
Net Ecosystem CO2 Exchange in Mountain Grasslands Is Seriously Endangered by the Temperature Increase in the Eastern Pyrenees


Abstract
1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Eddy Covariance Measurements
2.3. Data Analysis
3. Results
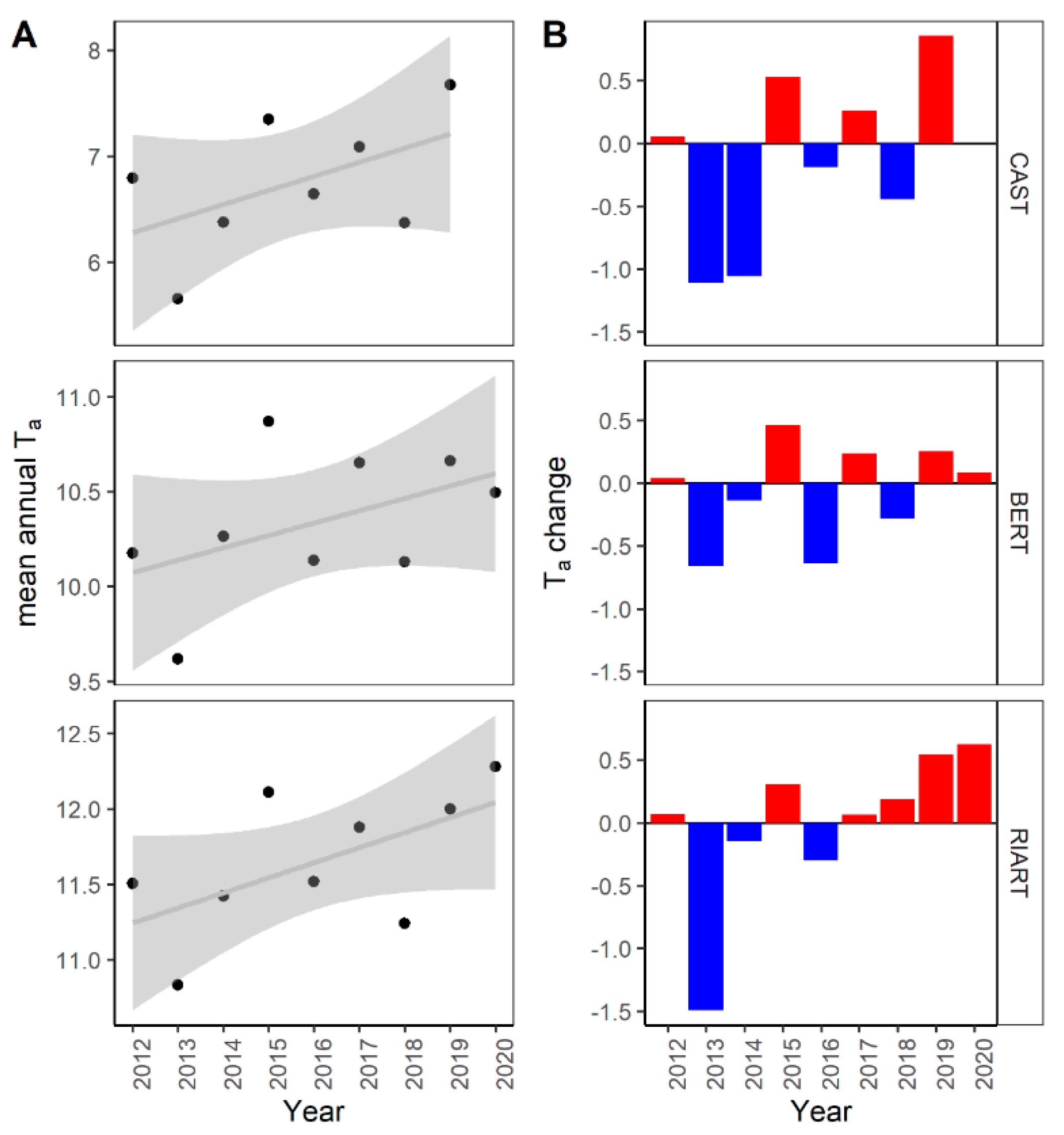
3.1. CO2 Flux Inter-Annual Dynamics
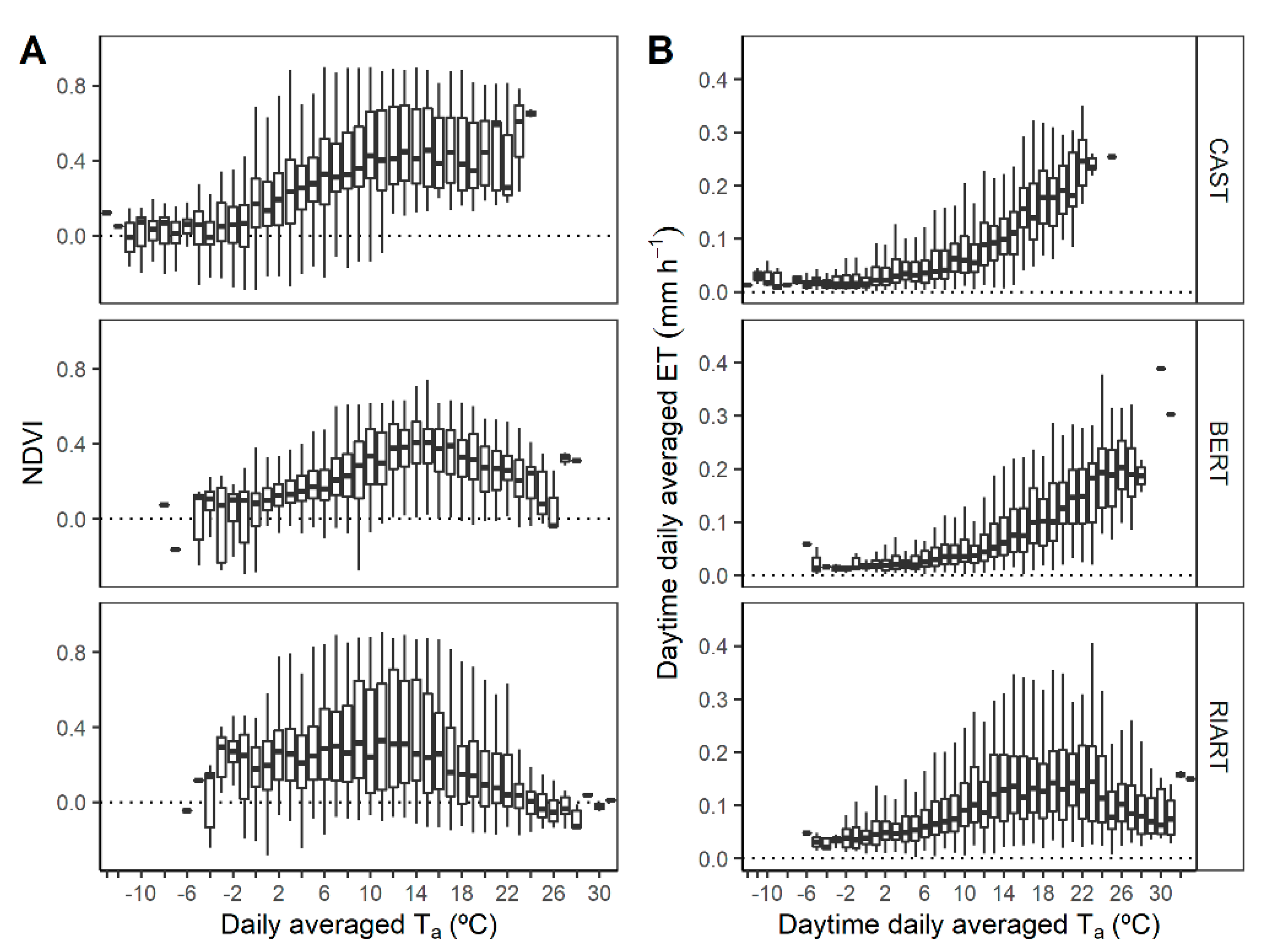
3.2. Intra-Annual CO2 Fluxes and Environmental and Phenological Dynamics
3.3. Environmental and Phenological Drivers of CO2 Fluxes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gerber, P.J.; Steinfeld, H.; Henderson, B.; Mottet, A.; Opio, C.; Dijkman, J.; Falcucci, A.; Tempio, G. Tackling Climate Change through Livestock: A Global Assessment of Emissions and Mitigation Opportunities; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2013; ISBN 9789251079201. [Google Scholar]
- Berninger, F.; Susiluoto, S.; Gianelle, D.; Bahn, M.; Wohlfahrt, G.; Sutton, M.; Garcia-Pausas, J.; Gimeno, C.; Sanz, M.J.; Dore, S.; et al. Management and Site Effects on Carbon Balances of European Mountain Meadows and Rangelands. Boreal Environ. Res. 2015, 20, 748–760. [Google Scholar]
- Hörtnagl, L.; Barthel, M.; Buchmann, N.; Eugster, W.; Butterbach-Bahl, K.; Díaz-Pinés, E.; Zeeman, M.; Klumpp, K.; Kiese, R.; Bahn, M.; et al. Greenhouse Gas Fluxes over Managed Grasslands in Central Europe. Glob. Chang. Biol. 2018, 24, 1843–1872. [Google Scholar] [CrossRef]
- FAO. Challenges and Opportunities for Carbon Sequestration in Grassland Systems. A Technical Report on Grassland Management and Climate Change Mitigation; Conant, R.T., Ed.; Plant Production and Protection Division Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2010; Volume 9, ISBN 9789251064948. [Google Scholar]
- OPCC-CTP. Climate Change in the Pyrenees: Impacts, Vulnerabilities and Adaptation. Bases of Knowledge for the Future Climate Change Adaptation Strategy in the Pyrenees; OPCC-CTP, Ed.; OPCC-CTP: Huesca, Spain, 2018; ISBN 978-84-09-06268-3. [Google Scholar]
- Smit, H.J.; Metzger, M.J.; Ewert, F. Spatial Distribution of Grassland Productivity and Land Use in Europe. Agric. Syst. 2008, 98, 208–219. [Google Scholar] [CrossRef]
- Porqueddu, C.; Ates, S.; Louhaichi, M.; Kyriazopoulos, A.P.; Moreno, G.; del Pozo, A.; Ovalle, C.; Ewing, M.A.; Nichols, P.G.H. Grasslands in “Old World” and “New World” Mediterranean-Climate Zones: Past Trends, Current Status and Future Research Priorities. Grass Forage Sci. 2016, 71, 1–35. [Google Scholar] [CrossRef]
- Rodríguez, A.; Canals, R.M.; Plaixats, J.; Albanell, E.; Debouk, H.; Garcia-Pausas, J.; San Emeterio, L.; Jimenez, J.J.; Sebastià, M.-T. Interactions between Biogeochemical and Management Factors Explain Soil Organic Carbon in Pyrenean Grasslands. Biogeosciences 2020, 17, 6033–6050. [Google Scholar] [CrossRef]
- Nogueira, C.; Werner, C.; Rodrigues, A.; Caldeira, M.C. A Prolonged Dry Season and Nitrogen Deposition Interactively Affect CO 2 Fluxes in an Annual Mediterranean Grassland. Sci. Total Environ. 2019, 654, 978–986. [Google Scholar] [CrossRef]
- Schaufler, G.; Kitzler, B.; Schindlbacher, A.; Skiba, U.; Sutton, M.A.; Zechmeister-Boltenstern, S. Greenhouse Gas Emissions from European Soils under Different Land Use: Effects of Soil Moisture and Temperature. Eur. J. Soil Sci. 2010, 61, 683–696. [Google Scholar] [CrossRef]
- Imer, D.; Merbold, L.; Eugster, W.; Buchmann, N. Temporal and Spatial Variations of Soil CO2, CH4 and N2O Fluxes at Three Differently Managed Grasslands. Biogeosciences 2013, 10, 5931–5945. [Google Scholar] [CrossRef]
- Ceschia, E.; Béziat, P.; Dejoux, J.F.; Aubinet, M.; Bernhofer, C.; Bodson, B.; Buchmann, N.; Carrara, A.; Cellier, P.; Di Tommasi, P.; et al. Management Effects on Net Ecosystem Carbon and GHG Budgets at European Crop Sites. Agric. Ecosyst. Environ. 2010, 139, 363–383. [Google Scholar] [CrossRef]
- Kutsch, W.L.; Aubinet, M.; Buchmann, N.; Smith, P.; Osborne, B.; Eugster, W.; Wattenbach, M.; Schrumpf, M.; Schulze, E.D.; Tomelleri, E.; et al. The Net Biome Production of Full Crop Rotations in Europe. Agric. Ecosyst. Environ. 2010, 139, 336–345. [Google Scholar] [CrossRef]
- Vuichard, N.; Ciais, P.; Viovy, N.; Li, L.; Ceschia, E.; Wattenbach, M.; Bernhofer, C.; Emmel, C.; Grünwald, T.; Jans, W.; et al. Simulating the Net Ecosystem CO2 Exchange and Its Components over Winter Wheat Cultivation Sites across a Large Climate Gradient in Europe Using the ORCHIDEE-STICS Generic Model. Agric. Ecosyst. Environ. 2016, 226, 1–17. [Google Scholar] [CrossRef]
- Xu, L.; Baldocchi, D.D. Seasonal Variation in Carbon Dioxide Exchange over a Mediterranean Annual Grassland in California. Agric. For. Meteorol. 2004, 1232, 79–96. [Google Scholar] [CrossRef]
- Jongen, M.; Pereira, J.S.; Aires, L.M.I.; Pio, C.A. The Effects of Drought and Timing of Precipitation on the Inter-Annual Variation in Ecosystem-Atmosphere Exchange in a Mediterranean Grassland. Agric. For. Meteorol. 2011, 151, 595–606. [Google Scholar] [CrossRef]
- Grant, R.F.; Baldocchi, D.D.; Ma, S. Ecological Controls on Net Ecosystem Productivity of a Seasonally Dry Annual Grassland under Current and Future Climates: Modelling with Ecosys. Agric. For. Meteorol. 2012, 152, 189–200. [Google Scholar] [CrossRef]
- Serrano-Ortiz, P.; Oyonarte, C.; Pérez-Priego, O.; Reverter, B.R.; Sánchez-Cañete, E.P.; Were, A.; Uclés, O.; Morillas, L.; Domingo, F. Ecological Functioning in Grass-Shrub Mediterranean Ecosystems Measured by Eddy Covariance. Oecologia 2014, 175, 1005–1017. [Google Scholar] [CrossRef]
- Ma, S.; Baldocchi, D.; Wolf, S.; Verfaillie, J. Slow Ecosystem Responses Conditionally Regulate Annual Carbon Balance over 15 Years in Californian Oak-Grass Savanna. Agric. For. Meteorol. 2016, 228–229, 252–264. [Google Scholar] [CrossRef]
- Pereira, J.S.; Mateus, J.A.; Aires, L.M.; Pita, G.; Pio, C.; David, J.S.; Andrade, V.; Banza, J.; David, T.S.; Paço, T.A.; et al. Net Ecosystem Carbon Exchange in Three Contrasting Mediterranean Ecosystems - The Effect of Drought. Biogeosciences 2007, 4, 791–802. [Google Scholar] [CrossRef]
- Rogger, J.; Hörtnagl, L.; Buchmann, N.; Eugster, W. Carbon Dioxide Fluxes of a Mountain Grassland: Drivers, Anomalies and Annual Budgets. Agric. For. Meteorol. 2022, 314, 108801. [Google Scholar] [CrossRef]
- Ibañez, M.; Altimir, N.; Ribas, À.; Eugster, W.; Sebastià, M.-T. Cereal-Legume Mixtures Increase Net CO2 Uptake in a Forage Crop System in the Eastern Pyrenees. F. Crop. Res. 2021, 272, 108262. [Google Scholar] [CrossRef]
- Ibañez, M.; Altimir, N.; Ribas, À.; Eugster, W.; Sebastià, M.-T. Phenology and Plant Functional Type Dominance Drive CO2 Exchange in Seminatural Grasslands in the Pyrenees. J. Agric. Sci. 2020, 158, 3–14. [Google Scholar] [CrossRef]
- Soil Survey Staff—NRCS/USDA. Keys to Soil Taxonomy, 12th ed.; United States Department of Agriculture: Washington, WA, USA, 2014; ISBN 0926487221. [Google Scholar]
- Moncrieff, J.; Clement, R.; Finnigan, J.; Meyers, T. Averaging, Detrending, and Filtering of Eddy Covariance Time Series. In Handbook of Micrometeorology. Atmospheric and Oceanographic Sciences Library; Lee, X., Massman, W., Law, B., Eds.; Springer: Dordrecht, The Netherlands, 2004; Volume 29. [Google Scholar]
- Moncrieff, J.B.; Massheder, J.M.; de Bruin, H.; Elbers, J.; Friborg, T.; Heusinkveld, B.; Kabat, P.; Scott, S.; Soegaard, H.; Verhoef, A. A System to Measure Surface Fluxes of Momentum, Sensible Heat, Water Vapour and Carbon Dioxide. J. Hydrol. 1997, 188–189, 589–611. [Google Scholar] [CrossRef]
- Webb, E.K.; Pearman, G.I.; Leuning, R. Correction of Flux Measurements for Density Effects Due to Heat and Water Vapour Transfer. Q. J. R. Meteorol. Soc. 1980, 106, 85–100. [Google Scholar] [CrossRef]
- Foken, T.; Gockede, M.; Mauder, M.; Mahrt, L.; Amiro, B.D.; Munger, J.W. Post-Field Quality Control. In Handbook of Micrometeorology. Atmospheric and Oceanographic Sciences Library; Lee, X., Massman, W., Law, B., Eds.; Springer: Dordrecht, The Netherlands, 2004; Volume 29, pp. 81–108. [Google Scholar]
- Papale, D. Data Gap Filling. In Eddy Covariance: A Practical Guide to Measurement and Data Analysis; Aubinet, M., Vesala, T., Papale, D., Eds.; Springer Dordrecht: Berlin/Heidelberg, Germany, 2012; pp. 159–172. ISBN 978-94-007-2350-4. [Google Scholar]
- Emmel, C.; Winkler, A.; Hörtnagl, L.; Revill, A.; Ammann, C.; D’Odorico, P.; Buchmann, N.; Eugster, W. Integrated Management of a Swiss Cropland Is Not Sufficient to Preserve Its Soil Carbon Pool in the Long Term. Biogeosciences 2018, 15, 5377–5393. [Google Scholar] [CrossRef]
- Aubinet, M.; Feigenwinter, C.; Heinesch, B.; Laffineur, Q.; Papale, D.; Reichstein, M.; Rinne, J.; Van Gorsel, E. Nighttime Flux Correction. In Eddy Covariance: A Practical Guide to Measurement and Data Analysis; Aubinet, M., Vesala, T., Papale, D., Eds.; Springer Dordrecht: Berlin/Heidelberg, Germany, 2012; pp. 133–157. ISBN 978-94-007-2351-1. [Google Scholar]
- Reichstein, M.; Falge, E.; Baldocchi, D.; Papale, D.; Aubinet, M.; Berbigier, P.; Bernhofer, C.; Buchmann, N.; Gilmanov, T.; Granier, A.; et al. On the Separation of Net Ecosystem Exchange into Assimilation and Ecosystem Respiration: Review and Improved Algorithm. Glob. Chang. Biol. 2005, 11, 1424–1439. [Google Scholar] [CrossRef]
- Muggeo, V.M.R. Segmented: An R Package to Fit Regression Models with Broken-Line Relationships. R News 2008, 8, 20–25. [Google Scholar]
- Baldocchi, D.; Chu, H.; Reichstein, M. Inter-Annual Variability of Net and Gross Ecosystem Carbon Fluxes: A Review. Agric. For. Meteorol. 2018, 249, 520–533. [Google Scholar] [CrossRef]
- Gilmanov, T.G.; Soussana, J.F.; Aires, L.; Allard, V.; Ammann, C.; Balzarolo, M.; Barcza, Z.; Bernhofer, C.; Campbell, C.L.; Cernusca, A.; et al. Partitioning European Grassland Net Ecosystem CO2 Exchange into Gross Primary Productivity and Ecosystem Respiration Using Light Response Function Analysis. Agric. Ecosyst. Environ. 2007, 121, 93–120. [Google Scholar] [CrossRef]
- Sebastià, M.-T. Plant Guilds Drive Biomass Response to Global Warming and Water Availability in Subalpine Grassland. J. Appl. Ecol. 2007, 44, 158–167. [Google Scholar] [CrossRef]
- García-González, R. Management of Natura 2000 Habitats. Alpine and Subalpine Calcareous Grasslands 6170; The European Commission: Brussels, Belgium, 2008; ISBN 9789279083266. [Google Scholar]
- Lin, D.; Xia, J.; Wan, S. Climate Warming and Biomass Accumulation of Terrestrial Plants: A Meta-Analysis. New Phytol. 2010, 188, 187–198. [Google Scholar] [CrossRef]
- Li, C.; Peng, F.; Xue, X.; You, Q.; Lai, C.; Zhang, W.; Cheng, Y. Productivity and Quality of Alpine Grassland Vary with Soil Water Availability under Experimental Warming. Front. Plant Sci. 2018, 9, 1790. [Google Scholar] [CrossRef]
- Leitinger, G.; Ruggenthaler, R.; Hammerle, A.; Lavorel, S.; Schirpke, U.; Clement, J.-C.; Lamarque, P.; Obojes, N.; Tappeiner, U. Impact of Droughts on Water Provision in Managed Alpine Grasslands in Two Climatically Different Regions of the Alps. Ecohydrology 2015, 8, 1600–1613. [Google Scholar] [CrossRef]
- Brilli, F.; Hörtnagl, L.; Hammerle, A.; Haslwanter, A.; Hansel, A.; Loreto, F.; Wohlfahrt, G. Leaf and Ecosystem Response to Soil Water Availability in Mountain Grasslands. Agric. For. Meteorol. 2011, 151, 1731–1740. [Google Scholar] [CrossRef]
- Yan, Y.; Wang, J.; Tian, D.; Luo, Y.; Xue, X.; Peng, F.; He, J.S.; Liu, L.; Jiang, L.; Wang, X.; et al. Sustained Increases in Soil Respiration Accompany Increased Carbon Input under Long-Term Warming across Global Grasslands. Geoderma 2022, 428, 116157. [Google Scholar] [CrossRef]
- Tiwari, P.; Bhattacharya, P.; Rawat, G.S.; Rai, I.D.; Talukdar, G. Experimental Warming Increases Ecosystem Respiration by Increasing Above-Ground Respiration in Alpine Meadows of Western Himalaya. Sci. Rep. 2021, 11, 2640. [Google Scholar] [CrossRef]
- Nyberg, M.; Hovenden, M.J. Warming Increases Soil Respiration in a Carbon-Rich Soil without Changing Microbial Respiratory Potential. Biogeosciences 2020, 17, 4405–4420. [Google Scholar] [CrossRef]
- Debouk, H.; De Bello, F.; Sebastia, M.T. Functional Trait Changes, Productivity Shifts and Vegetation Stability in Mountain Grasslands during a Short-Term Warming. PLoS One 2015, 10, 1–17. [Google Scholar] [CrossRef]
- Rodríguez, A.; Canals, R.M.; Sebastià, M.-T. Positive Effects of Legumes on Soil Organic Carbon Storage Disappear at High Legume Proportion across a Wide Range of Environmental Conditions in Grasslands in the Pyrenees. Ecosystems 2020, 25, 960–975. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Coordinates | Altitude (m a. s. l.) | MAT (°C) | MAP (mm) | Soil Type | Management Regime |
|---|---|---|---|---|---|---|
| Pla de Riart (RIART) | 42°03′48″ N, 1°30′48″ E | 1003 | 11.7 ± 7.4 | 664 ± 187 | Petrocalcic calcixerept | Mixed crop-livestock system. Crop rotation of cereal monocultures and cereal-legume mixtures. Cattle grazing (0.91 LSU ha−1) during the fallow period |
| La Bertolina (BERT) | 42°05′56” N, 1°39′40” E | 1275 | 10.2 ± 6.8 | 686 ± 193 | Udic calciustept | Cattle grazing (0.44 LSU ha−1), from early June to November |
| Castellar de n’Hug (CAST) | 42°18′18” N, 2°02′01” E | 2000 | 6.9 ± 6.8 | 944 ± 338 | Lithic udorthent | Cattle and sheep grazing (0.74 LSU ha−1), from late June to November |
| Site | Change Point | Slope 1 | Slope 2 | R2Adj | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Estimate | SE | Estimate | SE | p-Value | Estimate | SE | p-Value | |||
| NEE~Ta | RIART | 10.3 | 0.9 | −0.21 | 0.05 | *** | 0.23 | 0.03 | *** | 0.15 |
| BERT | 11.3 | 0.7 | −0.13 | 0.02 | *** | 0.16 | 0.03 | *** | 0.14 | |
| CAST | 16 | 2 | −0.08 | 0.02 | *** | −0.6 | 0.4 | *** | 0.09 | |
| NDVI~Ta | RIART | 11.4 | 0.8 | 0.020 | 0.005 | *** | −0.030 | 0.004 | *** | 0.15 |
| BERT | 12.6 | 0.6 | 0.030 | 0.003 | *** | −0.016 | 0.004 | *** | 0.33 | |
| CAST | 10 | 2 | 0.035 | 0.004 | *** | 0.011 | 0.009 | ns | 0.37 | |
| ET~Ta | RIART | 18.0 | 0.6 | 0.0062 | 0.0005 | *** | −0.011 | 0.003 | *** | 0.26 |
| BERT | 10 | 1 | 0.004 | 0.001 | ** | 0.0098 | 0.0009 | *** | 0.52 | |
| CAST | 9 | 1 | 0.0048 | 0.0006 | *** | 0.011 | 0.001 | *** | 0.58 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ibañez, M.; Sebastià, M.T. Net Ecosystem CO2 Exchange in Mountain Grasslands Is Seriously Endangered by the Temperature Increase in the Eastern Pyrenees. Atmosphere 2022, 13, 1980. https://doi.org/10.3390/atmos13121980
Ibañez M, Sebastià MT. Net Ecosystem CO2 Exchange in Mountain Grasslands Is Seriously Endangered by the Temperature Increase in the Eastern Pyrenees. Atmosphere. 2022; 13(12):1980. https://doi.org/10.3390/atmos13121980
Chicago/Turabian StyleIbañez, Mercedes, and Maria Teresa Sebastià. 2022. "Net Ecosystem CO2 Exchange in Mountain Grasslands Is Seriously Endangered by the Temperature Increase in the Eastern Pyrenees" Atmosphere 13, no. 12: 1980. https://doi.org/10.3390/atmos13121980
APA StyleIbañez, M., & Sebastià, M. T. (2022). Net Ecosystem CO2 Exchange in Mountain Grasslands Is Seriously Endangered by the Temperature Increase in the Eastern Pyrenees. Atmosphere, 13(12), 1980. https://doi.org/10.3390/atmos13121980