
Effect of Selected Meteorological Variables on Full Flowering of Some Forest Herbs in the Western Carpathians


Abstract
1. Introduction
2. Material and Methods
2.1. Study Site
2.2. Characteristics of Plant Species
2.3. Phenological Observations
2.4. Method for Calculation of Temperature and Cumulative Precipitation
2.5. Statistical Analysis
3. Results
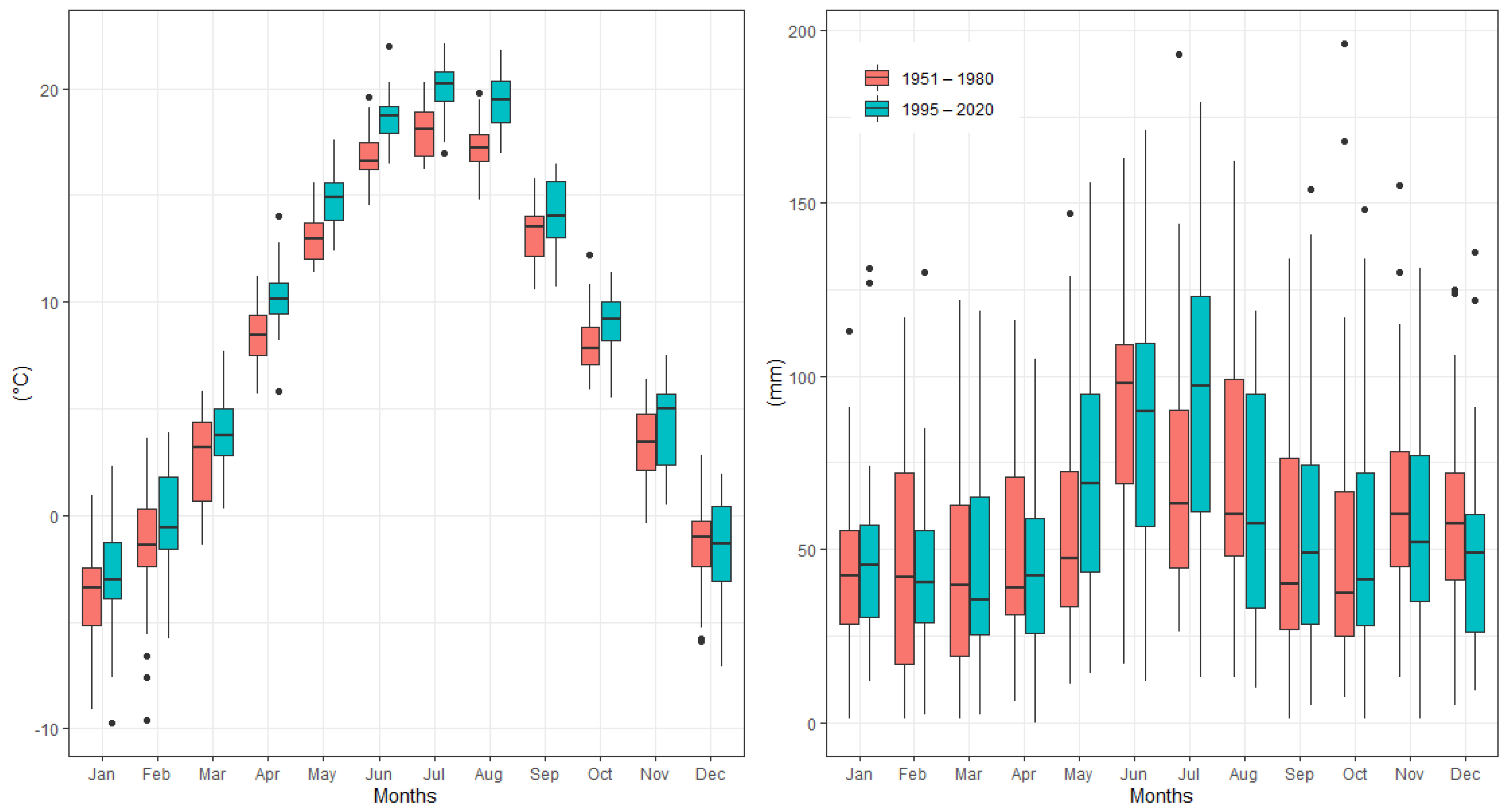
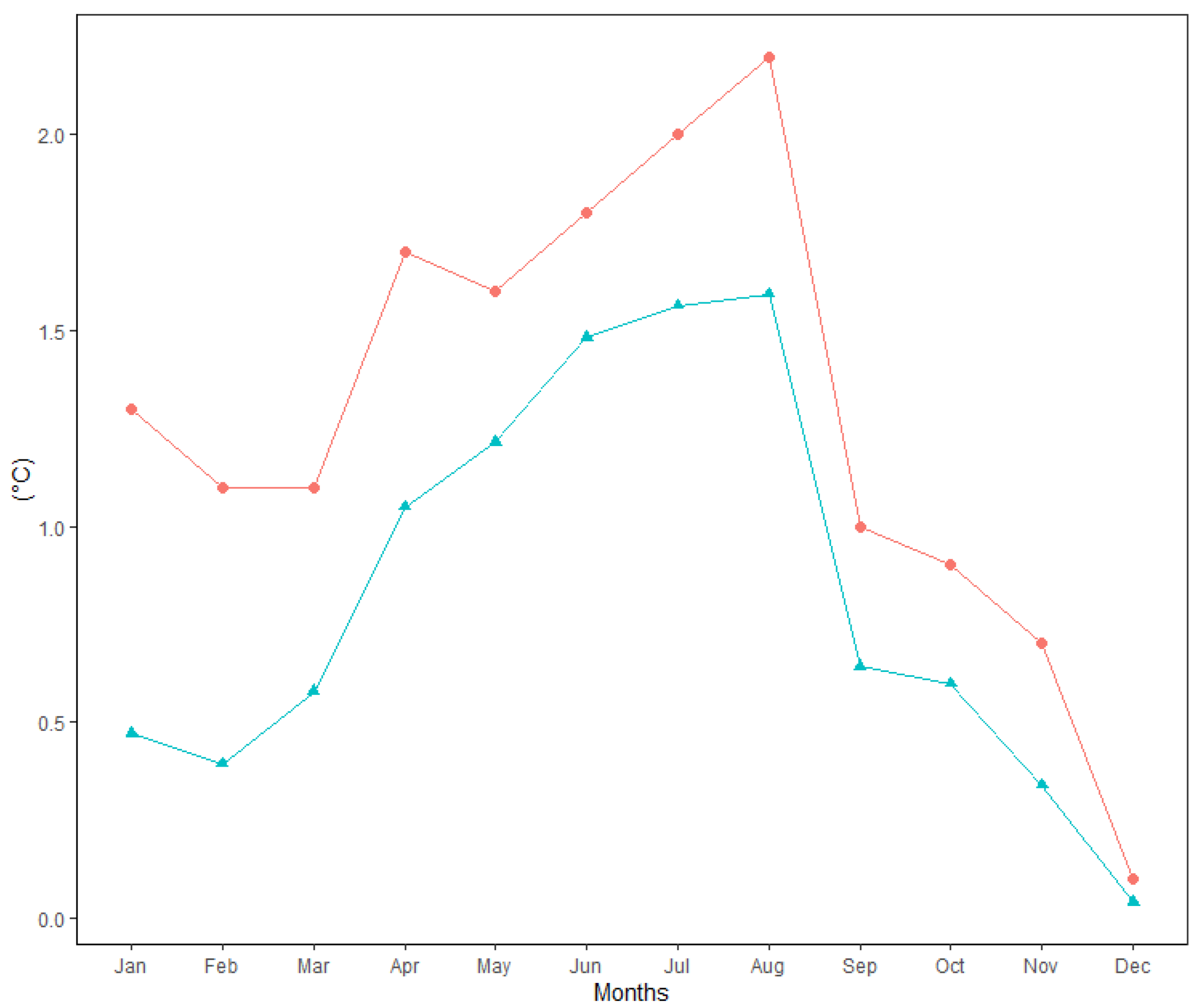
3.1. Analysis of Meteorological Variables
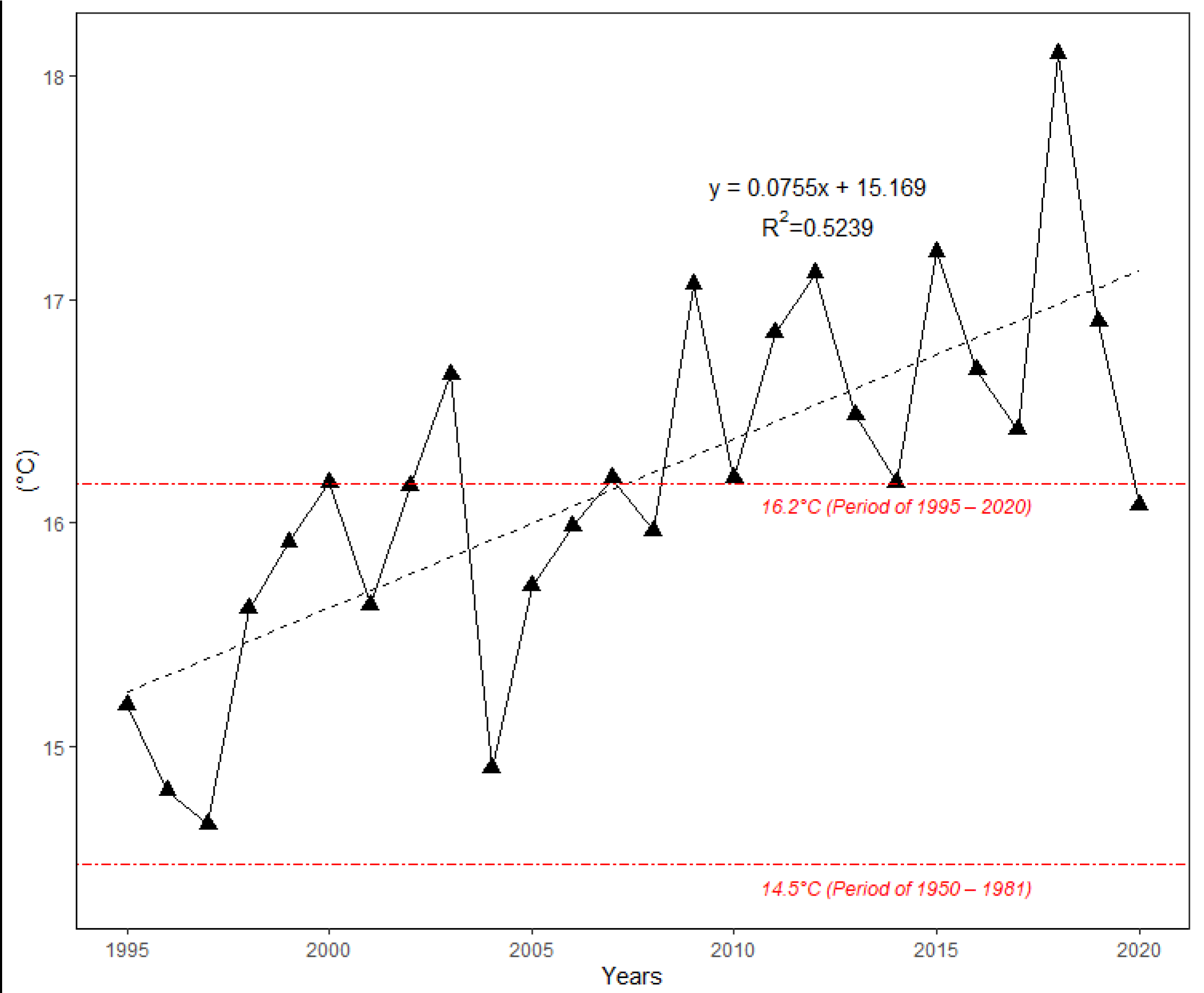
3.2. Interannual Variability Temporal Trends in the Onset of Flowering from 1995 to 2020
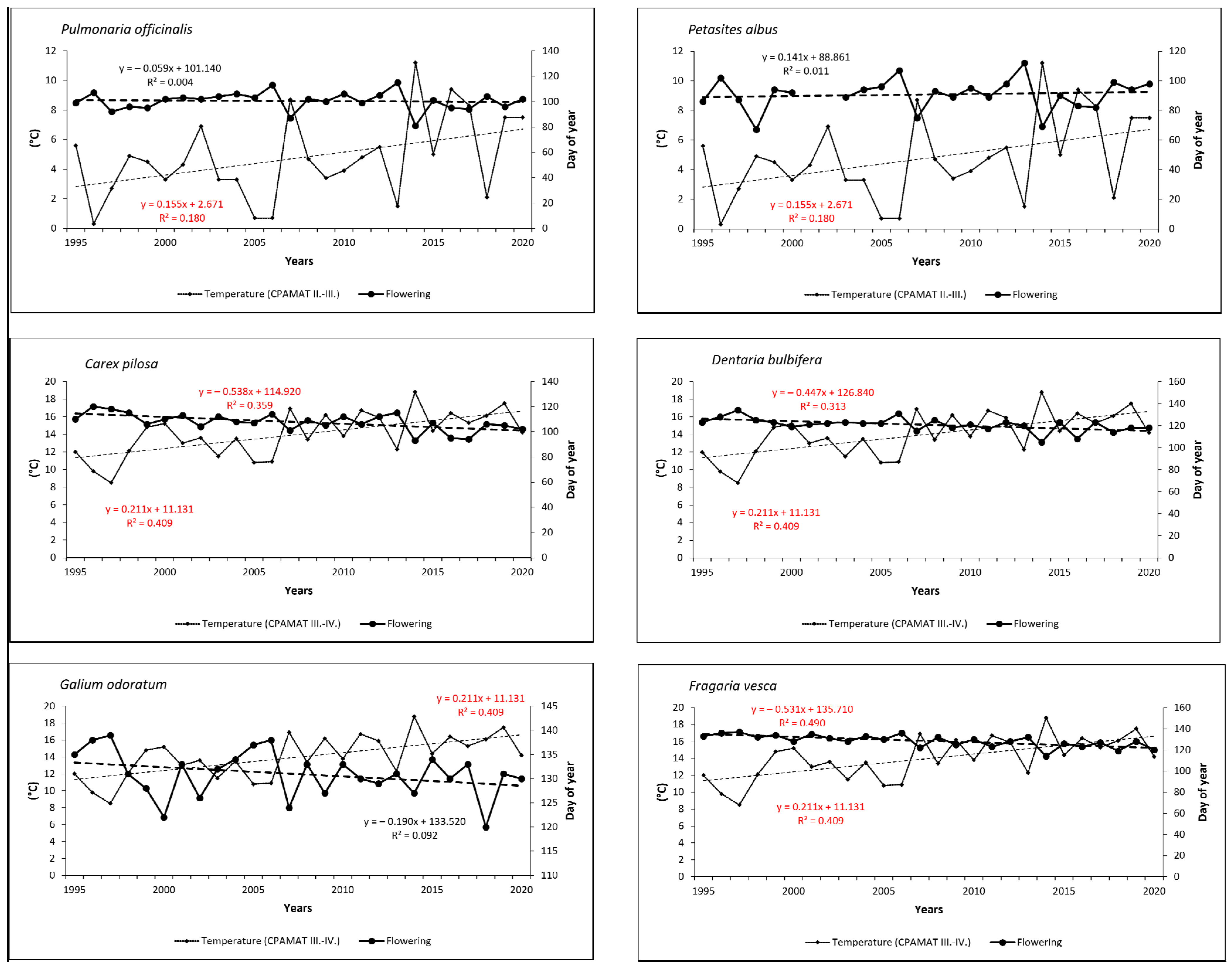
3.3. Flowering Phenology in Relation to the Monthly Sums of Air Temperature (CPAMAT)
3.4. Flowering Phenology in Relation to Monthly Cumulative Precipitation
3.5. Temperature-Sum Requirement for FF of the Species
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Amano, T.; Smithers, R.J.; Sparks, T.H.; Sutherland, W.J. A 250-year index of first flowering dates and its response to temperature changes. Proc. R. Soc. B 2010, 277, 2451–2457. [Google Scholar] [CrossRef] [PubMed]
- Boisvenue, C.; Running, S.W. Impacts of climate change on natural forest productivity—Evidence since the middle of the 20th century. Glob. Chang. Biol. 2006, 12, 862–882. [Google Scholar] [CrossRef]
- Parmesan, C.; Yohe, G. A globally coherent fingerprint of climate change impacts across natural systems. Nature 2003, 421, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Cheddadi, R.; Araújo, M.B.; Maiorano, L.; Edwards, M.; Guisan, A.; Carré, M.; Chevalier, M.; Pearman, P.B. Temperature range shifts for three European tree species over the last 10,000 years. Front. Plant Sci. 2016, 7, 1581. [Google Scholar] [CrossRef]
- Kozyr, I.V. Forest Vegetation dynamics along an altitudinal gradient in relation to the climate change in Southern Transbaikalia, Russia. Achiev. Life Sci. 2014, 8, 23–28. [Google Scholar] [CrossRef][Green Version]
- Thuiller, W.; Lavorel, S.; Araújo, M.B.; Sykes, M.T.; Prentice, I.C. Climate change threats to plant diversity in Europe. Proc. Natl. Acad. Sci. USA 2005, 102, 8245–8250. [Google Scholar] [CrossRef]
- Zimmermann, N.E.; Yoccoz, N.G.; Edwards, T.C.; Meier, E.S.; Thuiller, W.; Guisan, A.; Schmatz, D.R.; Pearman, P.B. Climatic extremes improve predictions of spatial patterns of tree species. Proc. Natl. Acad. Sci. USA 2009, 106, 19723–19728. [Google Scholar] [CrossRef]
- Menzel, A.; Estrella, N.; Fabian, P. Spatial and temporal variability of the phenological seasons in Germany from 1951 to 1996. Glob. Chang. Biol. 2001, 7, 657–666. [Google Scholar]
- Montgomery, R.A.; Rice, K.E.; Stefanski, A.; Rich, R.L.; Reich, P.B. Phenological responses of temperate and boreal trees to warming depend on ambient spring temperatures, leaf habit, and geographic range. Proc. Natl. Acad. Sci. USA 2020, 117, 10397–10405. [Google Scholar] [CrossRef]
- Schieber, B.; Kubov, M.; Janík, R. Effects of climate warming on vegetative phenology of the common beech Fagus sylvatica in a submontane forest of the Western Carpathians: Two-decade analysis. Pol. J. Ecol. 2017, 65, 339–351. [Google Scholar]
- Sierota, Z.; Grodzki, W.; Szczepkowski, A. Abiotic and biotic disturbances affecting forest health in Poland over the past 30 years: Impacts of climate and forest management. Forests 2019, 10, 75. [Google Scholar] [CrossRef]
- Lukasová, V.; Vido, J.; Škvareninová, J.; Bičárová, S.; Hlavatá, H.; Borsányi, P.; Škvarenina, J. Autumn phenological response of European Beech to summer drought and heat. Water 2020, 12, 2610. [Google Scholar] [CrossRef]
- Lukasová, V.; Škvareninová, J.; Bičárová, S.; Sitárová, Z.; Hlavatá, H.; Borsányi, P.; Škvarenina, J. Regional and altitudinal aspects in summer heatwave intensification in the Western Carpathians. Theor. Appl. Climatol. 2021, 146, 1111–1125. [Google Scholar] [CrossRef]
- Kirschbaum, M.U.F. Forest growth and species distribution in a changing climate. Tree Physiol. 2000, 20, 309–322. [Google Scholar] [CrossRef] [PubMed]
- Hartl-Meier, C.; Dittmar, C.; Zang, C.; Rothe, A. Mountain forest growth response to climate change in the Northern Limestone Alps. Trees 2014, 28, 819–829. [Google Scholar] [CrossRef]
- Ďurský, J.; Škvarenina, J.; Minďáš, J.; Miková, A. Regional analysis of climate change impact on Norway spruce (Picea abies L. Karst.) growth in Slovak mountain forests. J. Forest Sci. 2006, 52, 306–315. [Google Scholar] [CrossRef]
- Rafferty, N.E.; Diez, J.M.; Bertelsen, C.D. Changing climate drives divergent and nonlinear shifts in flowering phenology across elevations. Curr. Biol. 2020, 30, 432–441. [Google Scholar] [CrossRef]
- Bottlíková, A. Phenological characterization of selected phytocoenoses in the kettlehole of Liptov. Biologické Práce 1975, 21, 1–81. [Google Scholar]
- Davies, T.J.; Wolkovich, E.M.; Kraft, N.J.B.; Salamin, N.; Allen, J.M.; Ault, T.R.; Betancourt, J.L.; Bolmgren, K.; Cleland, E.E.; Cook, B.I.; et al. Phylogenetic conservatism in plant phenology. J. Ecol. 2013, 101, 1520–1530. [Google Scholar] [CrossRef]
- Tyler, G. Relationships between climate and flowering of eight herbs in a Swedish deciduous forest. Ann. Bot. 2001, 87, 623–630. [Google Scholar] [CrossRef][Green Version]
- Fitter, A.H.; Fitter, R.S.R. Rapid changes in flowering time in British plants. Science 2002, 296, 1689–1691. [Google Scholar] [CrossRef] [PubMed]
- Schieber, B. Changes of flowering phenology of six herbal species in a beech forest (Central Slovakia): A decade analysis. Pol. J. Ecol. 2007, 55, 233–244. [Google Scholar]
- Liu, H.; Dai, J.; Liu, J. Spatiotemporal variation in full-flowering dates of tree peonies in the middle and lower reaches of China’s Yellow River: A simulation through the panel data model. Sustainability 2017, 9, 1343. [Google Scholar] [CrossRef]
- Lee, H.K.; Lee, S.J.; Kim, M.K.; Lee, S.D. Prediction of plant phenological shift under climate change in South Korea. Sustainability 2020, 12, 9276. [Google Scholar] [CrossRef]
- Jiang, Y.; Li, B.; Yuan, Y.; Sun, Q.; Zhang, T.; Liu, Y.; Li, Y.; Li, R.; Li, F. Trends in flowering phenology of herbaceous plants and its response to precipitation and snow cover on the Qinghai-Tibetan Plateau from 1983 to 2017. Sustainability 2021, 13, 7640. [Google Scholar] [CrossRef]
- Khodorova, N.V.; Boitel-Conti, M. The role of temperature in the growth and flowering of geophytes. Plants 2013, 2, 699–711. [Google Scholar] [CrossRef]
- Szabó, B.; Vincze, E.; Czúcz, B. Flowering phenological changes in relation to climate change in Hungary. Int. J. Biometeorol. 2016, 60, 1347–1356. [Google Scholar] [CrossRef]
- Nam, B.E.; Kim, J.G. Flowering season of vernal herbs is shortened at elevated temperatures with reduced precipitation in early spring. Sci. Rep. 2020, 10, 17494. [Google Scholar] [CrossRef]
- Hájková, L.; Voženílek, V.; Tolasz, R.; Kohut, M.; Možný, M.; Nekovář, J.; Novák, M.; Reitschläger, J.D.; Richterová, D.; Stříž, M.; et al. Atlas of the Phenological Conditions in Czechia; CHMI Prague-UP Olomouc: Prague, Czech Republic, 2012; p. 312. [Google Scholar]
- Hájková, L.; Možný, M.; Oušková, V.; Bartošová, L.; Dížková, P.; Žalud, Z. Meteorological variables that affect the beginning of flowering of the winter oilseed rape in the Czech Republic. Atmosphere 2021, 12, 1444. [Google Scholar] [CrossRef]
- Craine, J.M.; Wolkovich, E.M.; Towne, E.G. The roles of shifting and filtering in generating community-level flowering phenology. Ecography 2012, 35, 1033–1038. [Google Scholar] [CrossRef]
- Song, Y.; Shim, J.; Kinmonth-Schultz, H.; Imaizumi, T. Photoperiodic flowering: Time measurement mechanisms in leaves. Annu. Rev. Plant Biol. 2015, 66, 441–464. [Google Scholar] [CrossRef] [PubMed]
- Mortensen, L.M. Effects of air humidity on growth, flowering, keeping quality and water relations of four short-day greenhouse species. Sci. Hortic. 2000, 86, 299–310. [Google Scholar] [CrossRef]
- Chauhan, Y.S.; Ryan, M.; Chandra, S.; Sadras, V.O. Accounting for soil moisture improves prediction of flowering time in chickpea and wheat. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- McMaster, G.S.; Wilhelm, W.W. Is soil temperature better than air temperature for predicting winter wheat phenology? Agron. J. 1998, 90, 602–607. [Google Scholar] [CrossRef]
- Fitter, H.; Fitter, R.S.R.; Harris, I.T.B.; Williamson, M.H. Relationships between first flowering date and temperature in the flora o a locality in central england. Funct. Ecol. 1995, 9, 55–60. [Google Scholar] [CrossRef]
- Mo, F.; Zhang, J.; Wang, J.; Cheng, Z.G.; Sun, G.J.; Ren, H.X.; Zhao, X.Z.; Cheruiyot, W.K.; Kavagi, L.; Wang, J.Y.; et al. Phenological evidence from China to address rapid shifts in global flowering times with recent climate change. Agric. For. Meteorol. 2017, 246, 22–30. [Google Scholar] [CrossRef]
- eLTER Networks. Available online: https://elter-ri.eu/national-lter-networks (accessed on 10 December 2021).
- Križová, E. Primary production of the aboveground biomass of a herb layer in selected forest types in EES Kováčová. Acta Fac. For. 1993, 35, 99–107. [Google Scholar]
- Kubov, M.; Schieber, B.; Janík, R. Seasonal dynamics of macronutrients in aboveground biomass of two herb-layer species in a beech forest. Biologia 2019, 74, 1415–1424. [Google Scholar] [CrossRef]
- Lapin, M.; Faško, P.; Melo, M.; Šťastný, P.; Tomlain, J. Atlas of the Landscape of the Slovak Republic, 1st ed.; MŽP: Bratislava, Slovakia, 2002; p. 344. [Google Scholar]
- Střelec, J. Influence of cutting intervention in a beech forest stand on changes in illumination. Lesnícky časopis-For. J. 1992, 38, 551–558. [Google Scholar]
- Diekmann, M. Relationship between flowering phenology of perennial herbs and meteorological data in deciduous forests of Sweden. Can. J. Bot. 1996, 74, 528–537. [Google Scholar] [CrossRef]
- Braslavská, O.; Kamenský, L. Phenological Observation of Forest Plants; Methodical Prescription; SHMI: Bratislava, Slovakia, 1996; p. 22. [Google Scholar]
- Meier, U. BBCH-Monograph: Growth Stages of Mono- and Dicotyledonous Plants, 2nd ed.; Technical Report; Federal Biological Research Centre for Agriculture and Forestry: Bonn, Germany, 2001. [Google Scholar]
- Braslavská, O.; Borsányi, P. Quality control of long series of phenological data with sum of cumulated average monthly air temperatures. In International Symposium on Applied Agrometeorology and Agroclimatology: Proceedings, Volos, Greece, 24–26 April 1996; Dalezios, N.R., Ed.; CEE: Luxembourg, 1996; pp. 305–310. [Google Scholar]
- Murray, M.B.; Cannell, G.R.; Smith, R.I. Date of budburst of fifteen tree species in Britain following climatic warming. J. Appl. Ecol. 1989, 26, 693–700. [Google Scholar] [CrossRef]
- Heide, O.M. Daylength and thermal time responses of budburst during dormancy release in some northern deciduous trees. Physiol. Plant. 1993, 88, 531–540. [Google Scholar] [CrossRef] [PubMed]
- Langvall, O.; Löfvenius, M.O. Long-term standardized forest phenology in Sweden: A climate change indicator. Int. J. Biometeorol. 2021, 65, 381–391. [Google Scholar] [CrossRef] [PubMed]
- R Core Team R. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017; Available online: https://www.project.org/ (accessed on 10 December 2021).
- Evans, J.D. Straightforward Statistics for the Behavioral Sciences; Brooks/Cole Publishing: Pacific Grove, CA, USA, 1996. [Google Scholar]
- WMO Report: 2020 Closes a Decade of Exceptional Heat. Available online: https://public.wmo.int/en/media/news/2020-closes-decade-of-exceptional-heat (accessed on 10 December 2021).
- IPCC 6th Report: Climate Change Widespread, Rapid, and Intensifying. Available online: https://www.ipcc.ch/2021/08/09/ar6-wg1-20210809-pr/ (accessed on 11 December 2021).
- Shorina, N.I.; Smirnova, O.V. The population biology of ephemeroids. In The Population Structure of Vegetation; White, J., Ed.; Dr. V. Junk Publishers: Dordrecht, The Netherlands, 1995; pp. 225–240. [Google Scholar]
- Lapointe, L.; Lerat, S. Annual growth of the spring ephemeral Erythronium americanum as a function of temperature and mycorrhizal status. Can. J. Bot. 2006, 84, 39–48. [Google Scholar] [CrossRef]
- Miller-Rushing, A.J.; Inouye, D.W.; Primack, R.B. How well do first flowering dates measure plant responses to climate change? The effects of population size and sampling frequency. J. Ecol. 2008, 96, 1289–1296. [Google Scholar] [CrossRef]
- Adhikari, B.S.; Kumar, R. Effect of snowmelt regime on phenology of herbaceous species at and around treeline in Western Himalaya, India. Not. Sci. Biol. 2020, 12, 901–919. [Google Scholar] [CrossRef]
- Willems, F.M.; Scheepens, J.F.; Ammer, C.; Block, S.; Bucharova, A.; Schall, P.; Sehrt, M.; Bossdorf, O. Spring understory herbs flower later in intensively managed forests. Ecol. Appl. 2021, 31, e02332. [Google Scholar] [CrossRef]
- Richardson, A.D.; O’Keefe, J. Phenological differences between understory and overstory: A case study using the long-term Harvard forest records. In Phenology and Ecosystem Processes; Noormets, A., Ed.; Springer Science + Business Media, LLC: New York, NY, USA, 2009; pp. 87–117. [Google Scholar]
- Chmielewski, F.M.; Müller, A.; Bruns, E. Climate changes and trends in phenology of fruit trees and field crops in Germany, 1961–2000. Agric. For. Meteorol. 2004, 121, 69–78. [Google Scholar] [CrossRef]
- Babálová, D.; Škvareninová, J.; Fazekaš, J.; Vyskot, I. The dynamics of the phenological development of four woody species in south-west and central Slovakia. Sustainability 2018, 10, 1497. [Google Scholar] [CrossRef]
- Gordo, O.; Sanz, J.J. Phenology and climate change: A long-term study in a Mediterranean locality. Oecologia 2005, 146, 484–495. [Google Scholar] [CrossRef]
- Moore, L.M.; Lauenroth, W.K. Differential effects of temperature and precipitation on early- vs. late- flowering species. Ecosphere 2017, 8, e01819. [Google Scholar] [CrossRef]
- Hájková, L.; Hubálek, Z.; Kožnarová, V.; Bartošová, L.; Možný, M. Flowering of allergenically important plant species in relation to the North Atlantic Oscillation system and thermal time in the Czech Republic. Aerobiologia 2018, 34, 157–169. [Google Scholar] [CrossRef]
- Von Wuehlisch, G.; Krusche, D.; Muhs, H.J. Variation in temperature sum requirement for flushing of beech provenances. Silvae Genet. 1995, 44, 343–346. [Google Scholar]
- Schemske, D.W.; Willson, M.F.; Melampy, M.N.; Miller, L.J.; Verner, L.; Schemske, K.M.; Best, L.B. Flowering ecology of some spring woodland herbs. Ecology 1978, 59, 351–366. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phenological Groups | Species | Earliest Onset DOY (year) | Latest Onset DOY (year) | Mean (DOY) | STDE (Days) | VR (Days) | CV (%) |
|---|---|---|---|---|---|---|---|
| Early spring | Petasites albus | 67 (1998) | 112 (2013) | 90.8 | 10.6 | 45 | 11.6 |
| Pulmonaria officinalis | 81 (2014) | 115 (2013) | 100.4 | 7.3 | 34 | 7.2 | |
| Mid spring | Carex pilosa | 93 (2014) | 120 (1996) | 107.7 | 6.9 | 27 | 6.4 |
| Dentaria bulbifera | 105 (2014) | 134 (1997) | 120.8 | 6.1 | 29 | 5.1 | |
| Late spring | Fragaria vesca | 114 (2014) | 137 (1997) | 128.5 | 5.8 | 23 | 4.5 |
| Galium odoratum | 120 (2018) | 139 (1997) | 131.0 | 4.8 | 19 | 3.7 | |
| Early summer | Veronica officinalis | 146 (2018) | 175 (1997) | 159.7 | 6.3 | 29 | 3.9 |
| Mid summer | Mycelis muralis | 171 (2018) | 191 (1997) | 182.7 | 4.9 | 20 | 2.7 |
| Late summer | Campanula trachelium | 194 (2015) | 222 (2004) | 203.4 | 6.5 | 28 | 3.2 |
| Phenological Group | Species | Period | Temperature (r Value) | Precipitation (r Value) |
|---|---|---|---|---|
| Early spring | Petasites albus | II.–III. | −0.658 *** | 0.207 |
| III. | −0.542 ** | 0.204 | ||
| Pulmonaria officinalis | II.–III. | −0.745 *** | 0.080 | |
| III. | −0.682 *** | 0.164 | ||
| Mid spring | Carex pilosa | III.–IV. | −0.751 *** | 0.255 |
| IV. | −0.289 | 0.309 | ||
| Dentaria bulbifera | III.–IV. | −0.816 *** | 0.114 | |
| IV. | −0.617 *** | 0.357 | ||
| Late spring | Galium odoratum | III.–IV. | −0.719 *** | 0.087 |
| IV. | −0.663 *** | 0.443 * | ||
| Fragaria vesca | III.–IV. | −0.780 *** | 0.322 | |
| IV. | −0.550 ** | 0.484 * | ||
| Early summer | Veronica officinalis | IV.–V. | −0.672 *** | 0.108 |
| V. | −0.430 * | 0.076 | ||
| Mid summer | Mycelis muralis | IV.–VI. | −0.771 *** | 0.357 |
| V.–VI. | −0.735 *** | 0.288 | ||
| VI. | −0.597 ** | 0.280 | ||
| Late summer | Campanula trachelium | IV.–VI. | −0.488 * | 0.515 ** |
| IV.–VII. | −0.612 *** | 0.268 | ||
| V.–VII. | −0.619 *** | 0.121 | ||
| VII. | −0.492 * | 0.347 |
| Phenological Groups | Species | Cumulative Daily Temperature Over 0 °C | ||||
|---|---|---|---|---|---|---|
| Average | Min | Max | CV (%) | STD | ||
| Early spring | Petasites albus | 223.4 | 177.9 | 287.7 | 16.9 | 37.8 |
| Pulmonaria officinalis | 294.2 | 268.1 | 325.4 | 6.1 | 17.8 | |
| Mid spring | Carex pilosa | 337.6 | 278.2 | 402.8 | 10.1 | 34.1 |
| Dentaria bulbifera | 483.4 | 362.6 | 559.9 | 10.2 | 49.4 | |
| Late spring | Fragaria vesca | 572.0 | 510.7 | 655.4 | 6.8 | 38.6 |
| Galium odoratum | 632.3 | 536.1 | 740.9 | 9.2 | 58.0 | |
| Early summer | Veronica officinalis | 996.3 | 907.4 | 1152.6 | 6.6 | 65.3 |
| Mid summer | Mycelis muralis | 1438.0 | 1333.1 | 1571.8 | 4.1 | 58.5 |
| Late summer | Campanula trachelium | 1820.8 | 1662.6 | 1992.2 | 5.2 | 94.4 |
| Phenological Groups | Species | Cumulative Daily Temperature Over 5 °C | ||||
|---|---|---|---|---|---|---|
| Average | Min | Max | CV (%) | STD | ||
| Early spring | Petasites albus | 142.9 | 65.5 | 227.3 | 33.0 | 47.2 |
| Pulmonaria officinalis | 210.6 | 135.8 | 257.5 | 18.6 | 39.1 | |
| Mid spring | Carex pilosa | 252.4 | 178.5 | 329.4 | 19.7 | 49.7 |
| Dentaria bulbifera | 395.5 | 269.5 | 486.5 | 14.5 | 57.5 | |
| Late spring | Fragaria vesca | 484.1 | 413.8 | 581.9 | 10.6 | 51.5 |
| Galium odoratum | 544.4 | 451.7 | 619.1 | 9.4 | 51.2 | |
| Early summer | Veronica officinalis | 908.4 | 814.4 | 1079.1 | 8.7 | 79.2 |
| Mid summer | Mycelis muralis | 1350.1 | 1240.1 | 1450.0 | 4.7 | 64.0 |
| Late summer | Campanula trachelium | 1732.9 | 1530.3 | 1898.5 | 6.1 | 106.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kubov, M.; Schieber, B.; Janík, R. Effect of Selected Meteorological Variables on Full Flowering of Some Forest Herbs in the Western Carpathians. Atmosphere 2022, 13, 195. https://doi.org/10.3390/atmos13020195
Kubov M, Schieber B, Janík R. Effect of Selected Meteorological Variables on Full Flowering of Some Forest Herbs in the Western Carpathians. Atmosphere. 2022; 13(2):195. https://doi.org/10.3390/atmos13020195
Chicago/Turabian StyleKubov, Martin, Branislav Schieber, and Rastislav Janík. 2022. "Effect of Selected Meteorological Variables on Full Flowering of Some Forest Herbs in the Western Carpathians" Atmosphere 13, no. 2: 195. https://doi.org/10.3390/atmos13020195
APA StyleKubov, M., Schieber, B., & Janík, R. (2022). Effect of Selected Meteorological Variables on Full Flowering of Some Forest Herbs in the Western Carpathians. Atmosphere, 13(2), 195. https://doi.org/10.3390/atmos13020195