
Landscape and Climate Changes in Southeastern Amazonia from Quaternary Records of Upland Lakes

 ,
,  , , , ,
, , , ,
Abstract
1. Introduction
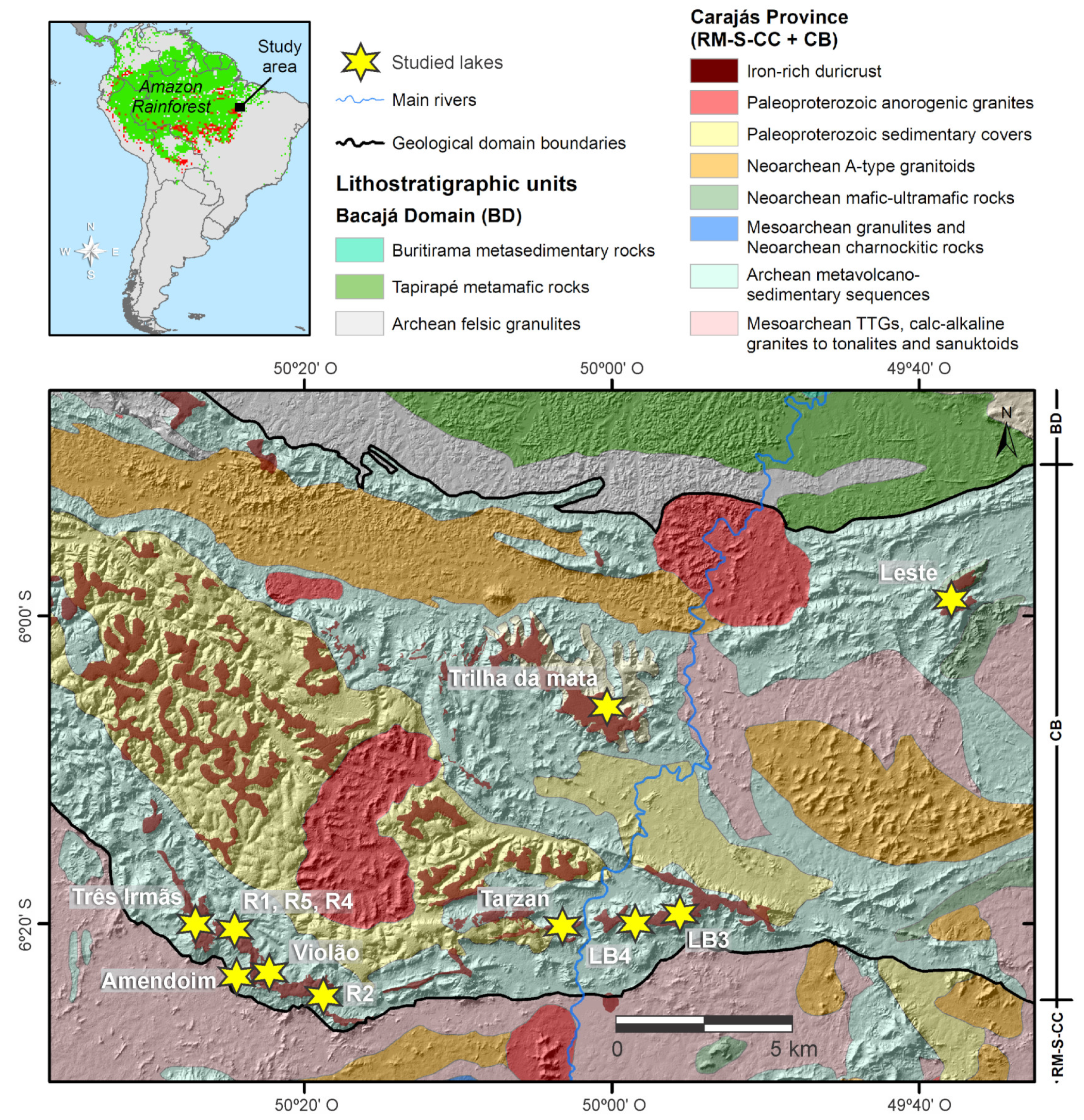
Geology, Physiography and Climate
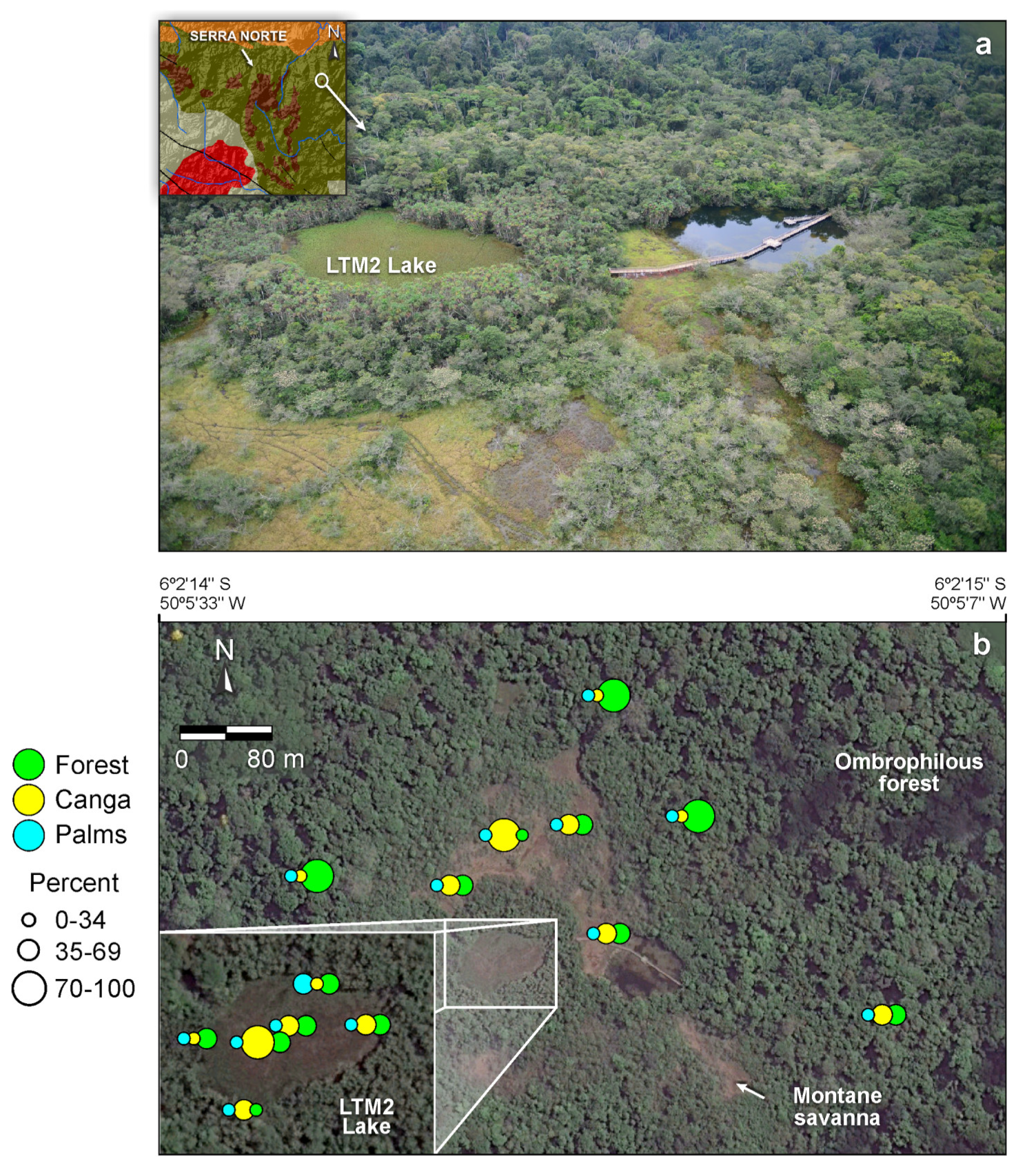
2. Lake Formation Processes
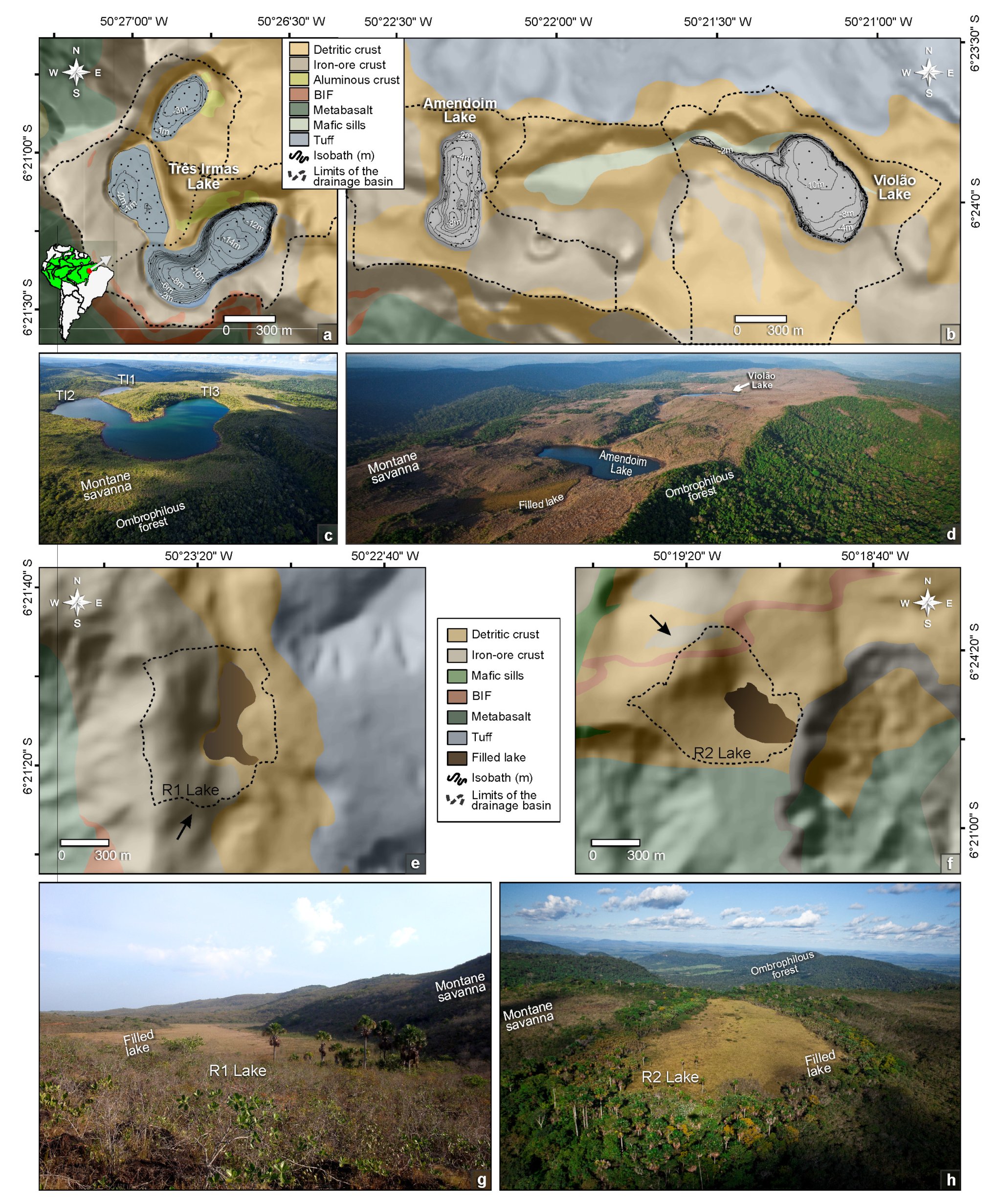
3. Surface Geology and Geobotany of the Catchment Basins
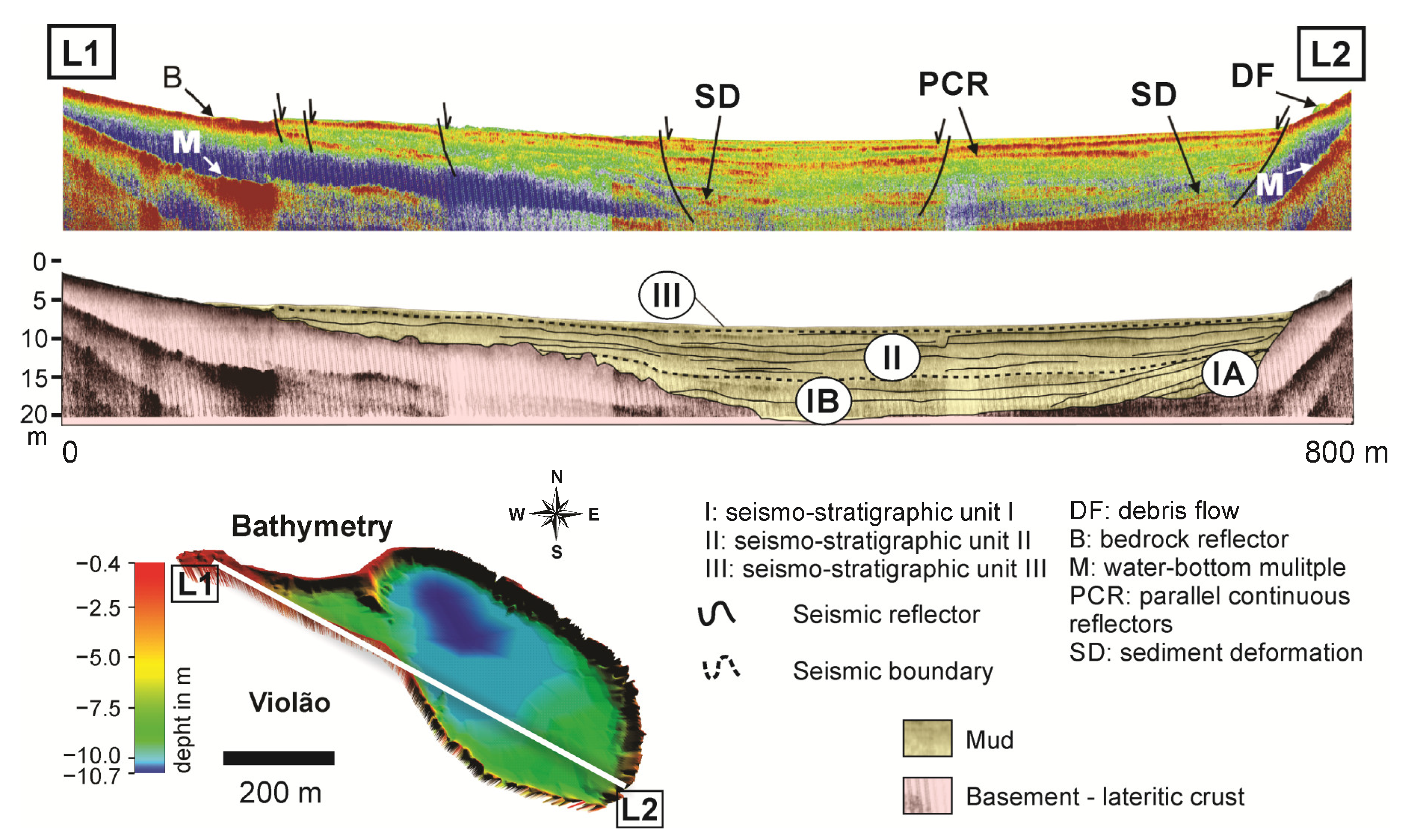
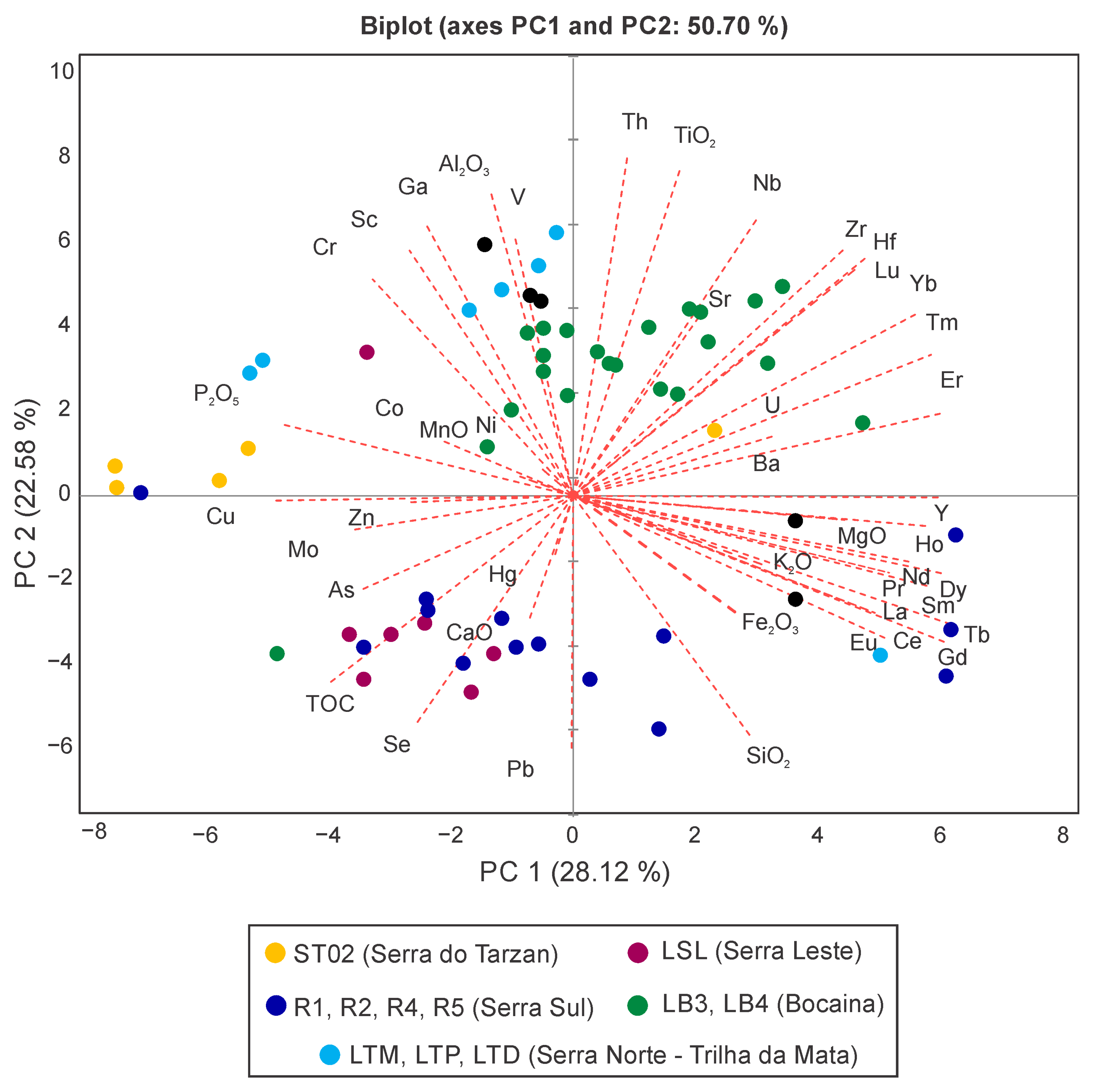
4. Modern Sedimentation Patterns: Basin Morphology and Source-to-Sink Relationship
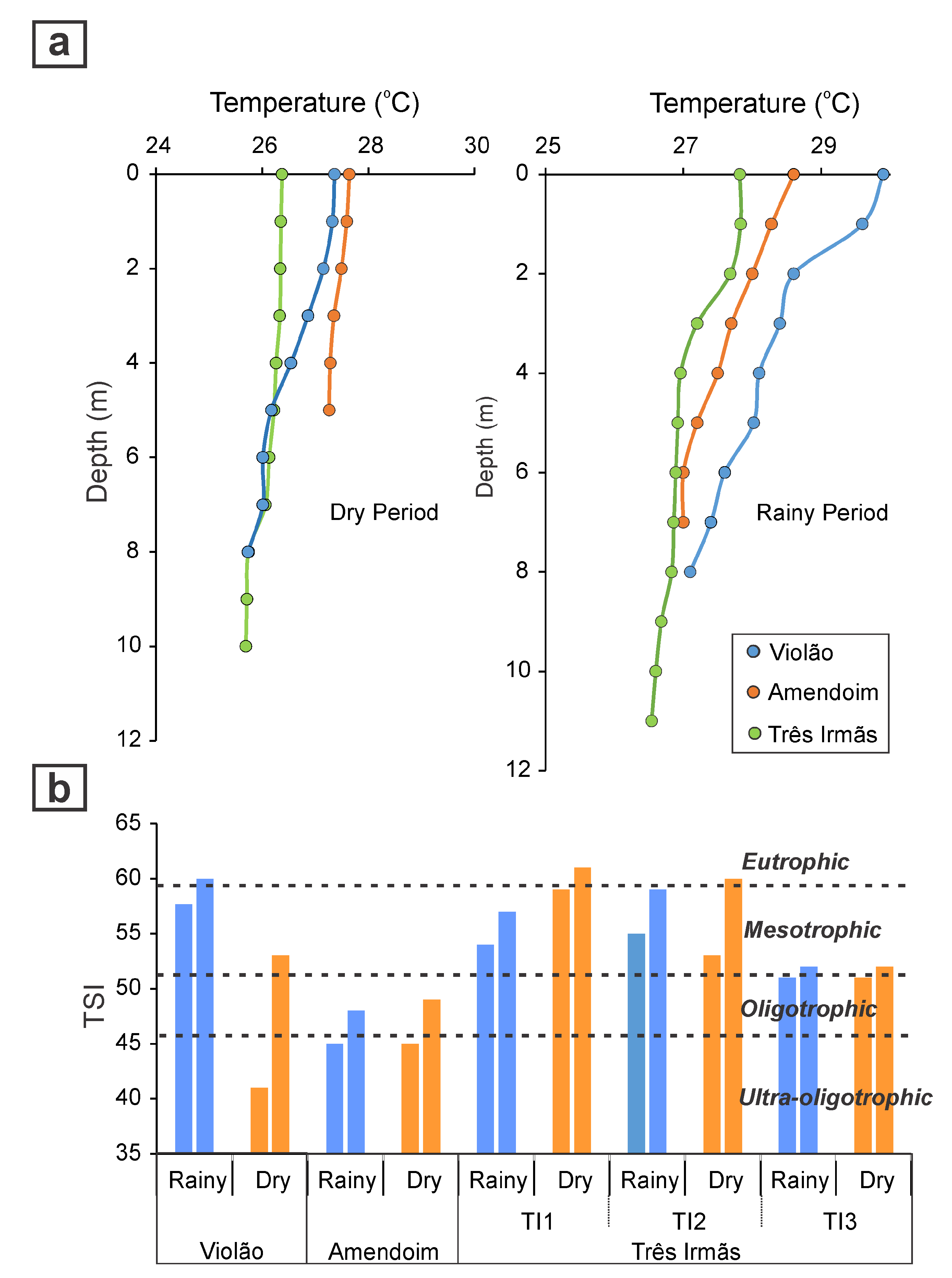
5. Environmental Influences on Limnology and Water Quality
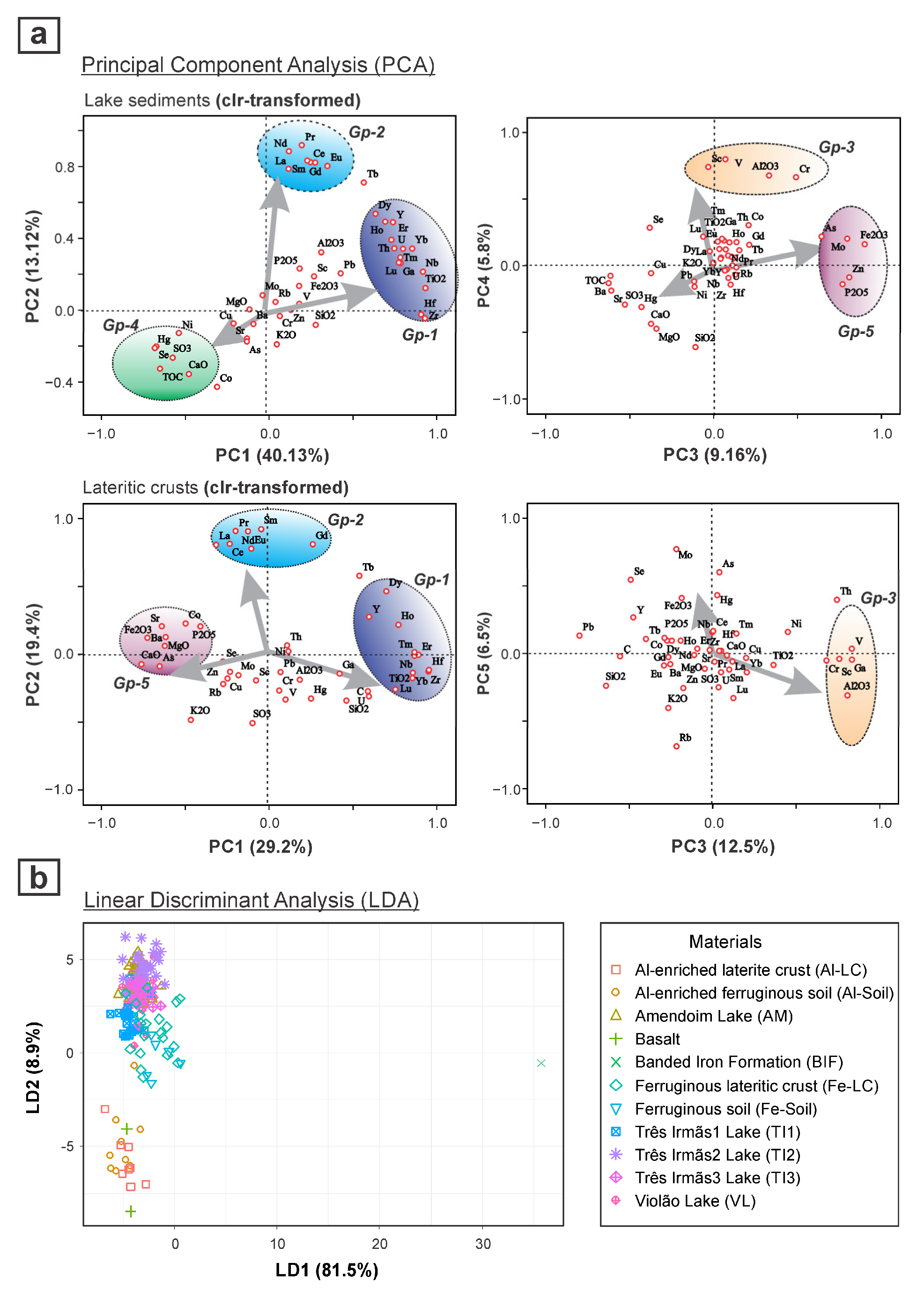
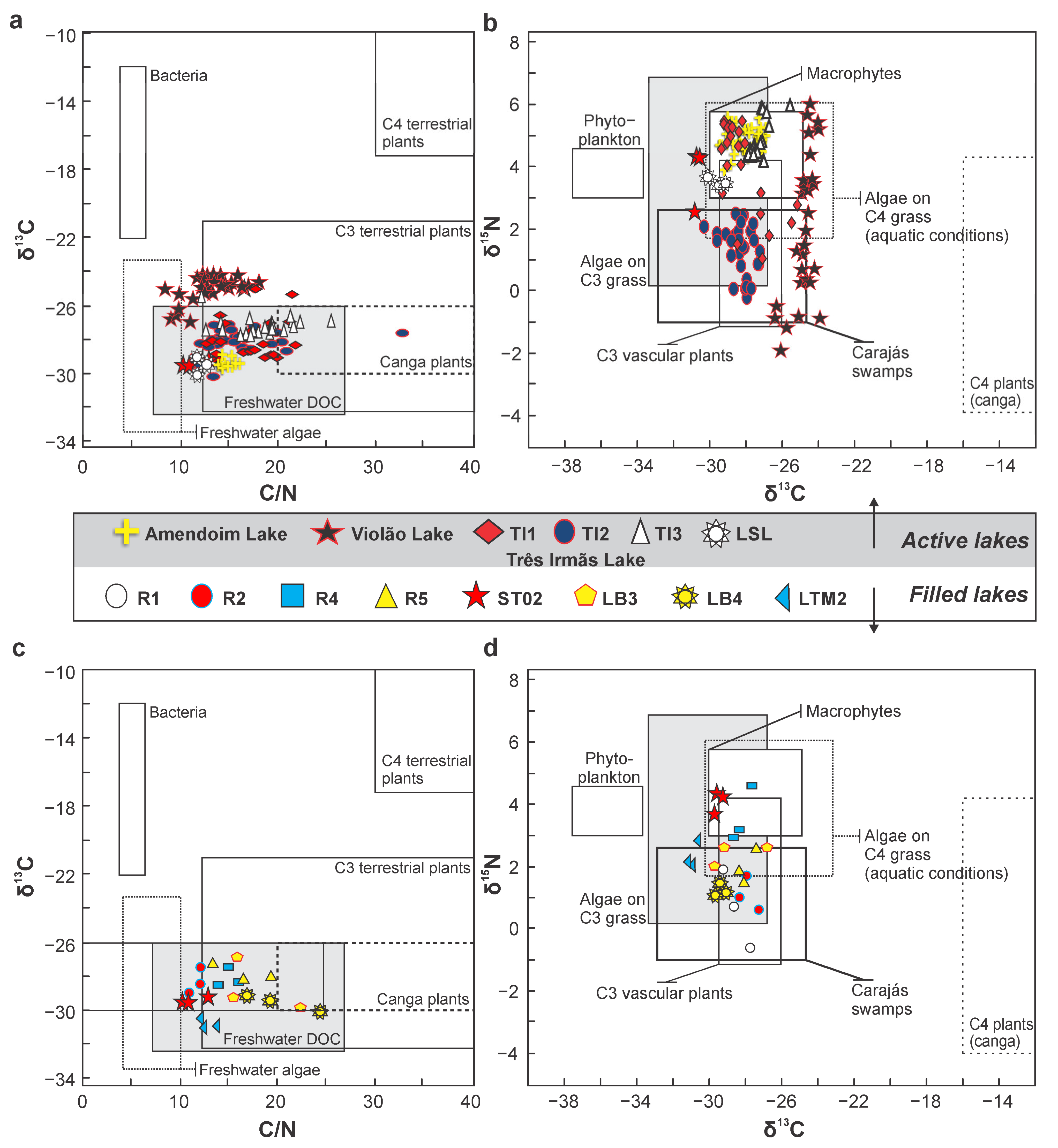
6. Role of Isotopes in Understanding the Sources of Sediment Organic Matter
7. Modern Pollen Rain Using UL Sediments
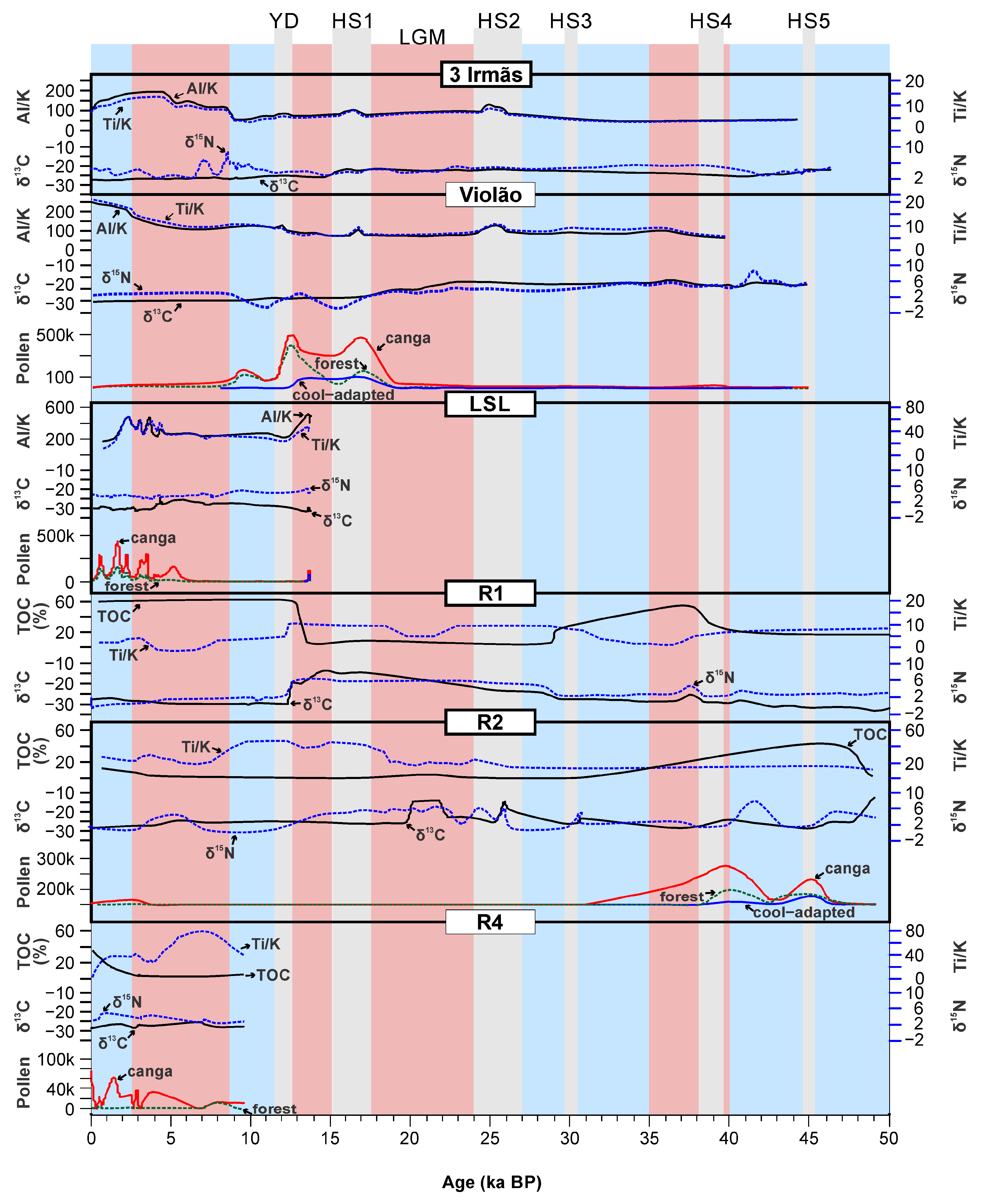
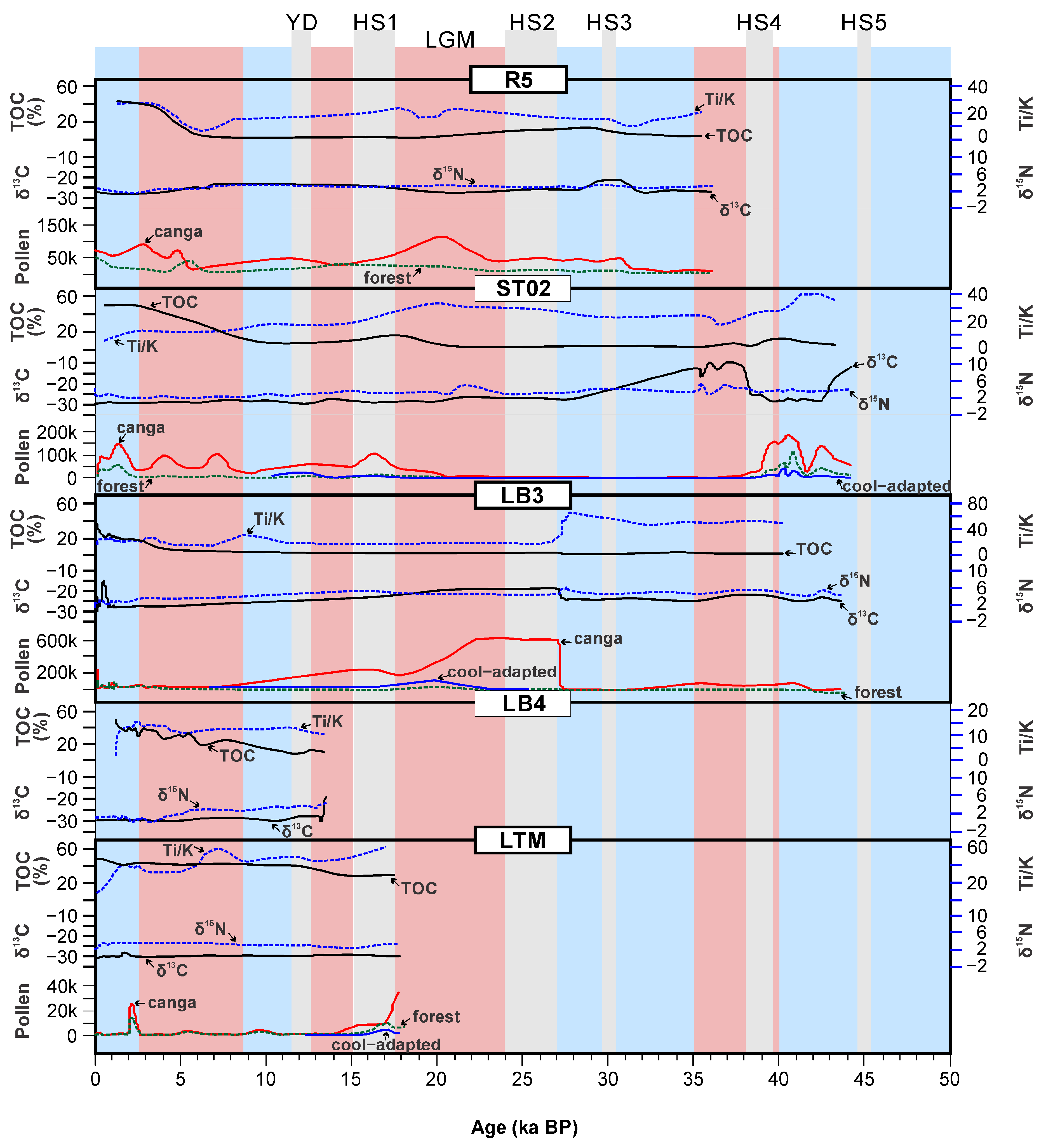
8. UL Sediments: Implications for Paleovegetation and Paleoenvironmental Changes
9. Current Approach and Future Perspectives of Paleoenvironmental Studies in Carajás
10. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Maurity, C.W.; Kotschoubey, B. Evolução recente da cobertura de alteraçao no platô N1-Serra dos Carajás-PA: Degradação, pseudocarstificaçao, espeleotemas. Bol. Mus. Para. Emilio Goeldi. Ser. Ciênc. Terra 1995, 7, 331–362. [Google Scholar]
- Costa, M.L.; Carmo, M.S.; Behling, H. Mineralogia e geoquímica de sedimentos lacustres com substrato laterítico na Amazônia Brasileira. Rev. Bras. Geociências 2005, 35, 165–176. [Google Scholar] [CrossRef]
- Souza-Filho, P.W.M.; Pinheiro, R.V.L.; Costa, F.R.; Guimarães, J.T.F.; Sahoo, P.K.; Silva, M.S.; Silva, C.G. The role of fault reactivation in the development of tropical montane lakes. Earth Surf. Process. Landforms 2020, 45, 3732–3746. [Google Scholar] [CrossRef]
- Reis, L.S.; Guimarães, J.T.F.; Souza-Filho, P.W.M.; Sahoo, P.K.; Figueiredo, M.M.J.C.; Souza, E.B.; Giannini, T.C. Environmental and vegetation changes in southeastern Amazonia during the late Pleistocene and Holocene. Quat. Int. 2017, 449, 83–105. [Google Scholar] [CrossRef]
- Sahoo, P.K.; Souza-Filho, P.W.; Guimarães, J.T.; Da Silva, M.S.; Costa, F.R.; De Oliveira Manes, C.L.; Oti, D.; Júnior, R.O.; Dall’Agnol, R. Use of multi-proxy approaches to determine the origin and depositional processes in modern lacustrine sediments: Carajás Plateau, Southeastern Amazon, Brazil. Appl. Geochem. 2015, 52, 130–146. [Google Scholar] [CrossRef]
- Sahoo, P.K.; Guimaraes, J.T.; Souza-Filho, P.W.; da Silva, M.S.; Maurity, C.W.; Powell, M.A.; Rodrigues, T.M.; Da Silva, D.F.; Mardegan, S.F.; Neto, A.E.; et al. Geochemistry of upland lacustrine sediments from Serra dos Carajás, Southeastern Amazon, Brazil: Implication for catchment weathering, provenance, and sedimentary processes. J. South Am. Earth Sci. 2016, 72, 178–190. [Google Scholar] [CrossRef]
- Sahoo, P.K.; Guimarães, J.T.; Souza-Filho, P.W.M.; Da Silva, M.S.; Júnior, R.O.S.; Pessim, G.; De Moraes, B.C.; Pessoa, P.F.; Rodrigues, T.M.; Da Costa, M.F.; et al. Influence of seasonal variation on the hydro-biogeochemical characteristics of two upland lakes in the Southeastern Amazon, Brazil. An. Acad. Bras. Ciências 2016, 88, 2211–2227. [Google Scholar] [CrossRef]
- Nunes, J.A.; Schaefer, C.E.G.R.; Ferreira Júnior, W.G.; Neri, A.V.; Correa, G.R.; Enright, N.J. Soil-vegetation relationships on a banded ironstone ‘island’, Carajás Plateau, Brazilian Eastern Amazonia. An. Acad. Bras. Ciênc. 2015, 87, 2097–2110. [Google Scholar] [CrossRef] [PubMed]
- Sahoo, P.K.; Guimarães, J.T.; Souza-Filho, P.W.; da Silva, M.S.; Júnior, W.N.; Powell, M.A.; Reis, L.S.; Pessenda, L.C.; Rodrigues, T.M.; da Silva, D.F.; et al. Geochemical characterization of the largest upland lake of the Brazilian Amazonia: Impact of provenance processes. J. South Am. Earth Sci. 2017, 80, 541–558. [Google Scholar] [CrossRef]
- Sahoo, P.K.; Guimarães, J.T.; Souza-filho, P.W.; Bozelli, R.L.; de Araujo, L.R.; de Souza Menezes, R.; Lopes, P.M.; da Silva, M.S.; Rodrigues, T.M.; da Costa, M.F.; et al. Limnological characteristics planktonic diversity of five tropical upland lakes from Brazilian Amazon. Ann. Limnol. Int. J. Limnol. 2017, 53, 467–483. [Google Scholar] [CrossRef]
- Colinvaux, P.A.; Irion, G.; Räsänen, M.E.; Bush, M.B.; Nunes de Mello, J.A.S. A paradigma to be discarded: Geological and paleoecological data falsify the HAFFER & PRANCE refuge hypothesis of Amazonian speciation. Amazoniana 2001, 16, 609–646. [Google Scholar]
- Bush, M.B.; Oliveira, P.E.; Colinvaux, P.A.; Miller, M.C.; Moreno, J.E. Amazonian palaeoecological histories: One hill three watersheds. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2004, 214, 359–393. [Google Scholar] [CrossRef]
- Sifeddine, A.; Martin, L.; Turcq, B.; Volkmer-Ribeiro, C.; Soubiès, F.; Cordeiro, R.C.; Suguio, K. Variations of the Amazonian rainforest environment: A sedimentological record covering 30,000 years. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2001, 168, 221–235. [Google Scholar] [CrossRef]
- Cordeiro, R.; Turcq, B.; Sifeddine, A.; Lacerda, L.; Filho, E.S.; Gueiros, B.; Potty, Y.; Santelli, R.; Pádua, E.; Patchinelam, S. Biogeochemical indicators of environmental changes from 50Ka to 10Ka in a humid region of the Brazilian Amazon. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2011, 299, 426–436. [Google Scholar] [CrossRef]
- D’Apolito, C.; Absy, M.L.; Latrubesse, E.M. The Hill of Six Lakes revisited: New data and re-evaluation of a key Pleistocene Amazon site. Quat. Sci. Rev. 2013, 76, 140–155. [Google Scholar] [CrossRef]
- Guimarães, J.T.F.; Sahoo, P.K.; Souza-Filho, P.W.M.; Maurity, C.W.; Silva, J.R.O.; Costa, F.R.; Dall’Agnol, R. Late Quaternary environmental climate changes registered in lacustrine sediments of the Serra Sul de Carajás southeast Amazonia. J. Quat. Sci. 2016, 31, 61–74. [Google Scholar] [CrossRef]
- Guimarães, J.T.F.; Sahoo, P.K.; De Figueiredo, M.M.J.C.; Lopes, K.S.; Gastauer, M.; Ramos, S.J.; Caldeira, C.F.; Souza-Filho, P.W.M.; Reis, L.S.; Da Silva, M.S.; et al. Lake sedimentary processes and vegetation changes over the last 45k cal a BP in the uplands of south-eastern Amazonia. J. Quat. Sci. 2021, 36, 255–272. [Google Scholar] [CrossRef]
- Reis, L.S.; Bouloubassi, I.; Mendez-Millan, M.; Guimarães, J.T.F.; Romeiro, L.D.A.; Sahoo, P.K.; Pessenda, L.C.R. Hydroclimate and vegetation changes in southeastern Amazonia over the past ∼25,000 years. Quat. Sci. Rev. 2022, 284, 107466. [Google Scholar] [CrossRef]
- Lopes, P.M.; Caliman, A.; Carneiro, L.S.; Bini, L.M.; Esteves, F.A.; Farjalla, V.; Bozelli, R.L. Concordance among assemblages of upland Amazonian lakes and the structuring role of spatial and environmental factors. Ecol. Indic. 2011, 11, 1171–1176. [Google Scholar] [CrossRef]
- Mormul, R.P.; Esteves, F.D.A.; Farjalla, V.F.; Bozelli, R.L. Space and seasonality effects on the aquatic macrophyte community of temporary Neotropical upland lakes. Aquat. Bot. 2015, 126, 54–59. [Google Scholar] [CrossRef]
- Silva, M.S.; Guimarães, J.T.F.; Souza Filho, P.W.M.; Nascimento Júnior, W.R.; Sahoo, P.K.; Costa, F.R.; Silva Júnior, R.O.; Rodrigues, T.M.; Costa, M.F. Morphology and morphometry of upland lakes over lateritic crust, Serra dos Carajás, southeastern Amazon region. An. Acad. Bras. Ciências 2018, 90, 1309–1325. [Google Scholar] [CrossRef]
- Schnurrenberger, D.; Russell, J.; Kelts, K. Classification of lacustrine sediments based on sedimentary components. J. Paleolimnol. 2003, 29, 141–154. [Google Scholar] [CrossRef]
- Walker, R.G. Facies, facies models and modern stratigraphic concepts. In Facies Models—Response to Sea Level Change, 2nd ed.; Walker, R.G., James, N.P., Eds.; Geological Association of Canada: Toronto, ON, Canada, 1992; pp. 1–14. [Google Scholar]
- Moraes, A.M.; Sahoo, P.K.; Guimarães, J.T.F.; Leite, A.S.; Salomão, G.N.; Souza-Filho, P.W.M.; Júniora, W.N.; Dall’Agnol, R. Multivariate statistics and geochemical approaches for understanding source-sink relationship—A case study from close-basin lakes in Southeast Amazon. J. S. Am. Earth Sci. 2020, 99, 102497. [Google Scholar] [CrossRef]
- Sahoo, P.K.; Guimarães, J.T.F.; Souza-Filho, P.W.M.; Powell, M.A.; da Silva, M.S.; Moraes, A.M.; Alves, R.; Leite, A.S.; Júnior, W.N.; Rodrigues, T.M.; et al. Statistical analysis of lake sediment geochemical data for understanding surface geological factors and processes: An example from Amazonian upland lakes, Brazil. Catena 2018, 175, 47–62. [Google Scholar] [CrossRef]
- Guimarães, J.T.G.; Sahoo, P.; Souza-Filho, P.W.M.; Silva, M.; Rodrigues, T.; Silva, E.; Reis, L.; Figueiredo, M.; Lopes, K.; Batista Junior, W.; et al. Upland Lakes of the Carajás Region: Origin and Development through Time. Preprints 2019, 2019050214. [Google Scholar] [CrossRef]
- Guimarães, J.T.; Sahoo, P.K.; Souza-Filho, P.W.; DE Figueiredo, M.M.C.; Reis, L.S.; Da Silva, M.S.; Rodrigues, T.M. Holocene history of a lake filling and vegetation dynamics of the Serra Sul dos Carajás, southeast Amazonia. An. Acad. Bras. Ciênc. 2019, 91, e20160916. [Google Scholar] [CrossRef]
- Da Silva, E.F.; Lopes, K.D.S.; Alves, R.; Carreira, L.M.M.; da Silva, D.F.; Romeiro, L.D.A.; Júnior, W.F.B.; Rodrigues, T.M.; Secco, R.D.S.; Guimarães, J.T.F. Hydroclimate influences on modern pollen rain of upland southeastern Amazonia. Holocene 2020, 30, 721–732. [Google Scholar] [CrossRef]
- Guimarães, J.T.F.; Souza-Filho, P.W.M.; Alves, R.; Souza, E.B.; Costa, F.R.; Reis, L.S.; Sahoo, P.K.; Manes, C.L.O.; Silva Junior, R.O.; Oti, D.; et al. Source and distribution of pollen and spores in surface sediments of a plateau lake in south-eastern Amazonia. Quat. Int. 2014, 352, 181–196. [Google Scholar] [CrossRef]
- Guimarães, J.T.F.; Rodrigues, T.M.R.; Reis, L.S.; de Figueiredo, M.M.J.C.; da Silva, D.F.; Alves, R.; Giannini, T.C.; Carreira, L.M.M.; Dias, A.C.R.; Silva, E.F.; et al. Modern pollen rain as a background for palaeoenvironmental studies in the Serra dos Carajás, southeastern Amazonia. Holocene 2017, 27, 1055–1066. [Google Scholar] [CrossRef]
- Faegri, K.; Iversen, J. Textbook of Pollen Analyses; Wiley: Chichester, UK, 1989. [Google Scholar]
- Blaauw, M.; Christen, J.A. Flexible palaeoclimate age-depth models using an autoregressive gamma process. Bayesian Anal. 2011, 6, 457–474. [Google Scholar] [CrossRef]
- Reimer, P.J.; Austin, W.E.N.; Bard, E.; Bayliss, A.; Blackwell, P.G.; Ramsey, C.B.; Butzin, M.; Cheng, H.; Edwards, R.L.; Friedrich, M.; et al. The IntCal20 Northern Hemisphere Radiocarbon Age Calibration Curve (0–55 cal kBP). Radiocarbon 2020, 62, 725–757. [Google Scholar] [CrossRef]
- EPA. Method 9060A—Total Organic Carbon. 5P. Revision 1; United States Environmental Protection Agency: Washington, DC, USA, 2004.
- SMEWW. 1060–Collection and Preservation of Samples; Standard Methods for the Examination of Water and Wastewater: Washington, DC, USA, 2005. [Google Scholar]
- Feio, G.; Dall’Agnol, R.; Dantas, E.; Macambira, M.; Santos, J.; Althoff, F.; Soares, J. Archean granitoid magmatism in the Canaã dos Carajás area: Implications for crustal evolution of the Carajás province, Amazonian craton, Brazil. Precambrian Res. 2013, 227, 157–185. [Google Scholar] [CrossRef]
- Moreto, C.P.N.; Monteiro, L.V.S.; Xavier, R.P.; Creaser, R.A.; DuFrane, S.A.; Tassinari, C.C.G.; Sato, K.; Kemp, A.I.S.; Amaral, W.S. Neoarchean and Paleoproterozoic Iron Oxide-Copper-Gold Events at the Sossego Deposit, Carajas Province, Brazil: Re-Os and U-Pb Geochronological Evidence. Econ. Geol. 2015, 110, 809–835. [Google Scholar] [CrossRef]
- Martins, P.L.G.; Toledo, C.L.B.; Silva, A.M.S.; Chemale, F., Jr.; Santos, J.O.S.; Assis, L.M. Neoarchean magmatism in the southeastern Amazonian Craton, Brazil: Petrography, geochemistry and tectonic significance of basalts from the Carajás Basin. Precambrian Res. 2017, 302, 340–357. [Google Scholar] [CrossRef]
- Dall’Agnol, R.; da Cunha, I.R.V.; Guimarães, F.V.; de Oliveira, D.C.; Teixeira, M.F.B.; Feio, G.R.L.; Lamarão, C.N. Mineralogy, geochemistry, and petrology of Neoarchean ferroan to magnesian granites of Carajás Province, Amazonian Craton: The origin of hydrated granites associated with charnockites. Lithos 2017, 277, 3–32. [Google Scholar] [CrossRef]
- Mansur, E.T.; Filho, C.F.F.; Oliveira, D.P. The Luanga deposit, Carajás Mineral Province, Brazil: Different styles of PGE mineralization hosted in a medium-size layered intrusion. Ore Geol. Rev. 2020, 118, 103340. [Google Scholar] [CrossRef]
- Nogueira, A.C.R.; Trunckenbrodt, W.; Pinheiro, R.L.V. Formação Águas Claras, Pré-cambriano da Serra dos Carajás: Redescrição e redefinição litoestratigráfica. Bol. Mus. Para. Emílio Goeldi 1995, 7, 177–197. [Google Scholar]
- Dall’Agnol, R.; Teixeira, N.P.; Rämö, O.T.; Moura, C.A.; Macambira, M.J.; de Oliveira, D.C. Petrogenesis of the Paleoproterozoic rapakivi A-type granites of the Archean Carajás metallogenic province, Brazil. Lithos 2005, 80, 101–129. [Google Scholar] [CrossRef]
- Vasquez, M.L.; Sousa, C.S.; Carvalho, K.M.A. Mapa Geológico e de Recursos Minerais do Estado do Pará, escala 1:1.000.000. Programa Geologia do Brasil (PGB), Integração, Atualização e Difusão de Dados da Geologia do Brasil, Mapas Geológicos Estaduais; Serviço Geológico do Brasil (CPRM): Sao Paolo, Brazil, 2008.
- Souza-Filho, P.W.M.; Guimarães, J.T.F.; Silva, M.S.; Costa, F.R.; Sahoo, P.K.; Maurity, C.W.; Dall’Agnol, R. Basin morphology, sedimentology and seismic stratigraphy of an upland lake from Serra dos Carajás, southeastern Amazon, Brazil. Bol. Mus. Para. Emílio Goeldi. Cienc. Nat. 2016, 11, 71–83. [Google Scholar] [CrossRef]
- Lopes, M.N.G.; Souza, E.B.; Ferreira, D.B.D.S. Climatologia regional da precipitação no estado do Pará. Rev. Bras. Climatol. 2013, 12, 84–102. [Google Scholar]
- Da Silva Júnior, R.O.; De Souza, E.B.; Tavares, A.L.; Mota, J.A.; Ferreira, D.B.; Souza-Filho, P.W.; Da Rocha, E.J. Three decades of reference evapotranspiration estimates for a tropical watershed in the eastern Amazon. An. Acad. Bras. Ciências 2017, 89, 1985–2002. [Google Scholar] [CrossRef]
- Tavares, A.L.; Carmo, A.M.C.; Silva Júnior, R.O.; Souza-Filho, P.W.M.; Silva, M.S.; Ferreira, D.B.S.; Nascimento Júnior, W.R.; Dall’Agnol, R. Climate indicators for a watershed in the eastern amazon. Rev. Bras. Climatol. 2018, 23, 389–410. [Google Scholar] [CrossRef]
- Golder. Anexo IV—Geologia. Estudo de Impacto Ambiental, EIA Projeto Ferro Carajás S11D. 2010. Available online: http://licenciamento.ibama.gov.br/Mineracao/Projeto%20Ferro%20Carajas%20S11D/EIA_RIMA/ (accessed on 13 April 2014).
- Skirycz, A.; Castilho, A.; Chaparro, C.; Carvalho, N.; Tzotzos, G.; Siqueira, J.O. Canga biodiversity, a matter of mining. Front. Plant Sci. 2014, 5, 653. [Google Scholar] [CrossRef]
- Schaefer, C.G.R.E.; Lima Neto, E.; Corrêa, G.R.; Simas, F.N.B.; Campos, J.F.; De Mendonça, B.A.F. Geoenviroments, soils and carbon stocks at Serra Sul of Carajás, Para State, Brazil. Bol. Mus. Para. Emílio Goeldi Cienc. Nat. 2016, 11, 85–101. [Google Scholar] [CrossRef]
- Mitre, S.K.; Mardegan, S.F.; Caldeira, C.F.; Ramos, S.J.; Neto, A.E.F.; Siqueira, J.O.; Gastauer, M. Nutrient and water dynamics of Amazonian canga vegetation differ among physiognomies and from those of other neotropical ecosystems. Plant Ecol. 2018, 219, 1341–1353. [Google Scholar] [CrossRef]
- Caldeira, C.F.; Abranches, C.B.; Gastauer, M.; Ramos, S.J.; Guimarães, J.T.F.; Pereira, J.B.S.; Siqueira, J.O. Sporeling regeneration and ex situ growth of Isoëtes cangae (Isoetaceae): Initial steps towards the conservation of a rare Amazonian quillwort. Aquat. Bot. 2018, 152, 51–58. [Google Scholar] [CrossRef]
- Viana, P.L.; Mota, N.F.O.; Gil, A.S.B.; Salino, A.; Zappi, D.C.; Harley, R.M.; Ilkiu-Borges, A.L.; Secco, R.; Almeida, T.E.; Watanabe, M.T.C.; et al. Flora of the cangas of the Serra dos Carajás, Pará, Brazil: History, study area and methodology. Rodriguesia 2016, 67, 1107–1124. [Google Scholar] [CrossRef]
- Wetzel, R.G. (Ed.) Oxygen. In Limnology; Academic Press: London, UK, 2001; pp. 151–168. [Google Scholar]
- Deines, P. The isotopic composition of reduced organic carbon. In Handbook of Environmental Isotope Geochemistry e the Terrestrial Environment; Fritz, P., Fontes, J.C., Eds.; Elsevier: Amsterdam, The Netherlands, 1980; pp. 329–406. [Google Scholar]
- Boutton, T.W. Stable carbon isotope ratios of natural materials: II. Atmospheric, terrestrial, marine, and freshwater environments. In Carbon Isotope Techniques; Coleman, D.C., Fry, B., Eds.; Academic Press Inc.: New York, NY, USA, 1991; pp. 173–185. [Google Scholar]
- Hamilton, S.K.; Lewis, W.M., Jr. Stable carbon and nitrogen isotopes in algae and detritus from the Orinoco River floodplain, Venezuela. Geochim. Cosmochim. Acta 1992, 56, 4237–4246. [Google Scholar] [CrossRef]
- Thornton, S.; McManus, J. Application of Organic Carbon and Nitrogen Stable Isotope and C/N Ratios as Source Indicators of Organic Matter Provenance in Estuarine Systems: Evidence from the Tay Estuary, Scotland. Estuar. Coast. Shelf Sci. 1994, 38, 219–233. [Google Scholar] [CrossRef]
- Meyers, P.A. Organic geochemical proxies of paleoceanographic, paleolimnologic, and paleoclimatic processes. Org. Geochem. 1997, 27, 213–250. [Google Scholar] [CrossRef]
- Brenner, M.; Whitmore, T.J.; Curtis, J.H.; Hodell, D.; Schelske, C.L. Stable isotope (δ13C and δ15N) signatures of sedimented organic matter as indicators of historic lake trophic state. J. Paleolimnol. 1999, 22, 205–221. [Google Scholar] [CrossRef]
- Troxler, T.G.; Richards, J.H. δ13C, δ15N, carbon, nitrogen and phosphorus as indicators of plant ecophysiology and organic matter pathways in Everglads deep slough, Florida, USA. Aquat. Bot. 2009, 91, 157–165. [Google Scholar] [CrossRef]
- Smith, C.B.; Cohen, M.C.L.; Pessenda, L.; França, M.; Guimarães, J. Holocenic proxies of sedimentary organic matter and the evolution of Lake Arari-Amazon Region. CATENA 2011, 90, 26–38. [Google Scholar] [CrossRef]
- Guimarães, J.T.F.; Sahoo, P.; Reis, L.S. Modern pollen rain raises doubts about the intensity and extension of the Last Glacial Cycle in Carajás: A reply to D’Apolito et al. Holocene 2017, 28, 332–335. [Google Scholar] [CrossRef]
- Pereira, J.B.D.S.; Salino, A.; Arruda, A.; Stützel, T. Two New Species of Isoetes (Isoetaceae) from northern Brazil. Phytotaxa 2016, 272, 141–148. [Google Scholar] [CrossRef]
- De Oliveira, S.M.B.; Saia, S.E.M.G.; Pessenda, L.C.R.; Favaro, D.I.T. Lacustrine sediments provide geochemical evidence of environmental change during the last millennium in southeastern Brazil. Geochemistry 2009, 69, 395–405. [Google Scholar] [CrossRef]
- Hodell, D.A.; Schelske, C.L. Production, sedimentation, and isotopic composition of organic matter in Lake Ontario. Limnol. Oceanogr. 1998, 43, 200–214. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, X.; Chiessi, C.M.; Mulitza, S.; Zhang, X.; Lohmann, G.; Prange, M.; Behling, H.; Zabel, M.; Govin, A.; et al. Equatorial Pacific forcing of western Amazonian precipitation during Heinrich Stadial 1. Sci. Rep. 2016, 6, 35866. [Google Scholar] [CrossRef] [PubMed]
- Fontes, D.; Cordeiro, R.; Martins, G.; Behling, H.; Turcq, B.; Sifeddine, A.; Seoane, J.; Moreira, L.; Rodrigues, R. Paleoenvironmental dynamics in South Amazonia, Brazil, during the last 35,000 years inferred from pollen and geochemical records of Lago do Saci. Quat. Sci. Rev. 2017, 173, 161–180. [Google Scholar] [CrossRef]
- Kimberley, M.M. Exhalative origins of iron formations. Ore Geol. Rev. 1989, 5, 13–145. [Google Scholar] [CrossRef]
- Lemos, V.P.; Costa, M.L.C.; Lemos, R.L.; de Faria, M.S.G. Vivianite and siderite in lateritic iron crust: An example of bioreduction. Quim. Nova 2007, 30, 36–40. [Google Scholar] [CrossRef]
- Maslin, M.A.; Ettwein, V.J.; Boot, C.S.; Bendle, J.; Pancost, R.D. Amazon Fan biomarker evidence against the Pleistocene rainforest refuge hypothesis? J. Quat. Sci. 2012, 27, 451–460. [Google Scholar] [CrossRef]
- Häggi, C.; Chiessi, C.M.; Merkel, U.; Mulitza, S.; Prange, M.; Schulz, M.; Schefuß, E. Response of the Amazon rainforest to late Pleistocene climate variability. Earth Planet. Sci. Lett. 2017, 479, 50–59. [Google Scholar] [CrossRef]
- Wang, X.; Edwards, R.L.; Auler, A.S.; Cheng, H.; Kong, X.; Wang, Y.; Cruz, F.W.; Dorale, J.A.; Chiang, H.-W. Hydroclimate changes across the Amazon lowlands over the past 45,000 years. Nature 2017, 541, 204–207. [Google Scholar] [CrossRef] [PubMed]
- Absy, M.L.; Cleef, A.M.; Fournier, M.; Martin, L.; Servant, M.; Sifeddine, A.; Silva da Soubiès, F.; Suguio, K.; Turcq, B.J.; Van der Hammen, T. Mise en évidence de quatre phases d’ouverture de la forêt dense dans le Sud-Est de l’Amazonie au cours des 60,000 dernières années: Première comparaison avec d’autres régions tropicales. Comptes Rendus L’académie Sci. 1991, 312, 673–678. [Google Scholar]
- Cordeiro, R.C.; Turcq, B.; Suguio, K.; Oliveira da Silva, A.; Sifeddine, A.; Volkmer, C. Holocene fires in East Amazonia (Carajás), new evidences, chronology and relation with palaeoclimate. Glob. Planet. Change 2008, 61, 49–62. [Google Scholar] [CrossRef]
- Hermanowski, B.; Da Costa, M.L.; Behling, H. Possible linkages of palaeofires in southeast Amazonia to a changing climate since the Last Glacial Maximum. Veg. Hist. Archaeobot. 2014, 24, 279–292. [Google Scholar] [CrossRef]
- Smith, R.J.; Mayle, F.E. Impact of mid- to late Holocene precipitation changes on vegetation across lowland tropical South America: A paleo-data synthesis. Quat. Res. 2017, 89, 134–155. [Google Scholar] [CrossRef]
- Cross, S.L.; Baker, P.A.; Seltzer, G.O.; Fritz, S.C.; Dunbar, R.B. A new estimate of the Holocene lowstand level of Lake Titicaca, central Andes, and implications for tropical palaeohydrology. Holocene 2000, 10, 21–32. [Google Scholar] [CrossRef]
- Seltzer, G.; Rodbell, D.; Burns, S. Isotopic evidence for late Quaternary climatic change in tropical South America. Geology 2000, 28, 35–38. [Google Scholar] [CrossRef]
- Mayle, F.E.; Power, M.J. Impact of a drier Early–Mid-Holocene climate upon Amazonian forests. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 1829–1838. [Google Scholar] [CrossRef]
- Guimarães, J.T.F.; Cohen, M.C.L.; Pessenda, L.C.R.; França, M.C.; Smith, C.B.; Nogueira, A.C.R. Mid and late Holocene sedimentary process and palaeovegetation changes near the mouth of the Amazon River. Holocene 2012, 22, 359–370. [Google Scholar] [CrossRef]
- Prado, L.F.; Wainer, I.; Chiessi, C.M.; Ledru, M.-P.; Turcq, B. A mid-Holocene climate reconstruction for eastern South America. Clim. Past 2013, 9, 2117–2133. [Google Scholar] [CrossRef]
- Maksic, J.; Shimizu, M.H.; De Oliveira, G.S.; Venancio, I.; Cardoso, M.; Ferreira, F.A. Simulation of the Holocene climate over South America and impacts on the vegetation. Holocene 2018, 29, 287–299. [Google Scholar] [CrossRef]
- Sifeddine, A.; Frohlich, F.; Fournier, M.; Martin, L.; Servant, M.; Soubiès, F.; Turcq, B.; Suguio, K.; Volkmer-Ribeiro, C. La sédimentation lacustre indicateur de changements des paléoenvironnements au cours des 30 000 dernières années (Caraias, Amazonie, Brésil). Académie Sci. Paris 1994, 318, 1645–1652. [Google Scholar]
- Pinakhina, D.V.; Chekunova, E.M. Environmental DNA: History of studies, current and perspective applications in fundamental and applied research. Ecol. Genet. 2020, 18, 493–509. [Google Scholar] [CrossRef]
- Thomsen, P.F.; Willerslev, E. Environmental DNA—An emerging tool in conservation for monitoring past and present biodiversity. Biol. Conserv. 2015, 183, 4–18. [Google Scholar] [CrossRef]
- Bohmann, K.; Evans, A.; Gilbert, M.T.P.; Carvalho, G.R.; Creer, S.; Knapp, M.; Yu, D.W.; De Bruyn, M. Environmental DNA for wildlife biology and biodiversity monitoring. Trends Ecol. Evol. 2014, 29, 358–367. [Google Scholar] [CrossRef] [PubMed]
- Haile, J.; Holdaway, R.; Oliver, K.; Bunce, M.; Gilbert, M.; Nielsen, R.; Munch, K.; Ho, S.Y.W.; Shapiro, B.; Willerslev, E. Ancient DNA Chronology within Sediment Deposits: Are Paleobiological Reconstructions Possible and Is DNA Leaching a Factor? Mol. Biol. Evol. 2007, 24, 982–989. [Google Scholar] [CrossRef] [PubMed]
- Willerslev, E.; Hansen, A.J.; Binladen, J.; Brand, T.B.; Gilbert, M.T.P.; Shapiro, B.; Bunce, M.; Wiuf, C.; Gilichinsky, D.A.; Cooper, A. Diverse Plant and Animal Genetic Records from Holocene and Pleistocene Sediments. Science 2003, 300, 791–795. [Google Scholar] [CrossRef]
- Yoccoz, N.; Bråthen, K.A.; Gielly, L.; Haile, J.; Edwards, M.E.; Goslar, T.; Von Stedingk, H.; Brysting, A.K.; Coissac, E.; Pompanon, F.; et al. DNA from soil mirrors plant taxonomic and growth form diversity. Mol. Ecol. 2012, 21, 3647–3655. [Google Scholar] [CrossRef] [PubMed]
- Giguet-Covex, C.; Pansu, J.; Arnaud, F.; Rey, P.-J.; Griggo, C.; Gielly, L.; Domaizon, I.; Coissac, E.; David, F.; Choler, P.; et al. Long livestock farming history and human landscape shaping revealed by lake sediment DNA. Nat. Commun. 2014, 5, 3211. [Google Scholar] [CrossRef] [PubMed]
- Correa-Metrio, A.; Bush, M.B.; Cabrera, K.R.; Sully, S.; Brenner, M.; Hodell, D.A.; Escobar, J.; Guilderson, T. Rapid climate change and no-analog vegetation in lowland Central America during the last 86,000 years. Quat. Sci. Rev. 2012, 38, 63–75. [Google Scholar] [CrossRef]
- Correa-Metrio, A.; Urrego, D.H.; Cabrera, K.R.; Bush, M.B. PaleoMAS: Paleo-ecological Analysis, R Package Version 2.0-1 ed. The R Project for Statistical Computing. 2011. Available online: http://CRAN.R-project.org/package=paleoMAS (accessed on 20 February 2019).
- Holtvoeth, J.; Whiteside, J.H.; Engels, S.; Freitas, F.S.; Grice, K.; Greenwood, P.; Johnson, S.; Kendall, I.; Lengger, S.K.; Lücke, A.; et al. The paleolimnologist’s guide to compound-specific stable isotope analysis—An introduction to principles and applications of CSIA for Quaternary lake sediments. Quat. Sci. Rev. 2019, 207, 101–133. [Google Scholar] [CrossRef]
- Sachse, D.; Billault, I.; Bowen, G.J.; Chikaraishi, Y.; Dawson, T.E.; Feakins, S.J.; Freeman, K.H.; Magill, C.R.; McInerney, F.A.; van der Meer, M.T.; et al. Molecular Paleohydrology: Interpreting the Hydrogen-Isotopic Composition of Lipid Biomarkers from Photosynthesizing Organisms. Annu. Rev. Earth Planet. Sci. 2012, 40, 221–249. [Google Scholar] [CrossRef]
- Piló, L.B.; Auler, A.S.; Martins, F. Carajás National Forest: Iron Ore Plateaus and Caves in Southeastern Amazon. In Landscapes and Landforms of Brazil, World Geomorphological Landscapes; Vieira, B.C., Salgado, A.A.R., Santos, L.J.C., Eds.; Springer: Berlin/Heidelberg, Germany, 2015; pp. 273–283. [Google Scholar]
- Piló, L.B.; Calux, A.; Scherer, R.; Bernard, E. Bats as ecosystem engineers in iron ore caves in the Carajás National Forest, Brazilian. bioRxiv 2022. [Google Scholar] [CrossRef]
- Bird, M.I.; Boobyer, E.M.; Bryant, C.; Lewis, H.A.; Paz, V.; Stephens, W.E. A long record of environmental change from bat guano deposits in Makangit Cave, Palawan, Philippines. Earth Environ. Sci. Trans. R. Soc. Edinb. 2007, 98, 59–69. [Google Scholar] [CrossRef]
- Widga, C.; Colburn, M. Paleontology and paleoecology of guano deposits in Mammoth Cave, Kentucky, USA. Quat. Res. 2015, 83, 427–436. [Google Scholar] [CrossRef]
- Wurster, C.; McFarlane, D.; Bird, M.; Ascough, P.; Athfield, N.B. Stable Isotopes of Subfossil Bat Guano as a Long-Term Environmental Archive: Insights from a Grand Canyon Cave Deposit. J. Cave Karst Stud. 2010, 72, 111–121. [Google Scholar] [CrossRef]
- Wurster, C.; Patterson, W.; McFarlane, D.A.; Wassenaar, L.; Hobson, K.A.; Athfield, N.B.; Bird, M. Stable carbon and hydrogen isotopes from bat guano in the Grand Canyon, USA, reveal Younger Dryas and 8.2 ka events. Geology 2008, 36, 683. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vegetation Type | Species |
|---|---|
| Canga | Dyckia duckei (Bromeliaceae), Ipomoea marabensis (Convolvulaceae), Erythroxylum nelson-rosae (Erythroxylaceae), Bauhinia pulchella, Mimosa acutistipula var. ferrea and M. skinneri var. carajarum (Fabaceae), Byrsonima chrysophylla (Malpighiaceae), Norantea goyasensis (Marcgraviaceae), Tibouchina edmundoi and Pleroma carajasensis (Melastomataceae), Sobralia liliastrum (Orchidaceae), Axonopus carajasensis, Panicum millegrana, Paspalum cangarum, P. carajasense and Sporobolus multiramosus (Poaceae), Borreria (Rubiaceae), Vellozia glauca (Velloziaceae), Callisthene microphylla (Vochysiaceae). |
| SDF | Licania blackii (Chrysobalanaceae), Aparisthmium cordatum and Maprounea guianensis (Euphorbiaceae), Myrcia splendens (Myrtaceae), Henriettea ramiflora, Miconia chrysophylla (Melastomataceae), Matayba inelegans (Sapindaceae). |
| HETF | Tapirira guianensis (Anacardiaceae), Guatteria punctata and G. tomentosa (Annonaceae), Oenocarpus distichus (Arecaceae), Cordia sellowiana (Boraginaceae), Licania laxiflora (Chrysobalanaceae), Deguelia negrensis, Pseudopiptadenia suaveolens (Fabaceae), Emmotum nitens (Icacinaceae), Sacoglottis mattogrossensis (Humiriaceae), Mezilaurus itauba (Lauraceae), Miconia piperifolia (Melastomataceae), Myrcia silvatica (Myrtaceae), Neea ovalifolia (Nyctaginaceae), Matayba arborescens (Sapindaceae), Erisma uncinatum (Vochysiaceae). |
| Filled lakes | Mauritiella armata (Arecaceae), Eleocharis pedroviane, Cyperus, Scleria (Cyperaceae), Hydrochorea corymbosa (Fabaceae), Miconia alternans (Melastomataceae), Styrax griseus (Styracaceae). |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guimarães, J.T.F.; Sahoo, P.K.; e Souza-Filho, P.W.M.; da Silva, M.S.; Rodrigues, T.M.; da Silva, E.F.; Reis, L.S.; de Figueiredo, M.M.J.C.; Lopes, K.d.S.; Moraes, A.M.; et al. Landscape and Climate Changes in Southeastern Amazonia from Quaternary Records of Upland Lakes. Atmosphere 2023, 14, 621. https://doi.org/10.3390/atmos14040621
Guimarães JTF, Sahoo PK, e Souza-Filho PWM, da Silva MS, Rodrigues TM, da Silva EF, Reis LS, de Figueiredo MMJC, Lopes KdS, Moraes AM, et al. Landscape and Climate Changes in Southeastern Amazonia from Quaternary Records of Upland Lakes. Atmosphere. 2023; 14(4):621. https://doi.org/10.3390/atmos14040621
Chicago/Turabian StyleGuimarães, José Tasso Felix, Prafulla Kumar Sahoo, Pedro Walfir Martins e Souza-Filho, Marcio Sousa da Silva, Tarcísio Magevski Rodrigues, Edilson Freitas da Silva, Luiza Santos Reis, Mariana Maha Jana Costa de Figueiredo, Karen da Silva Lopes, Aline Mamede Moraes, and et al. 2023. "Landscape and Climate Changes in Southeastern Amazonia from Quaternary Records of Upland Lakes" Atmosphere 14, no. 4: 621. https://doi.org/10.3390/atmos14040621
APA StyleGuimarães, J. T. F., Sahoo, P. K., e Souza-Filho, P. W. M., da Silva, M. S., Rodrigues, T. M., da Silva, E. F., Reis, L. S., de Figueiredo, M. M. J. C., Lopes, K. d. S., Moraes, A. M., Leite, A. S., da Silva Júnior, R. O., Salomão, G. N., & Dall’Agnol, R. (2023). Landscape and Climate Changes in Southeastern Amazonia from Quaternary Records of Upland Lakes. Atmosphere, 14(4), 621. https://doi.org/10.3390/atmos14040621