
The Productivity and Carbon Exchange of an Intensively Managed Pasture in Central Kentucky



Abstract
1. Introduction
2. Materials and Methods
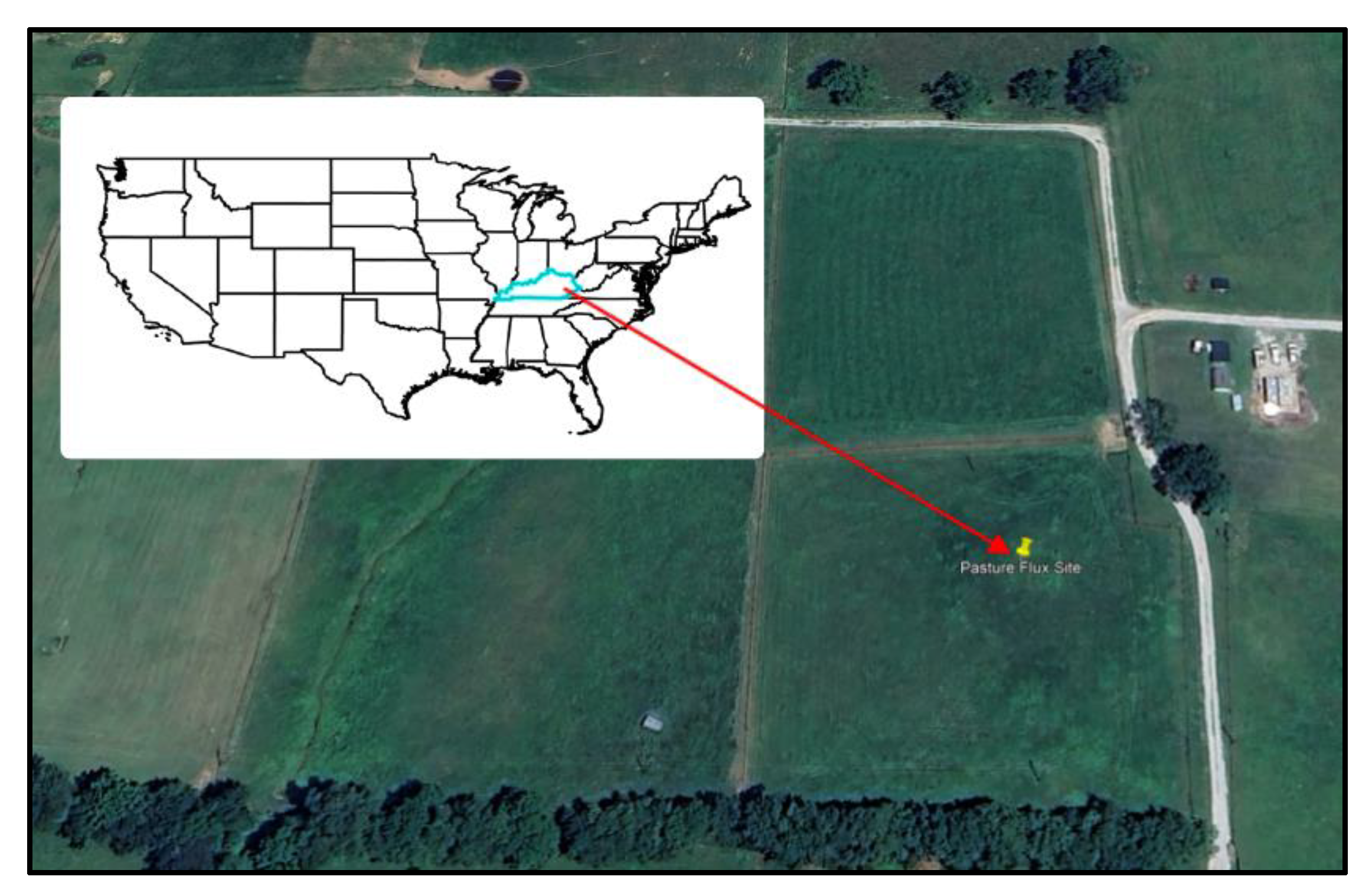
2.1. Site Characterization
2.2. Principles of Eddy Covariance
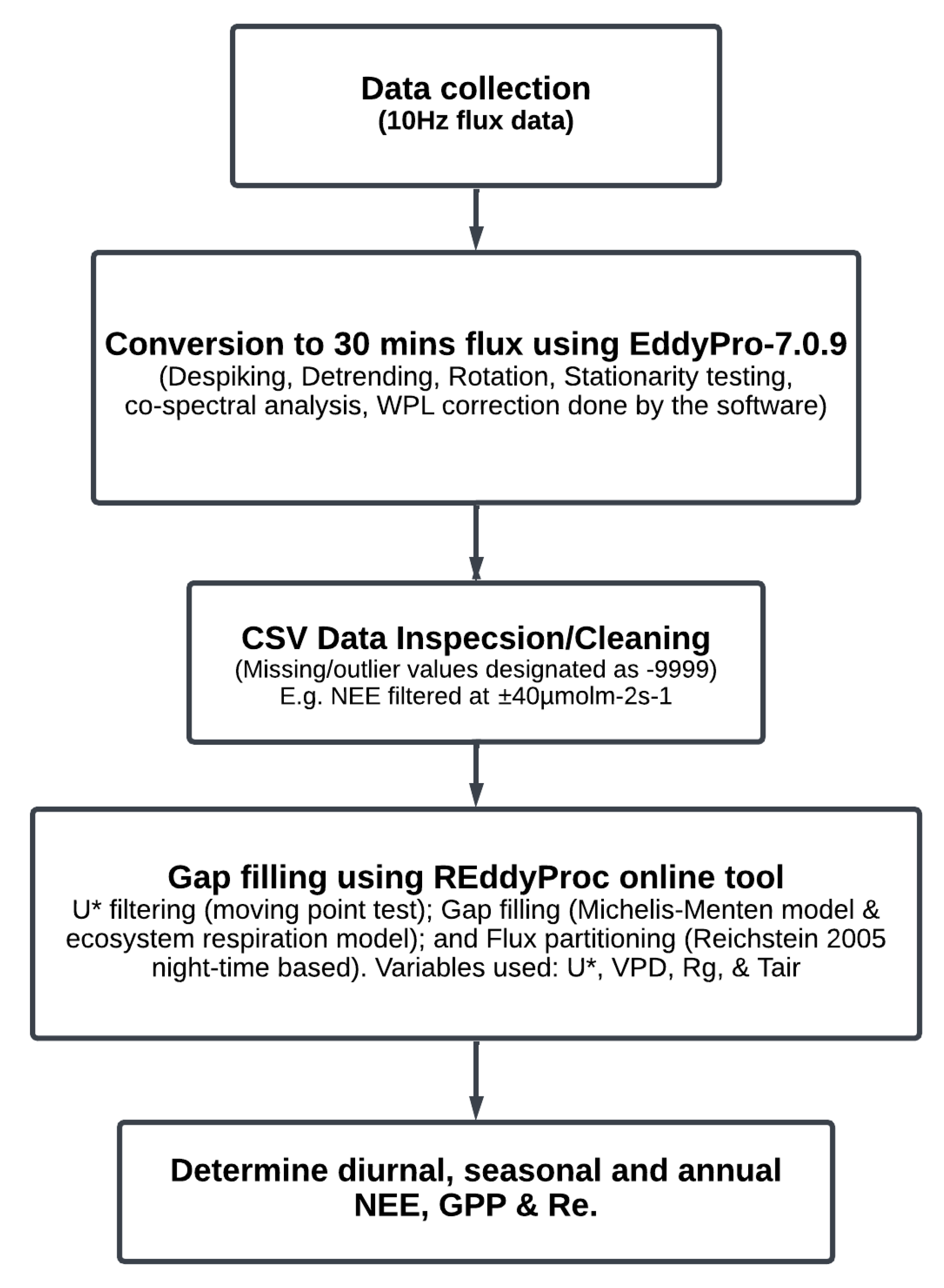
2.3. Data Collection
2.4. Data Filtering, Gap Filling, and Flux Correction
3. Results
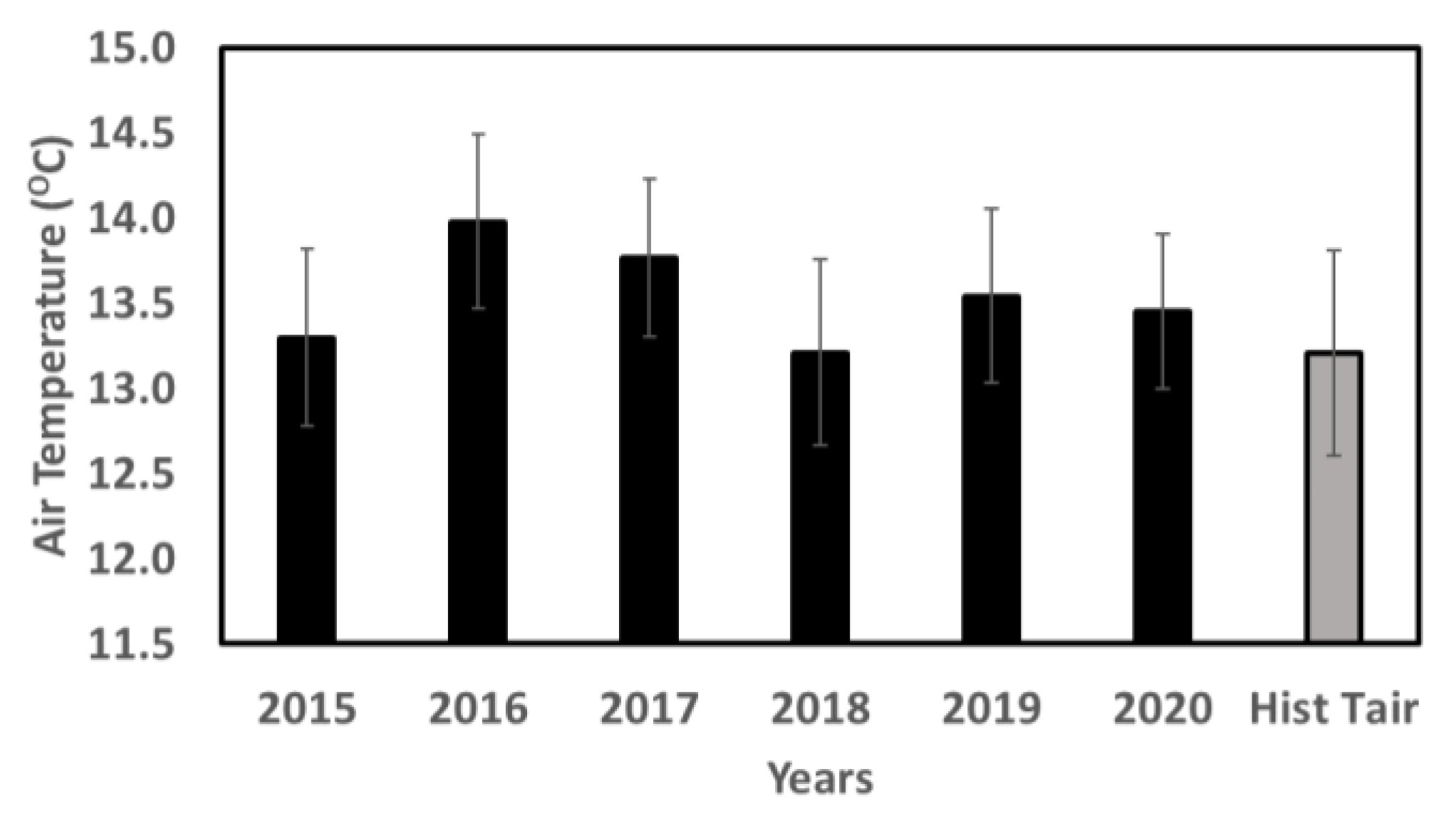
3.1. Meteorological Conditions in 2015–2020
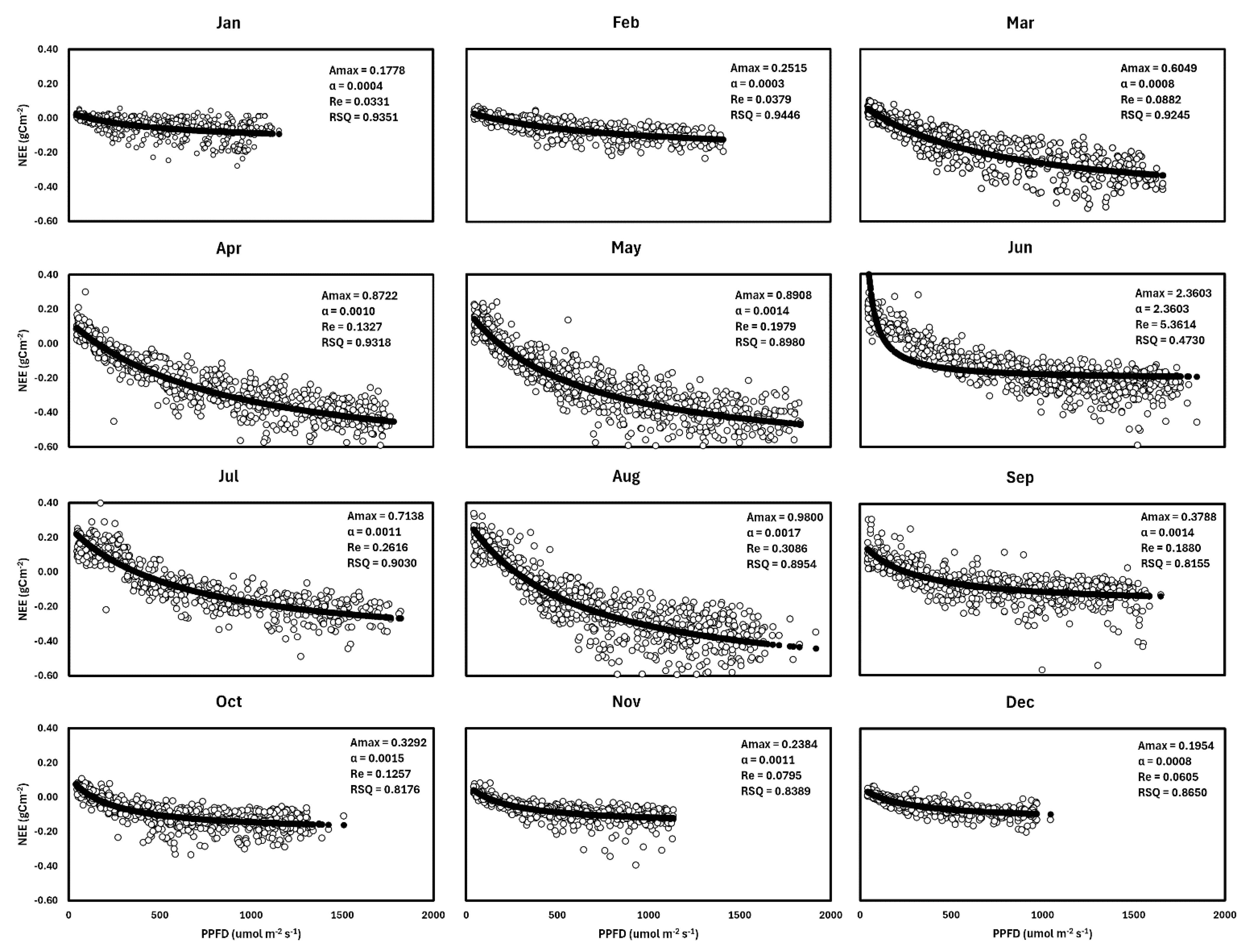
3.2. Light Response Curve
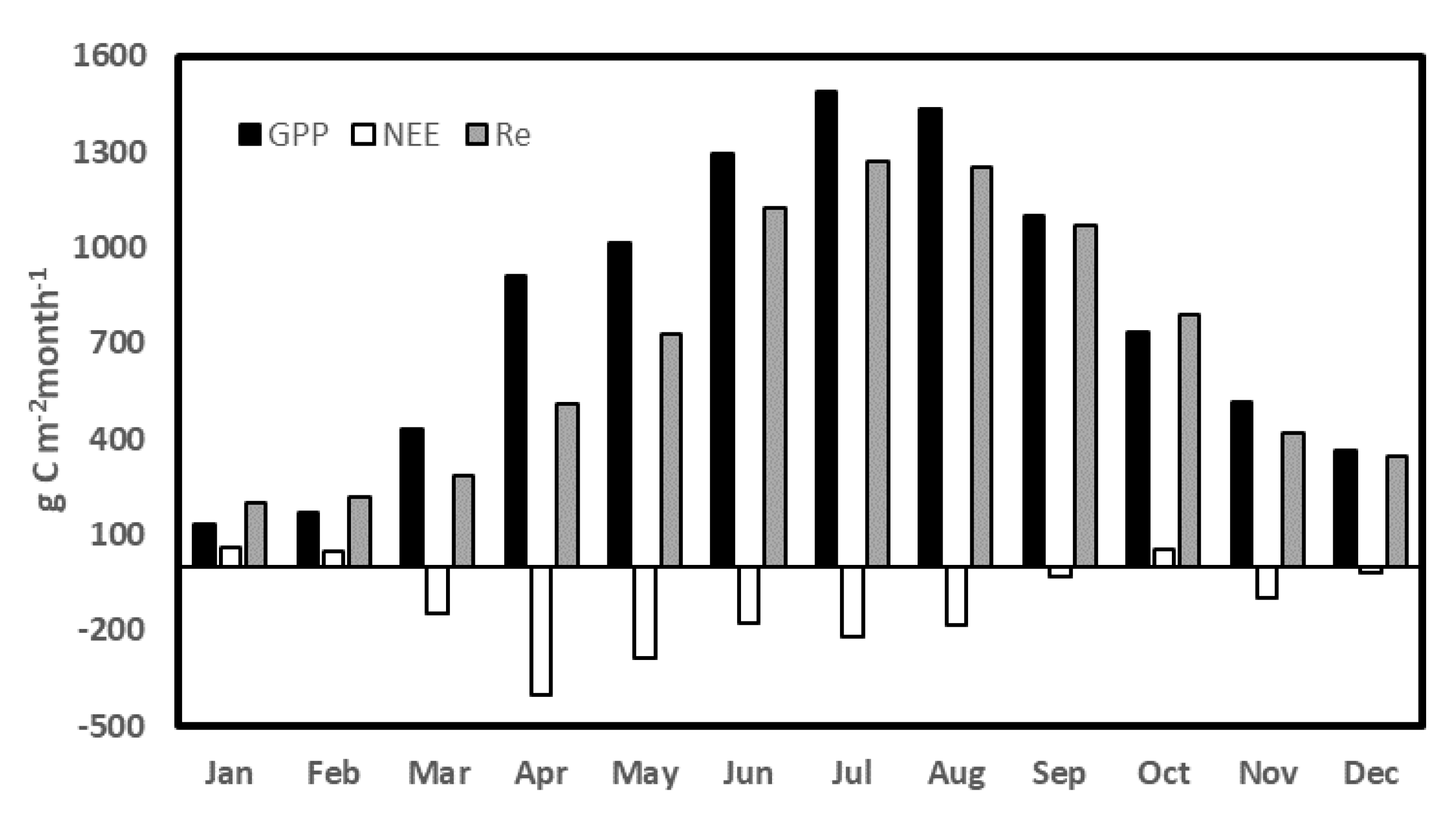
3.3. Daily, Diurnal and Seasonal Variations
3.3.1. Daily and Diurnal NEE
3.3.2. Seasonal NEE
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAO. Emissions from Agriculture and Forest Land: Global, Regional and Country Trends 1990–2019; FAO: Rome, Italy, 2021. [Google Scholar]
- Ritchie, H. How Much of the World’s Land Would We Need in Order to Feed the Global Population with the Average Diet of a Given Country; Our World in Data: Oxford, UK, 2017. [Google Scholar]
- FAO. The State of World Fisheries and Aquaculture 2020. In Sustainability in Action; FAO: Rome, Italy, 2020. [Google Scholar] [CrossRef]
- Willett, W.; Rockström, J.; Loken, B.; Springmann, M.; Lang, T.; Vermeulen, S.; Garnett, T.; Tilman, D.; DeClerck, F.; Wood, A.; et al. Food in the Anthropocene: The EAT–Lancet Commission on Healthy Diets from Sustainable Food Systems. Lancet 2019, 393, 447–492. [Google Scholar] [CrossRef]
- Farmland Innovation Center. Kentucky Data and Statistics. Available online: https://farmlandinfo.org/statistics/kentucky-statistics/ (accessed on 9 February 2024).
- Searchinger, T.; Herrero, M.; Yan, X.; Wang, J.; Beauchemin, K.; Kebreab, E. Opportunities to Reduce Methane Emissions from Global Agriculture; Faculty of Cornell University: Ithaca, NY, USA; Faculty of Princeton University: Princeton, NJ, USA, 2021. [Google Scholar]
- Jordon, M.W.; Buffet, J.C.; Dungait, J.A.J.; Galdos, M.V.; Garnett, T.; Lee, M.R.F.; Lynch, J.; Röös, E.; Searchinger, T.D.; Smith, P.; et al. A Restatement of the Natural Science Evidence Base Concerning Grassland Management, Grazing Livestock and Soil Carbon Storage. Proc. R. Soc. B 2024, 291, 20232669. [Google Scholar] [CrossRef] [PubMed]
- Steinfeld, H.; Gerber, P. Livestock Production and the Global Environment: Consume Less or Produce Better? Proc. Natl. Acad. Sci. USA 2010, 107, 18237–18238. [Google Scholar] [CrossRef] [PubMed]
- Henchion, M.; Moloney, A.P.; Hyland, J.; Zimmermann, J.; McCarthy, S. Review: Trends for Meat, Milk and Egg Consumption for the next Decades and the Role Played by Livestock Systems in the Global Production of Proteins. Animal 2021, 15, 100287. [Google Scholar] [CrossRef] [PubMed]
- Teague, R.; Kreuter, U. Managing Grazing to Restore Soil Health, Ecosystem Function, and Ecosystem Services. Front. Sustain. Food Syst. 2020, 4, 534187. [Google Scholar] [CrossRef]
- Baggio, R.; Overbeck, G.E.; Durigan, G.; Pillar, V.D. To Graze or Not to Graze: A Core Question for Conservation and Sustainable Use of Grassy Ecosystems in Brazil. Perspect. Ecol. Conserv. 2021, 19, 256–266. [Google Scholar] [CrossRef]
- Kgosikoma, E. Grazing Management Systems and Their Effects on Savanna Ecosystem Dynamics: A Review. J. Ecol. Nat. Environ. 2013, 5, 88–94. [Google Scholar] [CrossRef]
- Asner, G.P.; Elmore, A.J.; Olander, L.P.; Martin, R.E.; Harris, T. Grazing Systems, Ecosystem Responses, and Global Change. Annu. Rev. Environ. Resour. 2004, 29, 261–299. [Google Scholar] [CrossRef]
- Liebig, M.A.; Gross, J.R.; Kronberg, S.L.; Hanson, J.D.; Frank, A.B.; Phillips, R.L. Soil Response to Long-Term Grazing in the Northern Great Plains of North America. Agric. Ecosyst. Environ. 2006, 115, 270–276. [Google Scholar] [CrossRef]
- Post, W.M.; Emanuel, W.R.; Zinke, P.J.; Stangenberger, A.G. Soil Carbon Pools and World Life Zones. Nature 1982, 298, 156–159. [Google Scholar] [CrossRef]
- Eswaran, H.; Van Den Berg, E.; Reich, P. Organic Carbon in Soils of the World. Soil Sci. Soc. Am. J. 1993, 57, 192–194. [Google Scholar] [CrossRef]
- USDA. Crop Production 2012 Summary; No. January; National Agricultural Statistics Service: Washington, DC, USA, 2013.
- Silveira, M.; Hanlon, E.; Azenha, M.; Da Silva, H.M. Carbon Sequestration in Grazing Land Ecosystems. EDIS 2012, 2012, 1–4. [Google Scholar] [CrossRef]
- Conant, R.T.; Paustian, K.; Elliott, E.T. Grassland Management and Conversion into Grassland: Effects on Soil Carbon. Ecol. Appl. 2001, 11, 343–355. [Google Scholar] [CrossRef]
- Conant, R.T. Grassland Soil Organic Carbon Stocks: Status, Opportunities, Vulnerability. In Recarbonization of the Biosphere: Ecosystems and the Global Carbon Cycle; Springer: Berlin/Heidelberg, Germany, 2012. [Google Scholar] [CrossRef]
- Follett, R.F.; Kimble, J.M. The Potential of US Grazing Lands to Sequester Carbon and Mitigate the Greenhouse Effect; CRC Press: Boca Raton, FL, USA, 2000. [Google Scholar]
- Teague, W.R.; Apfelbaum, S.; Lal, R.; Kreuter, U.P.; Rowntree, J.; Davies, C.A.; Conser, R.; Rasmussen, M.; Hatfeld, J.; Wang, T.; et al. The Role of Ruminants in Reducing Agriculture’s Carbon Footprint in North America. J. Soil Water Conserv. 2016, 71, 156–164. [Google Scholar] [CrossRef]
- Henderson, B.B.; Gerber, P.J.; Hilinski, T.E.; Falcucci, A.; Ojima, D.S.; Salvatore, M.; Conant, R.T. Greenhouse Gas Mitigation Potential of the World’s Grazing Lands: Modeling Soil Carbon and Nitrogen Fluxes of Mitigation Practices. Agric. Ecosyst. Environ. 2015, 207, 91–100. [Google Scholar] [CrossRef]
- Gilmanov, T.G.; Verma, S.B.; Sims, P.L.; Meyers, T.P.; Bradford, J.A.; Burba, G.G.; Suyker, A.E. Gross Primary Production and Light Response Parameters of Four Southern Plains Ecosystems Estimated Using Long-Term CO2-Flux Tower Measurements. Glob. Biogeochem. Cycles 2003, 17, 1071. [Google Scholar] [CrossRef]
- Flanagan, L.B.; Wever, L.A.; Carlson, P.J. Seasonal and Interannual Variation in Carbon Dioxide Exchange and Carbon Balance in a Northern Temperate Grassland. Glob. Chang. Biol. 2002, 8, 599–615. [Google Scholar] [CrossRef]
- Dormaar, J.F.; Adams, B.W.; Willms, W.D. Impacts of Rotational Grazing on Mixed Prairie Soils and Vegetation. J. Range Manag. 1997, 50, 647. [Google Scholar] [CrossRef]
- Pineiro, G.; Paruelo, J.M.; Oesterheld, M.; Jobbágy, E.G. Pathways of Grazing Effects on Soil Organic Carbon and Nitrogen. Rangel. Ecol. Manag. 2010, 63, 109–119. [Google Scholar] [CrossRef]
- Reichstein, M.; Bahn, M.; Ciais, P.; Frank, D.; Mahecha, M.D.; Seneviratne, S.I.; Zscheischler, J.; Beer, C.; Buchmann, N.; Frank, D.C.; et al. Climate Extremes and the Carbon Cycle. Nature 2013, 500, 287–295. [Google Scholar] [CrossRef]
- Reverter, B.R.; Sánchez-Cañete, E.P.; Resco, V.; Serrano-Ortiz, P.; Oyonarte, C.; Kowalski, A.S. Analyzing the Major Drivers of NEE in a Mediterranean Alpine Shrubland. Biogeosciences 2010, 7, 2601–2611. [Google Scholar] [CrossRef]
- Jiao, L.; Lu, N.; Sun, G.; Ward, E.J.; Fu, B. Biophysical Controls on Canopy Transpiration in a Black Locust (Robinia Pseudoacacia) Plantation on the Semi-Arid Loess Plateau, China. Ecohydrology 2015, 9, 1068–1081. [Google Scholar] [CrossRef]
- Flanagan, L.B.; Johnson, B.G. Interacting Effects of Temperature, Soil Moisture and Plant Biomass Production on Ecosystem Respiration in a Northern Temperate Grassland. Agric. For. Meteorol. 2005, 130, 237–253. [Google Scholar] [CrossRef]
- Jones, L.-A.; Madani, N.; Kimball, J.-S.; Reichle, R.-H.; Colliander, A. Effects of Recent Regional Soil Moisture Variability on Global Net Ecosystem CO2 Exchange. In AGU Fall Meeting Abstracts; American Geophysical Union: Washington, DC, USA, 2017; Volume 2017, p. H21I-1586. [Google Scholar]
- Jung, M.; Reichstein, M.; Schwalm, C.R.; Huntingford, C.; Sitch, S.; Ahlström, A.; Arneth, A.; Camps-Valls, G.; Ciais, P.; Friedlingstein, P.; et al. Compensatory Water Effects Link Yearly Global Land CO2 Sink Changes to Temperature. Nature 2017, 541, 516–520. [Google Scholar] [CrossRef]
- Lu, Y.; Murase, J.; Watanabe, A.; Sugimoto, A.; Kimura, M. Linking Microbial Community Dynamics to Rhizosphere Carbon Flow in a Wetland Rice Soil. FEMS Microbiol. Ecol. 2004, 48, 179–186. [Google Scholar] [CrossRef]
- Chakravarthy, D.; Raghavendra, H.V.; Ratnam, J.; Sankaran, M. Soil Respiration Is Correlated with Rainfall and Soil Moisture at Multiple Temporal Scales in a Seasonal Wet Tropical Forest. bioRxiv 2024. [Google Scholar] [CrossRef]
- Baldocchi, D.D.; Hicks, B.B.; Meyers, T.P. Measuring Biosphere-Atmosphere Exchanges of Biologically Related Gases with Micrometeorological Methods. Ecology 1988, 69, 1331–1340. [Google Scholar] [CrossRef]
- Moncrieff, J.B.; Massheder, J.M.; De Bruin, H.; Elbers, J.; Friborg, T.; Heusinkveld, B.; Kabat, P.; Scott, S.; Soegaard, H.; Verhoef, A. A System to Measure Surface Fluxes of Momentum, Sensible Heat, Water Vapour and Carbon Dioxide. J. Hydrol. 1997, 188–189, 589–611. [Google Scholar] [CrossRef]
- Baldocchi, D.D. Assessing the Eddy Covariance Technique for Evaluating Carbon Dioxide Exchange Rates of Ecosystems: Past, Present and Future. Glob. Chang. Biol. 2003, 9, 479–492. [Google Scholar] [CrossRef]
- Law, B.E.; Falge, E.; Gu, L.; Baldocchi, D.D.; Bakwin, P.; Berbigier, P.; Davis, K.; Dolman, A.J.; Falk, M.; Fuentes, J.D.; et al. Environmental Controls over Carbon Dioxide and Water Vapor Exchange of Terrestrial Vegetation. Agric. For. Meteorol. 2002, 113, 97–120. [Google Scholar] [CrossRef]
- Verma, S.B.; Dobermann, A.; Cassman, K.G.; Walters, D.T.; Knops, J.M.; Arkebauer, T.J.; Suyker, A.E.; Burba, G.G.; Amos, B.; Yang, H.; et al. Annual Carbon Dioxide Exchange in Irrigated and Rainfed Maize-Based Agroecosystems. Agric. For. Meteorol. 2005, 131, 77–96. [Google Scholar] [CrossRef]
- Suyker, A.E.; Verma, S.B. Coupling of Carbon Dioxide and Water Vapor Exchanges of Irrigated and Rainfed Maize-Soybean Cropping Systems and Water Productivity. Agric. For. Meteorol. 2010, 150, 553–563. [Google Scholar] [CrossRef]
- Reynolds, O., IV. On the Dynamical Theory of Incompressible Viscous Fluids and the Determination of the Criterion. Philos. Trans. R. Soc. Lond. A 1895, 186, 123–164. [Google Scholar] [CrossRef]
- Derbyshire, S.H. Atmospheric Boundary Layer Flows—Their Structure and Measurement. by J. C. Kaimal and J. J. Finnigan. Oxford University Press. 1994. Pp. 289. Price £50.00 (Hardback). Isbn 0 19 506239 6. Q. J. R. Meteorol. Soc. 1995, 121, 1177. [Google Scholar] [CrossRef]
- Raupach, M.R.; Finnigan, J.J.; Brunet, Y. Coherent Eddies and Turbulence in Vegetation Canopies: The Mixing-Layer Analogy. Boundary-Layer Meteorol 1996, 78, 351–382. [Google Scholar] [CrossRef]
- Goulden, M.L.; Daube, B.C.; Fan, S.M.; Sutton, D.J.; Bazzaz, A.; Munger, J.W.; Wofsy, S.C. Physiological Responses of a Black Spruce Forest to Weather. J. Geophys. Res. Atmos. 1997, 102, 28987–28996. [Google Scholar] [CrossRef]
- Mauder, M.; Foken, T. Documentation and Instruction Manual of the Eddy-Covariance Software Package TK3 (Update); University of Bayreuth: Bayreuth, Germany, 2015. [Google Scholar]
- Lee, X.; Massman, W.; Law, B. Handbook of Micrometeorology A Guide for Surface Flux Measurement and Analysis; Springer: Berlin/Heidelberg, Germany, 2005. [Google Scholar]
- Falge, E.; Baldocchi, D.; Olson, R.; Anthoni, P.; Aubinet, M.; Bernhofer, C.; Burba, G.; Ceulemans, R.; Clement, R.; Dolman, H.; et al. Gap Filling Strategies for Defensible Annual Sums of Net Ecosystem Exchange. Agric. For. Meteorol. 2001, 107, 43–69. [Google Scholar] [CrossRef]
- Reichstein, M.; Falge, E.; Baldocchi, D.; Papale, D.; Aubinet, M.; Berbigier, P.; Bernhofer, C.; Buchmann, N.; Gilmanov, T.; Granier, A.; et al. On the Separation of Net Ecosystem Exchange into Assimilation and Ecosystem Respiration: Review and Improved Algorithm. Glob. Chang. Biol. 2005, 11, 1424–1439. [Google Scholar] [CrossRef]
- Schulze, E.D.; Lange, O.L.; Ziegler, H.; Gebauer, G. Carbon and Nitrogen Isotope Ratios of Mistletoes Growing on Nitrogen and Non-Nitrogen Fixing Hosts and on CAM Plants in the Namib Desert Confirm Partial Heterotrophy. Oecologia 1991, 88, 457–462. [Google Scholar] [CrossRef]
- Braswell, B.H.; Sacks, W.J.; Linder, E.; Schimel, D.S. Estimating Diurnal to Annual Ecosystem Parameters by Synthesis of a Carbon Flux Model with Eddy Covariance Net Ecosystem Exchange Observations. Glob. Chang. Biol. 2005, 11, 335–355. [Google Scholar] [CrossRef]
- Zhou, G.; Zhou, X.; He, Y.; Shao, J.; Hu, Z.; Liu, R.; Zhou, H.; Hosseinibai, S. Grazing Intensity Significantly Affects Belowground Carbon and Nitrogen Cycling in Grassland Ecosystems: A Meta-Analysis. Glob. Chang. Biol. 2016, 23, 1167–1179. [Google Scholar] [CrossRef] [PubMed]
- Von Randow, G. When the Centre Becomes Radical. JASSS 2003, 6, 28. [Google Scholar]
- Gaumont-Guay, D.; Black, T.A.; Griffis, T.J.; Barr, A.G.; Jassal, R.S.; Nesic, Z. Interpreting the Dependence of Soil Respiration on Soil Temperature and Water Content in a Boreal Aspen Stand. Agric. For. Meteorol. 2006, 140, 220–235. [Google Scholar] [CrossRef]
- Gebremedhin, M.T.; Loescher, H.W.; Tsegaye, T.D. Carbon Balance of No-till Soybean with Winter Wheat Cover Crop in the Southeastern United States. Agron. J. 2012, 104, 1321–1335. [Google Scholar] [CrossRef]
- Wen, X.F.; Yu, G.R.; Sun, X.M.; Li, Q.K.; Liu, Y.F.; Zhang, L.M.; Ren, C.Y.; Fu, Y.L.; Li, Z.Q. Soil Moisture Effect on the Temperature Dependence of Ecosystem Respiration in a Subtropical Pinus Plantation of Southeastern China. Agric. For. Meteorol. 2006, 137, 166–175. [Google Scholar] [CrossRef]
- Omidiran, Y.A. Net Ecosystem Exchange and Air Pollution in North Alabama. Ph.D. Thesis, Alabama Agricultural and Mechanical University, Huntsville, AL, USA, 2015. [Google Scholar]
- Joetzjer, E.; Maignan, F.; Chave, J.; Goll, D.; Poulter, B.; Barichivich, J.; Maréchaux, I.; Luyssaert, S.; Guimberteau, M.; Naudts, K.; et al. Effect of Tree Demography and Flexible Root Water Uptake for Modeling the Carbon and Water Cycles of Amazonia. Ecol. Model. 2022, 469, 109969. [Google Scholar] [CrossRef]
- Gilmanov, T.G.; Morgan, J.A.; Hanan, N.P.; Wylie, B.K.; Rajan, N.; Smith, D.P.; Howard, D.M. Productivity and CO 2 Exchange of Great Plains Ecoregions. I. Shortgrass Steppe: Flux Tower Estimates. Rangel. Ecol. Manag. 2017, 70, 700–717. [Google Scholar] [CrossRef]
- Wolf, S.; Eugster, W.; Potvin, C.; Turner, B.L.; Buchmann, N. Carbon Sequestration Potential of Tropical Pasture Compared with Afforestation in Panama. Glob. Chang. Biol. 2011, 17, 2763–2780. [Google Scholar] [CrossRef]
- Skinner, R.H. High Biomass Removal Limits Carbon Sequestration Potential of Mature Temperate Pastures. J. Environ. Qual. 2008, 37, 1319–1326. [Google Scholar] [CrossRef]
- Zhang, K.; Wang, Y.; Mamtimin, A.; Liu, Y. Carbon dioxide fluxes over grassland ecosystems in the Middle Tianshan region of China with eddy covariance method. Res. Sq. 2023. [Google Scholar] [CrossRef]
- Sun, J.; Zhou, T.C.; Liu, M.; Chen, Y.C.; Liu, G.H.; Xu, M.; Shi, P.L.; Peng, F.; Tsunekawa, A.; Liu, Y.; et al. Water and Heat Availability Are Drivers of the Aboveground Plant Carbon Accumulation Rate in Alpine Grasslands on the Tibetan Plateau. Glob. Ecol. Biogeogr. 2020, 29, 50–64. [Google Scholar] [CrossRef]
- Niu, Y.Y.; Li, Y.Q.; Wang, X.Y.; Gong, X.W.; Luo, Y.Q.; Tian, D.Y. Characteristics of Annual Variation in Net Carbon Dioxide Flux in a Sandy Grassland Ecosystem during Dry Years. Acta Prataculturae Sin. 2018, 27, 215–221. [Google Scholar] [CrossRef]
- Li, S.-G.; Asanuma, J.; Eugster, W.; Kotani, A.; Liu, J.-J.; Urano, T.; Oikawa, T.; Davaa, G.; Oyunbaatar, D.; Sugita, M. Net Ecosystem Carbon Dioxide Exchange over Grazed Steppe in Central Mongolia. Glob. Chang. Biol. 2005, 11, 1941–1955. [Google Scholar] [CrossRef]
- De la Motte, L.G.; Jérôme, E.; Mamadou, O.; Beckers, Y.; Bodson, B.; Heinesch, B.; Aubinet, M. Carbon Balance of an Intensively Grazed Permanent Grassland in Southern Belgium. Agric. For. Meteorol. 2016, 228–229, 370–383. [Google Scholar] [CrossRef]
- Hirata, R.; Miyata, A.; Mano, M.; Shimizu, M.; Arita, T.; Kouda, Y.; Matsuura, S.; Niimi, M.; Saigusa, T.; Mori, A.; et al. Carbon Dioxide Exchange at Four Intensively Managed Grassland Sites across Different Climate Zones of Japan and the Influence of Manure Application on Ecosystem Carbon and Greenhouse Gas Budgets. Agric. For. Meteorol. 2013, 177, 57–68. [Google Scholar] [CrossRef]
- Allard, V.; Soussana, J.F.; Falcimagne, R.; Berbigier, P.; Bonnefond, J.M.; Ceschia, E.; D’hour, P.; Hénault, C.; Laville, P.; Martin, C.; et al. The Role of Grazing Management for the Net Biome Productivity and Greenhouse Gas Budget (CO2, N2O and CH4) of Semi-Natural Grassland. Agric. Ecosyst. Environ. 2007, 121, 47–58. [Google Scholar] [CrossRef]
- Gilmanov, T.G.; Soussana, J.F.; Aires, L.; Allard, V.; Ammann, C.; Balzarolo, M.; Barcza, Z.; Bernhofer, C.; Campbell, C.L.; Cernusca, A.; et al. Partitioning European Grassland Net Ecosystem CO2 Exchange into Gross Primary Productivity and Ecosystem Respiration Using Light Response Function Analysis. Agric. Ecosyst. Environ. 2007, 121, 93–120. [Google Scholar] [CrossRef]
- Klumpp, K.; Bloor, J.M.G.; Ambus, P.; Soussana, J.-F. Effects of Clover Density on N2O Emissions and Plant-Soil N Transfers in a Fertilised Upland Pasture. Plant Soil 2011, 343, 97–107. [Google Scholar] [CrossRef]
- Wall, A.M.; Campbell, D.I.; Morcom, C.P.; Mudge, P.L.; Schipper, L.A. Quantifying Carbon Losses from Periodic Maize Silage Cropping of Permanent Temperate Pastures. Agric. Ecosyst. Environ. 2020, 301, 107048. [Google Scholar] [CrossRef]
- Rutledge, S.; Wall, A.M.; Mudge, P.L.; Troughton, B.; Campbell, D.I.; Pronger, J.; Joshi, C.; Schipper, L.A. The Carbon Balance of Temperate Grasslands Part II: The Impact of Pasture Renewal via Direct Drilling. Agric. Ecosyst. Environ. 2017, 239, 132–142. [Google Scholar] [CrossRef]
- Hunt, J.E.; Laubach, J.; Barthel, M.; Fraser, A.; Phillips, R.L. Carbon Budgets for an Irrigated Intensively Grazed Dairy Pasture and an Unirrigated Winter-Grazed Pasture. Biogeosciences 2016, 13, 2927–2944. [Google Scholar] [CrossRef]
- Campbell, D.I.; Wall, A.M.; Nieveen, J.P.; Schipper, L.A. Variations in CO2 Exchange for Dairy Farms with Year-Round Rotational Grazing on Drained Peatlands. Agric. Ecosyst. Environ. 2015, 202, 68–78. [Google Scholar] [CrossRef]
- Mudge, P.L.; Wallace, D.F.; Rutledge, S.; Campbell, D.I.; Schipper, L.A.; Hosking, C.L. Carbon Balance of an Intensively Grazed Temperate Pasture in Two Climatically: Contrasting Years. Agric. Ecosyst. Environ. 2011, 144, 271–280. [Google Scholar] [CrossRef]
- Nieveen, J.P.; Campell, D.I.; Schipper, L.A.; Blair, I.J. Carbon Exchange of Grazed Pasture on Drained Peat Soil. Glob. Chang. Biol. 2005, 11, 607–618. [Google Scholar] [CrossRef]
- Bezerra, B.G.; Santos e Silva, C.M.; Mendes, K.R.; Mutti, P.R.; Fernandes, L.S.; Marques, T.V.; Câmara e Silva, C.L.; Campos, S.; de Lima Vieira, M.M.; Urbano, S.A.; et al. CO2 Exchanges and Evapotranspiration of a Grazed Pasture under Tropical Climate Conditions. Agric. For. Meteorol. 2022, 323, 109088. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Measurement Parameter | Instrumentation | Model |
|---|---|---|
| H2O/CO2 EC-Flux, 10 Hz | Infrared Gas Analyzer | LI-7200, Closed Path (LI-COR Environmental GmbH, Bad Homburg, Germany) |
| Precipitation, total | Tipping bucket rain gauge | Texas Electronics TE525 (Texas Electronics, Dallas, TX, USA) |
| Photosynthetic Photon Flux Density (PPFD) | Quantum Sensor | LI-190R Quantum Sensor (LI-COR Environmental GmbH, Bad Homburg, Germany) |
| Soil, heat flux | Soil heat flux plate | REBS HFT (Radiation and Energy Balance Systems, Inc., Seattle, WA, USA) |
| Temperature, air | Temperature/humidity probe | Vaisala HMP 35C (Vaisala Oyj, Vantaa, Finland) |
| 3D Wind | 3D Sonic Anemometer | Gill (Gill Instruments Limited, Lymington, Hampshire, UK) |
| Amax | α | Re (gCm−2) | n | R2 | PPFD (KW/m2/Month) | |
|---|---|---|---|---|---|---|
| January | 0.1778 | 0.0004 | 0.0331 | 551 | 0.9351 | 5.80 |
| February | 0.2515 | 0.0003 | 0.0379 | 563 | 0.9446 | 6.44 |
| March | 0.6049 | 0.0008 | 0.0882 | 684 | 0.9245 | 9.49 |
| April | 0.8722 | 0.0010 | 0.1327 | 743 | 0.9318 | 13.24 |
| May | 0.8908 | 0.0014 | 0.1979 | 810 | 0.8980 | 12.85 |
| June | 2.3603 | 2.3603 | 5.3614 | 820 | 0.4730 | 16.46 |
| July | 0.7138 | 0.0011 | 0.2616 | 541 | 0.9030 | 8.97 |
| August | 0.9800 | 0.0017 | 0.3086 | 785 | 0.8954 | 13.26 |
| September | 0.3788 | 0.0014 | 0.1880 | 697 | 0.8155 | 12.01 |
| October | 0.3292 | 0.0015 | 0.1257 | 644 | 0.8176 | 9.23 |
| November | 0.2384 | 0.0011 | 0.0795 | 566 | 0.8389 | 6.96 |
| December | 0.1954 | 0.0008 | 0.0605 | 510 | 0.8650 | 4.40 |
| GPP (gCm−2yr−1) | RE (gCm−2yr−1) | NEE (gCm−2yr−1) | ||||||
|---|---|---|---|---|---|---|---|---|
| Year | GS | NGS | GS | NGS | GS | NGS | NEE | Status |
| 2015 | 1411.92 | 333.87 | 1261.98 | 239.62 | −149.94 | −94.25 | −244.19 | Sink |
| 2016 | 1008.99 | 330.33 | 1079.65 | 276.52 | 70.65 | −53.80 | 16.85 | Neutral |
| 2017 | 1651.25 | 340.42 | 1498.81 | 315.99 | −152.44 | −24.43 | −176.88 | Sink |
| 2018 | 1343.22 | 220.29 | 763.50 | 264.59 | −579.72 | 44.31 | −535.41 | Sink |
| 2019 | 1731.87 | 246.13 | 1672.11 | 277.50 | −59.75 | 31.37 | −28.38 | Sink |
| 2020 | 836.14 | 148.43 | 465.32 | 93.56 | −370.83 | −54.87 | −425.69 | Sink |
| Sub Total | 7983 ± 279 | 1619 ± 165 | 6741 ± 291 | 1468 ± 90 | −1242 ± 151 | −152 ± 91 | ||
| Cumulative | 9603 ± 479 | 8209 ± 410 | −1394 ± 146 | Sink | ||||
| Continent | Station | Cropping Management | Temp. (°C) | Precip. (mm) | NEE (g C yr−1) | Study Years | Location | Reference | |
|---|---|---|---|---|---|---|---|---|---|
| N. America | i. | Harold Benson Research Farm, Frankfort, KY | Managed Pasture | 13.54 | 1369.95 | −232.3 | 2015–2020 | 38.11° N, 84.88° W | Present Study |
| ii. | SGS ecoregion of the North American Great Plains, Colorado, USA | Heavy/moderate continuous grazing | −23/−59 | 2018 | [59] | ||||
| iii. | Sardinilla, N.E. Barro Colorado Island, Panama | 261 | 2008–2009 | 9°19′ N, 79°38′ W | [60] | ||||
| iv | Pennsylvania State Uni. Haller Research Farm | 9.7 | 1014 | 14/−5 | 2008 | 40.9° N; 77.8° W | [61] | ||
| Asia | i. | Ulastai station, Tien Shan Mountains, Xinjiang Uygur Autonomous Region. | Middle Tien Shan grassland ecosystem | 7.26 | 257.61 | −89.95 | 2018 | 43°28′ N, 87°12′ E | [62] |
| ii. | Yunnan-Guizhou Plateau, China | Grassland ecosystem | −116.06 | 2017–2018 | 27°46′ N, 107°28′ E | [63] | |||
| iii. | Horqin District, Inner Mongolia, China | Grassland ecosystem | −32.91 | 2015–2018 | 42°55′ N, 12°42′ E | [64] | |||
| iv. | Kherlenbayan-Ulaan, Hentiy province, central Mongolia | −41 | 2005 | [65] | |||||
| Europe | i. | Condroz region, Belgium | −141 | 2016 | [66] | ||||
| ii. | Grassland at Nakashibetsu, Shizunai, Nasushiobara, and Kobayashi., Japan | −245/−158 | 2013 | [67] | |||||
| iii. | Laqueuille, Puy de Döme, France | 1.32 | 2007 | [68] | |||||
| iv. | European grazed grassland | −32 | 2007 | [69] | |||||
| v. | Seminatural grassland site in Auvergne, ‘Laqueuille’, France | Extensively managed paddock | 7.07 | 997.5 | −225 | 2003–2008 | 45°38′ N, 2°44′ E | [70] | |
| vi. | “ | Intensively managed paddock | 7.07 | 997.5 | −209 | 2003–2008 | 45°38′ N, 2°44′ E | ‘’ | |
| Oceania | i. | Waikato region, New Zealand | Rotationally grazed pasture | 13.3 | 1249 | −114 | 2018 | 37°45.6′ S, 175°47.2′ E | [71] |
| ii. | “ | “ | 13.3 | 1249 | 11 | 2017 | 37°45.6′ S, 175°47.2′ E | ‘’ | |
| iii. | Troughton Farm, Waikato region, North Island, New Zealand | old pasture/renewed pasture | −187/−148 | 2017 | 37°45.6′ S, 175°47.2′ E | [72] | |||
| iv. | Canterbury, New Zealand | dryland/irrigated | −20/−408 | 2016 | 43.7542° S, 171.1637° E | [73] | |||
| v. | Hamilton Basin, Waikato region, North Island/Te Ika-a-Maui, New Zealand | Intense, year-round rotational grazing | 190 | 2015 | [74] | ||||
| vi. | Scott Farm, Waikato region, North Island, New Zealand | −195 | 2011 | [75] | |||||
| vii. | Rukuhia peatland, Waikato region, New Zealand | 4 | 2005 | [76] | |||||
| S. America | i. | Experimental station, GEFORP -EAJ-UFRN, Northeast coast of Brazil, Macaíba district, Brazil | Rotational grazing | 25.5 | 1100 | −215 ± 22 | 2015–2017 | 5◦53′34″ S. 35◦21′50″ W | [77] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Familusi, I.; Gebremedhin, M.; Ries, I.; Brown, J.; Gyawali, B. The Productivity and Carbon Exchange of an Intensively Managed Pasture in Central Kentucky. Atmosphere 2024, 15, 348. https://doi.org/10.3390/atmos15030348
Familusi I, Gebremedhin M, Ries I, Brown J, Gyawali B. The Productivity and Carbon Exchange of an Intensively Managed Pasture in Central Kentucky. Atmosphere. 2024; 15(3):348. https://doi.org/10.3390/atmos15030348
Chicago/Turabian StyleFamilusi, Ife, Maheteme Gebremedhin, Ian Ries, Jacob Brown, and Buddhi Gyawali. 2024. "The Productivity and Carbon Exchange of an Intensively Managed Pasture in Central Kentucky" Atmosphere 15, no. 3: 348. https://doi.org/10.3390/atmos15030348
APA StyleFamilusi, I., Gebremedhin, M., Ries, I., Brown, J., & Gyawali, B. (2024). The Productivity and Carbon Exchange of an Intensively Managed Pasture in Central Kentucky. Atmosphere, 15(3), 348. https://doi.org/10.3390/atmos15030348