
Which Is More Sensitive to Water Stress for Irrigation Scheduling during the Maturation Stage: Grapevine Photosynthesis or Berry Size?

 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant and Soil Materials
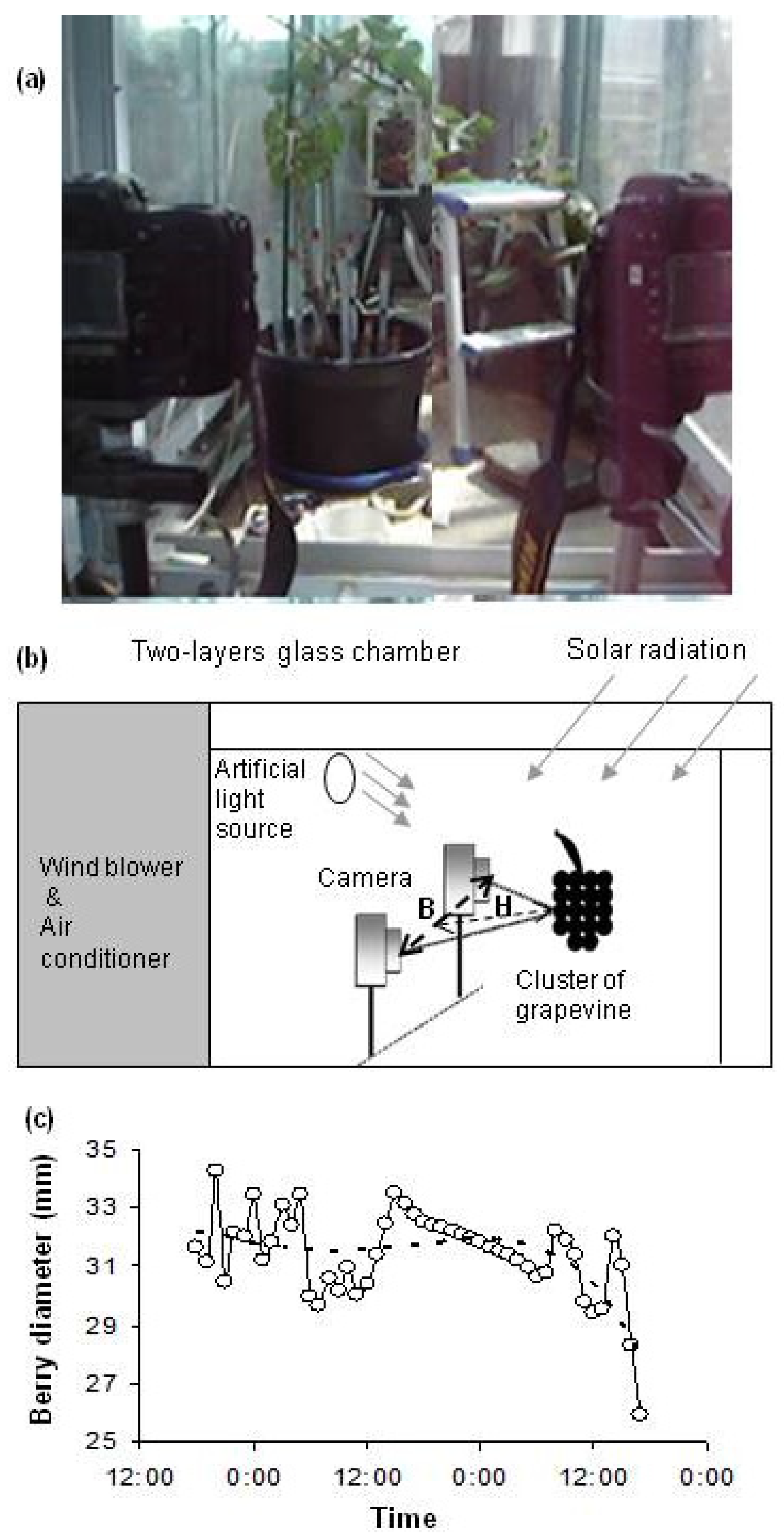
2.2. Experimental Design
2.3. Time Series Analysis of Berry Diameter Data
2.4. Statistical Analysis
3. Results and Discussion
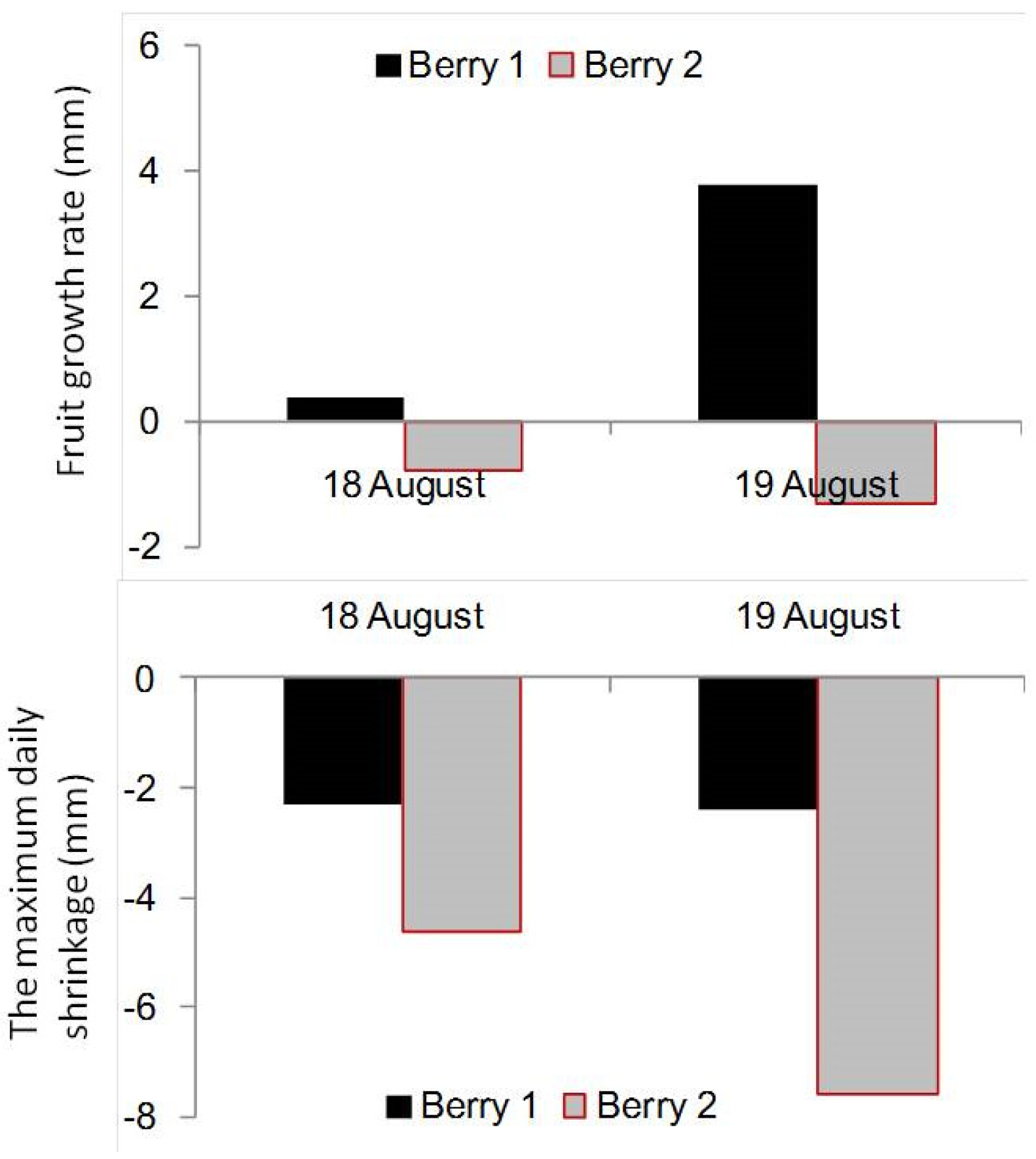
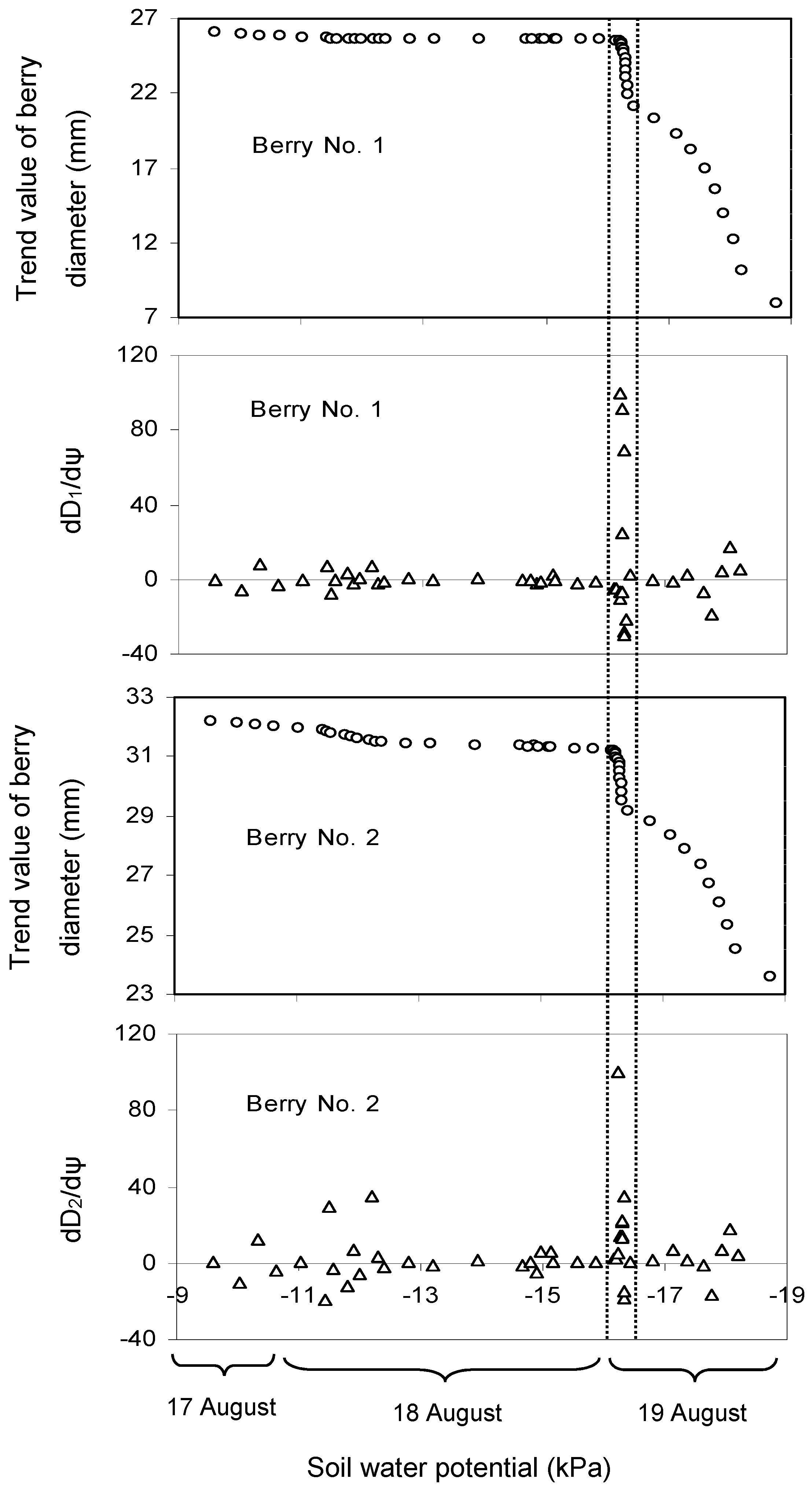
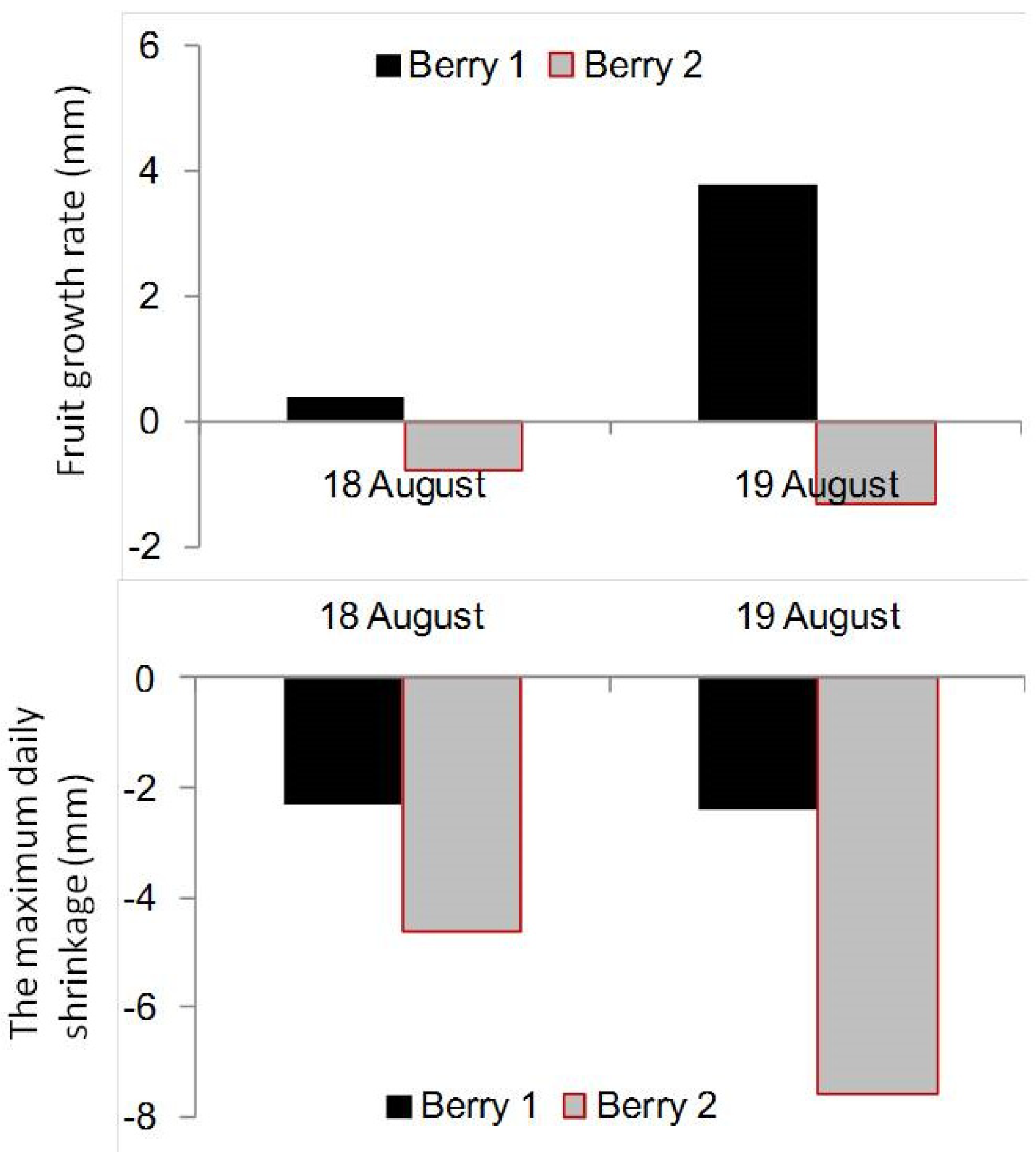
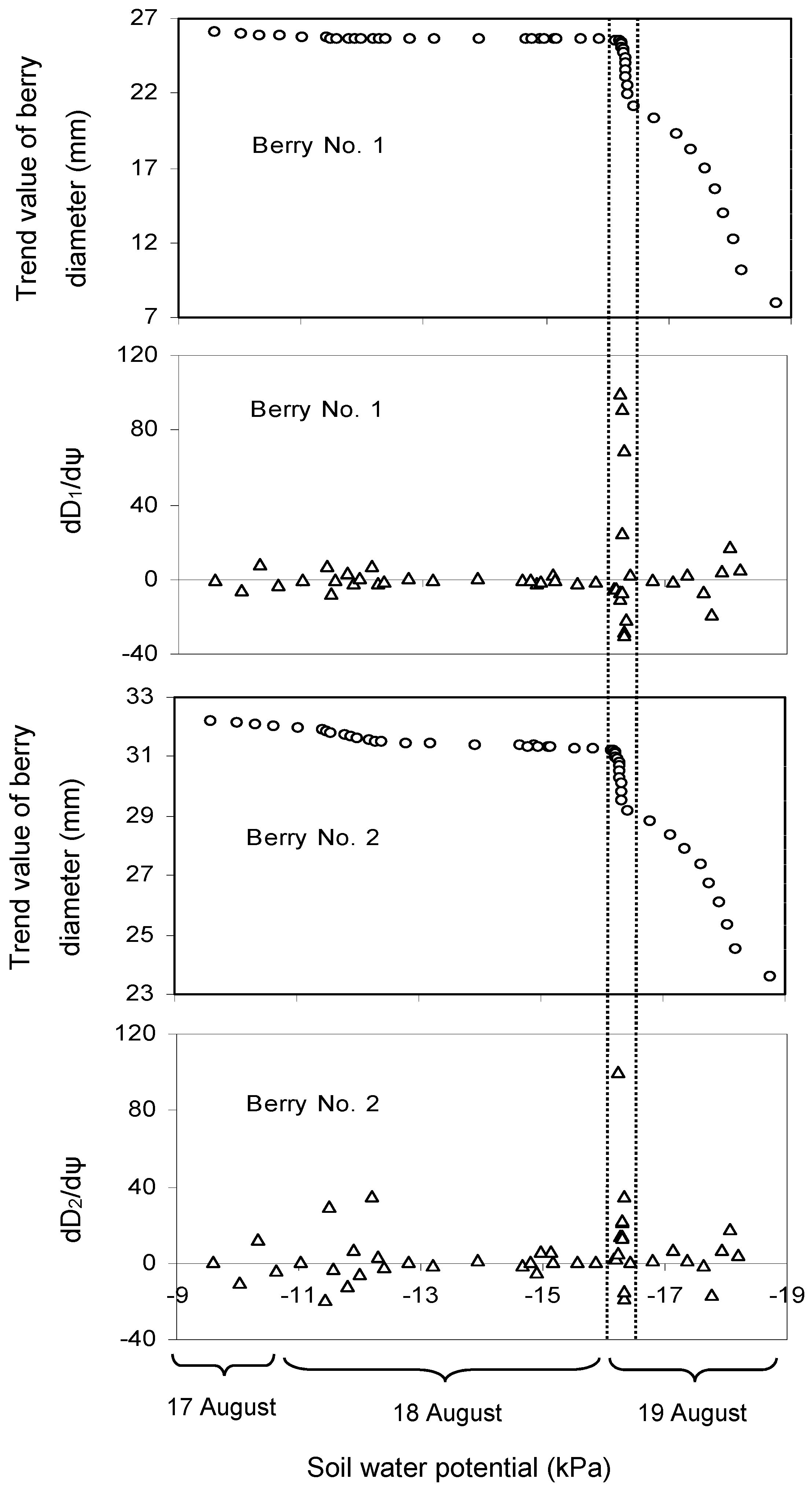
3.1. Effect of Soil Water Potential (ψ) on the Trend Value of Berry Diameter
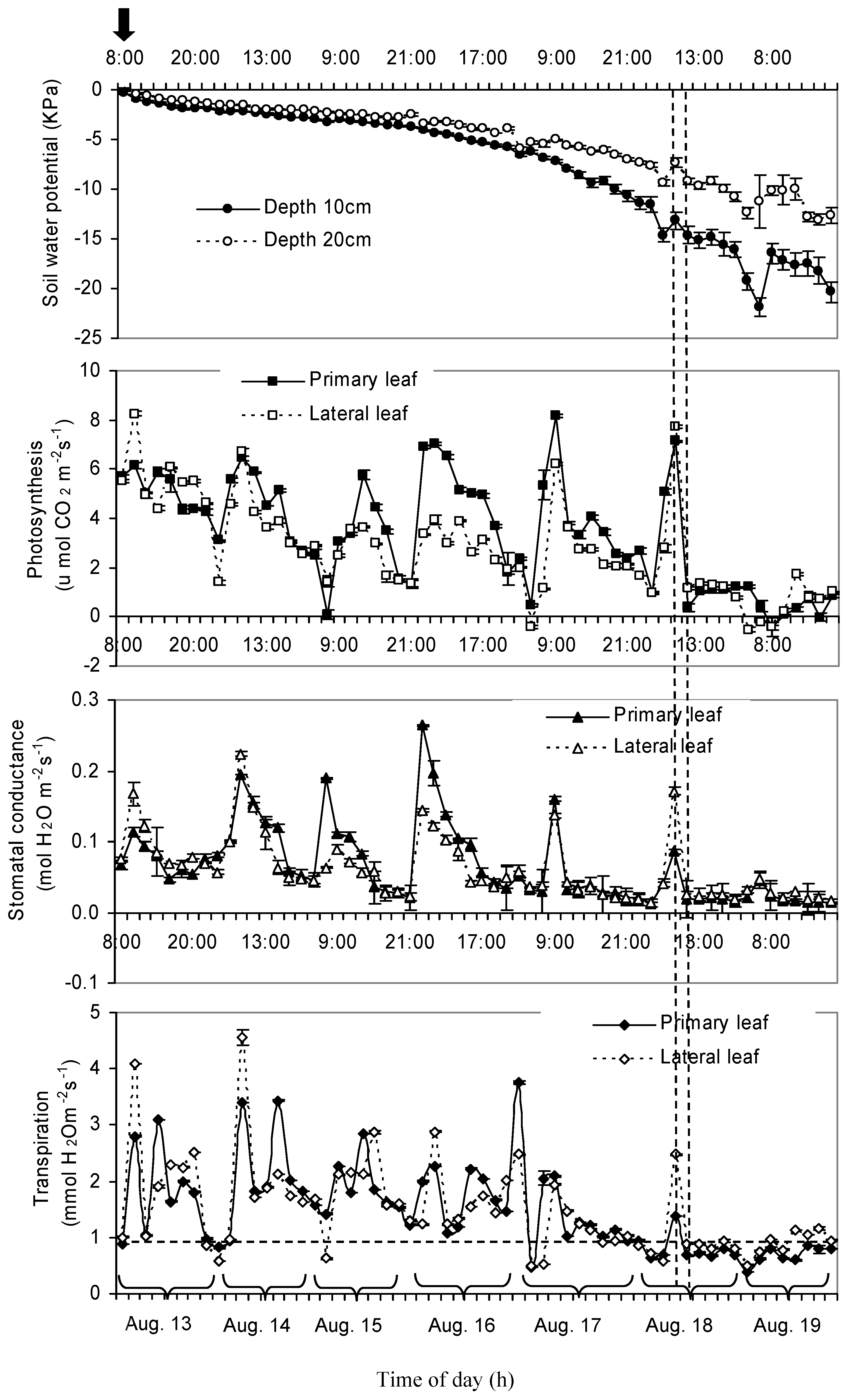
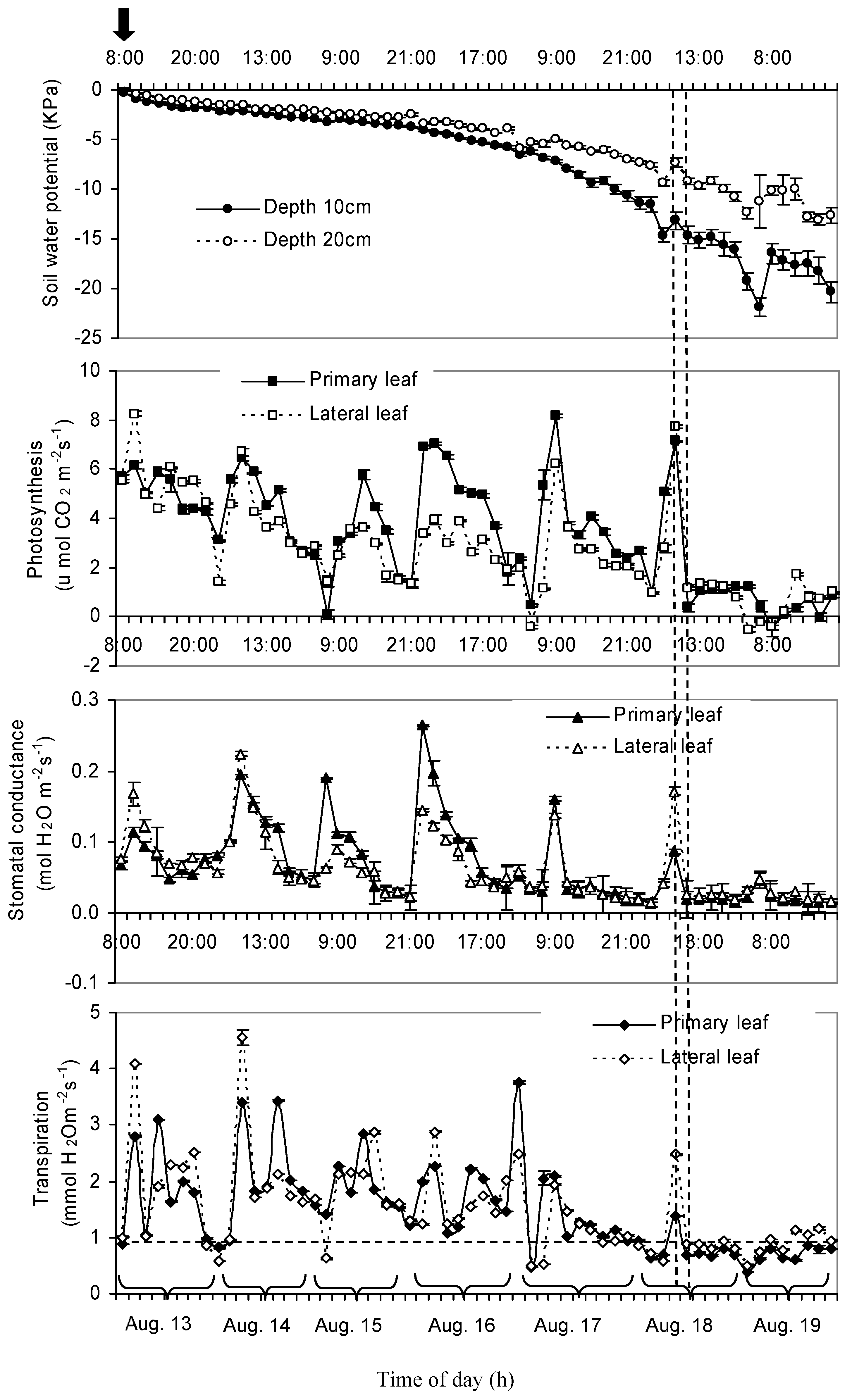
3.2. Daily Variation of Soil Water Potential, Stomatal Conductance, Net Photosynthesis, and Transpiration
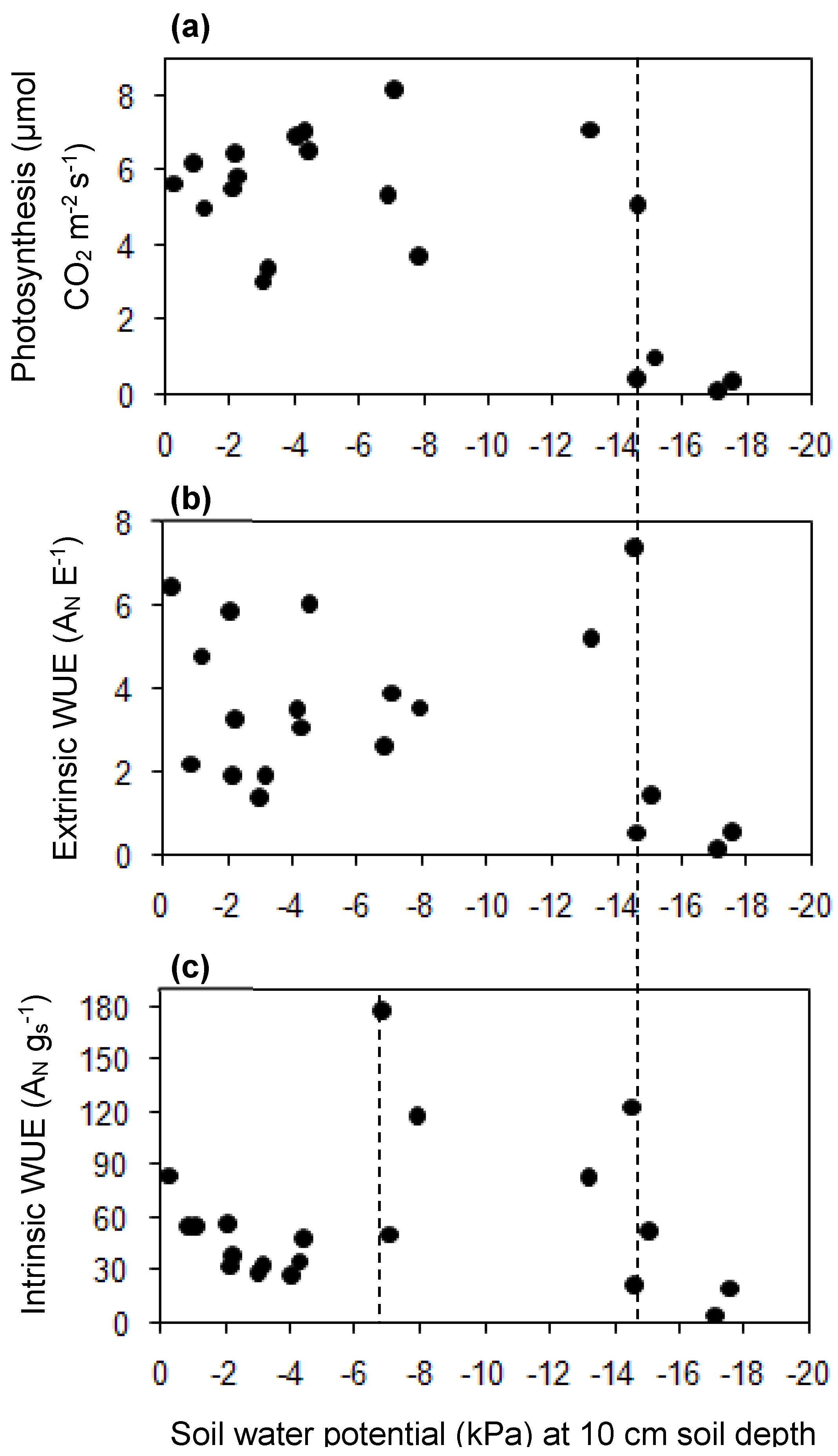
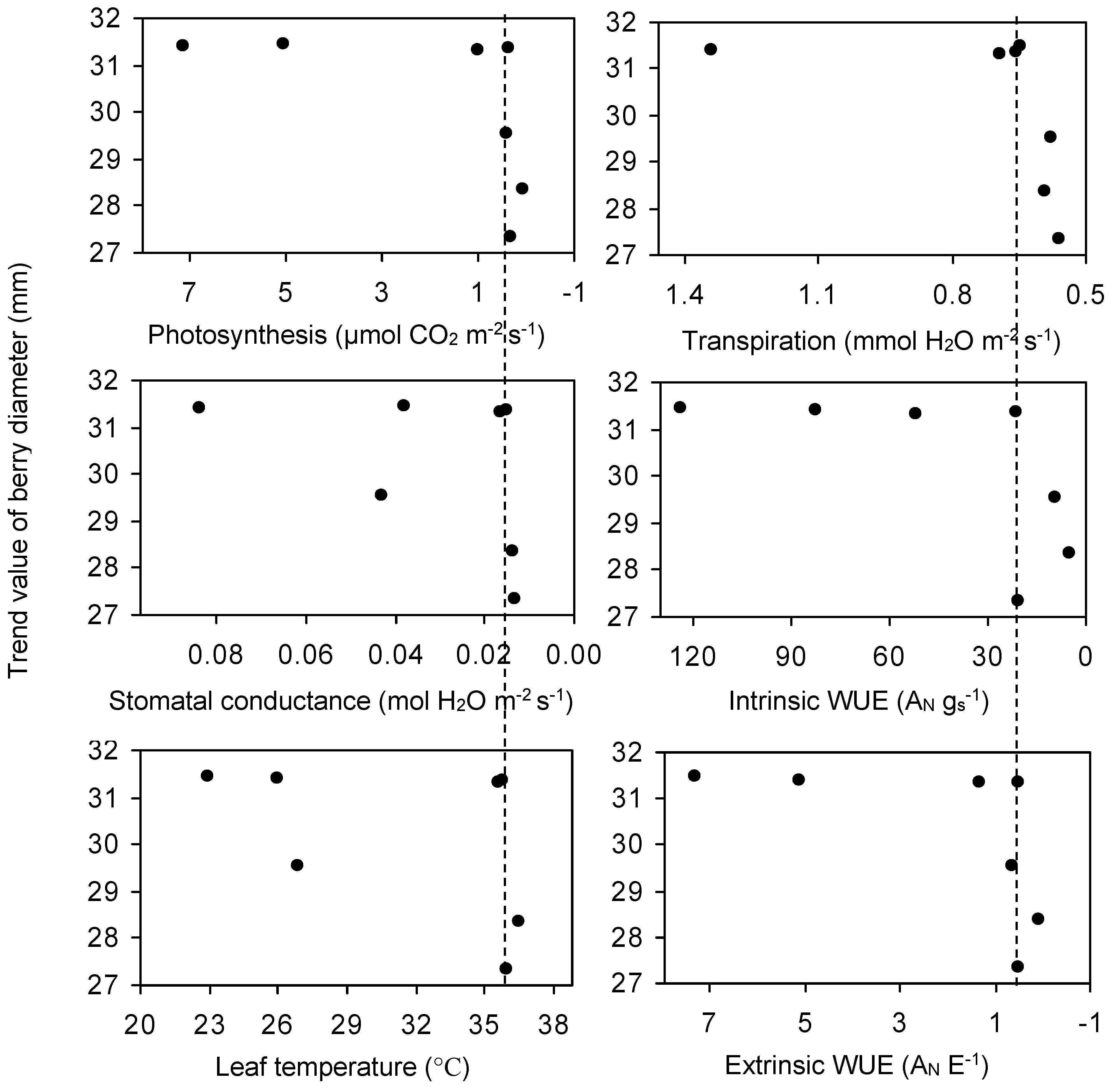
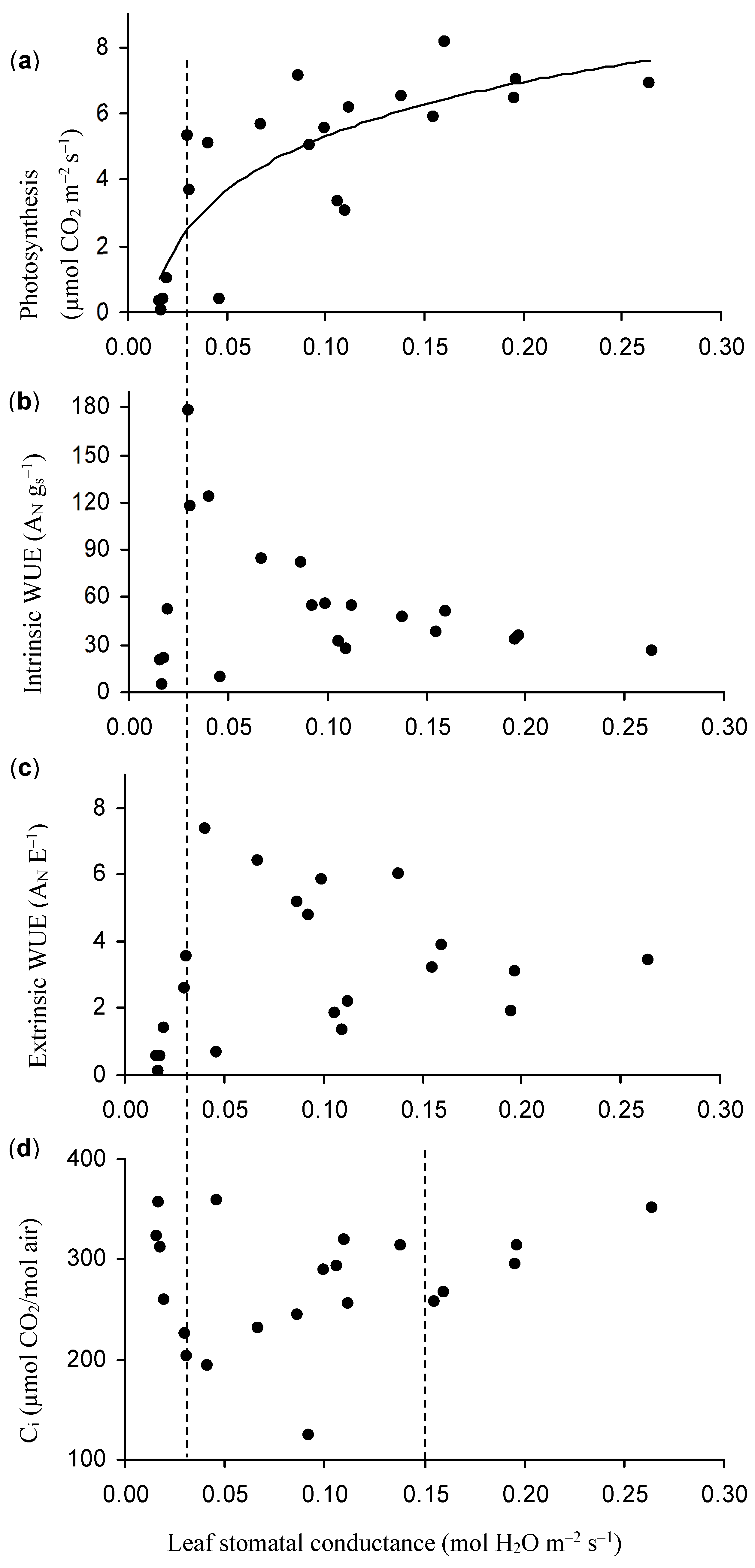
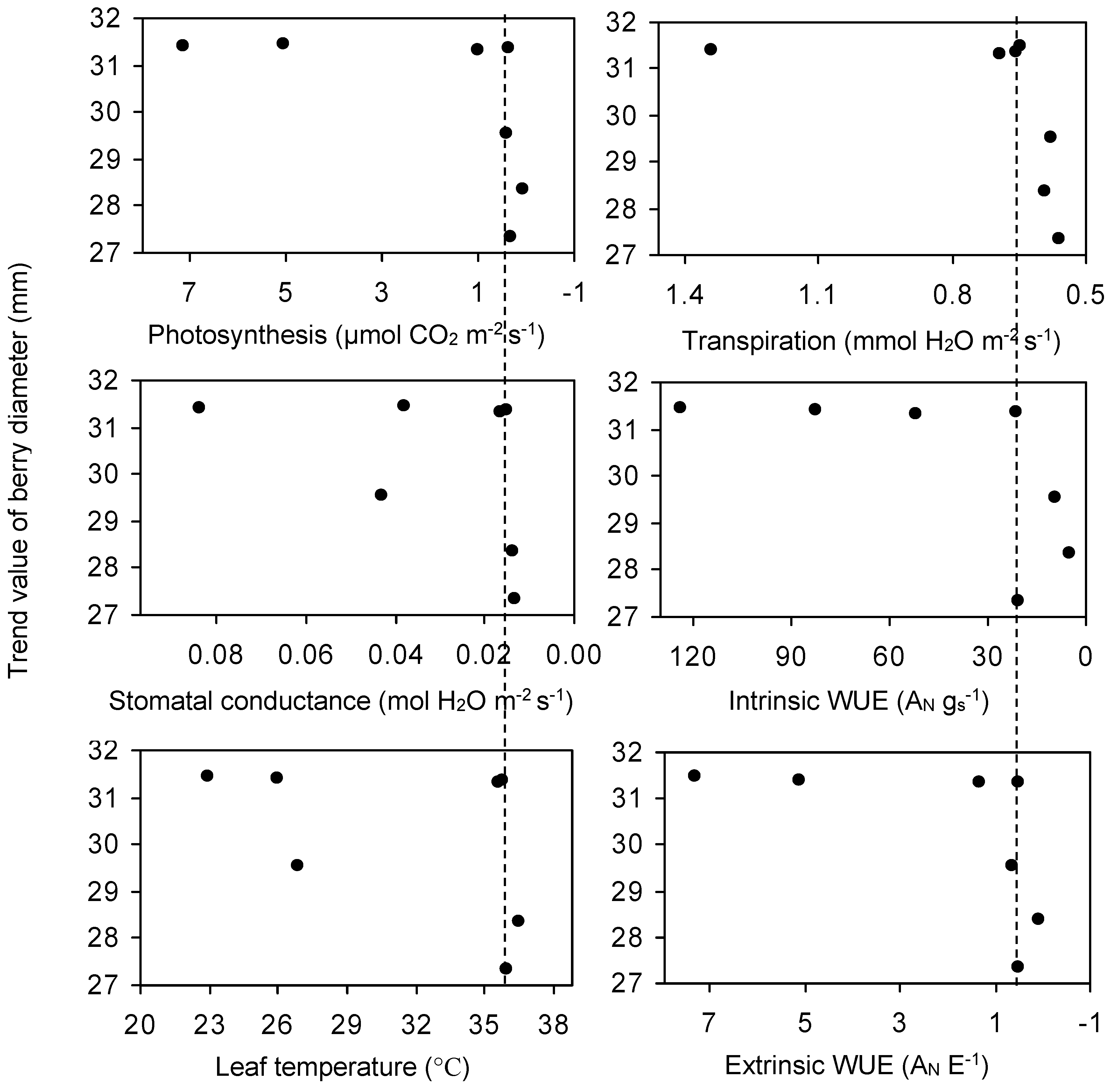
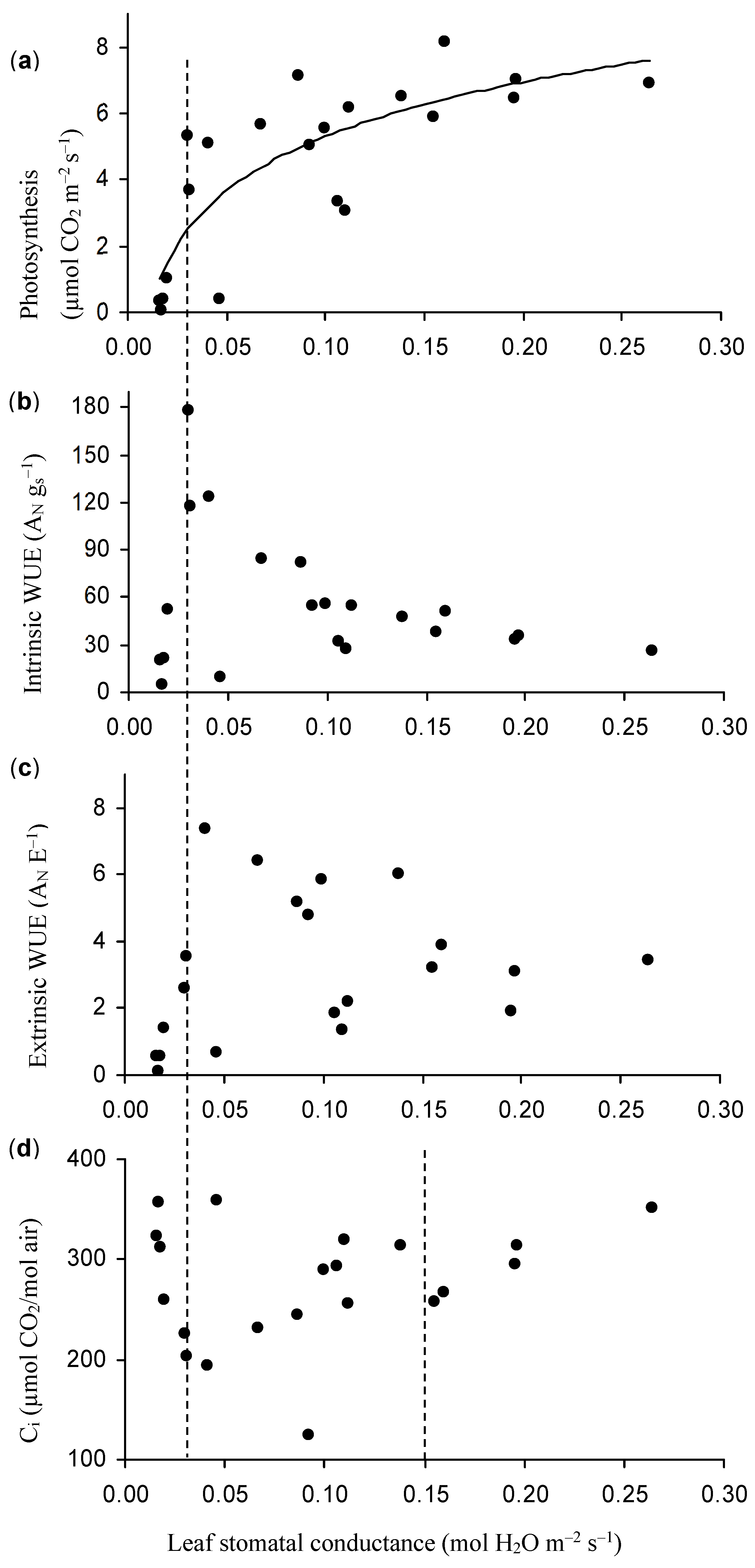
3.3. The Relationship among ψ, Berry Diameter, Leaf Photosynthesis, Stomatal Conductance, and Water Use
3.4. Suggestions for Irrigation Scheduling during the Maturation Stage of Grapevines
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Vörösmarty, C.J.; McIntyre, P.B.; Gessner, M.O.; Dudgeon, D.; Prusevich, A.; Green, P.; Glidden, S.; Bunn, S.E.; Sullivan, C.A.; Liermann, C.R.; et al. Global threats to human water security and river biodiversity. Natrue 2010, 467, 555–561. [Google Scholar] [CrossRef]
- Zhang, Q.T.; Wang, S.P.; Li, L.; Inoue, M.; Xiang, J.; Qiu, G.Y.; Jin, W.B. Effects of mulching and sub-surface irrigation on vine growth, berry sugar content and water use of grapevines. Agric. Water Manag. 2014, 143, 1–8. [Google Scholar] [CrossRef]
- Yan, N.N.; Wu, B.F.; Zhu, W.W. Assessment of agricultural water productivity in arid China. Water 2020, 12, 1161. [Google Scholar] [CrossRef]
- Conesa, M.R.; Dodd, I.C.; Temnani, A.; De la Rosa, J.M.; Pérez-Pastor, A. Physiological response of post-veraison deficit irrigation strategies and growth patterns of table grapes (cv. Crimson Seedless). Agric. Water Manag. 2018, 208, 363–372. [Google Scholar] [CrossRef] [Green Version]
- Santos, T.P.; Lopes, C.M.; Rodrigues, M.L.; de Souza, C.R.; Ricardo-da-Silva, J.M.; Maroco, J.P.; Pereira, J.S.; Chaves, M.M. Effects of deficit irrigation strategies on cluster microclimate for improving fruit composition of Moscatel field-grown grapevines. Sci. Hortic. 2007, 112, 321–330. [Google Scholar] [CrossRef]
- Schultz, H.R.; Stoll, M. Some critical issues in environmental physiology of grapevines: Future challenges and current limitations. Aust. J. Grape Wine Res. 2010, 16, 4–24. [Google Scholar] [CrossRef]
- Tomás, M.; Medrano, H.; Escalona, J.M.; Martorell, S.; Pou, A.; Ribas-Carbó, M.; Flexas, J. Variability of water use efficiency in grapevines. Environ. Exp. Bot. 2014, 103, 148–157. [Google Scholar] [CrossRef]
- Zhao, X.H.; Kang, L.F.; Wang, Q.; Lin, C.; Liu, W.; Chen, W.L.; Sang, T.; Yan, J. Water use efficiency and stress tolerance of the potential energy crop miscanthus lutarioriparius grown on the loess plateau of China. Plants 2021, 10, 544. [Google Scholar] [CrossRef]
- Poni, S.; Bernizzoni, F.; Civardi, S.; Gatti, M.; Porro, D.; Camin, F. Performance and water use efficiency (single-leaf vs. whole-canopy) of well-watered and half-stressed split-root Lambrusco grapevines grown in Po Valley (Italy). Agric. Ecosyst. Environ. 2009, 129, 97–106. [Google Scholar] [CrossRef]
- Chaves, M.M.; Zarrouk, O.; Francisco, R.; Costa, J.M.; Santos, T.; Regalado, A.P.; Rodrigues, M.L.; Lopes, C.M. Grapevine under deficit irrigation: Hints from physiolo-gical and molecular data. Ann. Bot. 2010, 105, 661–676. [Google Scholar] [CrossRef] [Green Version]
- Tomás, M.; Medrano, H.; Pou, A.; Escalona, J.M.; Martorell, S.; Ribas-Carbó, M.; Flexas, J. Water use efficiency in grapevine cultivars grown under controlled conditions: Effects of water stress at the leaf and whole-plant level. Aust. J. Grape Wine Res. 2012, 18, 164–172. [Google Scholar] [CrossRef]
- Atroosh, K.B.; Mukred, A.W.O.; Moustafa, A.T. Water requirement of grape (Vitis vinifera) in the northern highlands of Yemen. J. Agric. Sci. 2013, 5. [Google Scholar] [CrossRef] [Green Version]
- Centeno, A.; Baeza, P.; Lissarrague, J.R. Relationship between soil and plant water status in wine grapes under various water deficit regimes. HortTechnology 2010, 20, 585–593. [Google Scholar] [CrossRef] [Green Version]
- Lovisolo, C.; Perrone, I.; Hartung, W.; Schubert, A. An abscisic acid-related reduced transpiration promotes gradual embolism repair when grapevines are rehydrated after drought. New Phytol. 2008, 180, 642–651. [Google Scholar] [CrossRef]
- Perrone, I.; Gambino, G.; Chitarra, W.; Vitali, M.; Pagliarani, C.; Riccomagno, N.; Balestrini, R.; Kaldenhoff, R.; Uehlein, N.; Gribaudo, I.; et al. The grapevine root-Specific Aquaporin VvPIP2; 4N controls root hydraulic conductance and leaf gas exchange under well-watered conditions but not under water stress. Plant Physiol. 2012, 160, 965–977. [Google Scholar] [CrossRef] [Green Version]
- Bassoi, L.H.; Correia, J.D.; dos Santos, A.R.L.; Silva, J.A.; Costa, B.R.S. Deficit Irrigation in grapevine cv. Syrah during two growing seasons in the brazilian semiarid. Eng. Agric. 2015, 35, 430–441. [Google Scholar] [CrossRef] [Green Version]
- Fereres, E.; Goldhamer, D.A.; Parsons, L.R. Irrigation water management of horticultural crops. Hortscience 2004, 38, 1036–1042. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, A.H.D.; Sherer-Warren, M.; Hernandez, F.B.T.; Lopes, H.L. Water productivity assessment by using MODIS images and agrometeorological data in the Petrolina municipality, Brazil. Remote Sens. 2012, 8531. [Google Scholar] [CrossRef]
- Teixeira, A.H.D.; Hernandez, F.B.T.; Lopes, H.L. Up scaling table grape water requirements in the low-middle Sao Francisco river basin, Brazil. Acta. Hort. 2014, 1038, 655–662. [Google Scholar] [CrossRef]
- Konukcu, F.; Gowing, J.W.; Rose, D.A. Dry drainage: A sustainable solution to waterlogging and salinity problems in irrigation areas? Agric. Water Manag. 2006, 83, 1–12. [Google Scholar] [CrossRef]
- Romero, P.; Gil-Muñoz, R.; del Amor, F.M.; Valdés, E.; Fernández, J.I.; Martinez-Cutillas, A. Regulated deficit irrigation based upon optimum water status improves phenolic composition in Monastrell grapes and wines. Agric. Water Manag. 2013, 121, 85–101. [Google Scholar] [CrossRef]
- Jones, H.G. Irrigation scheduling: Advantages and pitfalls of plant-based methods. J. Exp. Bot. 2004, 55, 2427–2436. [Google Scholar] [CrossRef] [Green Version]
- Azorín, P.R.; García, J.G. The productive, economic, and social efficiency of vineyards using combined drought-tolerant rootstocks and efficient low water volume deficit irrigation techniques under mediterranean semiarid conditions. Sustainability 2020, 12, 1930. [Google Scholar] [CrossRef] [Green Version]
- Ton, Y.; Kopyt, M.; Zachs, I.; Ben-Ner, Z. Phytomonitoring technique for tuning irrigation of fruit trees. Acta. Hort. 2004, 646, 127–132. [Google Scholar] [CrossRef]
- Carr, M.K.V. The water relations and irrigation requirements of pineapple (Ananas comosus var. comosus): A review. Exp. Agric. 2011, 47, 27–51. [Google Scholar] [CrossRef]
- Goldhamer, D.A.; Fereres, E. Irrigation scheduling of almond trees with trunk diameter sensors. Irrig. Sci. 2004, 23, 11–19. [Google Scholar] [CrossRef]
- Fernández, J.E.; Cuevas, M.V. Irrigation scheduling from stem diameter variations: A review. Agric. For. Meteorol. 2010, 150, 135–151. [Google Scholar] [CrossRef]
- Avidan, A.; Hazan, A.; Kopyt, M.; Ton, Y.; Phytech, L. Application of the phytomonitoring technique for table grapes. In Proceedings of the International Workshop on Advances Ingrapevine and Wine Research, Venosa, Italy, 15–17 September 2005. [Google Scholar]
- Gratacos, E.; Gurovich, L. Phytomonitoring in kiwifruit orchards as a plant water status indicator and its use in irrigation scheduling. Cien. Inv. 2003, 30, 113–137. [Google Scholar] [CrossRef]
- Ton, Y.; Kopyt, M.; Nilov, N. Phytomonitoring technique for tuning irrigation of vineyards. Acta. Hort. 2004, 646, 133–139. [Google Scholar] [CrossRef]
- Ton, Y.; Kopyt, M. Phytomonitoring in realization of irrigation strategies for wine grapes. Acta. Hort. 2004, 652, 167–173. [Google Scholar] [CrossRef]
- Kopyt, M.; Ton, Y.; Tsadok, S. Chardonnay trunk diameter growth and microvariations: Four-year trial results and outlook for irrigation control. Aust. N. Z. Grapegrow. Winemak. 2005, 493, 23–25. [Google Scholar]
- Matthews, M.A.; Nuzzo, V. Berry size and yield paradigms on grapes and wines quality. Acta. Hort. 2007, 754, 423–435. [Google Scholar] [CrossRef] [Green Version]
- Imai, S.; Iwao, K.; Fujiwara, T. Measurements of plant physiological informations of vine tree and indexation of soil moisture control(3). Environ. Control Biol. 1991, 29, 19–26. [Google Scholar] [CrossRef]
- Greven, M.M.; Raw, V.; West, B.A. Effects of timing of water stress on yield and berry size. Water Sci. Technol. 2009, 60, 1249–1255. [Google Scholar] [CrossRef]
- Intrigliolo, D.S.; Castel, J.R. Evaluation of grapevine water status from trunk diameter variations. Irrig. Sci. 2007, 26, 49–59. [Google Scholar] [CrossRef] [Green Version]
- Chatelet, D.S.; Rost, T.L.; Matthews, M.A.; Shackel, K.A. The peripheral xylem of grapevine (Vitis vinifera) berries. 2. Anatomy and development. J. Expt. Bot. 2008, 59, 1997–2007. [Google Scholar] [CrossRef] [Green Version]
- Bittelli, M. Measuring soil water potential for water management in agriculture: A review. Sustainability 2010, 2, 1226–1251. [Google Scholar] [CrossRef] [Green Version]
- Barbagallo, M.G.; Vesco, G.; Di Lorenzo, R.; Lo Bianco, R.; Pisciotta, A. Soil and regulated deficit irrigation affect growth, yield and quality of ‘Nero d’Avola’ grapes in a semi-arid environment. Plants 2021, 10, 641. [Google Scholar] [CrossRef]
- Torres, R.; Ferrara, G.; Soto, F.; Lopez, J.A.; Sanchez, F.; Mazzeo, A.; Perez-Pastor, A.; Domingo, R. Effects of soil and climate in a table grape vineyard with cover crops. Irrigation management using sensors networks. Cienc. Tec. Vitivinic. 2017, 32, 72–81. [Google Scholar] [CrossRef] [Green Version]
- Cifre, J.; Bota, J.; Escalona, J.M.; Medrano, H.; Flexas, J. Physiological tools for irrigation scheduling in grapevine (Vitis vinifera L.): An open gate to improve water use efficiency? Agric. Ecosyst. Environ. 2005, 106, 159–170. [Google Scholar] [CrossRef]
- Zhang, Q.T.; Wang, S.P.; Inoue, M.; Moritani, S.; Tsuji, W.; Geng, S.; Qiu, G.Y.; Xie, Q. A new methodology for determining irrigation schedule of grapevines using photogrammetric measurement of berry diameter. J. Food Agric. Environ. 2012, 10, 582–587. [Google Scholar] [CrossRef]
- Chherawala, Y.; Lepage, R.; Doyon, G. Food grading/sorting based on color appearance trough machine vision: The case of fresh cranberries. Informat. Commun. Technol. 2006, 4, 1569–1574. [Google Scholar]
- Whalley, J.; Shanmuganathan, S. Applications of image processing in viticulture: A review. In Proceedings of the 20th International Congress on Modelling and Simulation, Adelaide, Australia, 1–6 December 2013; pp. 531–537. [Google Scholar]
- Herrero-Huerta, M.; Gonzalez-Aguilera, D.; Rodriguez-Gonzalvez, P.; Hernández-López, D. Vineyard yield estimation by automatic 3D bunch modelling in field conditi-ons. Comput. Electron. Agr. 2015, 110, 17–26. [Google Scholar] [CrossRef]
- Moritani, S.; Yamamoto, T.; Henintsoa, A.; Muraki, H. Monitoring of soil erosion using digital camera under simulated rainfall. Trans. JSIDRE 2006, 244, 545–551. [Google Scholar] [CrossRef]
- Wang, S.P.; Okamoto, G.; Hirano, K.; Lu, J.; Zhang, C.X. Effects of restricted rooting volume on vine growth and berry development of kyoho grapevines. Am. J. Enol. Vitic. 2001, 52, 248–253. [Google Scholar] [CrossRef]
- Marthaler, H.P.; Vogelsanger, W.; Richard, F.; Wierenga, P.J. A pressure transducer for field tensiometers. Soil Sci. Soc. Am. J. 1983, 47, 624–627. [Google Scholar] [CrossRef]
- Azooz, R.H.; Arshad, M.A. Laboratory calibration of pressure transducer-tensiometer system for hydraulic studies. Can. J. Soil. Sci. 1994, 74, 315–319. [Google Scholar] [CrossRef]
- Moran, M.S.; Scott, R.L.; Keefer, T.O.; Emmerich, W.E.; Hernandez, M.; Nearing, G.S.; Paige, G.B.; Cosh, M.H.; O’Neill, P.E. Partitioning evapotranspiration in semiaridgrassland and shrubland ecosystems using time series of soil surface temperature. Agric. For. Meteorol. 2009, 149, 59–72. [Google Scholar] [CrossRef]
- Conesa, M.R.; Torres, R.; Domingo, R.; Navarro, H.; Soto, F.; Perez-Pastor, A. Maximum daily trunk shrinkage and stem water potential reference equations for irrigation scheduling in table grapes. Agric. Water Manag. 2016, 172, 51–61. [Google Scholar] [CrossRef]
- Keller, M.; Smith, J.P.; Bondada, B.R. Ripening grape berries remain hydraulically connected to the shoot. J. Exp. Bot. 2006, 57, 2577–2587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meggio, F.; Trevisan, S.; Manoli, A.; Ruperti, B.; Quaggiotti, S. Systematic Investigation of the effects of a novel protein hydrolysate on the growth, physiological parameters, fruit development and yield of grapevine (Vitis Vinifera L., cv Sauvignon Blanc) under water stress conditions. Agronomy 2020, 10, 1785. [Google Scholar] [CrossRef]
- Zsόfi, Z.; Gal, L.; Szilagyi, Z.; Szucs, E.; Marschall, M.; Nagy, Z.; Balo, B. Use of stomatal conductance and pre-dawn water potential to classify terroir for the grape variety Kékfrankos. Aust. J. Grape Wine Res. 2009, 15, 36–47. [Google Scholar] [CrossRef]
- Goldhamer, D.A.; Fereres, E.; Mata, M.; Girona, J.; Cohen, M. Sensitivity of continuous and discrete plant and soil water status monitoring in peach trees subjected to deficit irrigation. J. Am. Soc. Hortic. Sci. 1999, 124, 437–444. [Google Scholar] [CrossRef] [Green Version]
- Boini, A.; Manfrini, L.; Bortolotti, G.; Corelli-Grappadelli, L.; Morandi, B. Monitoring fruit daily growth indicates the onset of mild drought stress in apple. Sci. Hortic. 2019, 256. [Google Scholar] [CrossRef]
- Resco, V.; Ewers, B.E.; Sun, W.; Huxman, T.E.; Weltzin, J.F.; Williams, D.G. Drought-induced hydraulic limitations constrain leaf gas exchange recovery after precipitation pulses in the C3 woody legume, prosopis velutina. New Phytol. 2009, 181, 672–682. [Google Scholar] [CrossRef]
- Santesteban, L.G.; Royo, J.B. Water status, leaf area and fruit load influence on berry weight and sugar accumulation of cv. ‘Tempranillo’ under semiarid conditions. Sci. Hortic. 2006, 109, 60–65. [Google Scholar] [CrossRef]
- Koundouras, S.; Tsialtas, I.T.; Zioziou, E.; Nikolaou, N. Rootstock effects on the adaptive strategies of grapevine (Vitis vinifera L. cv. Cabernet-Sauvignon) under contrasting water status: Leaf physiological and structural responses. Agric. Ecosyst. Environ. 2008, 128, 86–96. [Google Scholar] [CrossRef]
- Maroco, J.P.; Rodrigues, M.L.; Lopes, C.; Chaves, M.M. Limitations to leaf photosynthesis in field-grown grapevine under drought-metabolic and modelling approaches. Funct. Plant Biol. 2002, 29, 451–459. [Google Scholar] [CrossRef] [PubMed]
- Quick, W.P.; Chaves, M.M.; Wendler, R.; David, M.; Rodrigues, M.L.; Passaharinho, J.A.; Pereira, J.S.; Adcock, M.D.; Leegood, R.C.; Stitt, M. The effect of water stress on photosynthetic carbon metabolism in four species grown under field conditions. Plant Cell Environ. 1992, 15, 25–35. [Google Scholar] [CrossRef]
- Flexas, J.; Escalona, J.M.; Medrano, H. Water stress induces different levels of photosynthesis and electron transport rate regulation in grapevines. Plant Cell Environ. 1999, 22, 39–48. [Google Scholar] [CrossRef]
- Flexas, J.; Bota, J.; Escalona, J.M.; Sampol, B.; Medrano, H. Effects of drought on photosynthesis in grapevines under field conditions: An evaluation of stomatal and mesophyll limitations. Funct. Plant Biol. 2002, 29, 461–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamatiadis, S.; Christofides, C.; Tsadila, E.; Taskos, D.; Tsadilas, C.; Schepers, J.S. Relationship of leaf stable isotopes (delta 13C and delta 15N) to biomass production in two fertilized merlot vineyards. Am. J. Enol. Vitic. 2007, 58, 67–74. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Date | Time | 7:00 | 9:00 | 11:00 | 13:00 | 15:00 | 17:00 | 19:00 | 21:00 |
|---|---|---|---|---|---|---|---|---|---|
| 13 August | Ψ (kPa) | −0.33 | −0.93 | −1.18 | −1.33 | −1.69 | −1.91 | −1.86 | −1.86 |
| SD | 0.35 | 0.23 | 0.16 | 0.24 | 0.26 | 0.18 | 0.16 | 0.14 | |
| SE | 0.14 | 0.09 | 0.07 | 0.10 | 0.11 | 0.07 | 0.07 | 0.06 | |
| 14 August | Ψ (kPa) | −2.21 | −2.12 | −2.21 | −2.29 | −2.44 | −2.72 | −2.77 | −2.84 |
| SD | 0.17 | 0.21 | 0.12 | 0.18 | 0.14 | 0.12 | 0.09 | 0.06 | |
| SE | 0.09 | 0.05 | 0.07 | 0.06 | 0.05 | 0.04 | 0.03 | 0.05 | |
| 15 August | Ψ (kPa) | −3.22 | −3.02 | −3.15 | −3.29 | −3.40 | −3.60 | −3.65 | −3.72 |
| SD | 0.05 | 0.15 | 0.18 | 0.08 | 0.07 | 0.15 | 0.11 | 0.05 | |
| SE | 0.02 | 0.06 | 0.07 | 0.03 | 0.03 | 0.06 | 0.04 | 0.02 | |
| 16 August | Ψ (kPa) | −4.12 | −4.30 | −4.50 | −4.85 | −5.22 | −5.35 | −5.62 | −5.76 |
| SD | 0.15 | 0.15 | 0.23 | 0.14 | 0.26 | 0.20 | 0.28 | 0.38 | |
| SE | 0.06 | 0.06 | 0.09 | 0.06 | 0.11 | 0.08 | 0.11 | 0.16 | |
| 17 August | Ψ (kPa) | −6.87 | −7.14 | −7.92 | −8.57 | −9.37 | −9.17 | −10.04 | −10.65 |
| SD | 0.36 | 0.44 | 0.44 | 0.71 | 1.07 | 0.96 | 1.13 | 1.31 | |
| SE | 0.15 | 0.18 | 0.18 | 0.29 | 0.44 | 0.39 | 0.46 | 0.54 | |
| 18 August | Ψ (kPa) | −14.61 | −13.20 | −14.67 | −15.14 | −14.79 | −15.57 | −16.15 | −19.25 |
| SD | 1.72 | 2.01 | 2.05 | 1.79 | 1.86 | 2.84 | 1.89 | 2.05 | |
| SE | 0.70 | 0.82 | 0.84 | 0.73 | 0.76 | 1.16 | 0.77 | 0.84 | |
| 19 August | Ψ (kPa) | −21.87 | −16.44 | −17.14 | −17.62 | −17.45 | −18.22 | −20.39 | −21.87 |
| SD | 2.30 | 2.51 | 2.53 | 2.89 | 3.12 | 3.24 | 2.55 | 2.30 | |
| SE | 0.94 | 1.02 | 1.03 | 1.18 | 1.27 | 1.32 | 1.04 | 0.94 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Q.; Chen, Y.; Xiong, Y.; Moritani, S.; Wu, X.; Yan, C.; Chen, X. Which Is More Sensitive to Water Stress for Irrigation Scheduling during the Maturation Stage: Grapevine Photosynthesis or Berry Size? Atmosphere 2021, 12, 845. https://doi.org/10.3390/atmos12070845
Zhang Q, Chen Y, Xiong Y, Moritani S, Wu X, Yan C, Chen X. Which Is More Sensitive to Water Stress for Irrigation Scheduling during the Maturation Stage: Grapevine Photosynthesis or Berry Size? Atmosphere. 2021; 12(7):845. https://doi.org/10.3390/atmos12070845
Chicago/Turabian StyleZhang, Qingtao, Yixuan Chen, Yujiu Xiong, Shigeoki Moritani, Xinyu Wu, Chunhua Yan, and Xiaohong Chen. 2021. "Which Is More Sensitive to Water Stress for Irrigation Scheduling during the Maturation Stage: Grapevine Photosynthesis or Berry Size?" Atmosphere 12, no. 7: 845. https://doi.org/10.3390/atmos12070845
APA StyleZhang, Q., Chen, Y., Xiong, Y., Moritani, S., Wu, X., Yan, C., & Chen, X. (2021). Which Is More Sensitive to Water Stress for Irrigation Scheduling during the Maturation Stage: Grapevine Photosynthesis or Berry Size? Atmosphere, 12(7), 845. https://doi.org/10.3390/atmos12070845