
Potential Human and Plant Pathogenic Species in Airborne PM10 Samples and Relationships with Chemical Components and Meteorological Parameters

 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Monitoring Site Location and PM10 Sample Collection
2.2. Ions, Metals, and Organic and Elemental Carbon Analyses
2.3. DNA Extraction and 16SrRNA Gene High-Throughput Sequencing
2.4. Establishment of Potential Human and Plant Pathogenic Species and Non-Pathogenic Species
2.5. Statistical Methodologies for Data Analysis
3. Results and Discussion
3.1. Detection of Potential (Opportunistic) Human and Plant Pathogenic Species and Non-Pathogenic Species
3.2. Chemical Components and Potential (Opportunistic) Human and Plant Pathogens, and Non-Pathogenic Species in Winter and Spring Samples
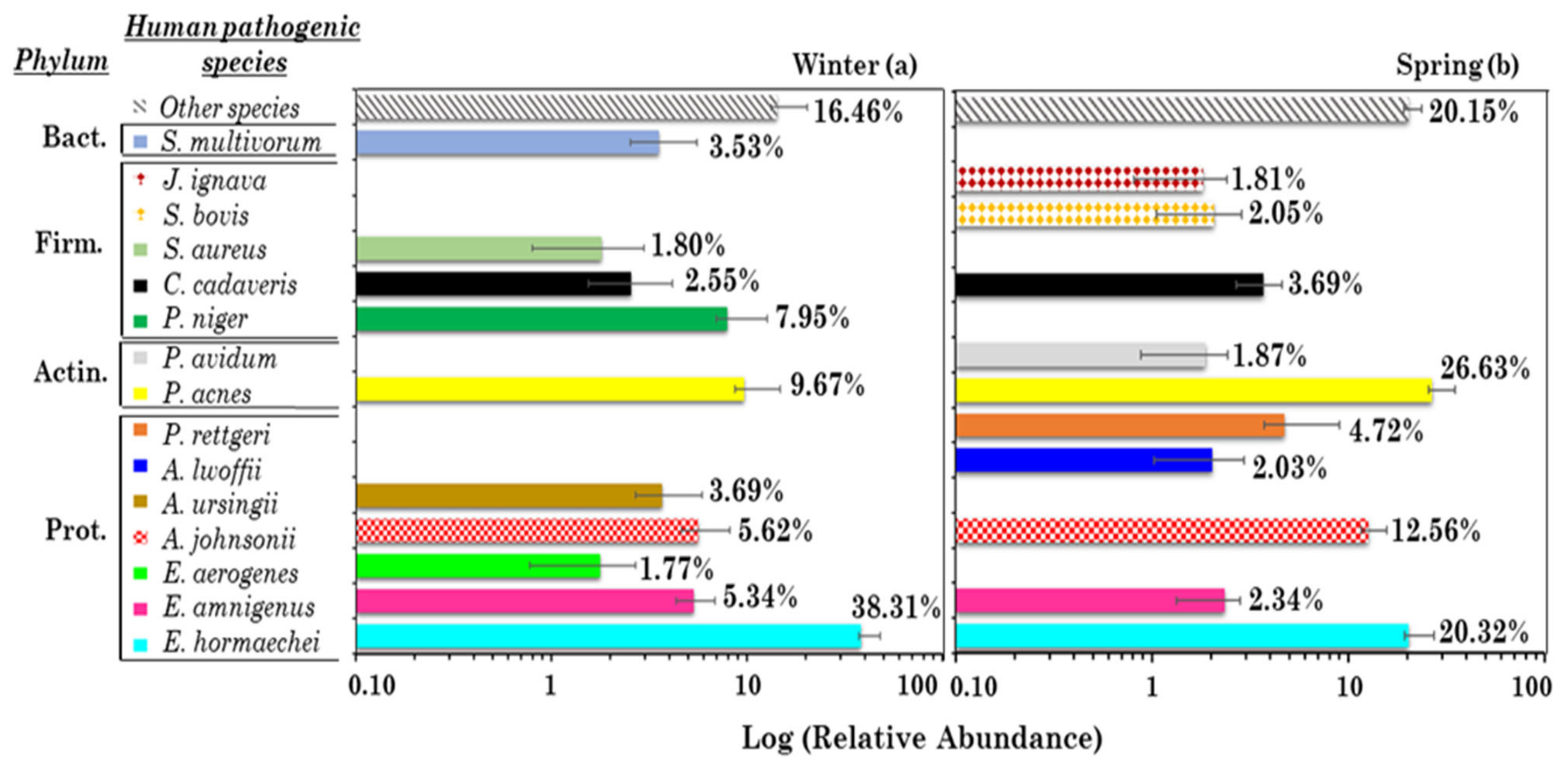
3.2.1. Analysis of the 10 Most Abundant and Pervasive Potential (Opportunistic) Human Pathogens in Winter and Spring
3.2.2. Correlations between the 10 Most Abundant and Pervasive Potential Human Pathogens, and with PM10 and Chemical Component Mass Concentrations, and Meteorological Parameters
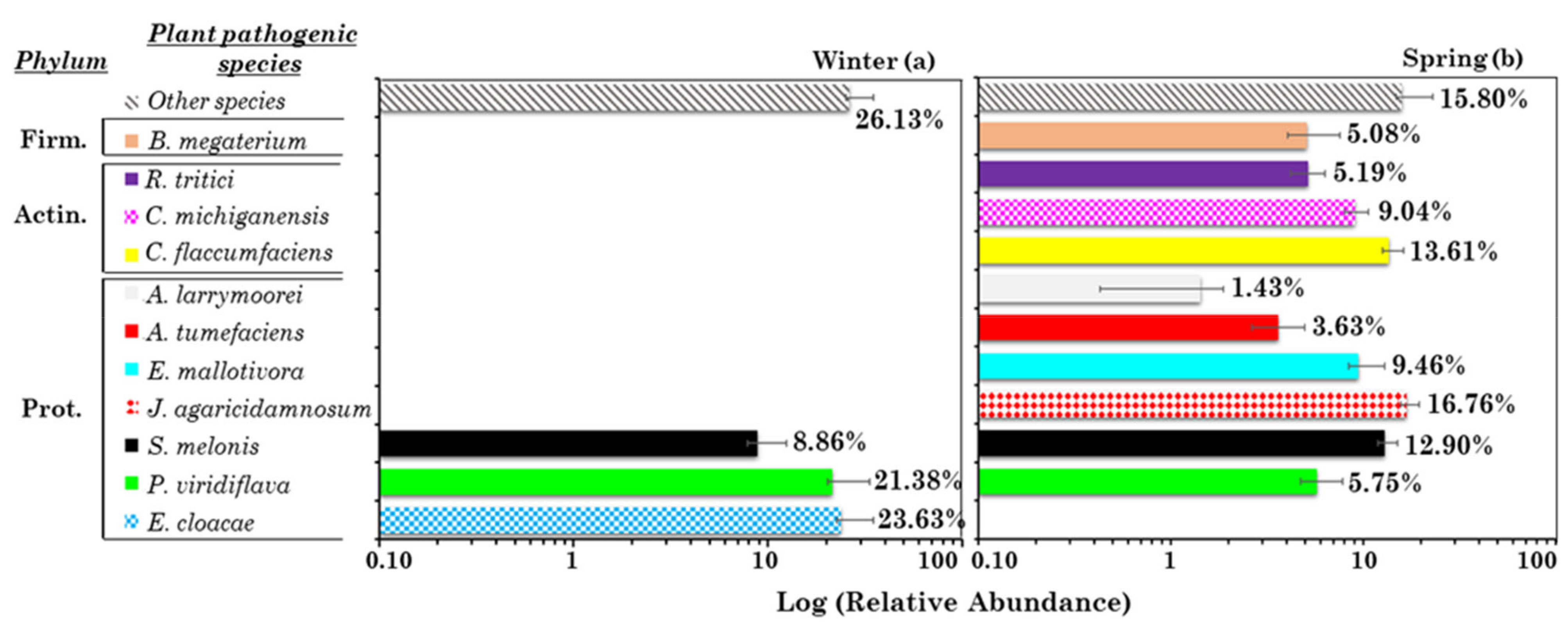
3.2.3. Analysis of the Most Abundant and Pervasive Potential (Opportunistic) Plant Pathogenic Species in Winter and Spring
3.2.4. Correlations between the Most Abundant and Pervasive Potential Plant Pathogens and with Human Pathogens, PM10 and Chemical Component Mass Concentrations, and Meteorological Parameters
3.2.5. Analysis of the 10 Most Abundant and Pervasive Potential Non-Pathogenic Species in Winter and Spring
3.2.6. Correlations between the Most Abundant and Pervasive Potential Non-Pathogenic Species, and with the Human and Plant Pathogens, PM10 and Chemical Component Mass Concentrations, and Meteorological Parameters
3.3. Carriers of Potential Non-Pathogenic Species in Winter and Spring
4. Conclusions
- Sample chemical composition and bacterial community varied from winter to spring. In particular, the total number of detected bacterial species increased more than twice from winter to spring. The stagnant and more favorable atmospheric conditions in spring for the bacterial survival and long-distance aerial dispersal likely contributed to this result, in addition to the seasonal dependence of the bacterial species-carriers.
- The number of strong relationships between potential (human and plant) pathogens and non-pathogens, chemical components, and meteorological parameters increased slightly from winter to spring, according to the Spearman coefficients.
- Rather few potential (opportunistic) human pathogens were significantly correlated with meteorological parameters. Conversely, many potential plant pathogens were strongly and positively correlated with wind direction and speed in winter and spring, suggesting that the dispersal of plant pathogens by the wind may likely contribute to the spreading of plant diseases.
- In winter, some potential human pathogens were correlated with chemical components that are tracers of marine and soil dust/mixed anthropogenic sources. Conversely, in spring, some potential human pathogens were mainly correlated with chemical components considered as marine aerosol tracers.
- We found that potential plant pathogens were not correlated among them in winter and that Enterobacter cloacae was the only plant pathogen significantly and positively correlated with the identified potential human pathogens. Few positive and significant correlations occurred between the plant pathogens and chemical component mass concentrations.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lelieveld, J.; Evans, J.S.; Fnais, M.; Giannadaki, D.; Pozzer, A. The Contribution of Outdoor Air Pollution Sources to Premature Mortality on a Global Scale. Nature 2015, 525, 367–371. [Google Scholar] [CrossRef]
- Wardeh, M.; Risley, C.; McIntyre, M.; Setzkorn, C.; Baylis, M. Database of host-pathogen and related species interactions, and their global distribution. Sci. Data 2015, 2, 150049. [Google Scholar] [CrossRef]
- Be, N.A.; Thissen, J.B.; Fofanov, V.Y.; Allen, J.E.; Rojas, M.; Golovko, G.; Fofanov, Y.; Koshinsky, H.; Jaing, C.J. Metagenomic Analysis of the Airborne Environment in Urban Spaces. Microb. Ecol. 2015, 69, 346–355. [Google Scholar] [CrossRef]
- Fröhlich-Nowoisky, J.; Kampf, C.J.; Weber, B.; Huffman, J.A.; Pöhlker, C.; Andreae, M.O.; Lang-Yona, N.; Burrows, S.M.; Gunthe, S.S.; Elbert, W.; et al. Bioaerosols in the earth system: Climate, health, and ecosystem interactions. Atmos. Res. 2016, 182, 346–376. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, X.; Zhang, H.; Yao, X.; Zhou, M.; Wang, J.; He, Z.; Zhang, H.; Lou, L.; Mao, W.; et al. Effect of air pollution on the total bacteria and pathogenic bacteria in different sizes of particulate matter. Environ. Pollut. 2018, 233, 483–493. [Google Scholar] [CrossRef]
- Fan, C.; Li, Y.; Liu, P.; Mu, F.; Xie, Z.; Lu, R.; Qi, Y.; Wang, B.; Jin, C. Characteristics of airborne opportunistic bacteria during autumn and winter in Xi’an, China. Sci. Total. Environ. 2019, 672, 834–845. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Kumari, D.; Achal, V. A Review on Airborne Microbes: The Characteristics of Sources, Pathogenicity and Geography. Atmosphere 2020, 11, 919. [Google Scholar] [CrossRef]
- Aylor, D.E. Spread of plant disease on a continental scale: Role of aerial dispersal of pathogens. Ecology 2003, 84, 1989–1997. [Google Scholar] [CrossRef]
- Van Leuken, J.P.G.; Swart, A.N.; Havelaar, A.H.; Van Pul, A.; Van der Hoek, W.; Heederik, D. Atmospheric dispersion modelling of bioaerosols that are pathogenic to humans and livestock—A review to inform risk assessment studies. Microb. Risk Analysis 2016, 1, 19–39. [Google Scholar]
- Kallos, G.; Astitha, M.; Katsafados, P.; Spyrou, C. Long-range transport of anthropogenically and naturally produced particulate matter in the Mediterranean and North Atlantic: Current state of knowledge. J. Appl. Meteorol. Climatol. 2007, 46, 1230–1251. [Google Scholar] [CrossRef]
- Maheras, P.; Xoplaki, E.; Davies, T.; Martin-Vide, J.; Bariendos, M.; Alcoforado, M.J. Warm and cold monthly anomalies across the Mediterranean basin and their relationship with circulation; 1860–1990. Int. J. Climatol. 1999, 19, 1697–1715. [Google Scholar] [CrossRef]
- Perrone, M.R.; Romano, S. Relationship between the planetary boundary layer height and the particle scattering coefficient at the surface. Atmos. Res. 2018, 213, 57–69. [Google Scholar] [CrossRef]
- Perrone, M.R.; Bertoli, I.; Romano, S.; Russo, M.; Rispoli, G.; Pietrogrande, M.C. PM2.5 and PM10 oxidative potential at a Central Mediterranean Site: Contrasts between dithiothreitol- and ascorbic acid-measured values in relation with particle size and chemical composition. Atmos. Environ. 2019, 210, 143–155. [Google Scholar] [CrossRef]
- Perrone, M.R.; Vecchi, R.; Romano, S.; Becagli, S.; Traversi, R.; Paladini, F. Weekly cycle assessment of PM mass concentrations and sources, and impacts on temperature and wind speed in Southern Italy. Atmos. Res. 2019, 218, 129–144. [Google Scholar] [CrossRef]
- Romano, S.; Di Salvo, M.; Rispoli, G.; Alifano, P.; Perrone, M.R.; Talà, A. Airborne Bacteria in the Central Mediterranean: Structure and role of meteorology and air mass transport. Sci. Total. Environ. 2019, 697, 134020. [Google Scholar] [CrossRef]
- Romano, S.; Becagli, S.; Lucarelli, F.; Rispoli, G.; Perrone, M.R. Airborne Bacteria Structure and Chemical Composition Relationships in Winter and Spring PM10 Samples over Southeastern Italy. Sci. Total Environ. 2020, 730, 138899. [Google Scholar] [CrossRef]
- Perrone, M.; Becagli, S.; Garcia Orza, J.A.G.; Vecchi, R.; Dinoi, A.; Udisti, R.; Cabello, M. The impact of long-range-transport on PM1 and PM2.5 at a Central Mediterranean site. Atmos. Environ. 2013, 71, 176–186. [Google Scholar] [CrossRef]
- Mykytczuk, N.C.; Wilhelm, R.C.; Whyte, L.G. Planococcus halocryophilus sp. nov., an extreme sub-zero species from high Arctic permafrost. Int. J. Syst. Evol. Microbiol. 2012, 62, 1937–1944. [Google Scholar] [CrossRef]
- Becagli, S.; Anello, F.; Bommarito, C.; Cassola, F.; Calzolai, G.; Di Iorio, T.; di Sarra, A.; Gómez-Amo, J.-L.; Lucarelli, F.; Marconi, M.; et al. Constraining the ship contribution to the aerosol of the central Mediterranean. Atmos. Chem. Phys. 2017, 17, 2067–2084. [Google Scholar] [CrossRef]
- Cavalli, F.; Viana, M.; Yttri, K.; Genberg, J.; Putaud, J. Toward a standardized thermal-optical protocol for measuring atmospheric organic and elemental carbon: The EUSAAR protocol. Atmos. Meas. Tech. 2010, 3, 79–89. [Google Scholar] [CrossRef]
- Lucarelli, F.; Nava, S.; Calzolai, G.; Chiari, M.; Udisti, R.; Marino, F. Is PIXE still a useful technique for the analysis of atmospheric aerosols? The LABEC experience. X-Ray Spectrom. 2011, 40, 162–167. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microb. 2007, 73, 5261e5267. [Google Scholar] [CrossRef] [PubMed]
- Werner, J.J.; Koren, O.; Hugenholtz, P.; DeSantis, T.Z.; Walters, W.A.; Caporaso, J.G.; Angenent, L.T.; Knight, R.; Ley, R.E. Impact of training sets on classification of high-throughput bacterial 16s rRNA gene surveys. ISME J. 2012, 6, 94–103. [Google Scholar] [CrossRef]
- Roura, A.; Doyle, S.R.; Nande, M.; Strugnell, J.M. You are what you eat: A genomic analysis of the gut microbiome of captive and wild Octopus vulgaris paralarvae and their zooplankton prey. Front. Physiol. 2017, 31, 362. [Google Scholar] [CrossRef]
- Ghaju Shrestha, R.; Tanaka, Y.; Malla, B.; Bhandari, D.; Tandukar, S.; Inoue, D.; Sei, K.; Sherchand, J.B.; Haramoto, E. Next-generation sequencing identification of pathogenic bacterial genes and their relationship with fecal indicator bacteria in different water sources in the Kathmandu Valley, Nepal. Sci. Total Environ. 2017, 1, 278–284. [Google Scholar] [CrossRef]
- Hugon, P.; Dufour, J.C.; Colson, P.; Fournier, P.E.; Sallah, K.; Raoult, D. A comprehensive repertoire of prokaryotic species identified in human beings. Lancet Infect. Dis. 2015, 15, 1211–1219. [Google Scholar] [CrossRef]
- Bull, C.T.; de Boer, S.H.; Denny, T.P.; Firrao, G.; Fischer-Le Saux, M.; Saddler, G.S.; Scortichini, M.; Stead, D.E.; Takikawa, Y. Comprehensive list of names of plant pathogenic bacteria, 1980–2007. Plant. Pathol. J. 2010, 92, 551–592. [Google Scholar]
- Bibby, K.; Viau, E.; Peccia, J. Pyrosequencing of the 16S rRNA gene to reveal bacterial pathogen diversity in biosolids. Water Res. 2010, 44, 4252–4260. [Google Scholar] [CrossRef]
- Guo, F.; Zhang, T. Profiling bulking and foaming bacteria in activated sludge by high throughput sequencing. Water Res. 2012, 46, 2772–2782. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, 590–596. [Google Scholar] [CrossRef] [PubMed]
- Ye, L.; Zhang, T. Pathogenic bacteria in sewage treatment plants as revealed by 454 pyrosequencing. Environ. Sci. Technol. 2011, 45, 7173–7179. [Google Scholar] [CrossRef]
- Hammer, O.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Sulaymon, I.D.; Zhang, Y.; Hopke, P.K.; Zhang, Y.; Hua, J.; Mei, X. COVID-19 pandemic in Wuhan: Ambient air quality and the relationships between criteria air pollutants and meteorological variables before, during, and after lockdown. Atmos. Res. 2021, 250, 105362. [Google Scholar] [CrossRef]
- Zuur, A.F.; Ieno, E.N.; Smith, G.M. Analysing Ecological Data. In Statistics for Biology and Health; Springer: New York, NY, USA, 2007. [Google Scholar]
- Romano, S.; Becagli, S.; Lucarelli, F.; Russo, M.; Pietrogrande, M.C. Oxidative Potential Sensitivity to Metals, Br, P, S, and Se in PM10 Samples: New Insights from a Monitoring Campaign in Southeastern Italy. Atmosphere 2020, 11, 367. [Google Scholar] [CrossRef]
- Van den Wollenberg, A.L. Redundancy analysis an alternative for canonical correlation analysis. Psychometrika 1977, 42, 207–219. [Google Scholar] [CrossRef]
- Paliy, O.; Shankar, V. Application of multivariate statistical techniques in microbial ecology. Mol. Ecol. 2016, 25, 1032–1057. [Google Scholar] [CrossRef]
- Hervé, M.R.; Nicolè, F.; Lê Cao, K.A. Multivariate Analysis of Multiple Datasets: A Practical Guide for Chemical Ecology. J. Chem. Ecol. 2018, 44, 215–234. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.L. Fathom Toolbox for MATLAB: Software for Multivariate Ecological and Oceanographic Data Analysis; College of Marine Science, University of South Florida: St. Petersburg, FL, USA, 2017; Available online: https://www.marine.usf.edu/research/matlab-resources/ (accessed on 26 April 2021).
- Sun, Y.; Xu, S.; Zheng, D.; Li, J.; Tian, H.; Wang, Y. Effects of haze pollution on microbial community changes and correlation with chemical components in atmospheric particulate matter. Sci. Total Environ. 2018, 637–638, 507–516. [Google Scholar] [CrossRef]
- Angeler, D.G.; Viedma, O.; Moreno, J.M. Statistical performance and information content of time lag analysis and redundancy analysis in time series modeling. Ecology 2009, 90, 3245–3257. [Google Scholar] [CrossRef] [PubMed]
- Warton, D.I. Many zeros does not mean zero inflation: Comparing the goodness-of-fit of parametric models to multivariate abundance data. Environmetrics 2005, 16, 275–289. [Google Scholar] [CrossRef]
- Herliansyah, R.; Fitria, I. Latent variable models for multi-species counts modeling in ecology. Biodiversitas 2018, 19, 1871–1876. [Google Scholar] [CrossRef]
- Caraka, R.E.; Chen, R.C.; Lee, Y.; Toharudin, T.; Rahmadi, C.; Tahmid, M.; Achmadi, A.S. Using multivariate generalized linear latent variable models to measure the difference in event count for stranded marine animals. Global J. Environ. Sci. Manag. 2021, 7, 117–130. [Google Scholar]
- Han, D.; Zhang, C.; Wang, C.; She, J.; Sun, Z.; Zhao, D.; Bian, Q.; Han, W.; Yin, L.; Sun, R.; et al. Differences in Response of Butterfly Diversity and Species Composition in Urban Parks to Land Cover and Local Habitat Variables. Forests 2021, 12, 140. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Hughes, D.D.; Mampage, C.B.A.; Jones, L.M.; Liu, Z.; Stone, E.A. Characterization of Atmospheric Pollen Fragments during Springtime Thunderstorms. Environ. Sci. Technol. Lett. 2020, 7, 409–414. [Google Scholar] [CrossRef]
- Ouyang, W.; Gao, B.; Cheng, H.; Zhang, L.; Wang, Y.; Lin, C.; Chen, J. Airborne bacterial communities and antibiotic gene dynamics in PM2.5 during rainfall. Environ. Int. 2020, 134, 105318. [Google Scholar] [CrossRef]
- Burrows, S.M.; Butler, T.; Jöckel, P.; Tost, H.; Kerkweg, A.; Pöschl, U.; Lawrence, M.G. Bacteria in the global atmosphere—Part 2: Modeling of emissions and transport between different ecosystems. Atmos. Chem. Phys. 2009, 9, 9281–9297. [Google Scholar] [CrossRef]
- Halda-Alija, L.; Hendricks, S.P.; Johnston, T.C. Spatial and temporal variation of Enterobacter genotypes in sediments and the underlying hyporheic zone of an agricultural stream. Microb. Ecol. 2001, 42, 286–294. [Google Scholar] [CrossRef] [PubMed]
- Paauw, A.; Caspers, M.P.; Leverstein-van Hall, M.A.; Schuren, F.H.; Montijn, R.C.; Verhoef, J.; Fluit, A.C. Identification of resistance and virulence factors in an epidemic Enterobacter hormaechei outbreak strain. Microbiology 2009, 155, 1478–1488. [Google Scholar] [CrossRef] [PubMed]
- Dessinioti, C.; Katsambas, A.D. The role of Propionibacterium acnes in acne pathogenesis: Facts and controversies. Clin. Dermatol. 2010, 28, 2–7. [Google Scholar] [CrossRef]
- McDowell, A.; Nagy, I.; Magyari, M.; Barnard, E.; Patrick, S. The Opportunistic Pathogen Propionibacterium acnes: Insights into Typing, Human Disease, Clonal Diversification and CAMP Factor Evolution. PLoS ONE 2013, 8, e70897. [Google Scholar] [CrossRef] [PubMed]
- Zeller, V.; Ghorbani, A.; Strady, C.; Leonard, P.; Mamoudy, P.; Desplaces, N. Propionibacterium acnes: An agent of prosthetic joint infection and colonization. J. Infect. 2007, 55, 119–124. [Google Scholar] [CrossRef]
- Murdoch, D.A. Gram-positive anaerobic cocci. Clin. Microbiol. Rev. 1998, 11, 81–120. [Google Scholar] [CrossRef] [PubMed]
- Polymenakou, P.N.; Mandalakis, M.; Stephanou, E.G.; Tselepides, A. Particle size distribution of airborne microorganisms and pathogens during an intense African dust event in the Eastern Mediterranean. Environ. Health Persp. 2008, 116, 292–296. [Google Scholar] [CrossRef] [PubMed]
- Mazar, Y.; Cytryn, E.; Erel, Y.; Rudich, Y. Effect of dust storms on the atmospheric microbiome in the Eastern Mediterranean. Environ. Sci. Technol. 2016, 50, 4194–4202. [Google Scholar] [CrossRef]
- Gat, D.; Mazar, Y.; Cytryn, E.; Rudich, Y. Origin-dependent variations in the atmospheric microbiome community in Eastern Mediterranean Dust Storms. Environ. Sci. Technol. 2017, 51, 6709–6718. [Google Scholar] [CrossRef]
- Romano, S.; Vecchi, R.; Perrone, M.R. Intensive optical parameters of pollution sources identified by the positive matrix factorization technique. Atmos. Res. 2020, 244, 105029. [Google Scholar] [CrossRef]
- Du, P.; Du, R.; Ren, W.; Lu, Z.; Zhang, Y.; Fu, P. Variations of bacteria and fungi in PM2.5 in Beijing, China. Atmos. Environ. 2018, 172, 55–64. [Google Scholar] [CrossRef]
- Perrone, M.R.; Romano, S.; Orza, J.A.G. Columnar and ground-level aerosol optical properties: Sensitivity to the transboundary pollution, daily and weekly patterns, and relationships. Environ. Sci. Pollut. R. 2015, 22, 16570–16589. [Google Scholar] [CrossRef]
- Hettema, J.M.; Prescott, C.A.; Kendler, K.S. Genetic and Environmental Sources of Covariation between Generalized Anxiety Disorder and Neuroticism. Am. J. Psychiatry 2004, 161, 1581–1587. [Google Scholar] [CrossRef]
- Bartoli, C.; Lamichhane, J.R.; Berge, O.; Varvaro, L.; Morris, C.E. Mutability in Pseudomonas viridiflava as a programmed balance between antibiotic resistance and pathogenicity. Mol. Plant. Pathol. 2015, 16, 860–869. [Google Scholar] [CrossRef] [PubMed]
- Balestra, G.M.; Mazzaglia, A.; Rossetti, A. Outbreak of bacterial blossom blight caused by Pseudomonas viridiflava on Actinidia chinensis kiwifruit plants in Italy. Plant. Dis. 2008, 92, 1707. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.K.M.; Hovmoller, M.S. Aerial dispersal of pathogens on the global and continental scales and its impact on plant desease. Science 2002, 297, 537–541. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Potential Human Pathogens | Spearman Correlation Coefficients | |
|---|---|---|
| Winter | Spring | |
| Enterobacter hormaechei | Enterobacter amnigenus (0.95 **) | Enterobacter amnigenus (0.79 **), Cr (0.74 *) |
| Enterobacter amnigenus | Enterobacter hormaechei (0.95 **) | Enterobacter hormaechei (0.79 **) |
| Enterobacter aerogenes | Acinetobacter ursingii (0.75 *), NH4+(0.71 *) | |
| Acinetobacter johnsonii | Acinetobacter ursingii (0.72 *), NH4+ (0.68 *), S (0.67 *) | |
| Acinetobacter ursingii | Acinetobacter johnsonii (0.72 *), Enterobacter aerogenes (0.75 *) | |
| Acinetobacter lwoffii | ||
| Providencia rettgeri | ||
| Propionibacterium acnes | Zn (0.68 *) | Propionibacterium avidum (0.93 **), Mg2+ (0.67*), MS− (0.74 *), Mo (0.74 *), Sr (0.66 *) |
| Propionibacterium avidum | Propionibacterium acnes (0.93 **), Clostridium cadaveris (0.71 *), MS− (0.70 *), Sr (0.83 **) | |
| Peptococcus niger | Na+ (0.66 *), T (0.70 *) | |
| Clostridium cadaveris | PM10 (0.82 **), As (0.75 *), Pb (0.67 *) | Streptococcus bovis (0.68 *), Propionibacterium avidum (0.71 *), Johnsonella ignava (0.87 **), MS− (0.66 *), V(0.68 *), Ca (0.67 *), Sr (0.72 *) |
| Staphylococcus aureus | ||
| Streptococcus bovis | Clostridium cadaveris (0.68 *), Johnsonella ignava (0.77 **) | |
| Johnsonella ignava | Clostridium cadaveris (0.87 **), Streptococcus bovis (0.77 **), PM10 (0.63 *), Ni (0.63 *), V (0.77 **), Ca (0.65 *), Sr (0.70 *) | |
| Sphingobacterium multivorum | ||
| Potential Plant Pathogens | Spearman Correlation Coefficients | |
|---|---|---|
| Winter | Spring | |
| Enterobacter cloacae | Enterobacter aerogenes (0.95 **), Acinetobacter johnsonii (0.65 *), Acinetobacter ursingii (0.85 **), NH4+ (0.65 *), WD (0.75 *) | |
| Pseudomonas viridiflava | Na+ (0.73 *), Cl− (0.75 *) | |
| Sphingomonas melonis | Al (0.65 *), Si (0.65 *), Ti (0.69 *), RH (0.67 *) | Clavibacter michiganensis (0.67 *), Mg2+ (0.79 **), T (0.64 *) |
| Janthinobacterium agaricidamnosum | WD (0.64 *) | |
| Erwinia mallotivora | Agrobacterium larrymoorei (0.67 *), Streptococcus bovis (0.74 *), Acinetobacter lwoffii (0.78 **), Cl− (0.75 *) | |
| Agrobacterium tumefaciens | Rathayibacter tritici (0.65 *), Propionibacterium acnes (0.63 *), Ni (0.66 *),V (0.78 **), WD (0.64 *), WS (0.64 *) | |
| Agrobacterium larrymoorei | Erwinia mallotivora (0.67 *), Propionibacterium avidum (0.68 *), Streptococcus bovis (0.70 *), Na+ (0.69 *), Cl− (0.75 *), NO3− (0.72 *), Mo (0.69 *) | |
| Curtobacterium flaccumfaciens | Clavibacter michiganensis (0.77 **), Propionibacterium avidum (0.70 *), WS (0.71 *) | |
| Clavibacter michiganensis | Curtobacterium flaccumfaciens (0.77 **), Sphingomonas melonis (0.67 *), T (0.65 *), WS (0.69 *) | |
| Rathayibacter tritici | Agrobacterium tumefaciens (0.65 *), Propionibacterium acnes (0.66 *), Clostridium cadaveris (0.83 **), Streptococcus bovis (0.65 *), Propionibacterium avidum (0.73 *), Johnsonella ignava (0.89 **), PM10 (0.72 *), MS− (0.71 *), Ni (0.80 **), V (0.91 **), Al (0.68 *), Si (0.71 *), Ca (0.70 *), Ti (0.69 *),Fe (0.68 *), Rb (0.66 *), Sr (0.77 **) | |
| Bacillus megaterium | ||
| Potential Non-Pathogenic Species | Spearman Correlation Coefficients | |
|---|---|---|
| Winter | Spring | |
| Calothrix parietina | Peptococcus niger (0.89 **) | Thiomonas thermosulfata (0.78 **), Sphingomonas oligophenolica (0.75 *), Acinetobacter lwoffii (0.66 *), Erwinia mallotivora (0.73 *) |
| Pseudomonas plecoglossicida | Pseudomonas entomophila (0.96 **), Arthrospira fusiformis (0.85 **), Pseudomonas putida (0.93 **), Enterobacter aerogenes (0.70 *) | Pseudomonas entomophila (0.82 **), Enterobacter aerogenes (0.80 **), Cd (0.70 *), Cr (0.72 *) |
| Stenotrophomonas geniculata | Staphylococcus aureus (0.71 *) | |
| Hyphomicrobium zavarzinii | Rhodococcus ruber (0.93 **) | Rhodococcus ruber (0.90 **) |
| Pseudomonas entomophila | Pseudomonas plecoglossicida (0.96 **), Arthrospira fusiformis (0.93 **), Chryseobacterium hispanicum (0.75 *), Pseudomonas putida (0.85 **), Acinetobacter johnsonii (0.63 *), Enterobacter aerogenes (0.80 **), Enterobacter cloacae (0.77 **) | |
| Thiomonas thermosulfata | Calothrix parietina (0.78 **), Sphingomonas oligophenolica (0.79 **), Propionibacterium acnes (0.78 **), Propionibacterium avidum (0.71 *), Mg2+ (0.79 **), Sr (0.66 *) | |
| Methylotenera mobilis | Clavibacter michiganensis (0.68 *), Mg2+ (0.70 *), Mo (0.69 *), Br (0.77 **), WS (0.65 *) | |
| Sphingomonas oligophenolica | Calothrix parietina (0.75 *), Thiomonas thermosulfata (0.79 **), Streptococcus bovis (0.84 **), Acinetobacter lwoffii (0.64 *), Johnsonella ignava (0.77 **), Erwinia mallotivora (0.66 *), Rathayibacter tritici (0.71 *), Sr (0.65 *) | |
| Arthrospira fusiformis | Pseudomonas plecoglossicida (0.85 **), Pseudomonas entomophila (0.93 **), Chryseobacterium hispanicum (0.90 **), Pseudomonas putida (0.73 *), Acinetobacter ursingii (0.77 **), Enterobacter aerogenes (0.87 **), Enterobacter cloacae (0.86 **) | Chryseobacterium hispanicum (0.90 **), Ba (0.82 **) |
| Chryseobacterium hispanicum | Pseudomonas entomophila (0.75 **), Arthrospira fusiformis (0.90 **), Acinetobacter johnsonii (0.67 *), Acinetobacter ursingii (0.89**), Enterobacter aerogenes (0.93 **), Enterobacter cloacae (0.93 **), Pb (0.65 *), P (0.64 *) | Arthrospira fusiformis (0.90 **), Ba (0.79 **), Cr (0.70 *) |
| Pseudomonas putida | Pseudomonas plecoglossicida (0.93 **), Pseudomonas entomophila (0.85 **), Arthrospira fusiformis (0.73 *), Enterobacter hormaechei (0.79 **), Enterobacter amnigenus (0.68 *) | |
| Rhodococcus ruber | Hyphomicrobium zavarzinii (0.93 **) | Hyphomicrobium zavarzinii (0.90 **), RH (0.74 *) |
| Bacillus badius | Sphingomonas melonis (0.71 *), Na+ (0.85 **), Mg2+ (0.78 **), Cl− (0.84 **), Al (0.86 **),Si(0.86 **), Ti(0.83 **), Br (0.67 *), WS (0.72 *) | |
| Bacillus aryabhattai | Bacillus megaterium (0.83 **) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Romano, S.; Fragola, M.; Alifano, P.; Perrone, M.R.; Talà, A. Potential Human and Plant Pathogenic Species in Airborne PM10 Samples and Relationships with Chemical Components and Meteorological Parameters. Atmosphere 2021, 12, 654. https://doi.org/10.3390/atmos12050654
Romano S, Fragola M, Alifano P, Perrone MR, Talà A. Potential Human and Plant Pathogenic Species in Airborne PM10 Samples and Relationships with Chemical Components and Meteorological Parameters. Atmosphere. 2021; 12(5):654. https://doi.org/10.3390/atmos12050654
Chicago/Turabian StyleRomano, Salvatore, Mattia Fragola, Pietro Alifano, Maria Rita Perrone, and Adelfia Talà. 2021. "Potential Human and Plant Pathogenic Species in Airborne PM10 Samples and Relationships with Chemical Components and Meteorological Parameters" Atmosphere 12, no. 5: 654. https://doi.org/10.3390/atmos12050654
APA StyleRomano, S., Fragola, M., Alifano, P., Perrone, M. R., & Talà, A. (2021). Potential Human and Plant Pathogenic Species in Airborne PM10 Samples and Relationships with Chemical Components and Meteorological Parameters. Atmosphere, 12(5), 654. https://doi.org/10.3390/atmos12050654