Ice Nucleating Particle Concentrations Increase When Leaves Fall in Autumn

Abstract
1. Introduction
2. Experiments
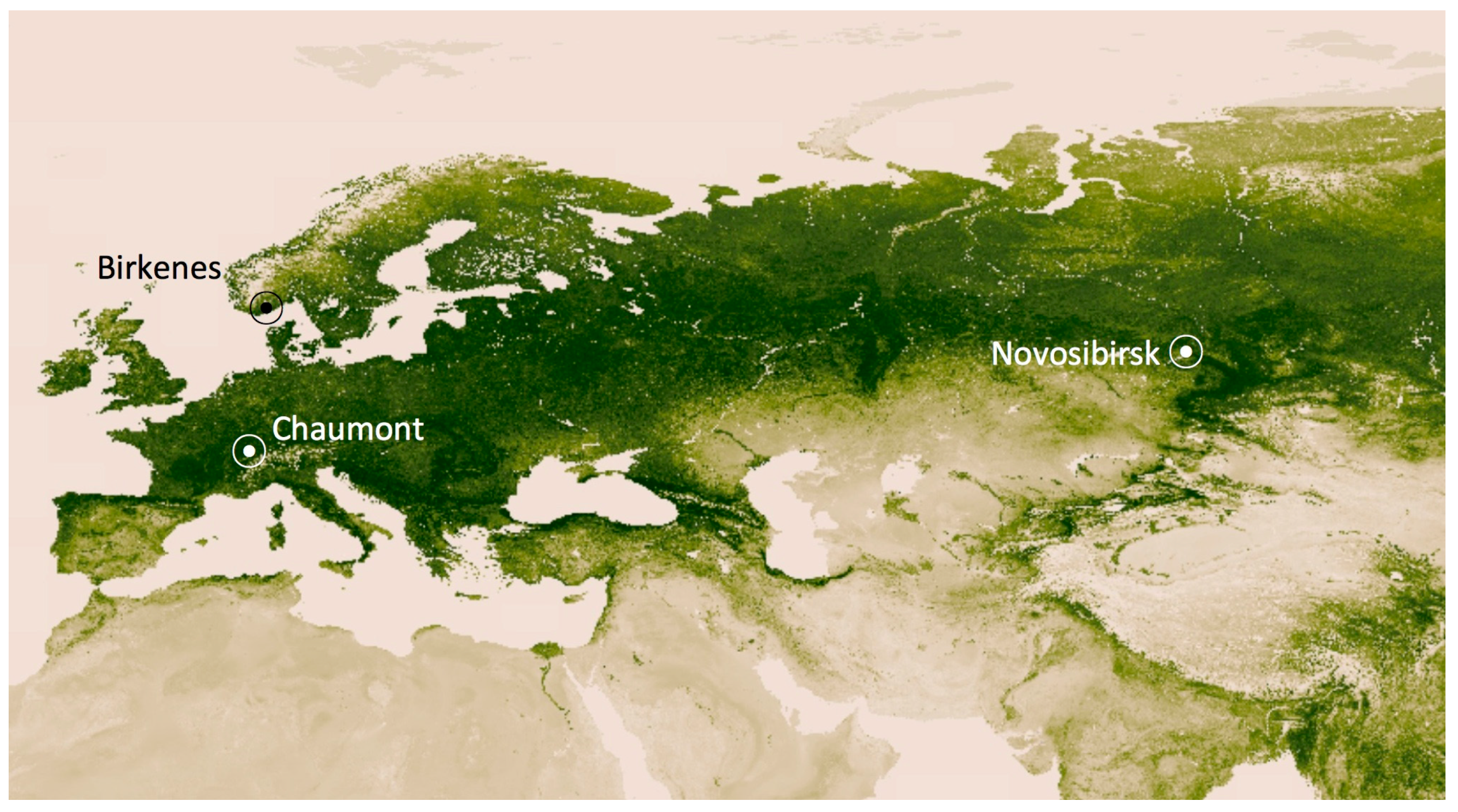
2.1. Altitudinal Gradient
2.2. Time Series
3. Results and Discussion
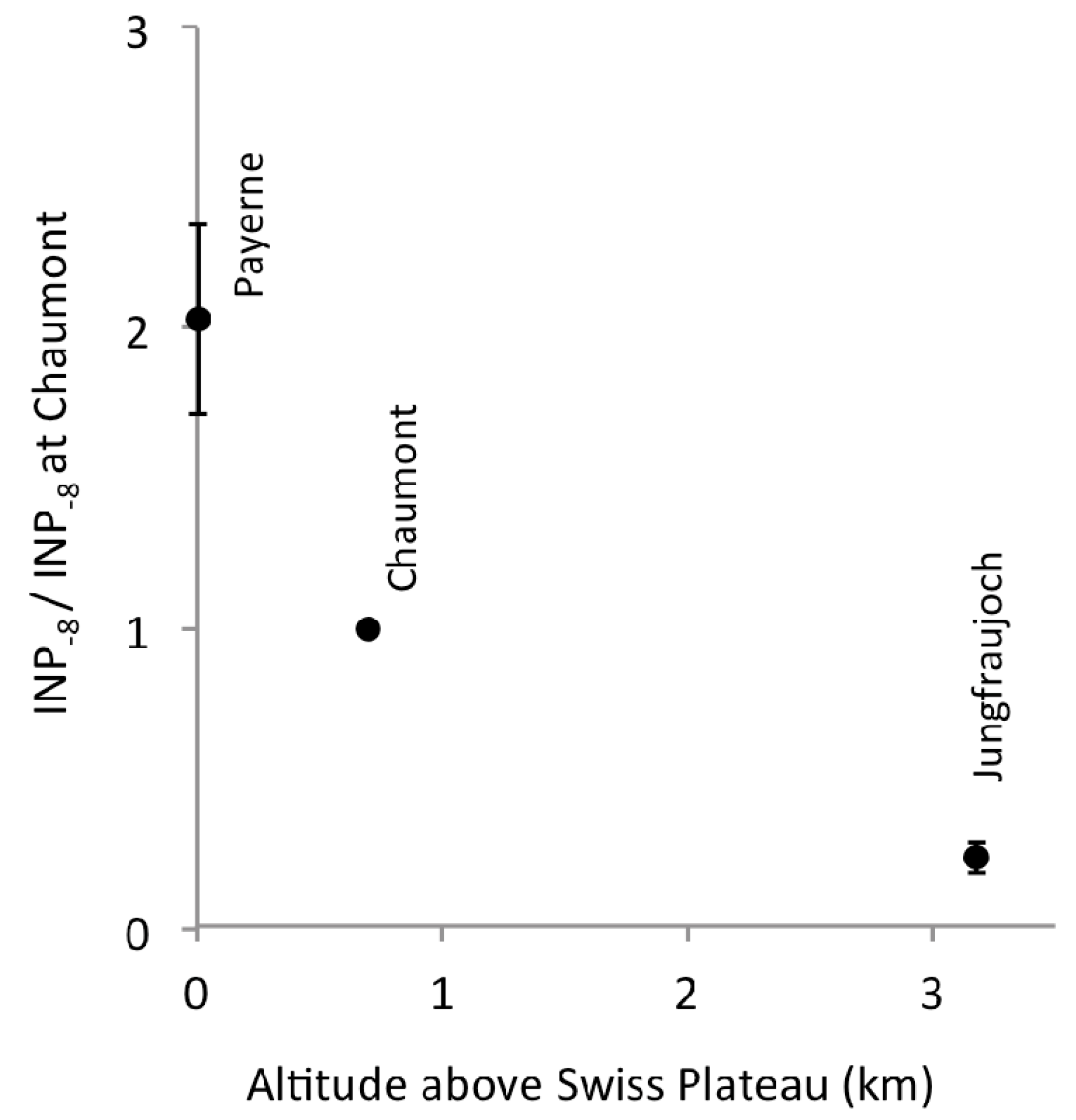
3.1. Altitudinal Gradient
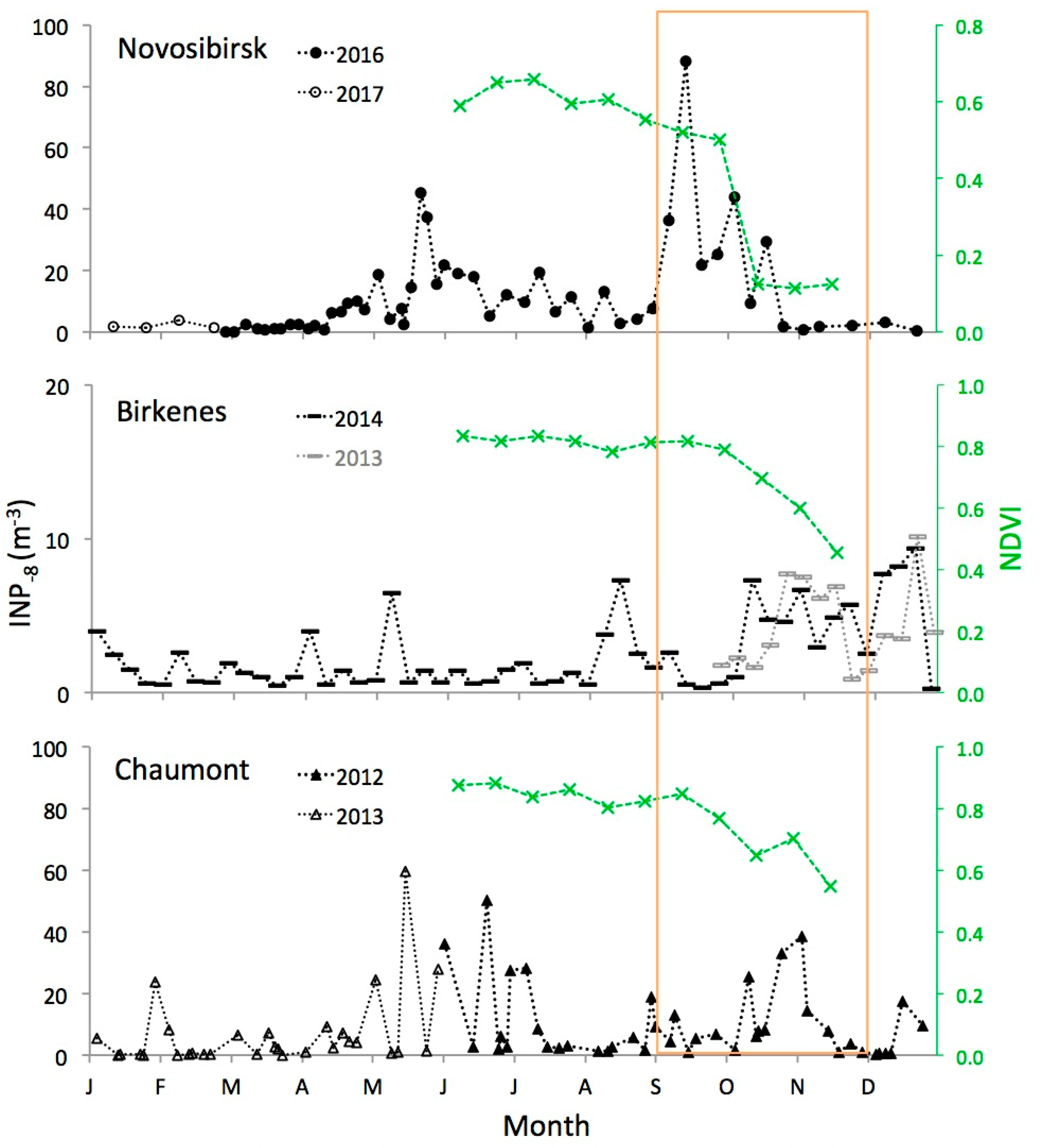
3.2. Autumn Peak
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Maki, L.R.; Galyan, E.L.; Chien, M.C.; Caldwell, D.R. Ice nucleation induced by Pseudomonas syringae. Appl. Microbiol. 1974, 28, 456–459. [Google Scholar] [PubMed]
- Sands, D.C.; Langhans, V.E.; Scharen, A.L.; de Smet, G. The association between bacteria and rain and possible resultant meteorological implications. J. Hung. Meteorol. Serv. 1982, 86, 148–152. [Google Scholar]
- Morris, C.E.; Conen, F.; Huffman, J.A.; Phillips, V.; Pöschl, U.; Sands, D.C. Bioprecipitation: A feedback cycle linking Earth history, ecosystem dynamics and land use through biological ice nucleators in the atmosphere. Glob. Chang. Biol. 2014, 20, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Stopelli, E.; Conen, F.; Guilbaud, C.; Zopfi, J.; Alewell, C.; Morris, C.E. Ice nucleators, bacterial cells and Pseudomonas syringae in precipitation at Jungfraujoch. Biogeosciences 2017, 14, 1189–1196. [Google Scholar] [CrossRef]
- Desprès, V.R.; Huffman, J.A.; Burrows, S.M.; Hoose, C.; Safatov, A.S.; Buryak, G.; Fröhlich-Nowoisky, J.; Elbert, W.; Andreae, M.O.; Pöschl, U.; et al. Primary biological aerosol particles in the atmosphere: A review. Tellus B 2012, 64, 15598. [Google Scholar] [CrossRef]
- Amato, P.; Joly, M.; Schaupp, C.; Attard, E.; Möhler, O.; Morris, C.E.; Brunet, Y.; Delort, A.M. Survival and ice nucleation activity of bacteria as aerosols in a cloud simulation chamber. Atmos. Chem. Phys. 2015, 15, 6455–6465. [Google Scholar] [CrossRef]
- Pummer, B.G.; Budke, C.; Augustin-Bauditz, S.; Niedermeier, D.; Felgitsch, L.; Kampf, C.J.; Huber, R.G.; Liedl, K.R.; Loerting, T.; Moschen, T.; et al. Ice nucleation by water-soluble macromolecules. Atmos. Chem. Phys. 2015, 15, 4077–4091. [Google Scholar] [CrossRef]
- Schnell, R.C.; Vali, G. Atmospheric ice nuclei from decomposing vegetation. Nature 1972, 236, 163–165. [Google Scholar] [CrossRef]
- Schnell, R.C.; Vali, G. World-wide source of leaf derived freezing nuclei. Nature 1973, 246, 212–213. [Google Scholar] [CrossRef]
- Thimonier, A.; Sedivy, I.; Schleppi, P. Estimating leaf area index in different types of mature forest stands in Switzerland: A comparison of methods. Eur. J. For. Res. 2010, 129, 543–562. [Google Scholar] [CrossRef]
- NOAA Environmental Visualisation Laboratory. Green: Vegetation on Our Planet. Available online: https://www.nnvl.noaa.gov/green.php (accessed on 22 May 2017).
- Conen, F.; Henne, S.; Morris, C.E.; Alewell, C. Atmospheric ice nucleators active ≥ −12 °C can be quantified on PM10 filters. Atmos. Meas. Tech. 2012, 5, 321–327. [Google Scholar] [CrossRef]
- Conen, F.; Rodriguez, S.; Hüglin, C.; Henne, S.; Herrmann, E.; Bukowiecki, N.; Alewell, C. Atmospheric ice nuclei at the high-altitude observatory Jungfraujoch, Switzerland. Tellus B 2015, 67, 25014. [Google Scholar] [CrossRef]
- Federal Office of Topography Swisstopo. Landsat Mosaic. Available online: https://shop.swisstopo.admin.ch/en/products/images/ortho_images/landsat (accessed on 10 October 2017).
- Conen, F.; Eckhardt, S.; Gundersen, H.; Stohl, A.; Yttri, K.E. Rainfall drives atmospheric ice nucleating particles in the maritime climate of Southern Norway. Atmos. Chem. Phys. Discuss. 2017. [Google Scholar] [CrossRef]
- Lukasova, V.; Lang, M.; Skvarenina, J. Seasonal changes in NDVI in relation to phenological phases, LAI and PAI of beech forests. Balt. For. 2014, 20, 248–262. [Google Scholar]
- Nagai, S.; Inoue, T.; Ohtsuka, T.; Kobayashi, H.; Kurumado, K.; Moraoka, H.; Nasahara, K.N. Relationship between spatio-temporal characteristics of leaf-fall phenology and seasonal variations in near surface-and satellite-observed vegetation indices in a cool-temperate deciduous broad-leaved forest in Japan. Int. J. Remote Sens. 2014, 35, 3520–3536. [Google Scholar] [CrossRef]
- Twohy, C.H.; McMeeking, G.R.; DeMott, P.J.; McCluskey, C.S.; Hill, T.C.J.; Burrows, S.M.; Kulkarni, G.R.; Tanarhte, M.; Kafle, D.N.; Toohey, D.W. Abundance of fluorescent biological aerosol particles at temperatures conducive to the formation of mixed-phase and cirrus clouds. Atmos. Chem. Phys. 2016, 16, 8205–8225. [Google Scholar] [CrossRef]
- Wang, Q.; Adiku, S.; Tenhunen, J.; Granier, A. On the relationship of NDVI with leaf area index in a deciduous forest site. Remote Sens. Environ. 2005, 94, 244–255. [Google Scholar] [CrossRef]
- Myoung, B.; Choi, Y.S.; Hong, S.; Park, S.K. Inter- and intra-annual variability of vegetation in the northern hemisphere and its association with precursory meteorological factors. Glob. Biogeochem. Cycles 2013, 27, 31–42. [Google Scholar] [CrossRef]
- Staebler, R.M.; Fitzjarrald, D.R. Measuring canopy structure and the kinematics of subcanopy flows in two forrests. J. Appl. Meteorol. 2005, 44, 1161–1179. [Google Scholar] [CrossRef]
- Limpert, E.; Stahel, W.A.; Abbt, M. Log-normal distributions across the sciences: Keys and clues. BioScience 2001, 51, 341–352. [Google Scholar] [CrossRef]
- Limpert, E.; Burke, J.; Galan, C.; del Mar Trigo, M.; West, J.S.; Stahel, W.A. Data, not only in aerobiology: How normal is the normal distribution? Aerobiologia 2008, 24, 121–124. [Google Scholar] [CrossRef]
- Boose, Y.; Kanji, Z.A.; Kohn, M.; Sireau, B.; Zipori, A.; Crawford, I.; Lloyd, G.; Bukowiecki, N.; Herrmann, E.; Kupiszewski, P.; et al. Ice nucleating particle measurements at 241 K during winter months at 3580 m MSL in the Swiss Alps. J. Atmos. Sci. 2016, 73, 2203–2228. [Google Scholar] [CrossRef]
- Schrod, J.; Weber, D.; Drücke, J.; Kelehis, C.; Pikridas, M.; Ebert, M.; Cvetkovic, B.; Nickovic, S.; Marinou, E.; Baars, H.; et al. Ice nucleating particles over the Eastern Mediterranean measured by unmanned aircraft systems. Atmos. Chem. Phys. 2017, 17, 4817–4835. [Google Scholar] [CrossRef]
- Hande, L.B.; Engler, C.; Hoose, C.; Tegen, I. Seasonal variability of Saharan desert dust and ice nucleating particles over Europe. Atmos. Chem. Phys. 2015, 15, 4389–4397. [Google Scholar] [CrossRef]
- Kanitz, T.; Seifert, P.; Ansman, A.; Engelmann, R.; Althausen, D.; Casiccia, C.; Rohwer, E.G. Contrasting the impact of aerosol at northern and southern midlatitudes on heterogeneous ice formation. Geophys. Res. Lett. 2011, 38, L17802. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Station | Novosibirsk | Chaumont | Birkenes |
|---|---|---|---|
| sampling period | 02.16–02.17 | 06.12–05.13 | 01.14–12.14 |
| (MM.YY) | |||
| season | number of samples | ||
| spring | 24 | 18 | 14 |
| summer | 14 | 18 | 13 |
| autumn | 11 | 18 | 13 |
| winter | 7 | 18 | 12 |
| median INP−8 (m−3) | |||
| spring | 3.4 | 3.4 | 1.0 |
| summer | 10.5 | 2.9 | 1.4 |
| autumn | 21.8 | 7.4 | 2.9 |
| winter | 1.6 | 0.4 | 2.0 |
| multiplicative standard deviation (s*) | |||
| spring | 4.6 | 3.9 | 1.8 |
| summer | 2.2 | 4.0 | 2.1 |
| autumn | 7.9 | 2.3 | 3.2 |
| winter | 2.1 | 8.2 | 4.5 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Conen, F.; Yakutin, M.V.; Yttri, K.E.; Hüglin, C. Ice Nucleating Particle Concentrations Increase When Leaves Fall in Autumn. Atmosphere 2017, 8, 202. https://doi.org/10.3390/atmos8100202
Conen F, Yakutin MV, Yttri KE, Hüglin C. Ice Nucleating Particle Concentrations Increase When Leaves Fall in Autumn. Atmosphere. 2017; 8(10):202. https://doi.org/10.3390/atmos8100202
Chicago/Turabian StyleConen, Franz, Mikhail V. Yakutin, Karl Espen Yttri, and Christoph Hüglin. 2017. "Ice Nucleating Particle Concentrations Increase When Leaves Fall in Autumn" Atmosphere 8, no. 10: 202. https://doi.org/10.3390/atmos8100202
APA StyleConen, F., Yakutin, M. V., Yttri, K. E., & Hüglin, C. (2017). Ice Nucleating Particle Concentrations Increase When Leaves Fall in Autumn. Atmosphere, 8(10), 202. https://doi.org/10.3390/atmos8100202