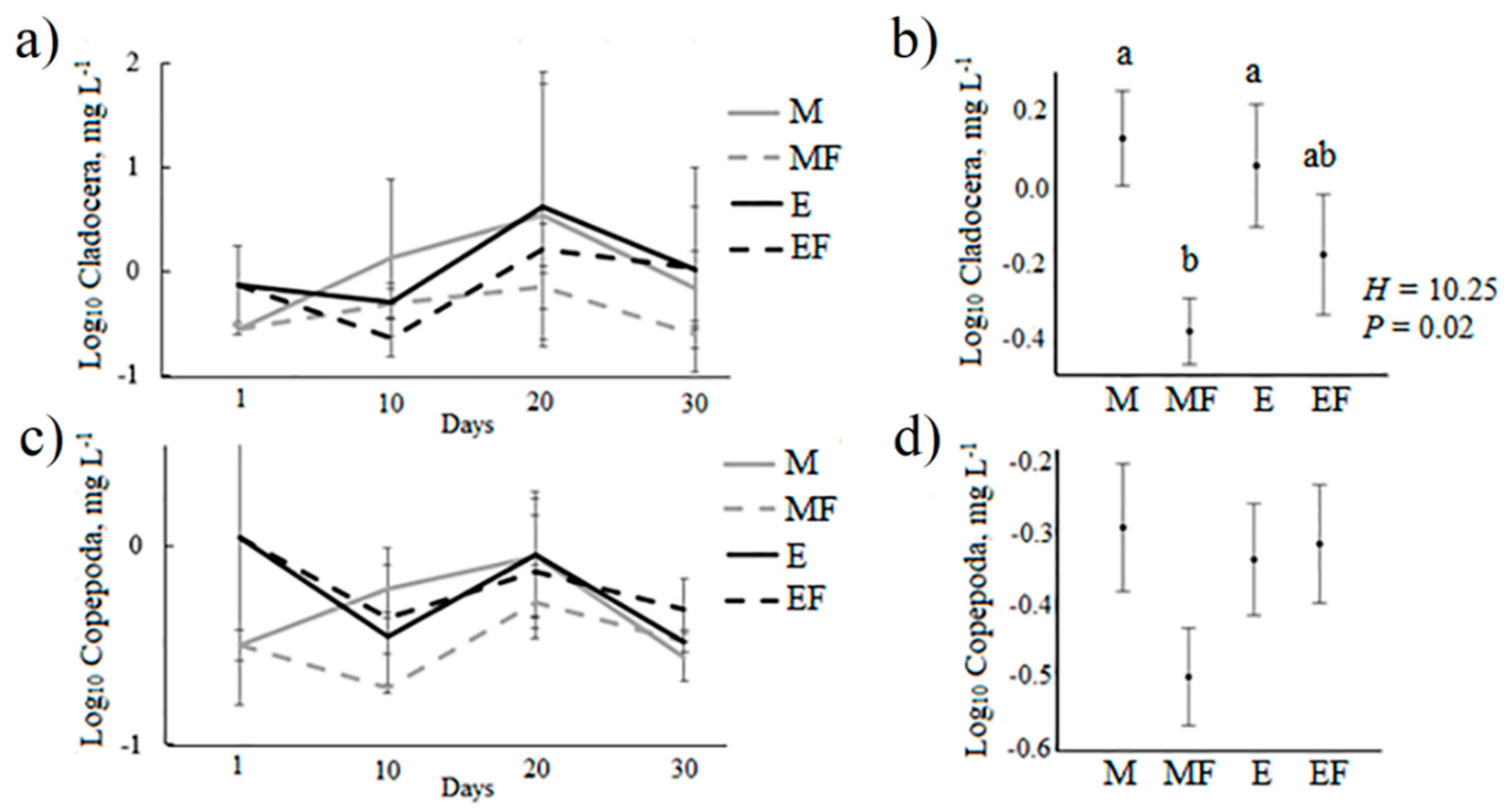
| H = 10.6 P = 0.01 | H = 10.6 P = 0.01 | H = 9.4 P = 0.02 | ||
| Fecundity | ||||
| M | 0a, * | 0a, * | 0.25 ± 0.01a, ** | H* = 7.6, P* = 0.02 |
| MF | 2.72 ± 1.52b, * | 2.13 ± 0.35b, * | 2.39 ± 0.43b, * | H* = 0.3, P* = 0.88 |
| E | 0a, * | 0a., * | 1.69 ± 0.53b, ** | H* = 7.6, P* = 0.02 |
| EF | 0.73 ± 0.34ab, * | 0.17 ± 0.03a, * | 3.77 ± 0.32c, ** | H* = 7.3, P* = 0.03 |
| H = 10.2 P = 0.02 | H = 10.7 P = 0.01 | H = 9.5 P = 0.02 | ||
| Time of first clutch | ||||
| M | 30 ± 0a, * | 30 ± 0a, * | 10 ± 1.15a, ** | H* = 7.6, P* = 0.02 |
| MF | 6.67 ± 0.67b, * | 7.33 ± 0.67b, * | 4 ± 0b, ** | H* = 6.2, P* = 0.04 |
| E | 30 ± 0a, * | 30 ± 0a, * | 4 ± 0b, ** | H* = 8.0, P* = 0.02 |
| EF | 9.33 ± 0.67c, * | 10 ± 0c, * | 4 ± 0b, ** | H* = 7.0, P* = 0.03 |
| H = 10.5 P = 0.01 | H = 10.9 P = 0.01 | H = 10.7 P = 0.01 | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feniova, I.; Sakharova, E.; Karpowicz, M.; Gladyshev, M.I.; Sushchik, N.N.; Dawidowicz, P.; Gorelysheva, Z.; Górniak, A.; Stroinov, Y.; Dzialowski, A. Direct and Indirect Impacts of Fish on Crustacean Zooplankton in Experimental Mesocosms. Water 2019, 11, 2090. https://doi.org/10.3390/w11102090
Feniova I, Sakharova E, Karpowicz M, Gladyshev MI, Sushchik NN, Dawidowicz P, Gorelysheva Z, Górniak A, Stroinov Y, Dzialowski A. Direct and Indirect Impacts of Fish on Crustacean Zooplankton in Experimental Mesocosms. Water. 2019; 11(10):2090. https://doi.org/10.3390/w11102090
Chicago/Turabian StyleFeniova, Irina, Ekaterina Sakharova, Maciej Karpowicz, Michail I. Gladyshev, Nadezhda N. Sushchik, Piotr Dawidowicz, Zoya Gorelysheva, Andrzej Górniak, Yaroslav Stroinov, and Andrew Dzialowski. 2019. "Direct and Indirect Impacts of Fish on Crustacean Zooplankton in Experimental Mesocosms" Water 11, no. 10: 2090. https://doi.org/10.3390/w11102090
APA StyleFeniova, I., Sakharova, E., Karpowicz, M., Gladyshev, M. I., Sushchik, N. N., Dawidowicz, P., Gorelysheva, Z., Górniak, A., Stroinov, Y., & Dzialowski, A. (2019). Direct and Indirect Impacts of Fish on Crustacean Zooplankton in Experimental Mesocosms. Water, 11(10), 2090. https://doi.org/10.3390/w11102090