
The Influence of pH Dynamics on Modeled Ammonia Emission Patterns of a Naturally Ventilated Dairy Cattle Building





Abstract
:1. Introduction
2. Materials and Methods
2.1. Farm Site and Data Collection
2.2. The Nested Model
2.3. Emission Value Derivation
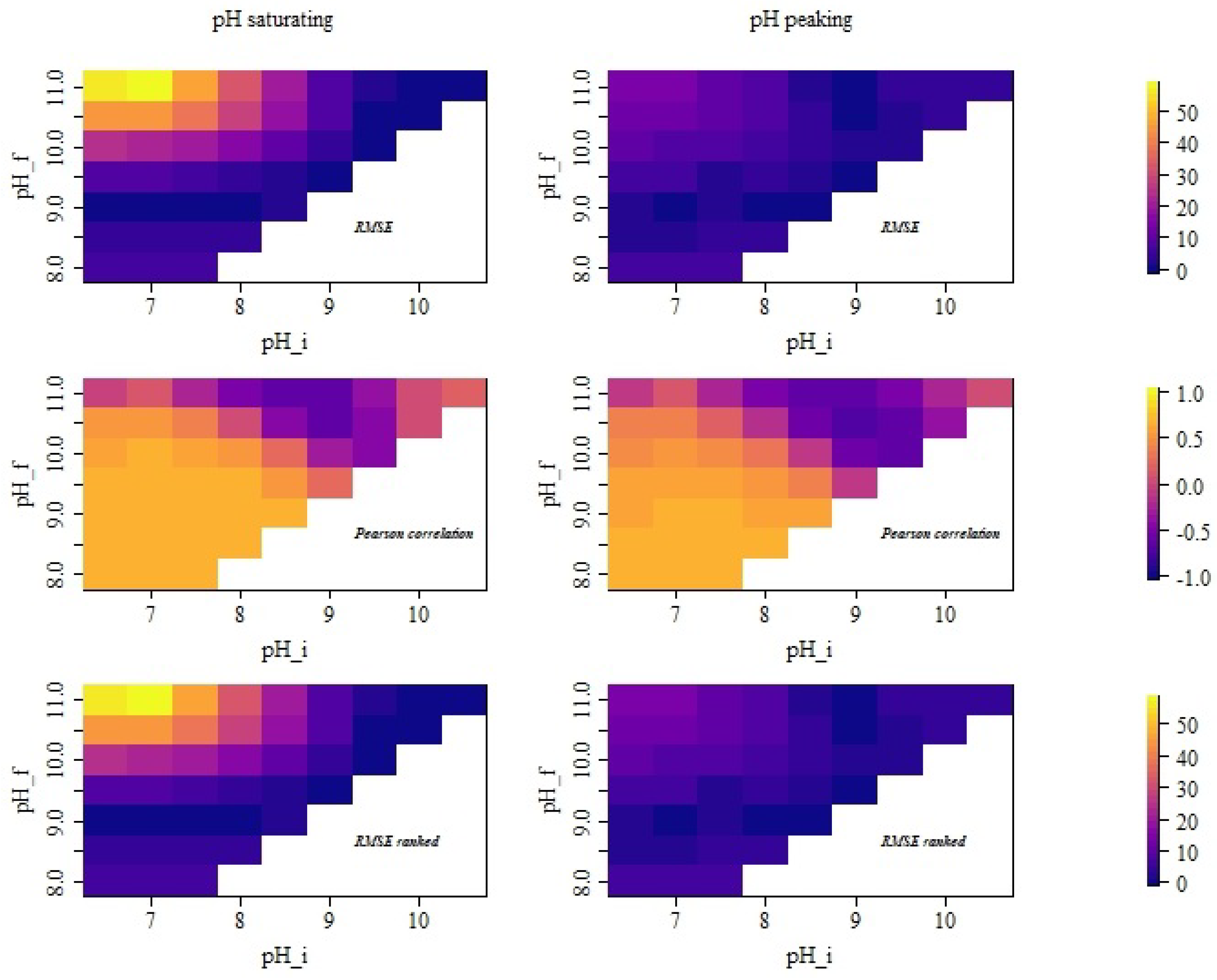
2.4. Data Analysis
3. Results
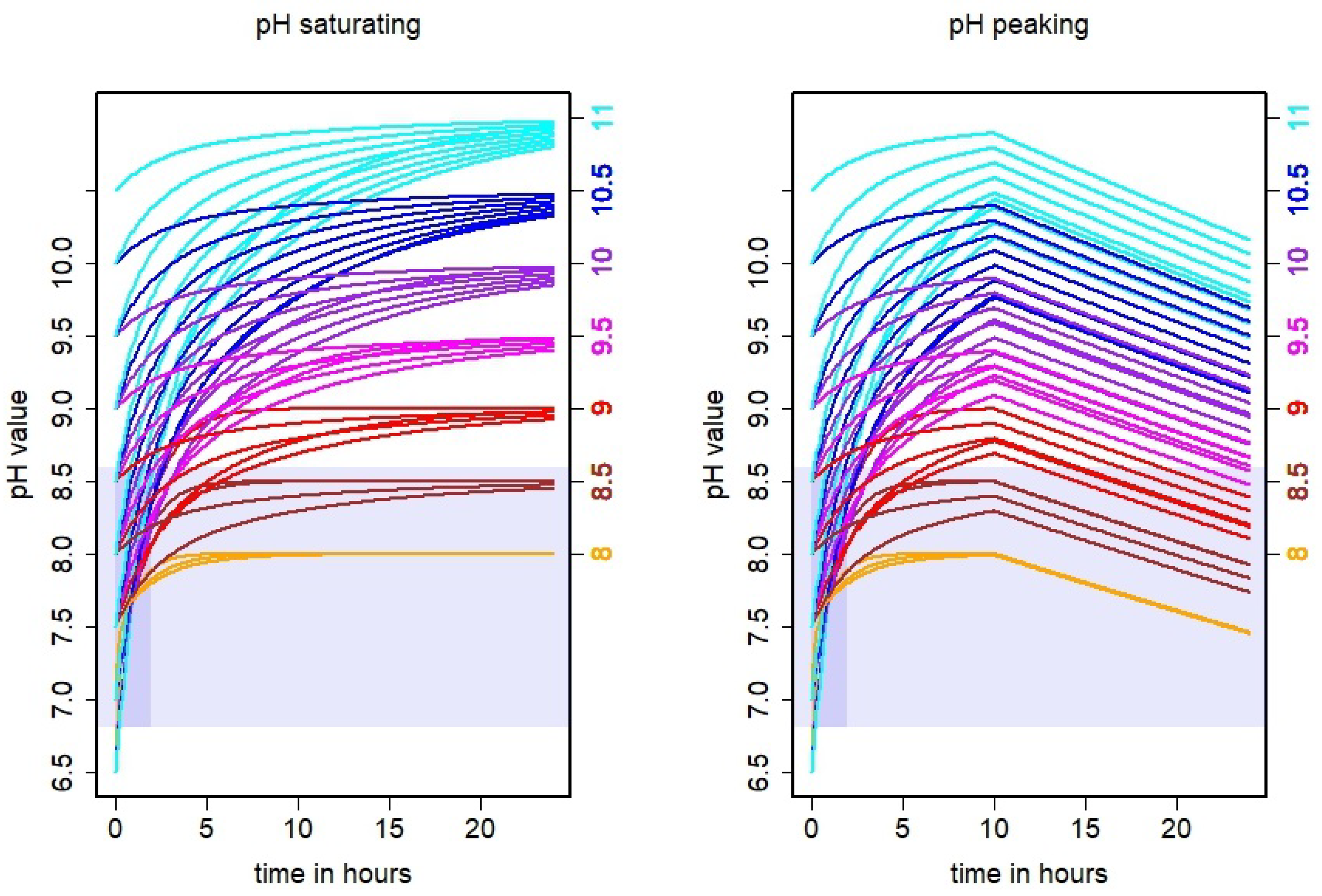
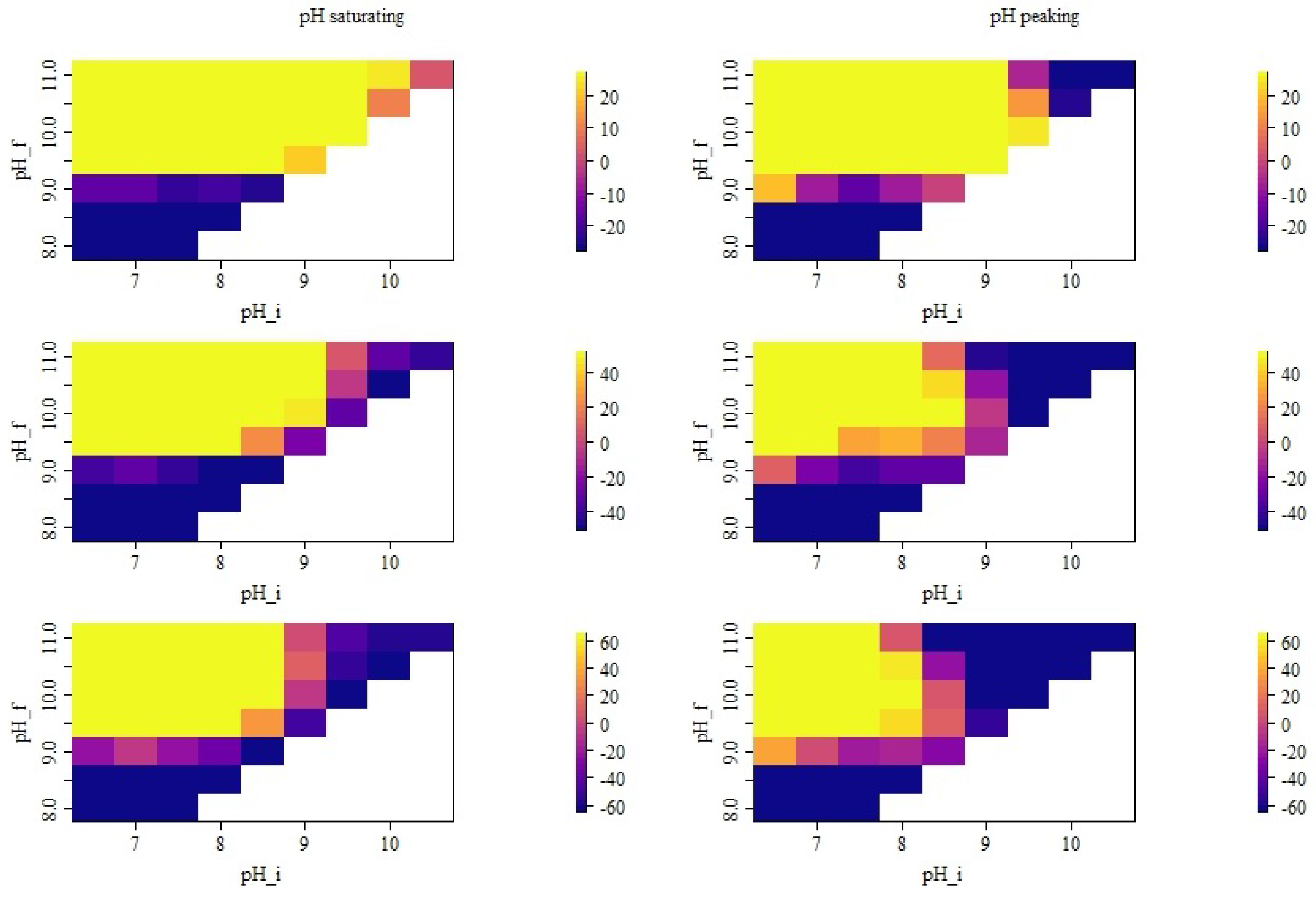
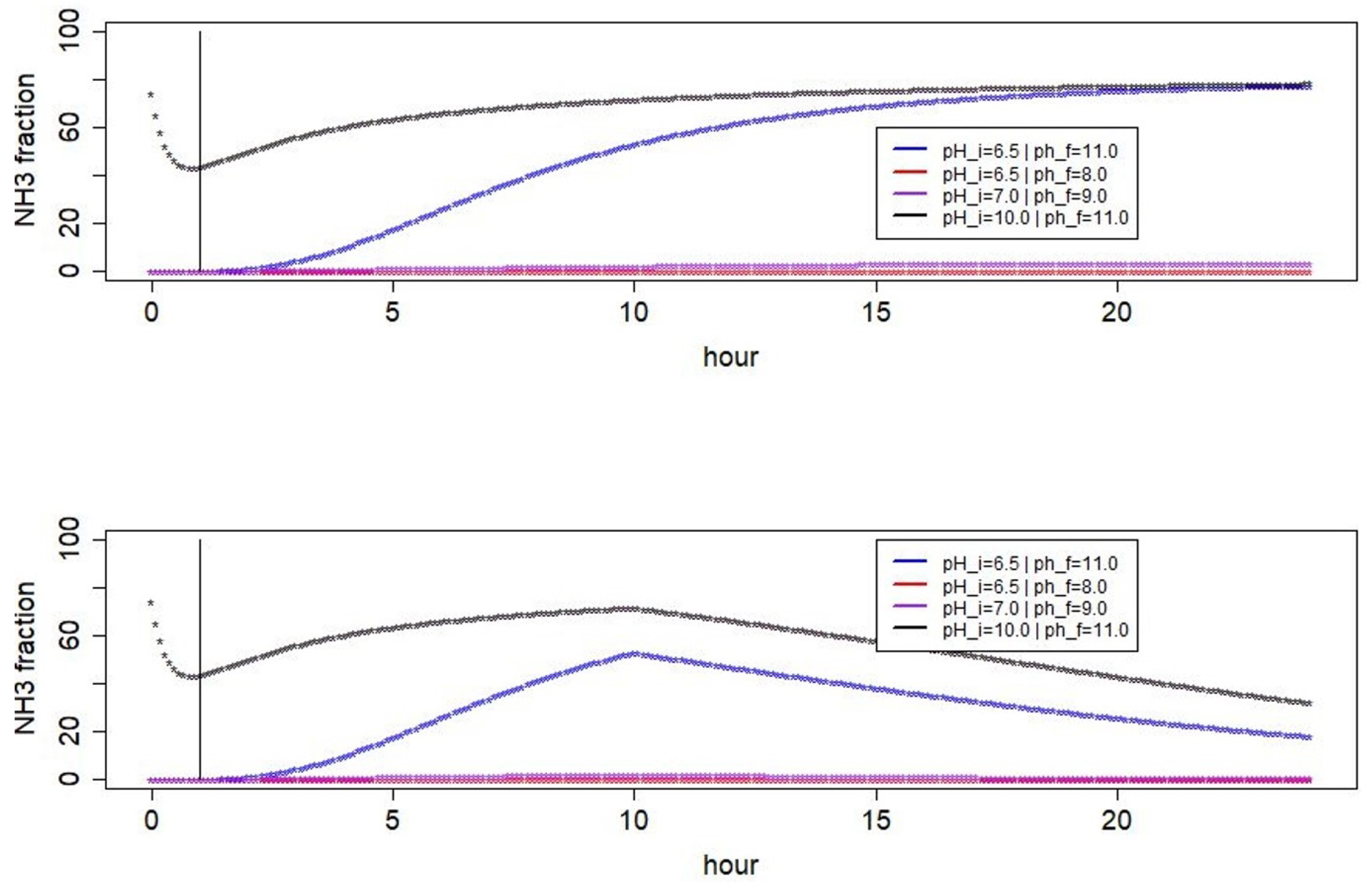
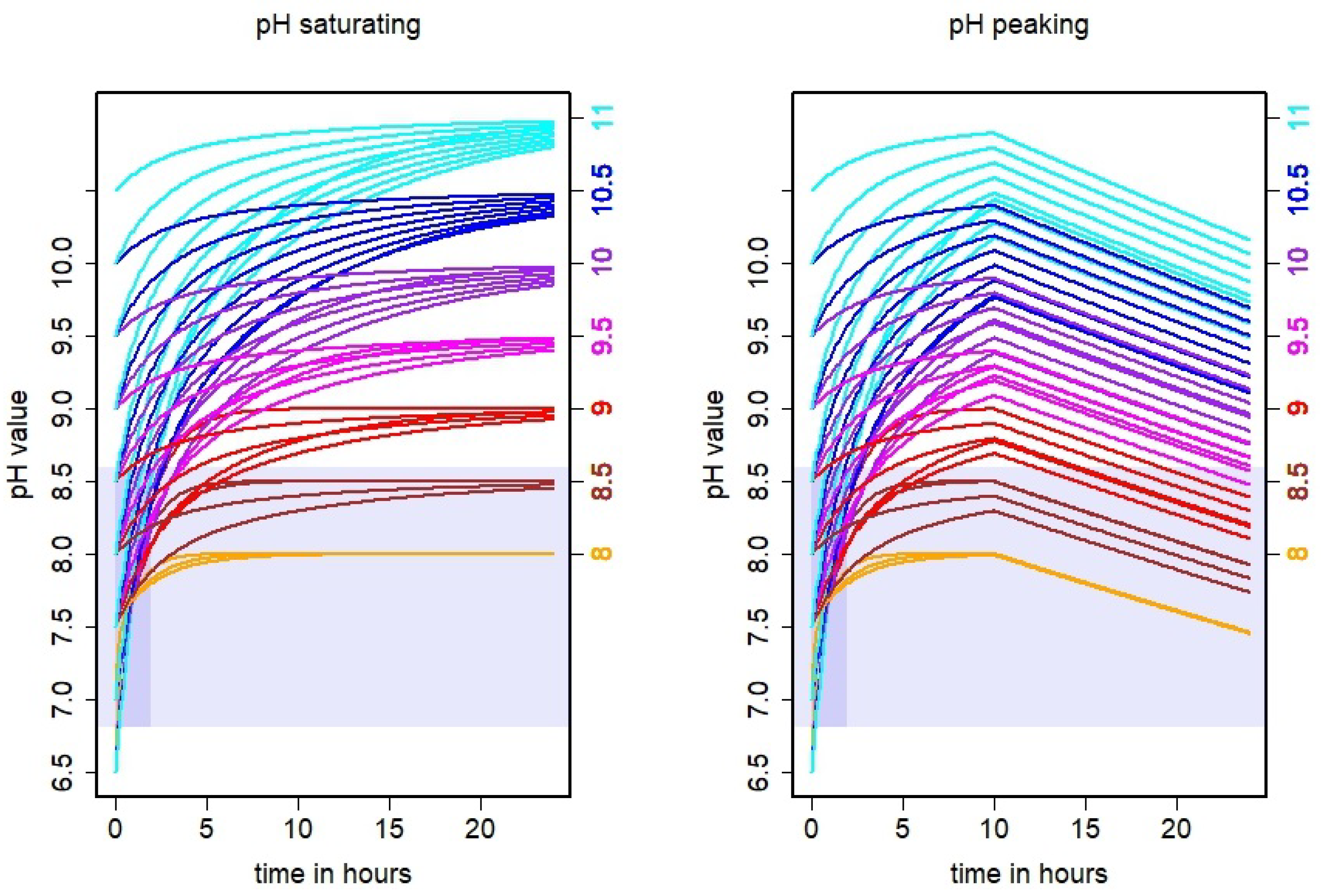
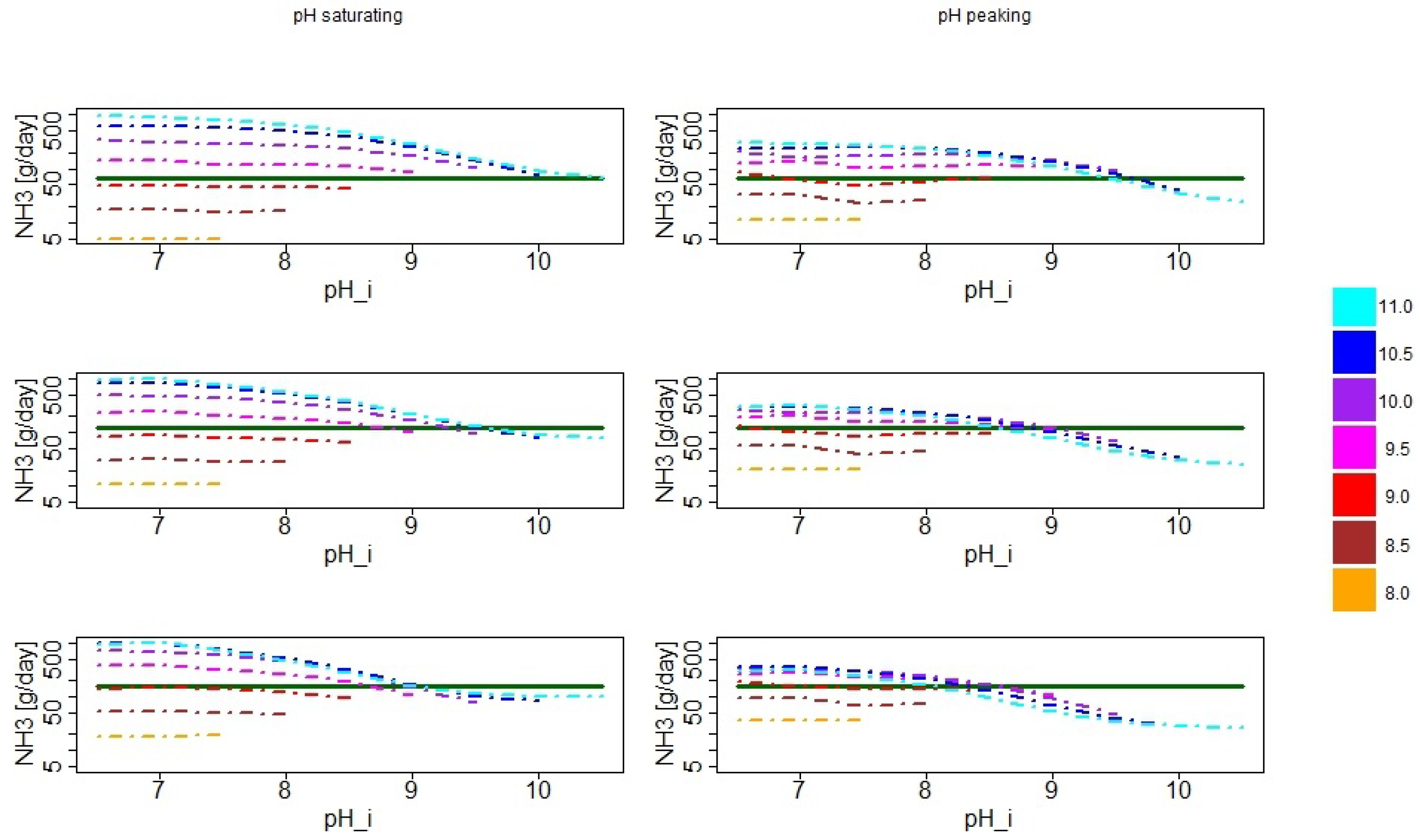
3.1. Simulated pH Dynamics
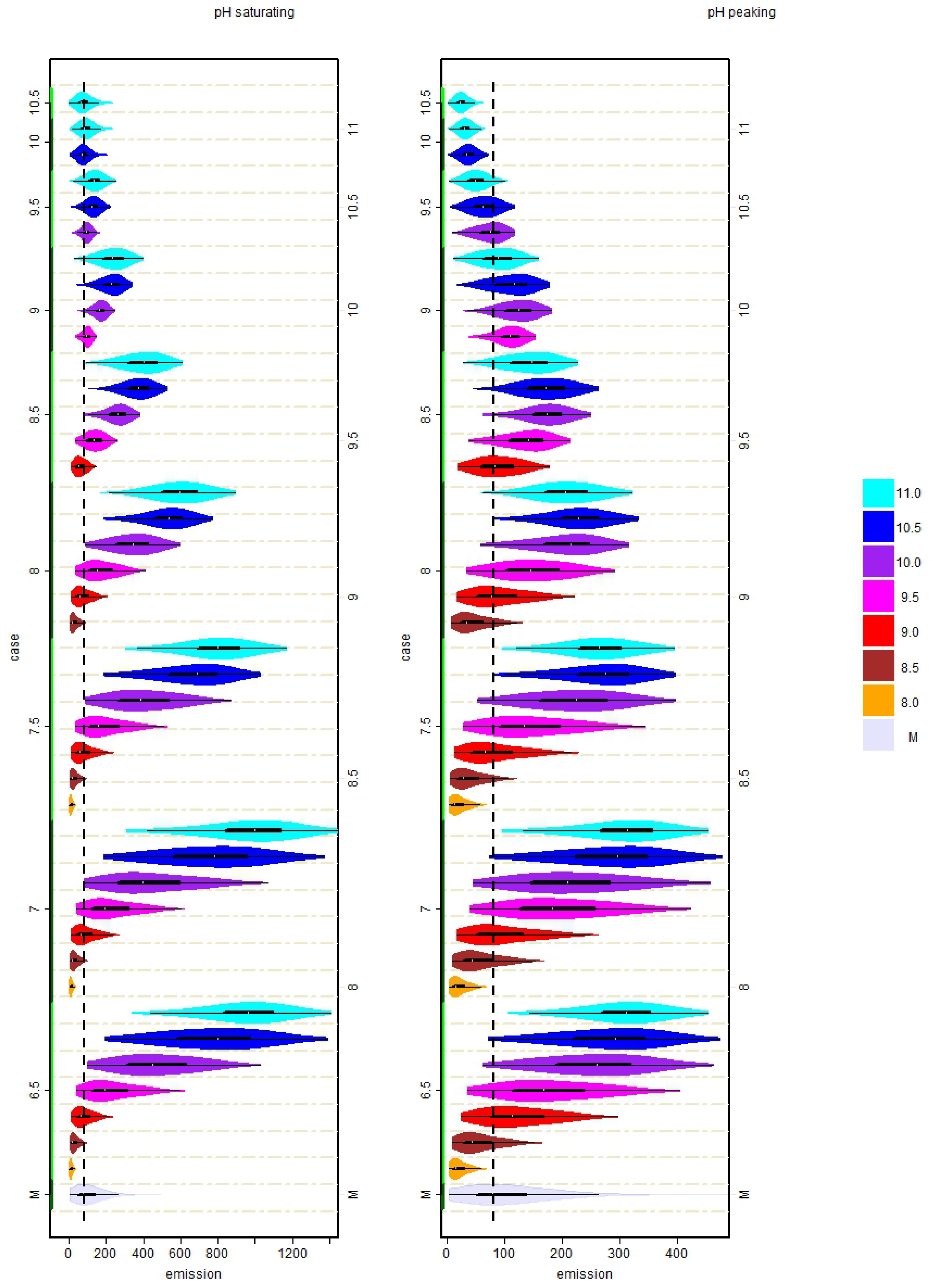
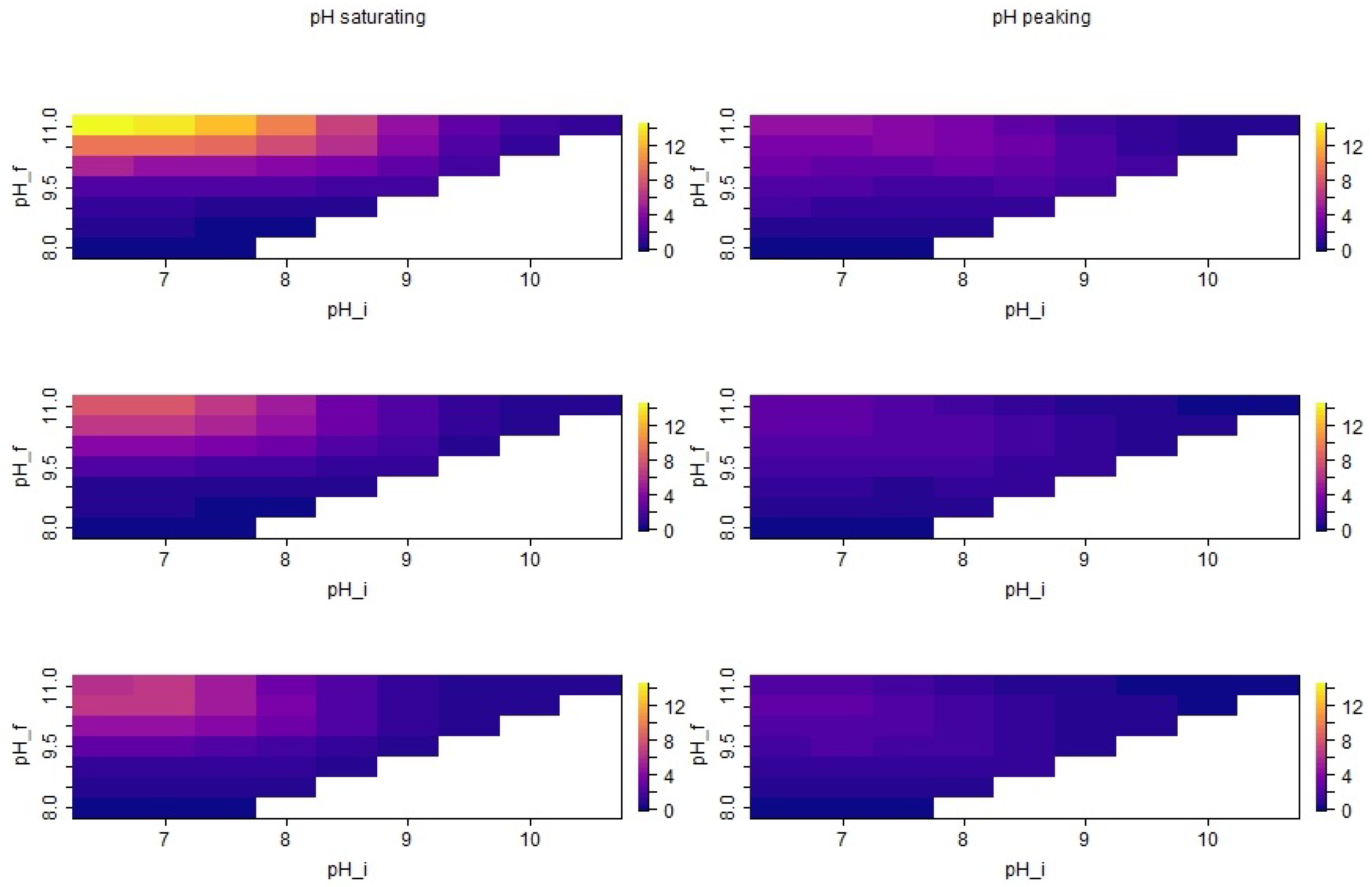
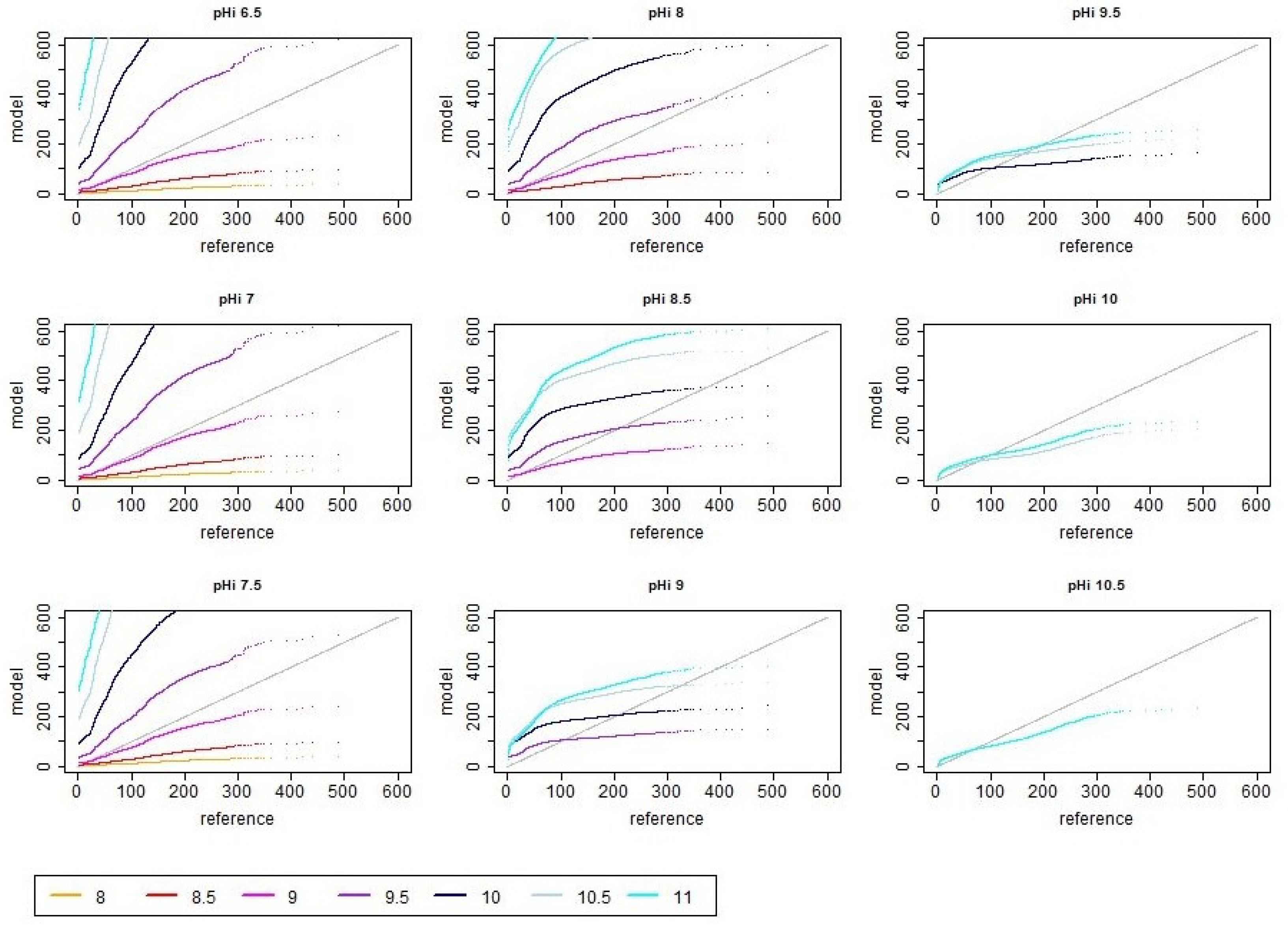
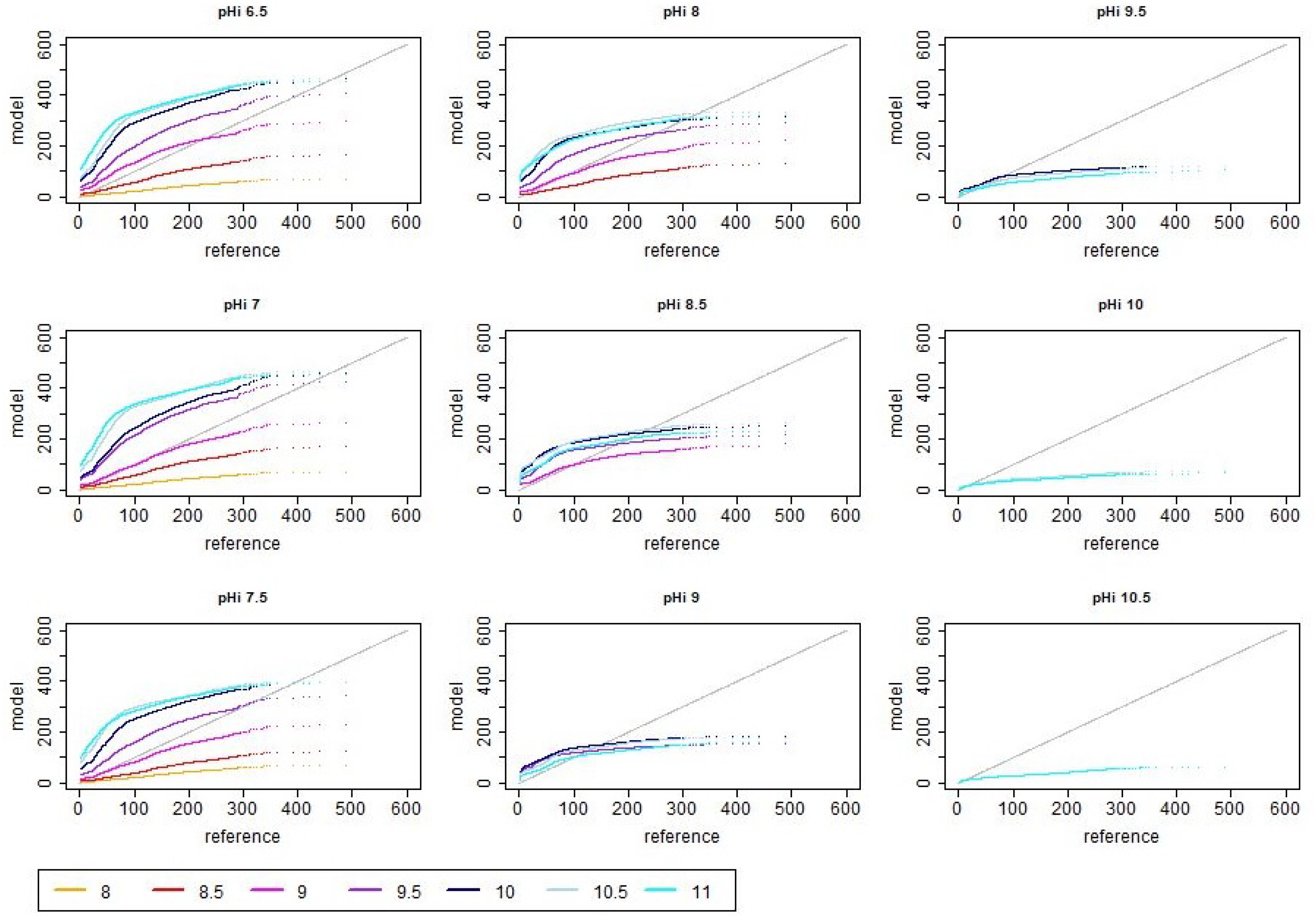
3.2. Distribution of Simulated Emission Values
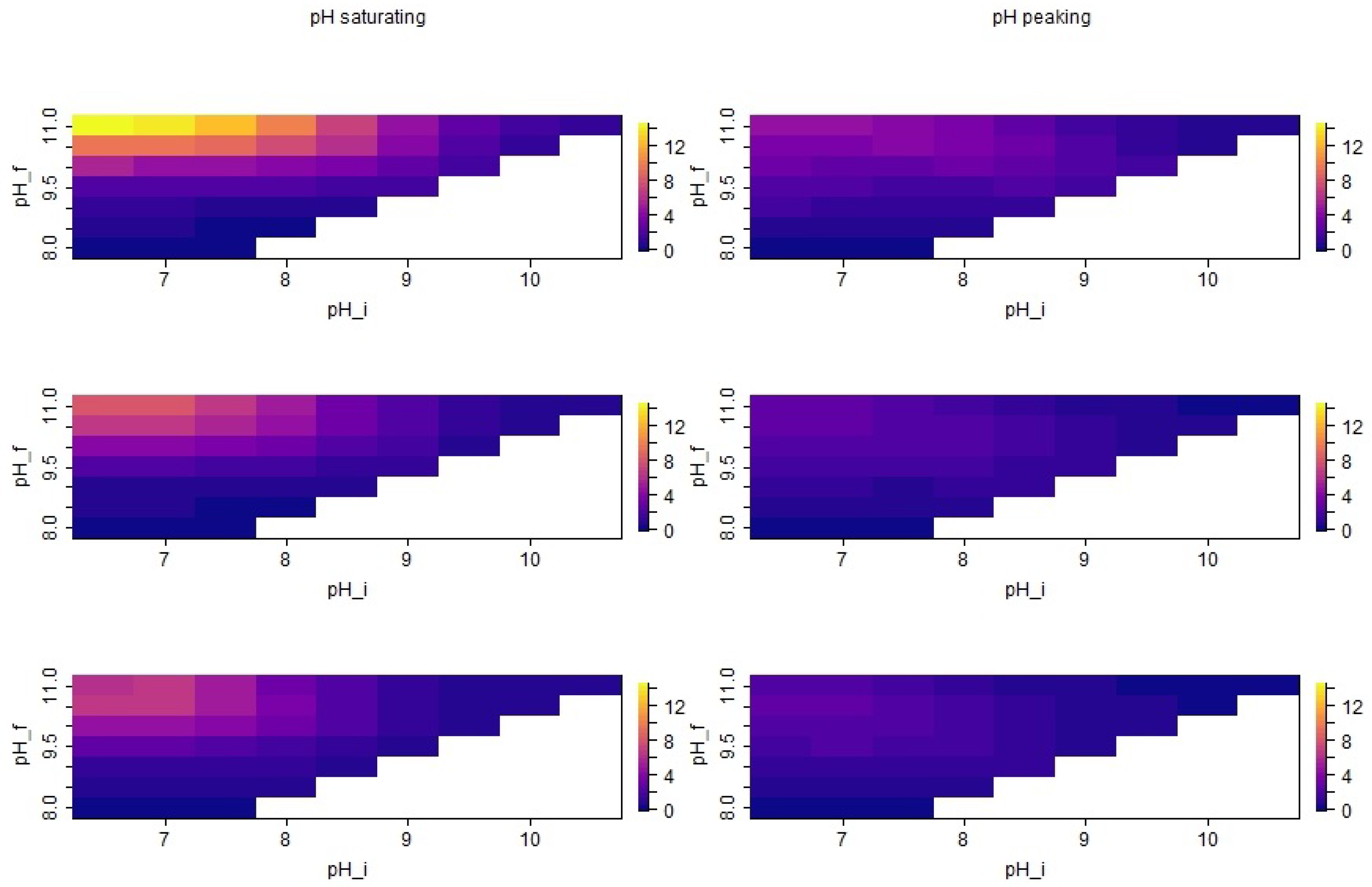
3.3. Seasonal Effects of pH Dynamics
4. Discussion
4.1. Conclusions on Actual pH Dynamics
4.2. Impact of pH Dynamics on Total Ammonia Emission
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| NH | Ammonia |
| CO | Carbon dioxide |
| NH | Ammonium |
| FTIR | Fourier transform infrared |
| DMI | Dry matter intake |
| TAN | Total ammoniacal nitrogen |
Appendix A
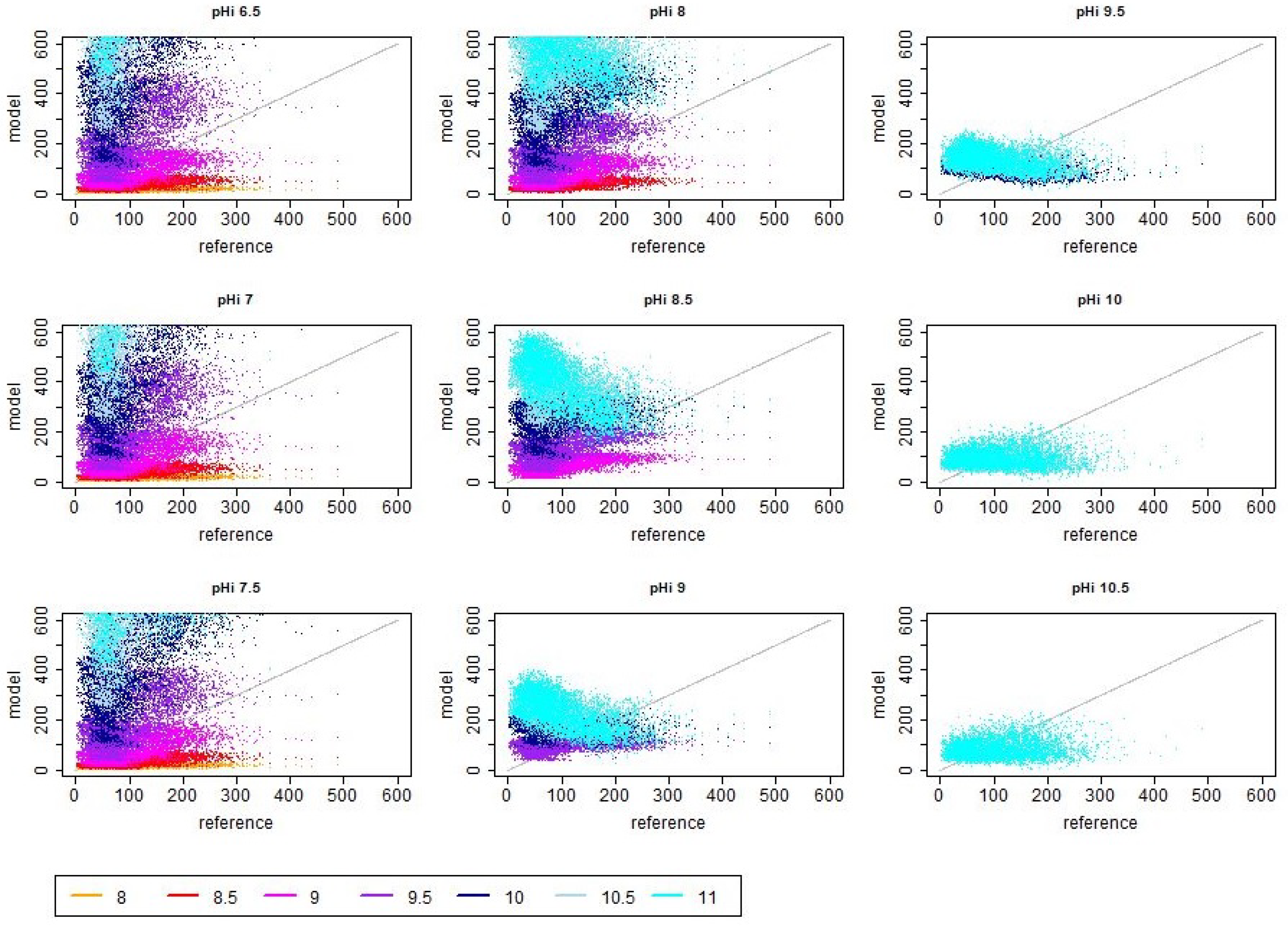
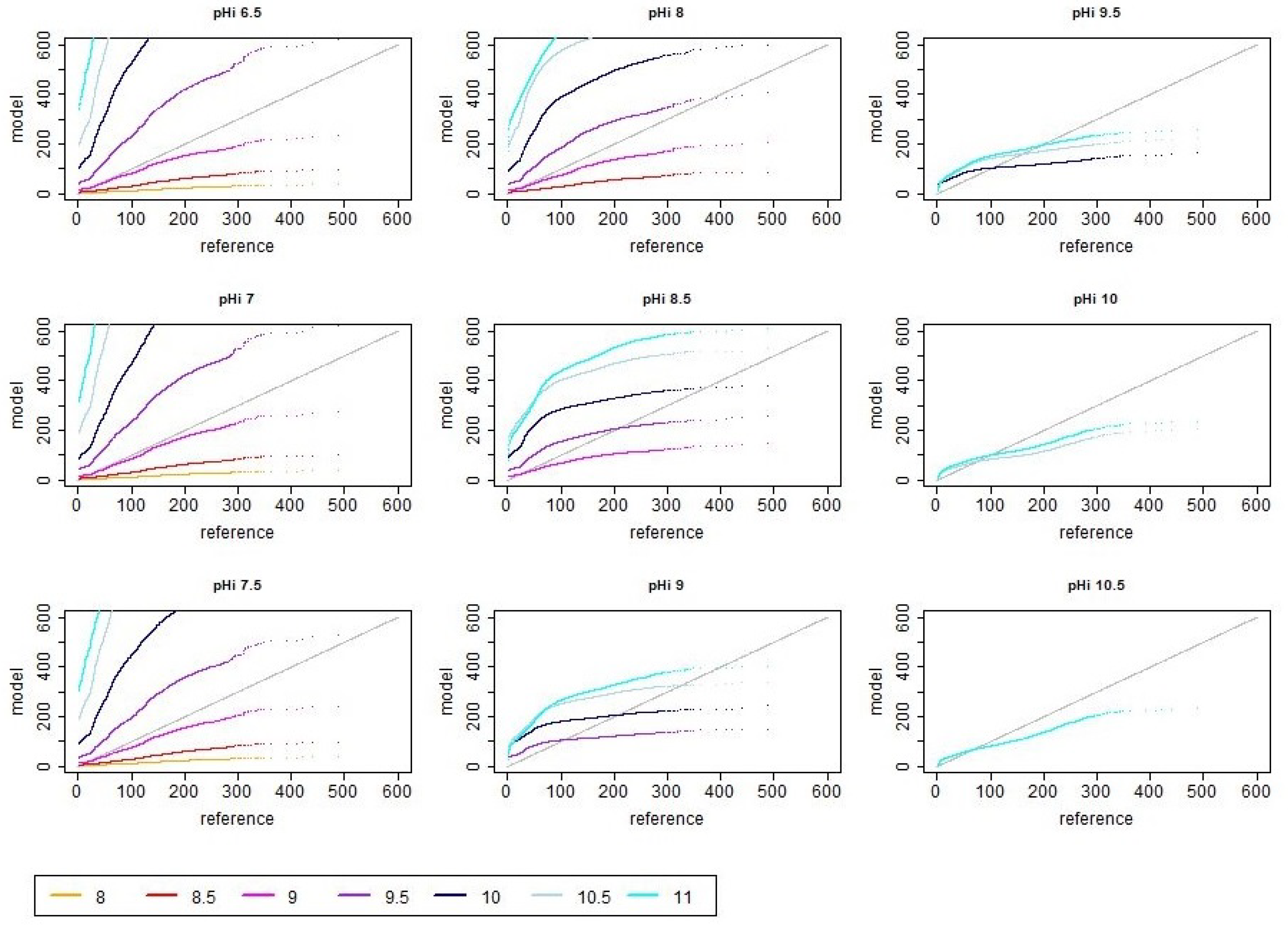
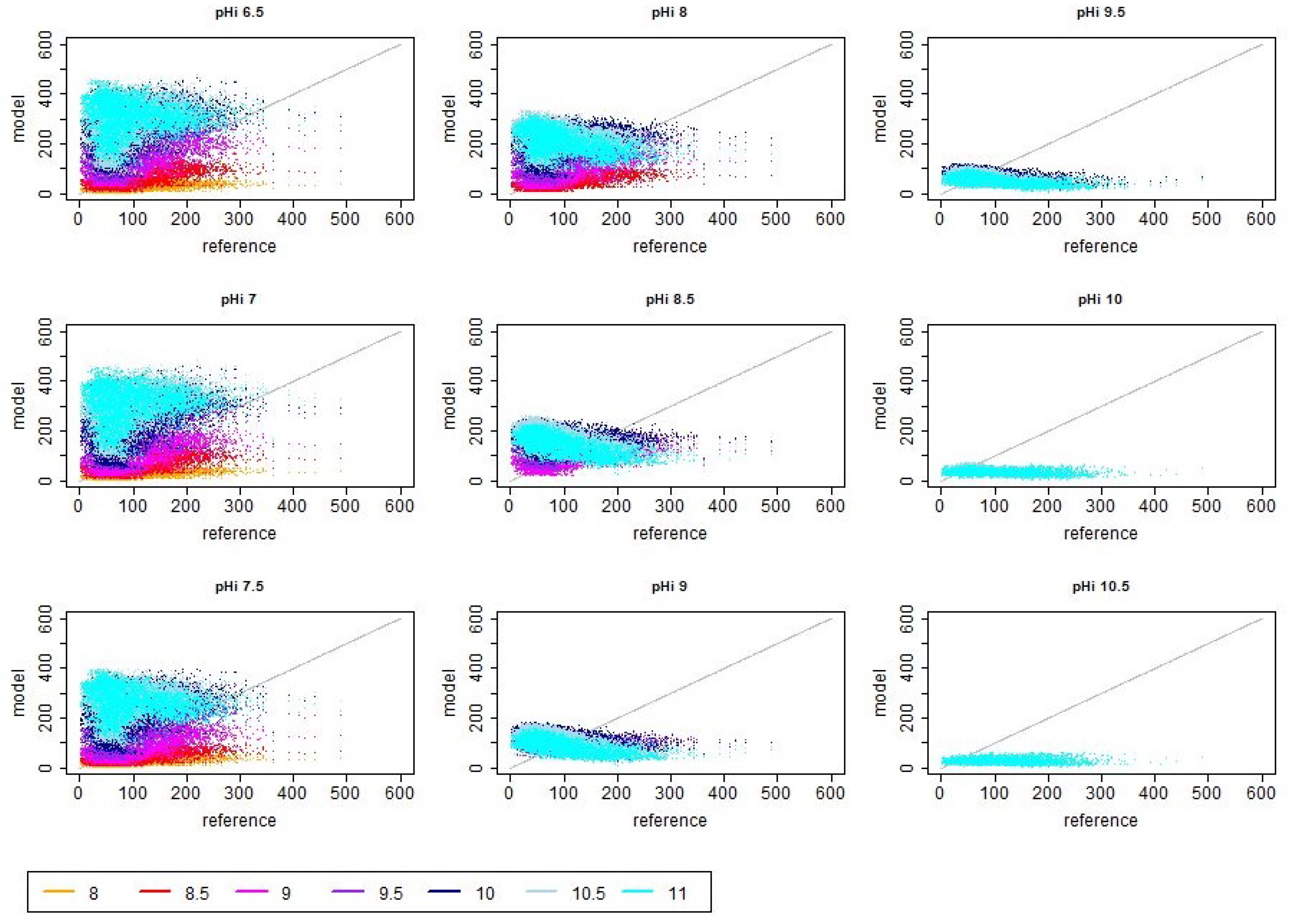
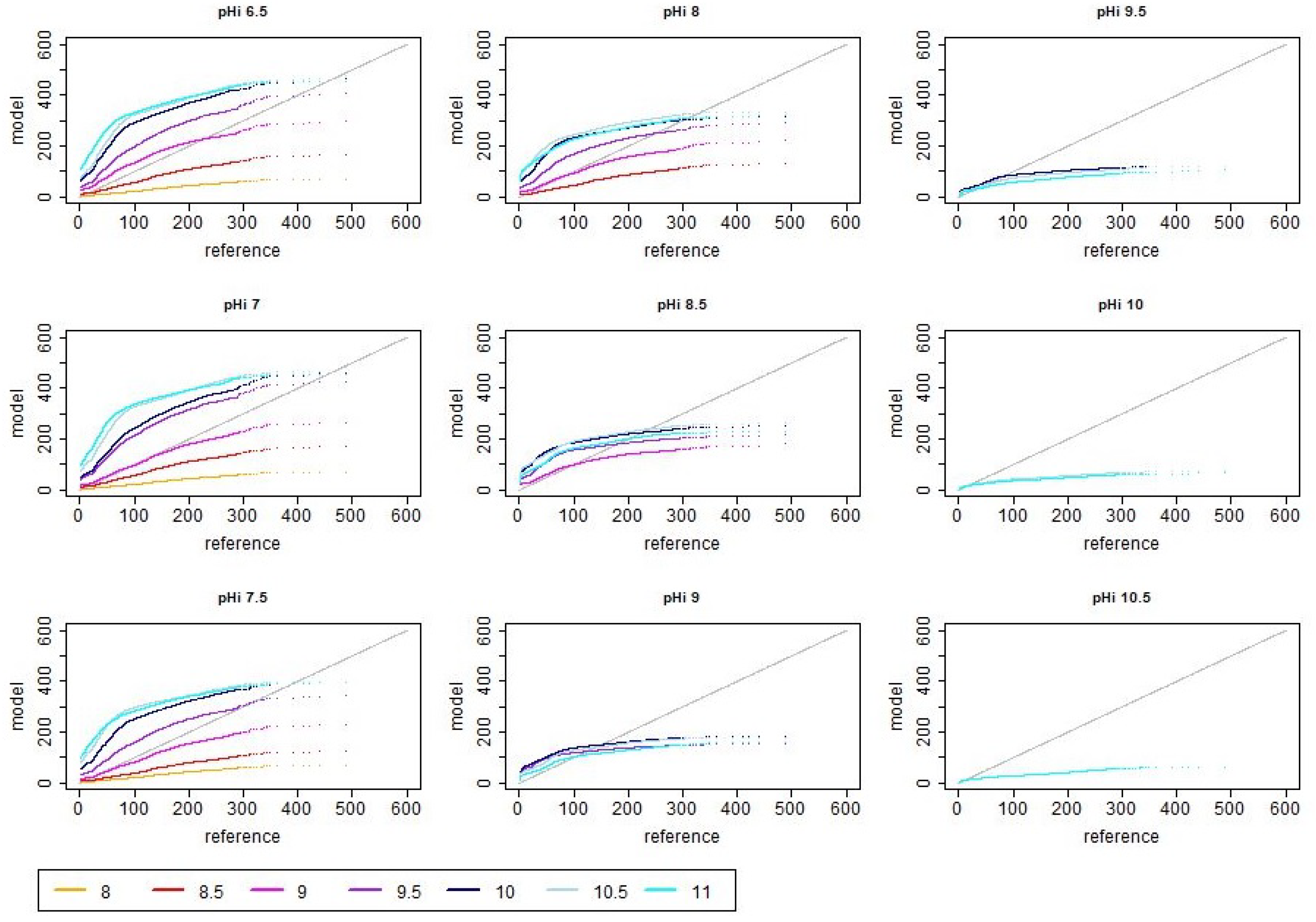
References
- Huang, T.; Gao, B.; Hu, X.K.; Lu, X.; Well, R.; Christie, P.; Bakken, L.R.; Ju, X.T. Ammonia-oxidation as an engine to generate nitrous oxide in an intensively managed calcareous Fluvo-aquic soil. Sci. Rep. 2014, 4, 3950. [Google Scholar] [CrossRef] [PubMed]
- Behera, S.N.; Sharma, M. Degradation of SO2, NO2 and NH3 leading to formation of secondary inorganic aerosols: An environmental chamber study. Atmos. Environ. 2011, 45, 4015–4024. [Google Scholar] [CrossRef]
- OECD. Trends and Drivers of Agri-Environmental Performance in OECD Countries; OECD Publishing: Paris, France, 2019; p. 105. [Google Scholar] [CrossRef]
- European Environment Agency (EEA). European Union Emission Inventory Report 1990–2017; European Environment Agency (EEA): Copenhagen, Denmark, 2019; Volume 8.
- Dai, X.; Karring, H. A determination and comparison of urease activity in feces and fresh manure from pig and cattle in relation to ammonia production and pH changes. PLoS ONE 2014, 9, e110402. [Google Scholar] [CrossRef] [PubMed]
- Vaddella, V.; Ndegwa, P.; Jiang, A. An empirical model of ammonium ion dissociation in liquid dairy manure. Trans. ASABE 2011, 54, 1119–1126. [Google Scholar] [CrossRef]
- Hempel, S.; Ouatahar, L.; Janke, D.; Doumbia, E.M.; Willink, D.; Amon, B.; Bannink, A.; Amon, T. Ammonia emission prediction for dairy cattle housing from reaction kinetic modeling to the barn scale. Comput. Electron. Agric. 2022, 199, 107168. [Google Scholar] [CrossRef]
- Mielcarek-Bocheńska, P.; Rzeźnik, W. Odors and ammonia emission from a mechanically ventilated fattening piggery on deep litter in Poland. Arch. Environ. Prot. 2022, 48, 86–94. [Google Scholar]
- Snell, H.; Seipelt, F.; Van den Weghe, H. Ventilation rates and gaseous emissions from naturally ventilated dairy houses. Biosyst. Eng. 2003, 86, 67–73. [Google Scholar] [CrossRef]
- Janke, D.; Willink, D.; Ammon, C.; Hempel, S.; Schrade, S.; Demeyer, P.; Hartung, E.; Amon, B.; Ogink, N.; Amon, T. Calculation of ventilation rates and ammonia emissions: Comparison of sampling strategies for a naturally ventilated dairy barn. Biosyst. Eng. 2020, 198, 15–30. [Google Scholar] [CrossRef]
- Bannink, A.; Spek, W.J.; Dijkstra, J.; Šebek, L.B. A Tier 3 method for enteric methane in dairy cows applied for fecal N digestibility in the ammonia inventory. Front. Sustain. Food Syst. 2018, 2, 66. [Google Scholar] [CrossRef]
- Bjerg, B.; Cascone, G.; Lee, I.B.; Bartzanas, T.; Norton, T.; Hong, S.W.; Seo, I.H.; Banhazi, T.; Liberati, P.; Marucci, A.; et al. Modelling of ammonia emissions from naturally ventilated livestock buildings. Part 3: CFD modelling. Biosyst. Eng. 2013, 116, 259–275. [Google Scholar] [CrossRef]
- Saha, C.K.; Zhang, G.; Ni, J.Q. Airflow and concentration characterisation and ammonia mass transfer modelling in wind tunnel studies. Biosyst. Eng. 2010, 107, 328–340. [Google Scholar] [CrossRef]
- Rong, L.; Nielsen, P.V.; Zhang, G. Effects of airflow and liquid temperature on ammonia mass transfer above an emission surface: Experimental study on emission rate. Bioresour. Technol. 2009, 100, 4654–4661. [Google Scholar] [CrossRef] [PubMed]
- Elzing, A.; Monteny, G. Modeling and experimental determination of ammonia emissions rates from a scale model dairy-cow house. Trans. ASAE 1997, 40, 721–726. [Google Scholar] [CrossRef]
- Snoek, D.J.; Stigter, J.D.; Ogink, N.W.; Koerkamp, P.W.G. Sensitivity analysis of mechanistic models for estimating ammonia emission from dairy cow urine puddles. Biosyst. Eng. 2014, 121, 12–24. [Google Scholar] [CrossRef]
- Melendez, P.; Bartolome, J.; Roeschmann, C.; Soto, B.; Arevalo, A.; Möller, J.; Coarsey, M. The association of prepartum urine pH, plasma total calcium concentration at calving and postpartum diseases in Holstein dairy cattle. Animal 2021, 15, 100148. [Google Scholar] [CrossRef] [PubMed]
- Snoek, D.J.; Stigter, J.D.; Kupers, G.C.; Koerkamp, P.W.G.; Ogink, N.W. Assessing fresh urine puddle chemistry in commercial dairy cow houses. Biosyst. Eng. 2017, 159, 143–153. [Google Scholar] [CrossRef]
- Kume, S.; Sato, T.; Murai, I.; Kitagawa, M.; Nonaka, K.; Oshita, T. Relationships between urine pH and electrolyte status in cows fed forages. Anim. Sci. J. 2011, 82, 456–460. [Google Scholar] [CrossRef] [PubMed]
- Gulhane, H.; Nakanekar, A.; Mahakal, N.; Bhople, S.; Salunke, A. Gomutra (cow urine): A multidimensional drug review article. Int. J. Res. Ayurveda Pharm. 2017, 8, 1–6. [Google Scholar] [CrossRef]
- Hempel, S.; Ouatahar, L.; Janke, D.; Amon, T. A nested semi-mechanistic model to predict the temporal dynamics of ammonia emissions from a solid floor naturally ventilated dairy cattle building—Parameter estimation and associated uncertainties. In Proceedings of the International Conference on Agricultural Engineering AgEng-LAND.TECHNIK 2022, Berlin, Germany, 22–23 November 2022; VDI Wissensforum GmbH: Düsseldorf, Germany, 2022; Volume 2406, pp. 197–202, ISBN 978-3-18092406-9. [Google Scholar]
- Tabase, R.K. Mimicking indoor climate dynamics and ammonia emissions in a pig housing compartment using artificial pigs and an automatic urea spraying installation. Agric. Eng. Int. Cigr J. 2019, 21, 40–50. [Google Scholar]
- Qu, Q.; Groot, J.C.; Zhang, K.; Schulte, R.P. Effects of housing system, measurement methods and environmental factors on estimating ammonia and methane emission rates in dairy barns: A meta-analysis. Biosyst. Eng. 2021, 205, 64–75. [Google Scholar] [CrossRef]
- Cortus, E.; Lemay, S.; Barber, E.; Hill, G.; Godbout, S. A dynamic model of ammonia emission from urine puddles. Biosyst. Eng. 2008, 99, 390–402. [Google Scholar] [CrossRef]
- Snoek, D.J.; Ogink, N.W.; Stigter, J.D.; Agricola, S.; Van de Weijer, T.M.; Koerkamp, P.W.G. Dynamic behavior of PH in fresh urine puddles of dairy cows. Trans. ASABE 2016, 59, 1403–1411. [Google Scholar]
- Sommer, S.; Sherlock, R. pH and buffer component dynamics in the surface layers of animal slurries. J. Agric. Sci. 1996, 127, 109–116. [Google Scholar] [CrossRef]
- Doumbia, E.M.; Janke, D.; Yi, Q.; Amon, T.; Kriegel, M.; Hempel, S. CFD modelling of an animal occupied zone using an anisotropic porous medium model with velocity depended resistance parameters. Comput. Electron. Agric. 2021, 181, 105950. [Google Scholar] [CrossRef]
- Piccione, G.; Caola, G.; Refinetti, R. Daily and estrous rhythmicity of body temperature in domestic cattle. BMC Physiol. 2003, 3, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Kadzere, C.T.; Murphy, M.; Silanikove, N.; Maltz, E. Heat stress in lactating dairy cows: A review. Livest. Prod. Sci. 2002, 77, 59–91. [Google Scholar] [CrossRef]
- Danscher, A.M.; Li, S.; Andersen, P.H.; Khafipour, E.; Kristensen, N.B.; Plaizier, J.C. Indicators of induced subacute ruminal acidosis (SARA) in Danish Holstein cows. Acta Vet. Scand. 2015, 57, 39. [Google Scholar] [CrossRef]
- Roche, J.R.; Dalley, D.E.; O’Mara, F.P. Effect of a metabolically created systemic acidosis on calcium homeostasis and the diurnal variation in urine pH in the non-lactating pregnant dairy cow. J. Dairy Res. 2007, 74, 34–39. [Google Scholar] [CrossRef]
- Ni, J. Mechanistic models of ammonia release from liquid manure: A review. J. Agric. Eng. Res. 1999, 72, 1–17. [Google Scholar] [CrossRef]
- Hashimoto, A.; Ludington, D. Ammonia desorption from concentrated chicken manure slurries. In Livestock Waste Management and Pollution Abatement; ASABE: St. Joseph, MI, USA, 1971; pp. 117–121. [Google Scholar]
- Afsahi, A.; Ahmadi-Hamedani, M.; Khodadi, M. Comparative evaluation of urinary dipstick and pH-meter for cattle urine pH measurement. Heliyon 2020, 6, e03316. [Google Scholar] [CrossRef]
- Petersen, S.O.; Andersen, A.J.; Eriksen, J. Effects of cattle slurry acidification on ammonia and methane evolution during storage. J. Environ. Qual. 2012, 41, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Fangueiro, D.; Hjorth, M.; Gioelli, F. Acidification of animal slurry–a review. J. Environ. Manag. 2015, 149, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Misselbrook, T.; Hunt, J.; Perazzolo, F.; Provolo, G. Greenhouse gas and ammonia emissions from slurry storage: Impacts of temperature and potential mitigation through covering (pig slurry) or acidification (cattle slurry). J. Environ. Qual. 2016, 45, 1520–1530. [Google Scholar] [CrossRef] [PubMed]
- Larsen, T.A.; Riechmann, M.E.; Udert, K.M. State of the art of urine treatment technologies: A critical review. Water Res. X 2021, 13, 100114. [Google Scholar] [CrossRef]
- Overmeyer, V.; Holtkamp, F.; Clemens, J.; Büscher, W.; Trimborn, M. Dynamics of different buffer systems in slurries based on time and temperature of storage and their visualization by a new mathematical tool. Animals 2020, 10, 724. [Google Scholar] [CrossRef]
- Mohammed-Nour, A.; Al-Sewailem, M.; El-Naggar, A.H. The influence of alkalization and temperature on ammonia recovery from cow manure and the chemical properties of the effluents. Sustainability 2019, 11, 2441. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | I | II | III | IV |
|---|---|---|---|---|
| Average number of cows | 120 | 120 | 70 | 70 |
| Dry matter intake in g d | 24,078 | 23,897 | 22,485 | 20,451 |
| Total gross energy intake in MJ d | 444.094 | 449.093 | 423.297 | 389.725 |
| Fraction of ash | 81 | 81 | 77 | 81 |
| Nitrogen intake in g N d | 642.9 | 681.9 | 576.0 | 527.8 |
| Milk N per growth N | 213.2 | 207.8 | 181.2 | 186.5 |
| Milk (kg Fat- and protein-corrected milk d) | 40.7 | 39.6 | 34.6 | 35.6 |
| Total excreted nitrogen in g N d | 429.7 | 474.1 | 394.8 | 341.3 |
| Total urine nitrogen in g N d | 239.5 | 282.0 | 205.4 | 171.4 |
| Total fecal nitrogen in g N d | 190.2 | 192.1 | 189.4 | 169.9 |
| Volume urine in l d | 46.69 | 46.98 | 43.94 | 40.13 |
| Mean near-surface wind speed in m s | 1.03 ± 0.54 | 1.02 ± 0.52 | 1.07 ± 0.51 | 1.03 ± 0.57 |
| Max near-surface wind speed in m s | 2.77 | 2.72 | 2.77 | 3.09 |
| Mean air temperature in C | 10.51 ± 7.13 | 10.51 ± 7.13 | 10.51 ± 7.13 | 10.51 ± 7.13 |
| Min air temperature in C | −4.30 | −4.30 | −4.30 | −4.30 |
| Max air temperature in C | 32.3 | 32.3 | 32.3 | 32.3 |
| Mean inflow in m s | 2.01 ± 1.36 | 2.01 ± 1.36 | 2.01 ± 1.36 | 2.01 ± 1.36 |
| Max inflow in m s | 8.59 | 8.59 | 8.59 | 8.59 |
| 6.5 | 8 | 0.675 | 0.825 | 4.54 | 1.03 | 8 | 8 | 7.46 |
| 6.5 | 8.5 | 0.9 | 1.1 | 1.2 | 0.76 | 8.5 | 8.5 | 7.92 |
| 6.5 | 9 | 1.125 | 1.375 | 0.72 | 0.63 | 9 | 9 | 8.39 |
| 6.5 | 9.5 | 1.35 | 1.65 | 1.2 | 0.18 | 9.23 | 9.48 | 8.6 |
| 6.5 | 10 | 1.575 | 1.925 | 0.9 | 0.16 | 9.61 | 9.96 | 8.96 |
| 6.5 | 10.5 | 1.8 | 2.2 | 0.8 | 0.11 | 9.77 | 10.34 | 9.11 |
| 6.5 | 11 | 2.025 | 2.475 | 0.57 | 0.15 | 10.44 | 10.93 | 9.74 |
| 7 | 8 | 0.45 | 0.55 | 6.33 | 0.58 | 8 | 8 | 7.46 |
| 7 | 8.5 | 0.675 | 0.825 | 0.72 | 0.57 | 8.5 | 8.5 | 7.92 |
| 7 | 9 | 0.9 | 1.1 | 0.92 | 0.16 | 8.78 | 8.98 | 8.18 |
| 7 | 9.5 | 1.125 | 1.375 | 0.54 | 0.19 | 9.29 | 9.49 | 8.66 |
| 7 | 10 | 1.35 | 1.65 | 0.52 | 0.1 | 9.39 | 9.85 | 8.75 |
| 7 | 10.5 | 1.575 | 1.925 | 0.5 | 0.1 | 9.78 | 10.33 | 9.12 |
| 7 | 11 | 1.8 | 2.2 | 0.5 | 0.1 | 10.18 | 10.8 | 9.49 |
| 7.5 | 8 | 0.225 | 0.275 | 0.69 | 0.33 | 7.99 | 8 | 7.45 |
| 7.5 | 8.5 | 0.45 | 0.55 | 0.5 | 0.1 | 8.29 | 8.45 | 7.73 |
| 7.5 | 9 | 0.675 | 0.825 | 0.5 | 0.1 | 8.69 | 8.93 | 8.1 |
| 7.5 | 9.5 | 0.9 | 1.1 | 0.5 | 0.1 | 9.09 | 9.4 | 8.47 |
| 7.5 | 10 | 1.125 | 1.375 | 0.5 | 0.1 | 9.49 | 9.88 | 8.85 |
| 7.5 | 10.5 | 1.35 | 1.65 | 0.5 | 0.1 | 9.88 | 10.35 | 9.22 |
| 7.5 | 11 | 1.575 | 1.925 | 0.5 | 0.1 | 10.28 | 10.83 | 9.59 |
| 8 | 8.5 | 0.225 | 0.275 | 0.5 | 0.1 | 8.4 | 8.48 | 7.83 |
| 8 | 9 | 0.45 | 0.55 | 0.5 | 0.1 | 8.79 | 8.95 | 8.2 |
| 8 | 9.5 | 0.675 | 0.825 | 0.5 | 0.1 | 9.19 | 9.43 | 8.57 |
| 8 | 10 | 0.9 | 1.1 | 0.5 | 0.1 | 9.59 | 9.9 | 8.94 |
| 8 | 10.5 | 1.125 | 1.375 | 0.5 | 0.1 | 9.99 | 10.38 | 9.31 |
| 8 | 11 | 1.35 | 1.65 | 0.5 | 0.1 | 10.38 | 10.85 | 9.68 |
| 8.5 | 9 | 0.225 | 0.275 | 0.5 | 0.1 | 8.9 | 8.98 | 8.3 |
| 8.5 | 9.5 | 0.45 | 0.55 | 0.5 | 0.1 | 9.29 | 9.45 | 8.67 |
| 8.5 | 10 | 0.675 | 0.825 | 0.5 | 0.1 | 9.69 | 9.93 | 9.04 |
| 8.5 | 10.5 | 0.9 | 1.1 | 0.5 | 0.1 | 10.09 | 10.4 | 9.41 |
| 8.5 | 11 | 1.125 | 1.375 | 0.5 | 0.1 | 10.49 | 10.88 | 9.78 |
| 9 | 9.5 | 0.225 | 0.275 | 0.5 | 0.1 | 9.4 | 9.48 | 8.76 |
| 9 | 10 | 0.45 | 0.55 | 0.5 | 0.1 | 9.79 | 9.95 | 9.13 |
| 9 | 10.5 | 0.675 | 0.825 | 0.5 | 0.1 | 10.19 | 10.43 | 9.5 |
| 9 | 11 | 0.9 | 1.1 | 0.5 | 0.1 | 10.59 | 10.9 | 9.87 |
| 9.5 | 10 | 0.225 | 0.275 | 0.5 | 0.1 | 9.9 | 9.98 | 9.23 |
| 9.5 | 10.5 | 0.45 | 0.55 | 0.5 | 0.1 | 10.29 | 10.45 | 9.6 |
| 9.5 | 11 | 0.675 | 0.825 | 0.5 | 0.1 | 10.69 | 10.93 | 9.97 |
| 10 | 10.5 | 0.225 | 0.275 | 0.5 | 0.1 | 10.4 | 10.48 | 9.69 |
| 10 | 11 | 0.45 | 0.55 | 0.5 | 0.1 | 10.79 | 10.95 | 10.06 |
| 10.5 | 11 | 0.225 | 0.275 | 0.5 | 0.1 | 10.9 | 10.98 | 10.16 |
| 8.0 | 8.5 | 9.0 | 9.5 | 10.0 | 10.5 | 11.0 | |
|---|---|---|---|---|---|---|---|
| 6.5 | 5/11/18 | 17/31/52 | 49/85/135 | 143/240/375 | 349/512/708 | 632/826/1029 | 939/970/949 |
| 7.0 | 5/11/18 | 17/32/53 | 49/91/151 | 144/242/376 | 299/470/693 | 616/811/1015 | 923/1004/1035 |
| 7.5 | 5/11/19 | 15/29/50 | 44/81/135 | 120/204/320 | 295/434/601 | 577/700/796 | 810/809/746 |
| 8.0 | -/-/- | 16/29/48 | 44/76/121 | 118/183/268 | 276/367/456 | 505/548/544 | 653/589/477 |
| 8.5 | -/-/- | -/-/- | 42/66/95 | 107/147/189 | 235/272/288 | 393/376/317 | 464/379/275 |
| 9.0 | -/-/- | -/-/- | -/-/- | 86/100/106 | 171/171/151 | 259/221/168 | 283/220/160 |
| 9.5 | -/-/- | -/-/- | -/-/- | -/-/- | 102/93/79 | 143/121/102 | 154/129/114 |
| 10.0 | -/-/- | -/-/- | -/-/- | -/-/- | -/-/- | 75/77/83 | 90/92/101 |
| 10.5 | -/-/- | -/-/- | -/-/- | -/-/- | -/-/- | -/-/- | 68/81/98 |
| 8.0 | 8.5 | 9.0 | 9.5 | 10.0 | 10.5 | 11.0 | |
|---|---|---|---|---|---|---|---|
| 6.5 | 11/21/37 | 33/58/95 | 85/132/194 | 128/191/271 | 208/269/337 | 241/296/353 | 306/310/305 |
| 7.0 | 11/21/37 | 33/59/96 | 59/100/158 | 141/206/287 | 162/231/314 | 244/299/356 | 293/313/325 |
| 7.5 | 11/21/36 | 22/40/67 | 49/85/136 | 102/156/227 | 177/233/295 | 245/276/297 | 271/265/246 |
| 8.0 | -/-/- | 26/46/75 | 58/93/141 | 113/159/212 | 183/216/243 | 230/231/216 | 231/203/163 |
| 8.5 | -/-/- | -/-/- | 65/94/128 | 118/145/168 | 171/176/166 | 193/169/131 | 173/136/94 |
| 9.0 | -/-/- | -/-/- | -/-/- | 108/112/105 | 138/121/92 | 136/105/69 | 110/80/52 |
| 9.5 | -/-/- | -/-/- | -/-/- | -/-/- | 90/69/47 | 79/57/39 | 61/45/34 |
| 10.5 | -/-/- | -/-/- | -/-/- | -/-/- | -/-/- | 41/34/29 | 34/30/29 |
| 10.5 | -/-/- | -/-/- | -/-/- | -/-/- | -/-/- | -/-/- | 24/25/27 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hempel, S.; Vu, H.; Amon, T.; Janke, D. The Influence of pH Dynamics on Modeled Ammonia Emission Patterns of a Naturally Ventilated Dairy Cattle Building. Atmosphere 2023, 14, 1534. https://doi.org/10.3390/atmos14101534
Hempel S, Vu H, Amon T, Janke D. The Influence of pH Dynamics on Modeled Ammonia Emission Patterns of a Naturally Ventilated Dairy Cattle Building. Atmosphere. 2023; 14(10):1534. https://doi.org/10.3390/atmos14101534
Chicago/Turabian StyleHempel, Sabrina, Huyen Vu, Thomas Amon, and David Janke. 2023. "The Influence of pH Dynamics on Modeled Ammonia Emission Patterns of a Naturally Ventilated Dairy Cattle Building" Atmosphere 14, no. 10: 1534. https://doi.org/10.3390/atmos14101534
APA StyleHempel, S., Vu, H., Amon, T., & Janke, D. (2023). The Influence of pH Dynamics on Modeled Ammonia Emission Patterns of a Naturally Ventilated Dairy Cattle Building. Atmosphere, 14(10), 1534. https://doi.org/10.3390/atmos14101534