


Response Mechanisms of Vegetation Productivity to Water Variability in Arid and Semi-Arid Areas of China: A Decoupling Analysis of Soil Moisture and Precipitation

Abstract
1. Introduction
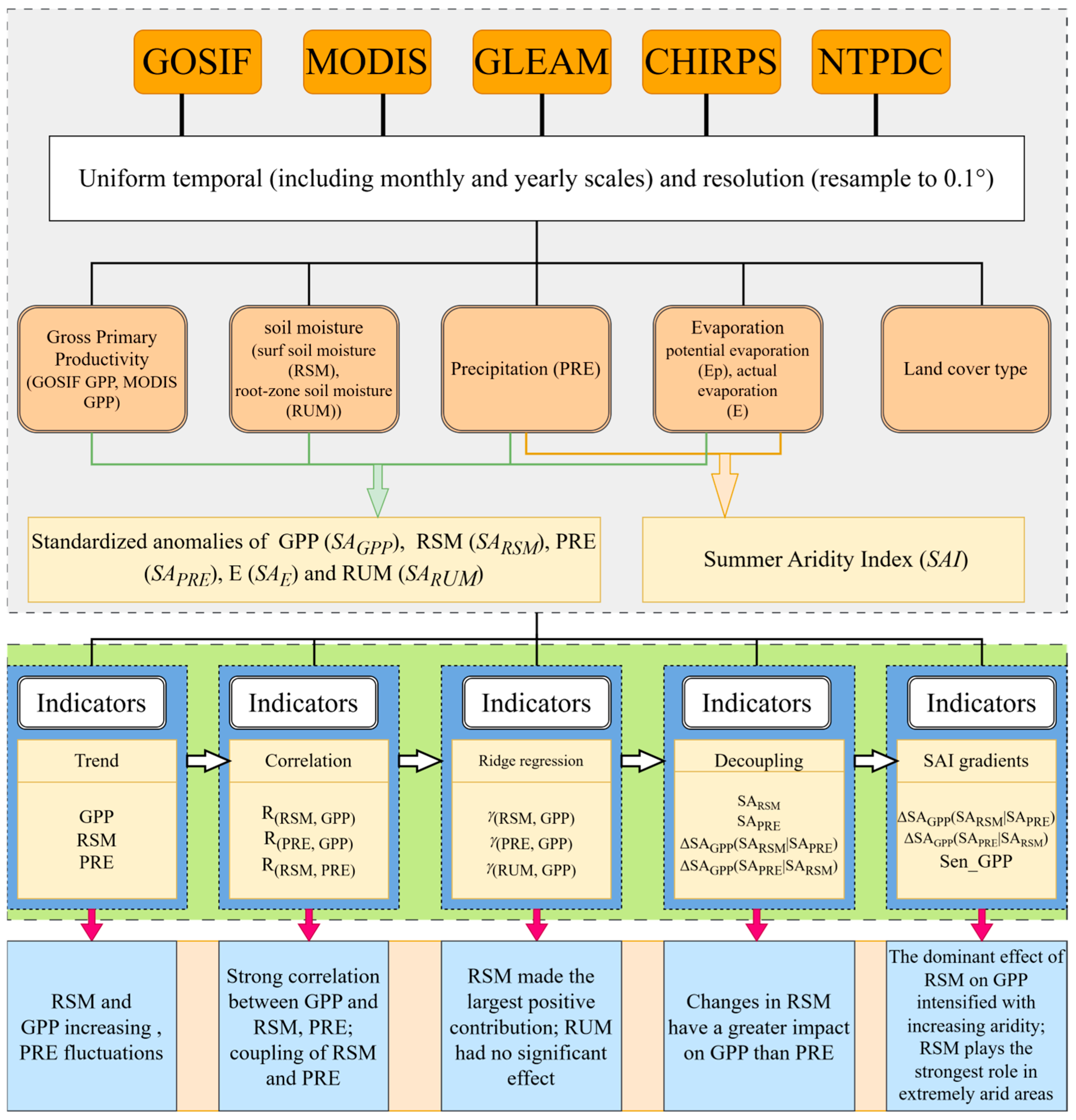
2. Materials and Methods
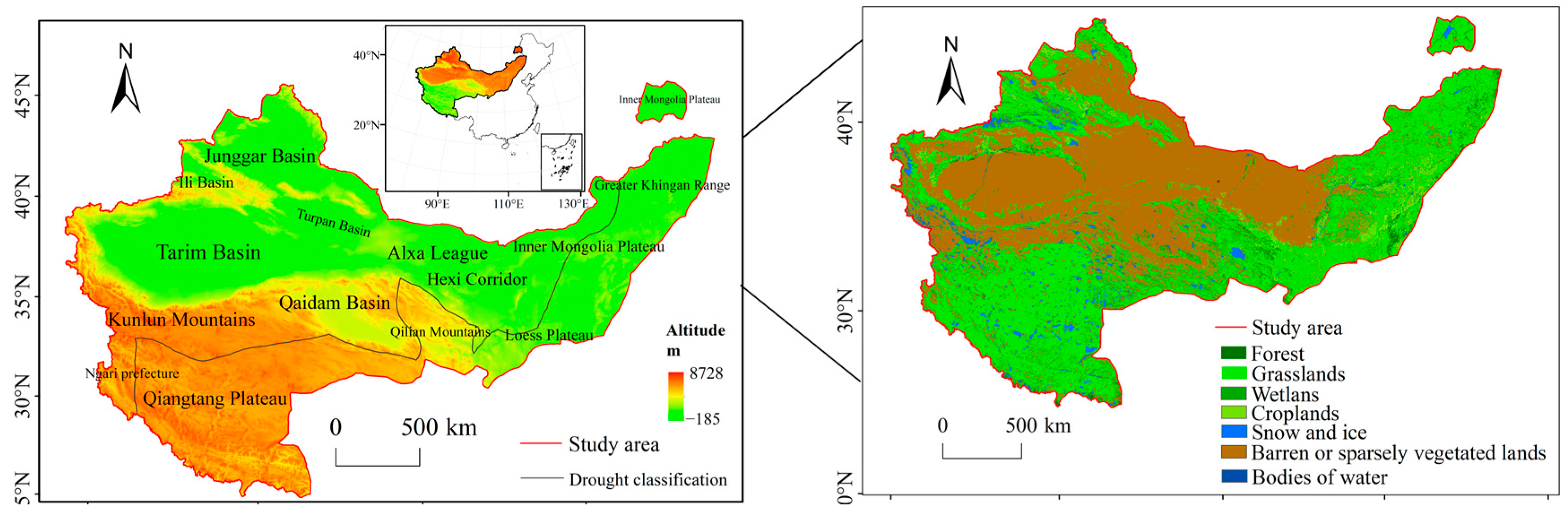
2.1. The Study Area
2.2. Data Source
2.2.1. GPP and Land Use Data
2.2.2. Auxiliary Data
2.3. Methods
2.3.1. Aridity Calculation
2.3.2. Trend Variation and Correlation Analysis
2.3.3. Standardized Anomaly
2.3.4. Data Binning
2.3.5. Ridge Regression
3. Results
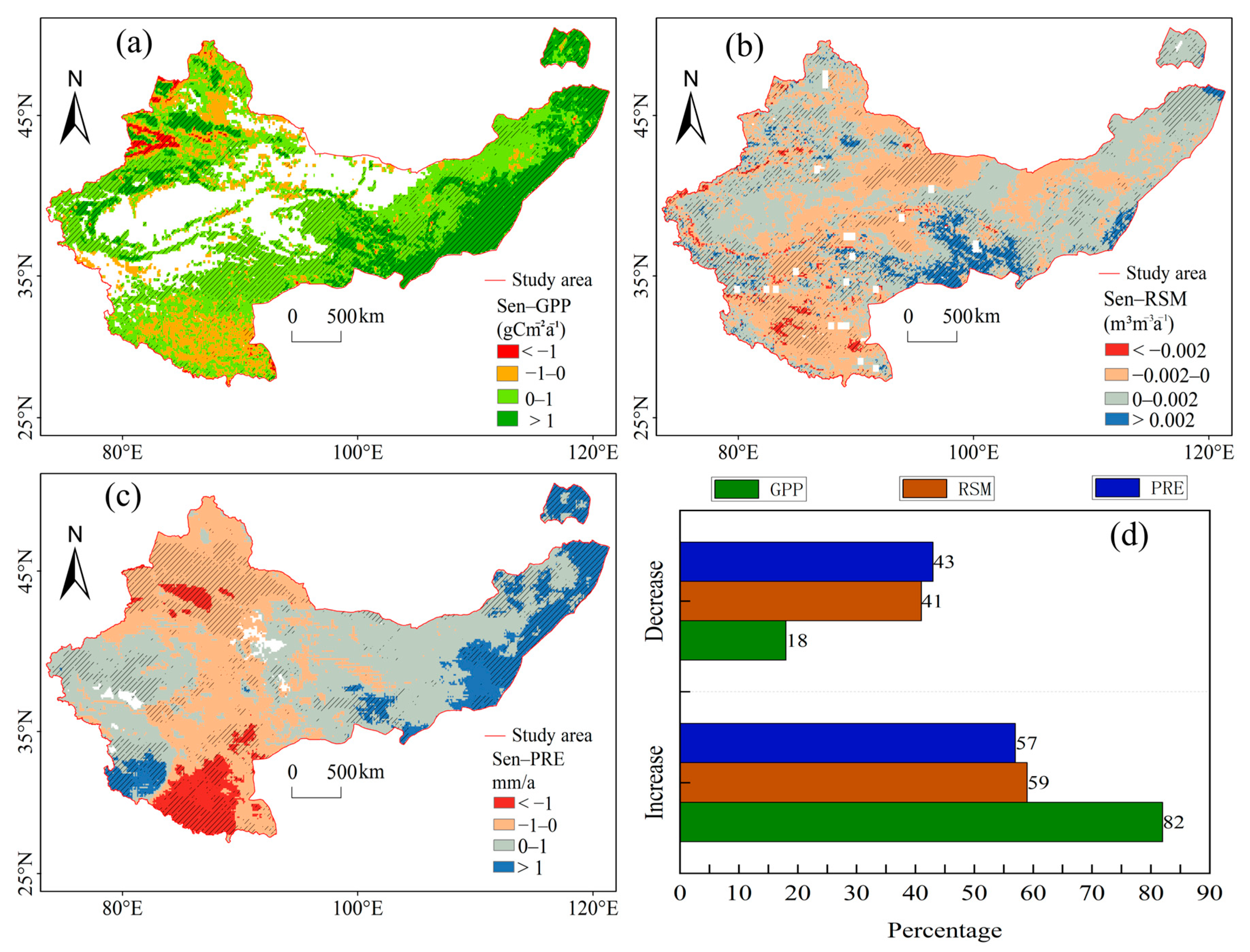
3.1. Spatiotemporal Trends of RSM, PRE, and GPP
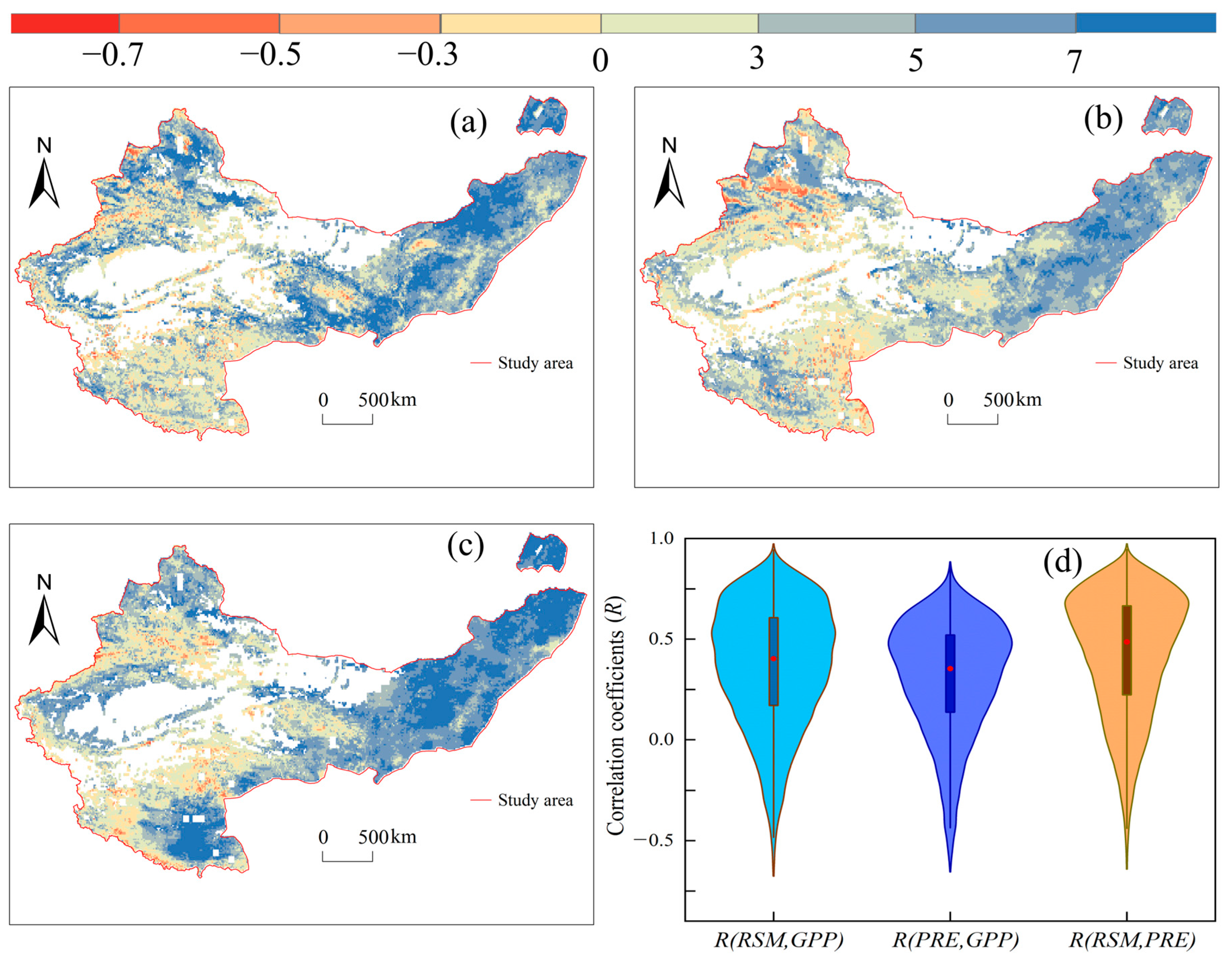
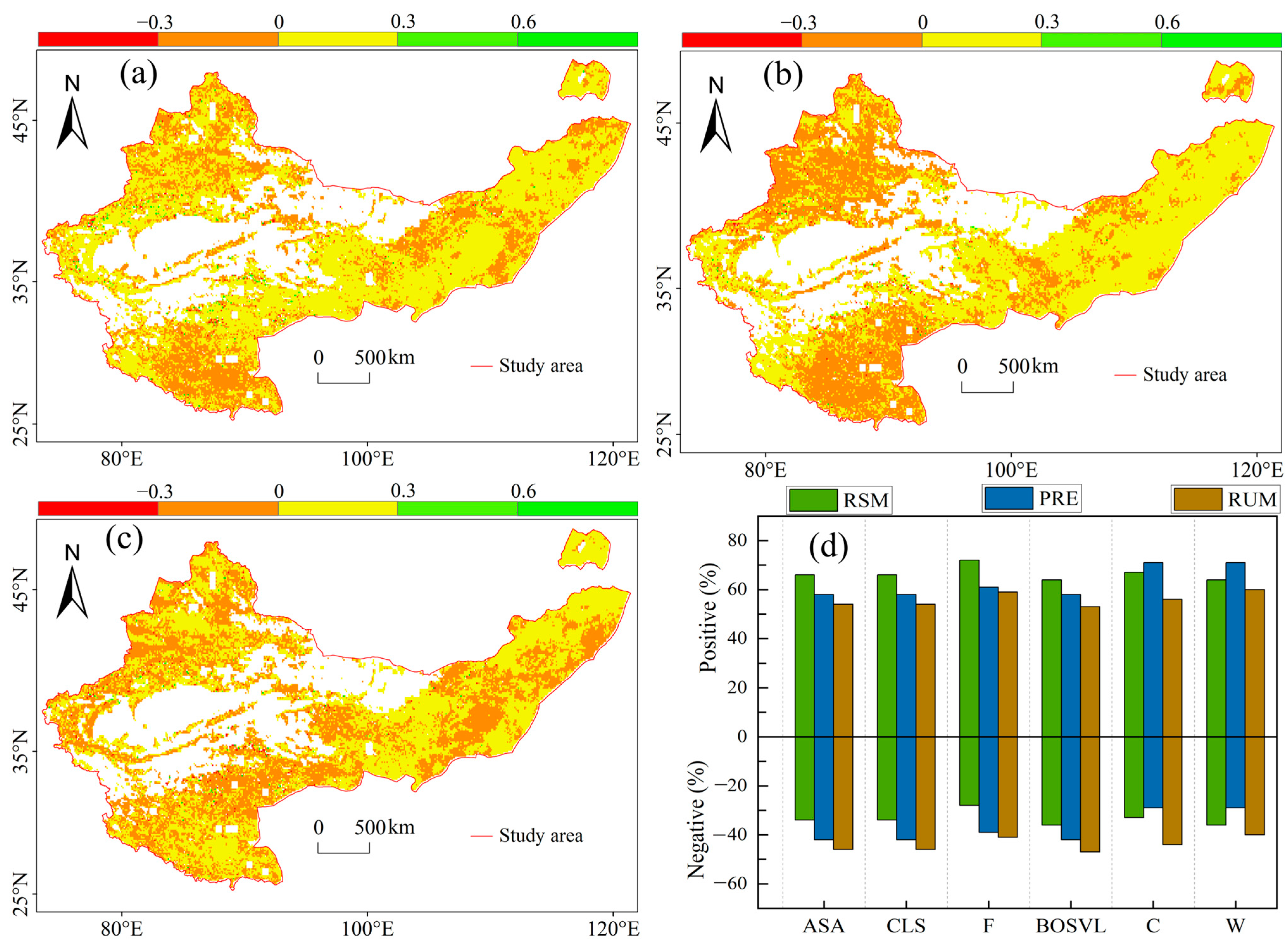
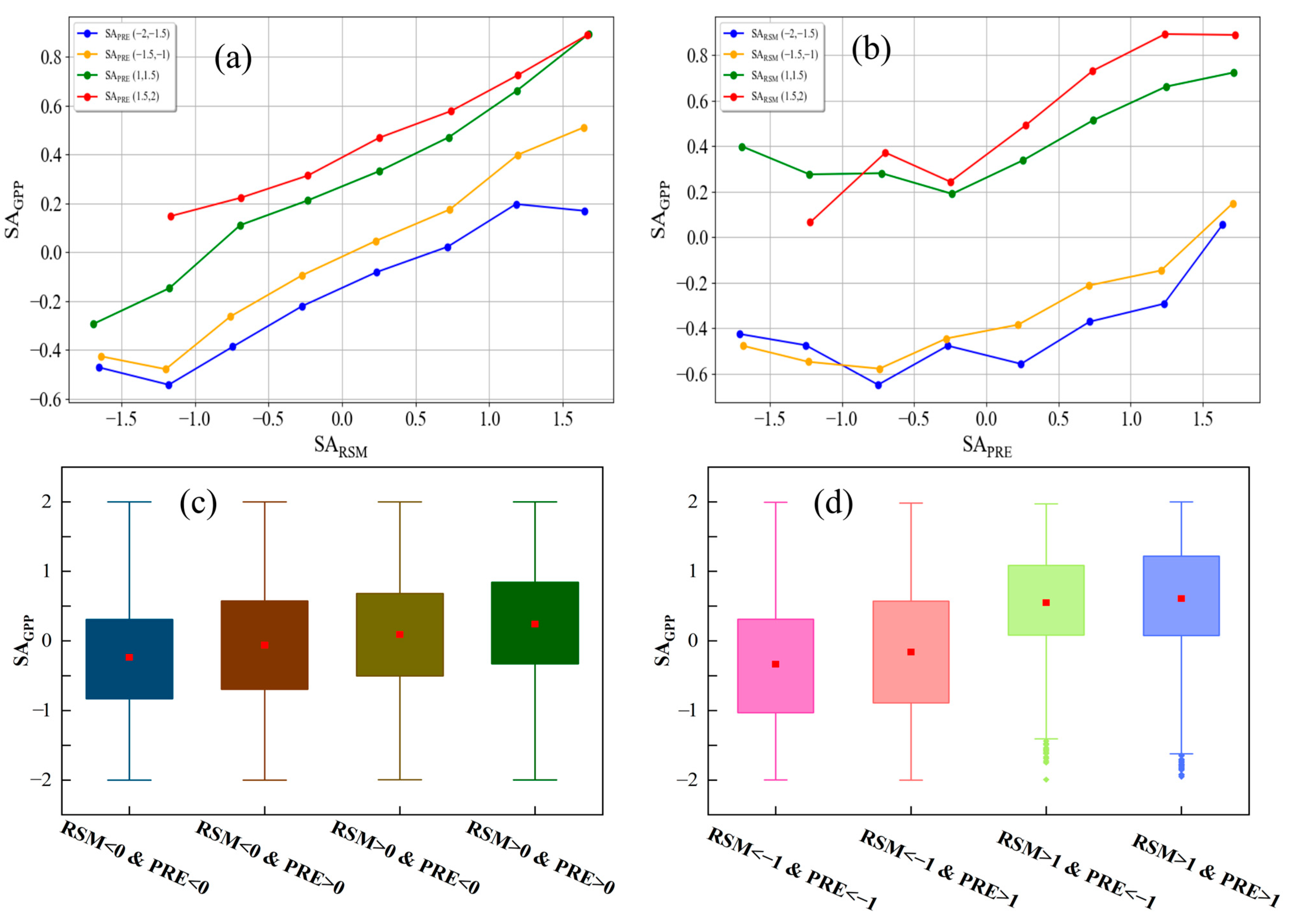
3.2. Coupling Effects of RSM and PRE on GPP
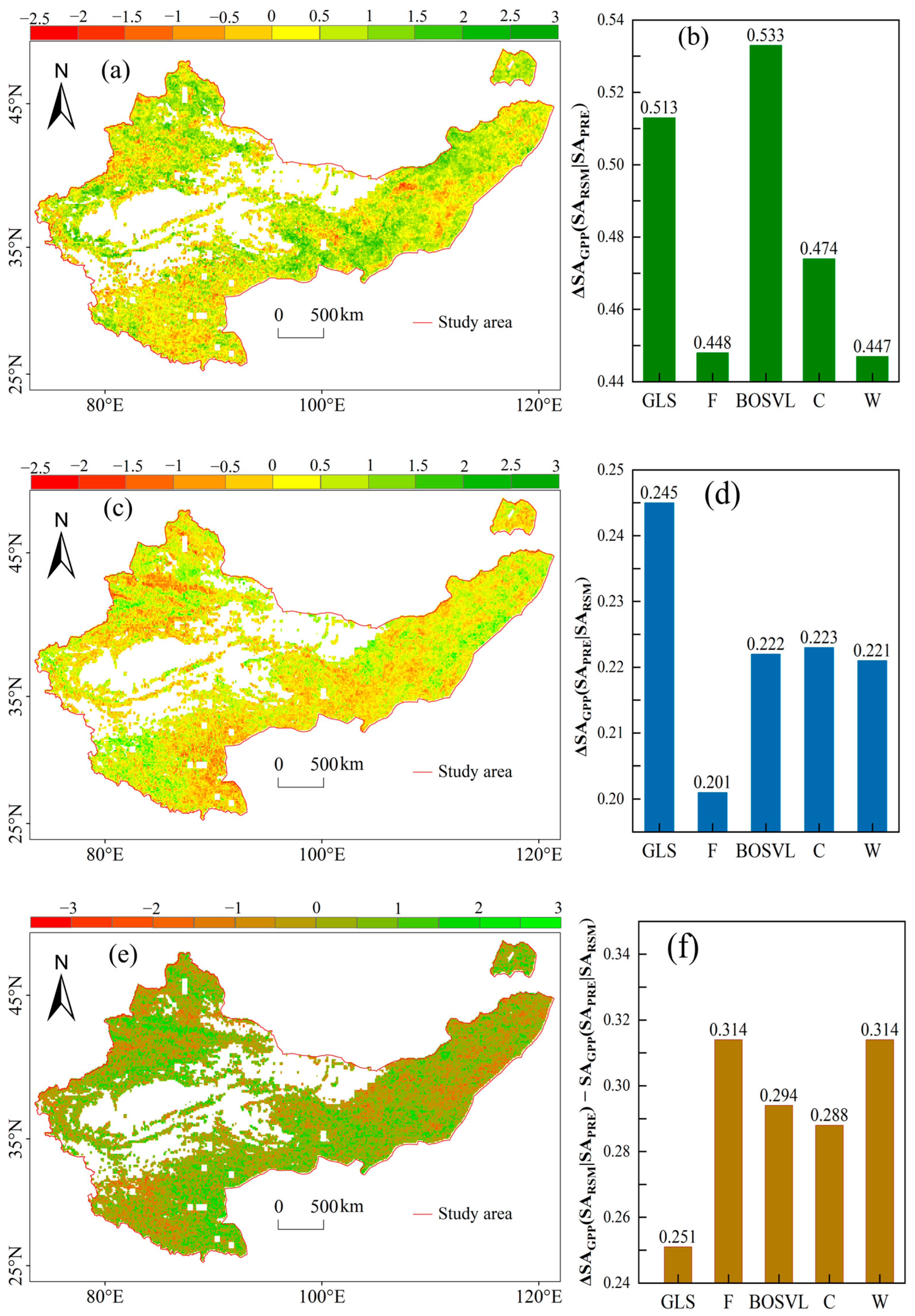
3.3. The Independent Effects of RSM and PRE on GPP
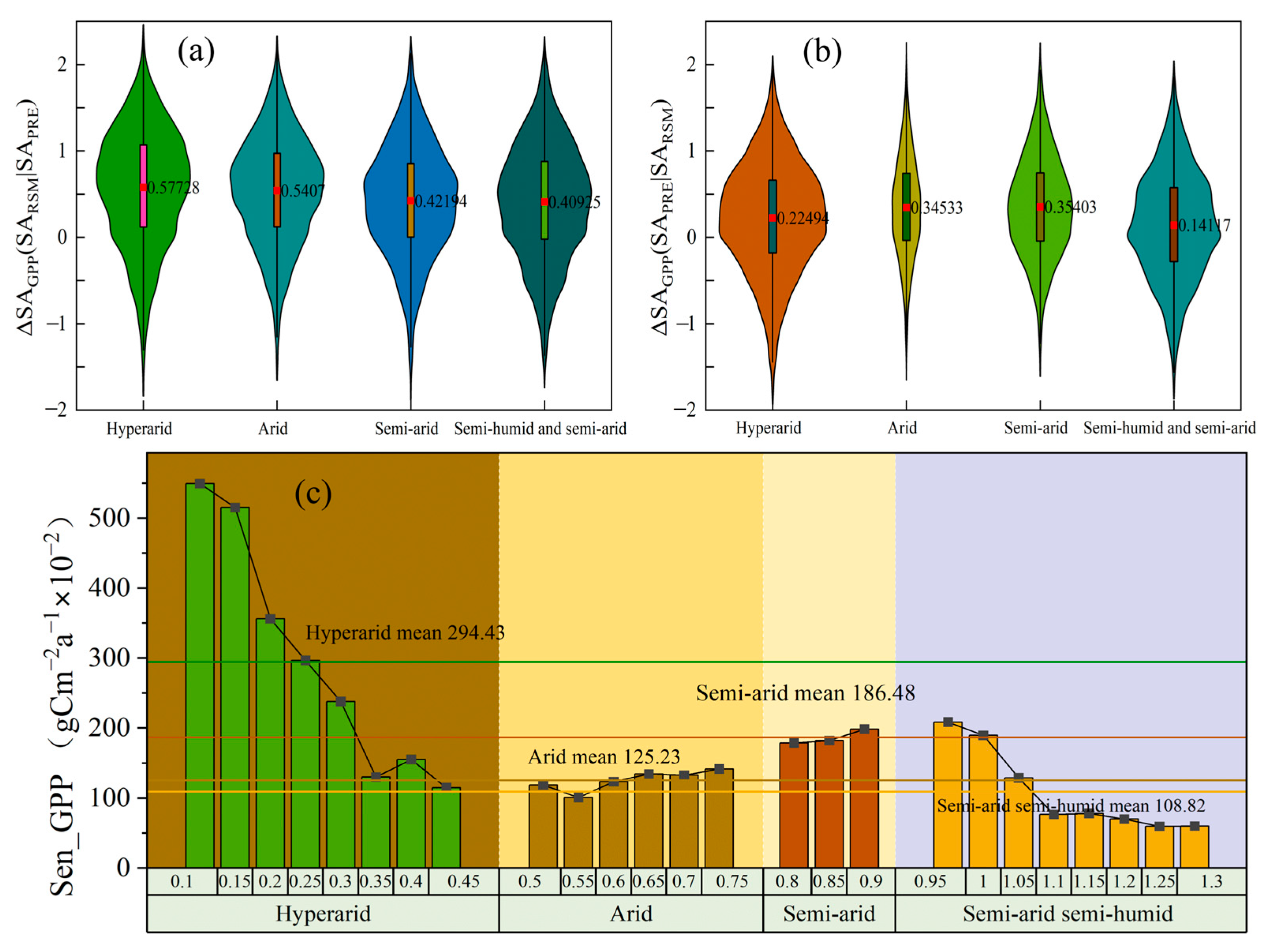
3.4. The Sensitivity of RSM and PRE to Aridity Levels
4. Discussion
4.1. Spatiotemporal Variations and Correlations of RSM, PRE, and GPP
4.2. The Decoupled Effects of RSM and PRE on GPP
4.3. The Sensitivity of RSM and PRE to Drought
4.4. Uncertainty Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhou, S.; Williams, A.P.; Lintner, B.R.; Berg, A.M.; Zhang, Y.; Keenan, T.F.; Cook, B.I.; Hagemann, S.; Seneviratne, S.I.; Gentine, P. Soil moisture–atmosphere feedbacks mitigate declining water availability in drylands. Nat. Clim. Change 2021, 11, 38–44. [Google Scholar] [CrossRef]
- Humphrey, V.; Berg, A.; Ciais, P.; Gentine, P.; Jung, M.; Reichstein, M.; Seneviratne, S.I.; Frankenberg, C. Soil moisture-atmosphere feedback dominates land carbon uptake variability. Nature 2021, 592, 65–69. [Google Scholar] [CrossRef]
- Wang, L.; She, D.; Jun, X.; Meng, L.; Li, L. Revegetation affects the response of land surface henology to climate in Loess Plateau, China. Sci. Total Environ. 2023, 860, 160383. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Williams, A.P.; Lintner, B.R.; Findell, K.L.; Keenan, T.F.; Zhang, Y.; Gentine, P. Diminishing seasonality of subtropical water availability in a warmer world dominated by soil moisture-atmosphere feedbacks. Nat. Commun. 2022, 13, 5756. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Liu, Y.; Chen, Y.; Gao, L.; Yuan, W.; Bryan, B.A.; He, B.; Duan, W.; Fang, G.; Li, Y.; et al. Future soil moisture will slowdown the advancement of ecosystem productivity in the Northern Hemisphere. Sci. Bull. 2025, 70, 347–350. [Google Scholar] [CrossRef]
- Piao, S.; Wang, X.; Park, T.; Chen, C.; Lian, X.; He, Y.; Bjerke, J.W.; Chen, A.; Ciais, P.; Tømmervik, H.; et al. Characteristics, drivers and feedbacks of global greening. Nat. Rev. Earth Environ. 2019, 1, 14–27. [Google Scholar] [CrossRef]
- Tang, X.; Li, H.; Desai, A.R.; Nagy, Z.; Luo, J.; Kolb, T.E.; Olioso, A.; Xu, X.; Yao, L.; Kutsch, W.; et al. How is water-use efficiency of terrestrial ecosystems distributed and changing on Earth? Sci. Rep. 2014, 4, 7483. [Google Scholar] [CrossRef]
- Yang, Y.; Guan, H.; Batelaan, O.; McVicar, T.R.; Long, D.; Piao, S.; Liang, W.; Liu, B.; Jin, Z.; Simmons, C.T. Contrasting responses of water use efficiency to drought across global terrestrial ecosystems. Sci. Rep. 2016, 6, 23284. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Luo, S.; Lyu, S.; Cheng, C.; Li, Z.; Zhang, S. Numerical modeling of the responses of soil temperature and soil moisture to climate change over the Tibetan Plateau, 1961–2010. Int. J. Climatol. 2021, 41, 4134–4150. [Google Scholar] [CrossRef]
- Deng, Y.; Wang, S.; Bai, X.; Luo, G.; Wu, L.; Cao, Y.; Li, H.; Li, C.; Yang, Y.; Hu, Z.; et al. Variation trend of global soil moisture and its cause analysis. Ecol. Indic. 2020, 110, 5939. [Google Scholar] [CrossRef]
- Wang, R.; Li, L.; Chen, L.; Ning, L.; Yuan, L.; Lü, G. Respective contributions of precipitation and potential evapotranspiration to long—term changes in global drought duration and intensity. Int. J. Climatol. 2022, 42, 10126–10137. [Google Scholar] [CrossRef]
- He, Q.; Ju, W.; Li, X. Response of Global Terrestrial Carbon Fluxes to Drought from 1981 to 2016. Atmosphere 2023, 14, 229. [Google Scholar] [CrossRef]
- Zhao, H.; Jia, G.; Xu, X.; Zhang, A. Earlier Precipitation Enhances Dryland Annual Primary Production. J. Geophys. Res. Biogeosci. 2023, 128, e2022JG007127. [Google Scholar] [CrossRef]
- Fang, Q.; Wang, G.; Xue, B.; Liu, T.; Kiem, A. How and to what extent does precipitation on multi-temporal scales and soil moisture at different depths determine carbon flux responses in a water-limited grassland ecosystem? Sci. Total Environ. 2018, 635, 1255–1266. [Google Scholar] [CrossRef]
- Humphrey, V.; Zscheischler, J.; Ciais, P.; Gudmundsson, L.; Sitch, S.; Seneviratne, S.I. Sensitivity of atmospheric CO2 growth rate to observed changes in terrestrial water storage. Nature 2018, 560, 628–631. [Google Scholar] [CrossRef] [PubMed]
- Smith, W.K.; Dannenberg, M.P.; Yan, D.; Herrmann, S.; Barnes, M.L.; Barron-Gafford, G.A.; Biederman, J.A.; Ferrenberg, S.; Fox, A.M.; Hudson, A.; et al. Remote sensing of dryland ecosystem structure and function Progress, challenges, and opportunities. Remote Sens. Environ. 2019, 233, 111401. [Google Scholar] [CrossRef]
- Green, J.K.; Seneviratne, S.I.; Berg, A.M.; Findell, K.L.; Hagemann, S.; Lawrence, D.M.; Gentine, P. Large influence of soil moisture on long-term terrestrial carbon uptake. Nature 2019, 565, 476–479. [Google Scholar] [CrossRef]
- Dang, C.; Shao, Z.; Huang, X.; Qian, J.; Cheng, G.; Ding, Q.; Fan, Y. Assessment of the importance of increasing temperature and decreasing soil moisture on global ecosystem productivity using solar-induced chlorophyll fluorescence. Glob. Change Biol. 2022, 28, 2066–2080. [Google Scholar] [CrossRef]
- Liu, X.; Feng, X.; Fu, B. Changes in global terrestrial ecosystem water use efficiency are closely related to soil moisture. Sci. Total Environ. 2020, 698, 134165. [Google Scholar] [CrossRef]
- Peng, J.; Tang, J.; Xie, S.; Wang, Y.; Liao, J.; Chen, C.; Sun, C.; Mao, J.; Zhou, Q.; Niu, S. Evidence for the acclimation of ecosystem photosynthesis to soil moisture. Nat. Commun. 2024, 15, 9795. [Google Scholar] [CrossRef]
- Chang, Q.; He, H.; Ren, X.; Zhang, L.; Feng, L.; Lv, Y.; Zhang, M.; Xu, Q.; Liu, W.; Zhang, Y.; et al. Soil moisture drives the spatiotemporal patterns of asymmetry in vegetation productivity responses across China. Sci. Total Environ. 2023, 855, 158819. [Google Scholar] [CrossRef]
- Sun, W.; Zhou, S.; Yu, B.; Zhang, Y.; Keenan, T.; Fu, B. Soil moisture-atmosphere interactions drive terrestrial carbon-water trade-offs. Commun. Earth Environ. 2025, 6, 169. [Google Scholar] [CrossRef]
- Shi, J.; Liu, M.; Li, Y.; Guan, C. Response of total primary productivity of vegetation to meteorological drought in arid and semi-arid regions of China. J. Arid. Environ. 2025, 228, 105346. [Google Scholar] [CrossRef]
- Xu, S.; Gentine, P.; Li, L.; Wang, L.; Yu, Z.; Dong, N.; Ju, Q.; Zhang, Y. Response of ecosystem productivity to high vapor pressure deficit and low soil moisture: Lessons learned from the global eddy—Covariance observations. Earth’s Future 2023, 11, e2022EF003252. [Google Scholar] [CrossRef]
- Zhao, M.; Heinsch, F.A.; Nemani, R.R.; Running, S.W. Improvements of the MODIS terrestrial gross and net primary production global data set. Remote Sens. Environ. 2005, 95, 164–176. [Google Scholar] [CrossRef]
- Zhang, Y.; Ye, A. Would the obtainable gross primary productivity (GPP) products stand up? A critical assessment of 45 global GPP products. Sci. Total Environ. 2021, 783, 146965. [Google Scholar] [CrossRef]
- Qiu, R.; Han, G.; Ma, X.; Xu, H.; Shi, T.; Zhang, M. A Comparison of OCO-2 SIF, MODIS GPP, and GOSIF Data from Gross Primary Production (GPP) Estimation and Seasonal Cycles in North America. Remote Sens. 2020, 12, 258. [Google Scholar] [CrossRef]
- Frankenberg, C.; Fisher, J.B.; Worden, J.; Badgley, G.; Saatchi, S.S.; Lee, J.-E.; Toon, G.C.; Butz, A.; Jung, M.; Kuze, A.; et al. New global observations of the terrestrial carbon cycle from GOSAT: Patterns of plant fluorescence with gross primary productivity. Geophys. Res. Lett. 2011, 38, 17. [Google Scholar] [CrossRef]
- Cai, Y.; Zhang, F.; Duan, P.; Jim, C.Y.; Chan, N.W.; Shi, J.; Liu, C.; Wang, J.; Bahtebay, J.; Ma, X. Vegetation cover changes in China induced by ecological restoration-protection projects and land-use changes from 2000 to 2020. Catena 2022, 217, 106530. [Google Scholar] [CrossRef]
- Zhu, B.; Zhang, Q.; Yang, J.-H.; Li, C.-H. Response of Potential Evapotranspiration to Warming and Wetting in Northwest China. Atmosphere 2022, 13, 353. [Google Scholar] [CrossRef]
- Niu, J.Q.; Liu, Z.J.; Chen, F.Y.; Liu, G.J.; Zhou, J.L.; Zhou, P.; Li, H.R.; Li, M.Y. Variations of soil moisture and its influencing factors in arid and semi-arid areas, China. J. Arid. Land 2025, 17, 624–643. [Google Scholar] [CrossRef]
- Li, X.; Xiao, J. Mapping Photosynthesis Solely from Solar-Induced Chlorophyll Fluorescence: A Global, Fine-Resolution Dataset of Gross Primary Production Derived from OCO-2. Remote Sens. 2019, 11, 2563. [Google Scholar] [CrossRef]
- Xiao, J.; Li, X.; He, B.; Arain, M.A.; Beringer, J.; Desai, A.R.; Emmel, C.; Hollinger, D.Y.; Krasnova, A.; Mammarella, I.; et al. Solar-induced chlorophyll fluorescence exhibits a universal relationship with gross primary productivity across a wide variety of biomes. Glob. Change Biol. 2019, 25, e4–e6. [Google Scholar] [CrossRef]
- Baia, J.; Zhanga, H.; Suna, R.; Lic, X.; Xiao, J.; Wang, Y. Estimation of global GPP from GOME-2 and OCO-2 SIF by considering the dynamic variations of GPP-SIF relationship. Agric. For. Meteorol. 2022, 326, 109180. [Google Scholar] [CrossRef]
- Qi, G.; She, D.; Xia, J.; Song, J.; Jiao, W.; Li, J.; Liu, Z. Soil moisture plays an increasingly important role in constraining vegetation productivity in China over the past two decades. Agric. For. Meteorol. 2024, 356, 110193. [Google Scholar] [CrossRef]
- Ran, Y.H.; Li, X.; Lu, L.; Li, Z.Y. Large-scale land cover mapping with the integration of multi-source information based on the Dempster–Shafer theory. Int. J. Geogr. Inf. Sci. 2012, 26, 169–191. [Google Scholar] [CrossRef]
- Miralles, D.G.; Bonte, O.; Koppa, A.; Baez-Villanueva, O.M.; Tronquo, E.; Zhong, F.; Beck, H.E.; Hulsman, P.; Dorigo, W.; Verhoest, N.E.C.; et al. GLEAM4: Global land evaporation and soil moisture dataset at 0.1 resolution from 1980 to near present. Sci. Data 2025, 12, 416. [Google Scholar] [CrossRef]
- Yang, S.; Zeng, J.; Fan, W.; Cui, Y. Evaluating Root-Zone Soil Moisture Products from GLEAM, GLDAS, and ERA5 Based on In Situ Observations and Triple Collocation Method over the Tibetan Plateau. J. Hydrometeorol. 2022, 23, 1861–1878. [Google Scholar] [CrossRef]
- Liu, J.; Zhao, J.; He, J.; Qu, J.; Xing, Y.; Du, R.; Chen, S.; Tang, X.; Wang, L.; Yue, C. Spatiotemporal Trend and Influencing Factors of Surface Soil Moisture in Eurasian Drylands over the Past Four Decades. Forests 2025, 16, 589. [Google Scholar] [CrossRef]
- Li, H.; Liu, F.; Zhang, S.; Zhang, C.; Zhang, C.; Ma, W.; Luo, J. Drying–Wetting Changes of Surface Soil Moisture and the Influencing Factors in Permafrost Regions of the Qinghai-Tibet Plateau, China. Remote Sens. 2022, 14, 2915. [Google Scholar] [CrossRef]
- Funk, C.; Peterson, P.; Landsfeld, M.; Pedreros, D.; Verdin, J.; Shukla, S.; Husak, G.; Rowland, J.; Harrison, L.; Hoell, A.; et al. The climate hazards infrared precipitation with stations--a new environmental record for monitoring extremes. Sci. Data 2015, 2, 150066. [Google Scholar] [CrossRef]
- Gao, F.; Zhang, Y.; Ren, X.; Yao, Y.; Hao, Z.; Cai, W. Evaluation of CHIRPS and its application for drought monitoring over the Haihe River Basin, China. Nat. Hazards 2018, 92, 155–172. [Google Scholar] [CrossRef]
- Du, H.; Tan, M.L.; Zhang, F.; Chun, K.P.; Li, L.; Kabir, M.H. Evaluating the effectiveness of CHIRPS data for hydroclimatic studies. Theor. Appl. Climatol. 2023, 155, 1519–1539. [Google Scholar] [CrossRef]
- Ocampo-Marulanda, C.; Fernández-Álvarez, C.; Cerón, W.L.; Canchala, T.; Carvajal-Escobar, Y.; Alfonso-Morales, W. A spatiotemporal assessment of the high-resolution CHIRPS rainfall dataset in southwestern Colombia using combined principal component analysis. Ain Shams Eng. J. 2022, 13, 101739. [Google Scholar] [CrossRef]
- Sen, P.K. Estimates of the Regression Coefficient Based on Kendall’s Tau. J. Am. Stat. Assoc. 1968, 63, 1379–1389. [Google Scholar] [CrossRef]
- Ali, A.; Lebel, T. The Sahelian standardized rainfall index revisited. Int. J. Climatol. 2008, 29, 1705–1714. [Google Scholar] [CrossRef]
- Nguyen, H.; Wheeler, M.C.; Otkin, J.A.; Cowan, T.; Frost, A.; Stone, R. Using the evaporative stress index to monitor flash drought in Australia. Environ. Res. Lett. 2019, 14, 064016. [Google Scholar] [CrossRef]
- Deng, Y.; Wang, X.; Wang, K.; Ciais, P.; Tang, S.; Jin, L.; Li, L.; Piao, S. Responses of vegetation greenness and carbon cycle to extreme droughts in China. Agric. For. Meteorol. 2021, 298-299, 108307. [Google Scholar] [CrossRef]
- Kang, J.; Jin, R.; Li, X.; Zhang, Y.; Zhu, Z. Spatial Upscaling of Sparse Soil Moisture Observations Based on Ridge Regression. Remote Sens. 2018, 10, 192. [Google Scholar] [CrossRef]
- Wang, Z.; Peñuelas, J.; Tagesson, T.; Smith, W.K.; Wu, M.; He, W.; Sitch, S.; Wang, S. Evolution of Global Terrestrial Gross Primary Productivity Trend. Ecosyst. Health Sustain. 2024, 10, 0278. [Google Scholar] [CrossRef]
- Wang, S.; Fu, B.; Wei, F.; Piao, S.; Maestre, F.T.; Wang, L.; Jiao, W.; Liu, Y.; Li, Y.; Li, C.; et al. Drylands contribute disproportionately to observed global productivity increases. Sci. Bull. 2023, 68, 224–232. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Shi, H. Temporal and Spatial Variation Characteristics of Soil Moisture in Spring in the Arid Regions of Northwest China in the Past 60s years. J. Geosci. Environ. Prot. 2022, 10, 273–282. [Google Scholar] [CrossRef]
- Xu, L.; Zheng, C.; Ma, Y. Variations in precipitation extremes in the arid and semi-arid regions of China. Int. J. Climatol. 2020, 41, 1542–1554. [Google Scholar] [CrossRef]
- Shang, K.; Liu, X.; Xie, X.; Sha, Y.; Zhao, X.; Liu, J.; Wang, A. Synergistic Impact of Midlatitude Westerly and East Asian Summer Monsoon on Mid-Summer Precipitation in North China. Atmosphere 2025, 16, 658. [Google Scholar] [CrossRef]
- Zhang, G.; Nan, Z.; Zhao, L.; Liang, Y.; Cheng, G. Qinghai-Tibet Plateau wetting reduces permafrost thermal responses to climate warming. Earth Planet. Sci. Lett. 2021, 562, 116858. [Google Scholar] [CrossRef]
- Sun, Y.; Zhao, G.; Zheng, Z.; Zhu, Y.; Zhu, J.; Di, Y.; Gao, J.; Cai, M.; Zhang, Y. Wetting-warming climate increases ecosystem carbon use efficiency of Chinese arid and semi-arid ecosystem. Agric. For. Meteorol. 2024, 351, 110018. [Google Scholar] [CrossRef]
- Yu, T.; Jiapaer, G.; Bao, A.; Yuan, Y.; Bao, J.; Van de Voorde, T. Soil moisture dominates gross primary productivity variation during severe droughts in Central Asia. Ecol. Inform. 2025, 86, 103076. [Google Scholar] [CrossRef]
- Zhao, H.; Jia, G.; Xu, X.; Zhang, A.; Wang, H. Divergent effects of intensified precipitation on primary production in global drylands. Sci. Total Environ. 2023, 892, 164736. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Wang, L.; Jiang, Q.; Wang, Z. Sensitivity of gross primary production to precipitation and the driving factors in China’s agricultural ecosystems. Sci. Total Environ. 2024, 948, 174938. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Chen, J.; Lee, S.-C.; Xiong, L.; Su, T.; Lin, Q.; Xu, C.-Y. Response and recovery times of vegetation productivity under drought stress: Dominant factors and relationships. J. Hydrol. 2025, 655, 132945. [Google Scholar] [CrossRef]
- Xu, S.; Ji, X.; Jin, B.; Zhang, J. Root distribution of three dominant desert shrubs and their water uptake dynamics. J. Plant Ecol. 2016, 10, 780–790. [Google Scholar] [CrossRef]
- Xue, S.; Wu, G. A Study of the Dependence between Soil Moisture and Precipitation in different Ecoregions of the Northern Hemisphere. EGUsphere 2025, 2025, 1–39. [Google Scholar]
- Prudhomme, C.; Giuntoli, I.; Robinson, E.L.; Clark, D.B.; Arnell, N.W.; Dankers, R.; Fekete, B.M.; Franssen, W.; Gerten, D.; Gosling, S.N.; et al. Hydrological droughts in the 21st century, hotspots and uncertainties from a global multimodel ensemble experiment. Proc. Natl. Acad. Sci. USA 2014, 111, 3262–3267. [Google Scholar] [CrossRef]
- Dai, Y.; Zeng, X.; Dickinson, R.E.; Baker, I.; Bonan, G.B.; Bosilovich, M.G.; Denning, A.S.; Dirmeyer, P.A.; Houser, P.R.; Niu, G. The common land model. Bull. Am. Meteorol. Soc. 2003, 84, 1013–1024. [Google Scholar] [CrossRef]
- Huang, J.; Yu, H.; Guan, X.; Wang, G.; Guo, R. Accelerated dryland expansion under climate change. Nat. Clim. Change 2015, 6, 166–171. [Google Scholar] [CrossRef]
- Yang, S.; Chen, K.; Zhu, B.; Tian, Y.; Zeng, Z.; Liu, M.; Zheng, C. How does irrigation alter the water, carbon, and nitrogen budgets in a large endorheic river basin? J. Hydrol. 2022, 613, 128317. [Google Scholar] [CrossRef]
- Wang, X.; Chen, G.; Wu, Q.; Cao, L.; Awange, J.; Song, Y.; Wu, M. Spatio-temporal patterns and drivers of carbon–water coupling in frozen soil zones across the gradients of freezing over the Qinghai-Tibet Plateau. J. Hydrol. 2023, 621, 129674. [Google Scholar] [CrossRef]
- Liao, D.; Niu, J.; Ciais, P.; Du, T.; Zhang, B.; Kang, S. Changing Climate Threatens Irrigation Benefits of Maize Gross Primary Productivity in China. Earth’s Future 2024, 12, e2022EF003474. [Google Scholar] [CrossRef]
- Zhou, T.; Yang, H.; Qiu, X.; Sun, H.; Song, P.; Yang, W. China’s grassland ecological compensation policy achieves win-win goals in Inner Mongolia. Environ. Res. Commun. 2023, 5, 031007. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | Influence | ASA | GLS | F | BOSVL | C | W |
|---|---|---|---|---|---|---|---|
| ΔSAGPP(SARSM|SAPRE) | Positive | 78.12 | 79.19 | 74.69 | 77.91 | 76.74 | 75.54 |
| Negative | 21.88 | 20.81 | 25.31 | 22.09 | 23.26 | 24.46 | |
| ΔSAGPP(SAPRE|SARSM) | Positive | 65.53 | 66.28 | 62.22 | 64.48 | 64.23 | 61.16 |
| Negative | 34.47 | 33.72 | 37.78 | 35.52 | 35.77 | 38.84 | |
| ΔSAGPP(SARSM|SAPRE) − ΔSAGPP(SAPRE|SARSM) | Positive | 62.13 | 61.24 | 65.11 | 62.97 | 61.79 | 66.97 |
| Negative | 37.87 | 38.76 | 34.89 | 37.21 | 38.21 | 33.03 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Z.; Lin, H.; Li, H.; Li, M.; Zhou, P.; Wang, Z.; Niu, J. Response Mechanisms of Vegetation Productivity to Water Variability in Arid and Semi-Arid Areas of China: A Decoupling Analysis of Soil Moisture and Precipitation. Atmosphere 2025, 16, 933. https://doi.org/10.3390/atmos16080933
Liu Z, Lin H, Li H, Li M, Zhou P, Wang Z, Niu J. Response Mechanisms of Vegetation Productivity to Water Variability in Arid and Semi-Arid Areas of China: A Decoupling Analysis of Soil Moisture and Precipitation. Atmosphere. 2025; 16(8):933. https://doi.org/10.3390/atmos16080933
Chicago/Turabian StyleLiu, Zijian, Hao Lin, Hongrui Li, Mengyang Li, Peng Zhou, Ziyu Wang, and Jiqiang Niu. 2025. "Response Mechanisms of Vegetation Productivity to Water Variability in Arid and Semi-Arid Areas of China: A Decoupling Analysis of Soil Moisture and Precipitation" Atmosphere 16, no. 8: 933. https://doi.org/10.3390/atmos16080933
APA StyleLiu, Z., Lin, H., Li, H., Li, M., Zhou, P., Wang, Z., & Niu, J. (2025). Response Mechanisms of Vegetation Productivity to Water Variability in Arid and Semi-Arid Areas of China: A Decoupling Analysis of Soil Moisture and Precipitation. Atmosphere, 16(8), 933. https://doi.org/10.3390/atmos16080933