Reduction of Bioaerosols Emitted from a Swine Confinement Building by a Percolating Biofilter During a 10-Month Period


Abstract
1. Introduction
1.1. Potential Costs of Disease and Mitigation
1.2. Biofiltration of Air
2. Experiments
2.1. Air Treatment Unit Design and Operation
2.2. Air Sampling and Data Acquisition
2.3. Statistical Analysis
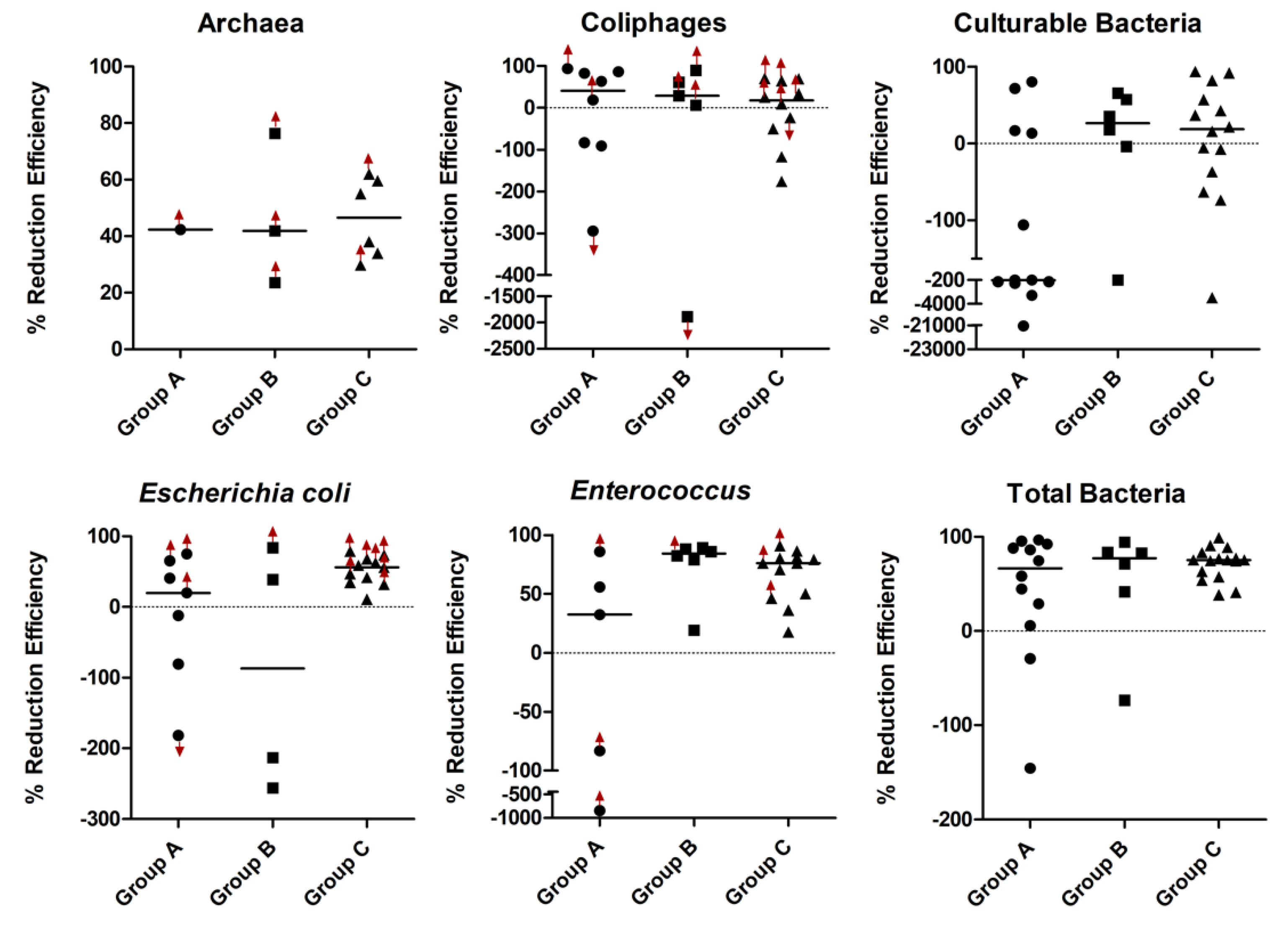
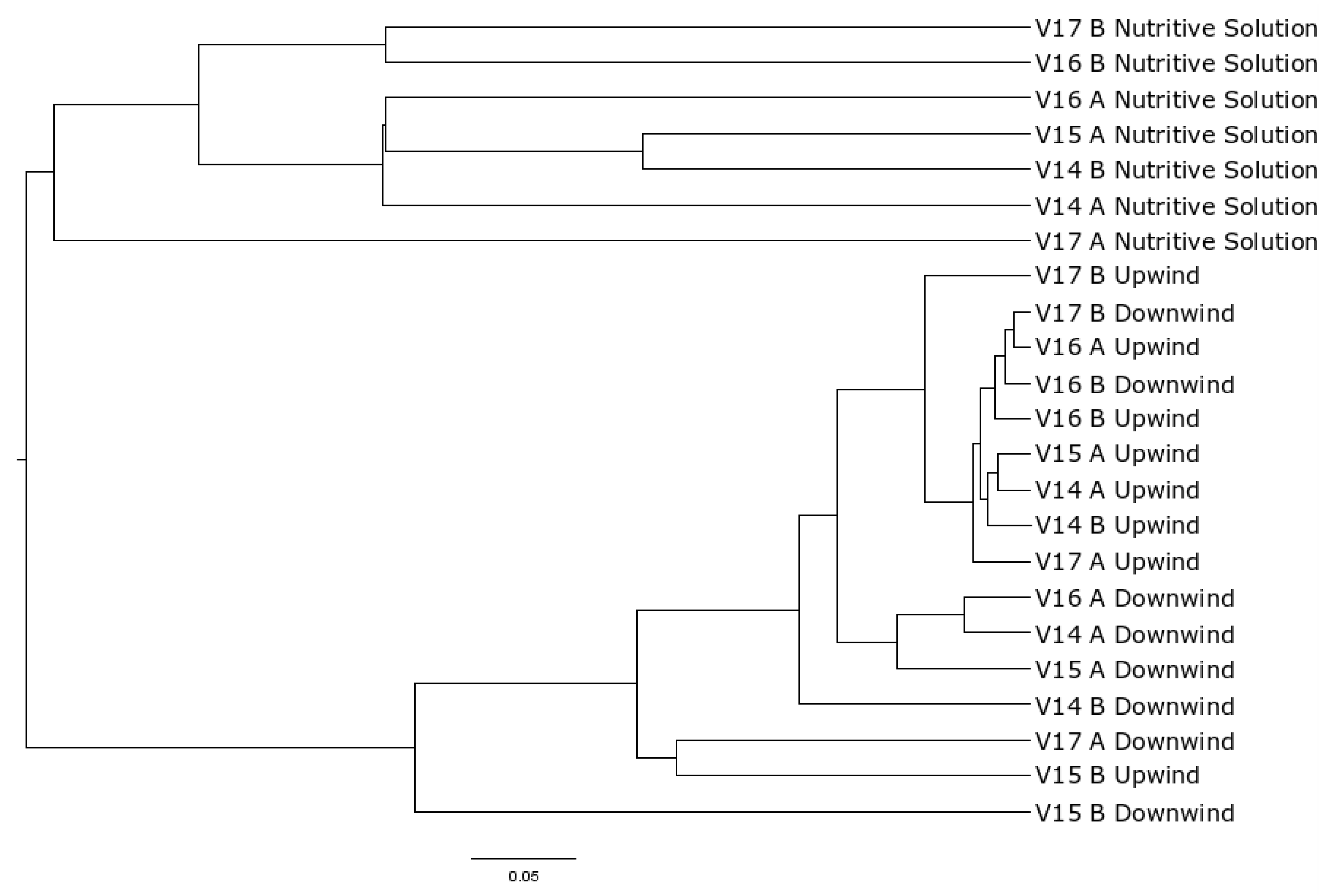
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Canadian Pork Council. Canadian Live Swine and Pork Exports. Available online: http://www.cpc-ccp.com/canadian-pork-and-live-swine-exports (accessed on 27 July 2018).
- Brisson, Y. The Changing Face of the Canadian Hog Industry; Statistics Canada: Ottawa, ON, Canada, 2014. [Google Scholar]
- Walker, J.T.; Aneja, V.P.; Dickey, D.A. Atmospheric transport and wet deposition of ammonium in North Carolina. Atmos. Environ. 2000, 34, 3407–3418. [Google Scholar] [CrossRef]
- Cambra-López, M.; Aarnink, A.J.; Zhao, Y.; Calvet, S.; Torres, A.G. Airborne particulate matter from livestock production systems: A review of an air pollution problem. Environ. Pollut. 2010, 158, 1–17. [Google Scholar] [CrossRef]
- Kristiansen, A.; Saunders, A.M.; Hansen, A.A.; Nielsen, P.H.; Nielsen, J.L. Community structure of bacteria and fungi in aerosols of a pig confinement building. FEMS Microbiol. Ecol. 2012, 80, 390–401. [Google Scholar] [CrossRef] [PubMed]
- Nehme, B.; Létourneau, V.; Forster, R.J.; Veillette, M.; Duchaine, C. Culture-independent approach of the bacterial bioaerosol diversity in the standard swine confinement buildings, and assessment of the seasonal effect. Environ. Microbiol. 2008, 10, 665–675. [Google Scholar] [CrossRef] [PubMed]
- Létourneau, V.; Nehmé, B.; Mériaux, A.; Massé, D.; Cormier, Y.; Duchaine, C. Human pathogens and tetracycline-resistant bacteria in bioaerosols of swine confinement buildings and in nasal flora of hog producers. Int. J. Hyg. Environ. Health 2010, 213, 444–449. [Google Scholar] [CrossRef]
- Bonifait, L.; Veillette, M.; Létourneau, V.; Grenier, D.; Duchaine, C. Detection of Streptococcus suis in bioaerosols of swine confinement buildings. Appl. Environ. Microbiol. 2014, 3296–3304. [Google Scholar] [CrossRef]
- Cole, D.; Todd, L.; Wing, S. Concentrated swine feeding operations and public health: A review of occupational and community health effects. Environ. Health Perspect. 2000, 108, 685–699. [Google Scholar] [CrossRef] [PubMed]
- Schiffman, S.S.; Miller, E.A.S.; Suggs, M.S.; Graham, B.G. The effect of environmental odors emanating from commercial swine operations on the mood of nearby residents. Brain Res. Bull. 1995, 37, 369–375. [Google Scholar] [CrossRef]
- Lim, T.-T.; Heber, A.J.; Ni, J.-Q.; Kendall, D.C.; Richert, B.T. Effects of manure removal strategies on odor and gas emission from swine finishing. Trans. ASAE 2004, 47, 2041–2050. [Google Scholar] [CrossRef]
- Girard, M.; Belzile, M.; Lemay, S.; Feddes, J.; Godbout, S. Innovative Air Treatment Unit for Swine Exhaust Air–Laboratory-Scale Tests. In Proceedings of the CSBE/SCGAB 2013 Annual Conference, Saskatoon, SK, Canada, 7–10 July 2013. [Google Scholar]
- Belzile, M.; Lemay, S.P.; Zegan, D.; Feddes, J.J.; Godbout, S.; Larouche, J.-P.; Martel, M. Reduction of gas and odour emissions from a swine building using a biotrickling filter. In Proceedings of the XVIIth World Congress of the International Commission of Agricultural Engineering, Québec City, QC, Canada, 13–17 June 2010. [Google Scholar]
- Morales, A.S.; Fragoso de Araújo, J.; de Moura Gomes, V.T.; Reis Costa, A.T.; Prazeres Rodrigues, D.D.; Porfida Ferreira, T.S.; de Lima Filsner, P.H.N.; Felizardo, M.R.; Micke Moreno, A. Colistin resistance in Escherichia coli and Salmonella enterica strains isolated from swine in Brazil. Sci. World J. 2012, 2012, 1–4. [Google Scholar] [CrossRef]
- Harada, K.; Asai, T.; Kojima, A.; Oda, C.; Ishihara, K.; Takahashi, T. Antimicrobial susceptibility of pathogenic Escherichia coli isolated from sick cattle and pigs in Japan. J. Vet. Med. Sci. 2005, 67, 999–1003. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kempf, I.; Jouy, E.; Chauvin, C. Colistin use and colistin resistance in bacteria from animals. Int. J. Antimicrob. Agents 2016, 48, 598–606. [Google Scholar] [CrossRef] [PubMed]
- Fairbrother, J.M.; Nadeau, É.; Gyles, C.L. Escherichia coli in postweaning diarrhea in pigs: An update on bacterial types, pathogenesis, and prevention strategies. Anim. Health Res. Rev. 2005, 6, 17–39. [Google Scholar] [CrossRef] [PubMed]
- Kolotilin, I.; Kaldis, A.; Devriendt, B.; Joensuu, J.; Cox, E.; Menassa, R. Production of a subunit vaccine candidate against porcine post-weaning diarrhea in high-biomass transplastomic tobacco. PLoS ONE 2012, 7, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Agriculture and Agri-Food Canada. Evaluation of the Control of Diseases in the Hog Industry. Available online: http://www.agr.gc.ca/eng/about-us/offices-and-locations/office-of-audit-and-evaluation/evaluation-reports/evaluation-of-the-control-of-diseases-in-the-hog-industry/?id=1435868137804 (accessed on 5 September 2019).
- Mussell, A.; Oginskyy, A.; Grier, K.; Morin, M.; Lachance, M.; Whittington, L.; Friendship, R. A Risk, Benefit, Strength, Weakness, Opportunity and Threat Analysis for the Control and Possible Eradication of Porcine Reproductive and Respiratory Syndrome (PRRS) Virus Within the Canadian Swine Herd; The George Morris Center: Guelph, ON, Canada, 2011. [Google Scholar]
- Holtkamp, D.J.; Kliebenstein, J.B.; Neumann, E.; Zimmerman, J.J.; Rotto, H.; Yoder, T.K.; Wang, C.; Yeske, P.; Mowrer, C.L.; Haley, C.A. Assessment of the economic impact of porcine reproductive and respiratory syndrome virus on United States pork producers. J. Swine Health Prod. 2013, 21, 72–84. [Google Scholar]
- Otake, S.; Dee, S.; Corzo, C.; Oliveira, S.; Deen, J. Long-distance airborne transport of infectious PRRSV and Mycoplasma hyopneumoniae from a swine population infected with multiple viral variants. Vet. Microbiol. 2010, 145, 198–208. [Google Scholar] [CrossRef] [PubMed]
- Alonso, C.; Goede, D.P.; Morrison, R.B.; Davies, P.R.; Rovira, A.; Marthaler, D.G.; Torremorell, M. Evidence of infectivity of airborne porcine epidemic diarrhea virus and detection of airborne viral RNA at long distances from infected herds. Vet. Res. 2014, 45, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Dee, S.; Spronk, G.; Reicks, D.; Ruen, P.; Deen, J. Further assessment of air filtration for preventing PRRSV infection in large breeding pig herds. Vet. Rec. 2010, 167, 976–977. [Google Scholar] [CrossRef] [PubMed]
- Dee, S.A.; Philips, R. Using vaccination and unidirectional pig flow to control PRRSV transmission. J. Swine Health Prod. 1998, 6, 21–25. [Google Scholar]
- Van der Heyden, C.; Demeyer, P.; Volcke, E.I. Mitigating emissions from pig and poultry housing facilities through air scrubbers and biofilters: State-of-the-art and perspectives. Biosyst. Eng. 2015, 134, 74–93. [Google Scholar] [CrossRef]
- Guo, H.; Dehod, W.; Agnew, J.; Laguë, C.; Feddes, J.; Pang, S. Annual odor emission rate from different types of swine production buildings. Trans. Asabe 2006, 49, 517–525. [Google Scholar] [CrossRef][Green Version]
- Martens, W.; Martinec, M.; Zapirain, R.; Stark, M.; Hartung, E.; Palmgren, U. Reduction potential of microbial, odour and ammonia emissions from a pig facility by biofilters. Int. J. Hyg. Environ. Health 2001, 203, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Cho, M.S.; Ko, H.J.; Kim, D.; Kim, K.Y. On-site application of air cleaner emitting plasma ion to reduce airborne contaminants in pig building. Atmos. Environ. 2012, 63, 276–281. [Google Scholar] [CrossRef]
- Aarnink, A.; Landman, W.; Melse, R.; Zhao, Y.; Ploegaert, J.; Huynh, T. Scrubber capabilities to remove airborne microorganisms and other aerial pollutants from the exhaust air of animal houses. Trans. ASABE 2011, 54, 1921–1930. [Google Scholar] [CrossRef]
- Lemay, S.P.; Girard, M.; Belzile, M.; Hogue, R.; Duchaine, C.; Létourneau, V.; Martel, M.; Jeanne, T.; Feldes, J.; Godbout, S. Un Concept Innovateur Pour Traiter L’air émis des Batiments porcins Réduisant L’impact Environnemental et Favorisant la Cohabitation; Rapport Final; IRDA: Québec, QC, Canada, 2012; p. 125. [Google Scholar]
- Walser, S.M.; Brenner, B.; Wunderlich, A.; Tuschak, C.; Huber, S.; Kolb, S.; Niessner, R.; Seidel, M.; Höller, C.; Herr, C.E. Detection of Legionella-contaminated aerosols in the vicinity of a bio-trickling filter of a breeding sow facility—A pilot study. Sci. Total Environ. 2017, 575, 1197–1202. [Google Scholar] [CrossRef] [PubMed]
- Kumari, P.; Choi, H.L. Seasonal variability in airborne biotic contaminants in swine confinement buildings. PLoS ONE 2014, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez de Evgrafov, M.; Kõll, P.; Frank, D.N.; Baumgartner, L.K.; Robertson, C.E.; Hernández, M.T.; Pace, N.R. Molecular analysis of bacterial and circovirus bioaerosols in concentrated animal feeding operations. Aerosol Sci. Technol. 2013, 47, 755–766. [Google Scholar] [CrossRef]
- Vestergaard, D.V.; Holst, G.J.; Basinas, I.; Elholm, G.; Schlünssen, V.; Linneberg, A.; Šantl-Temkiv, T.; Finster, K.; Sigsgaard, T.; Marshall, I.P.G. Pig farmers’ homes harbor more diverse airborne bacterial communities than pig stables or suburban homes. Front. Microbiol. 2018, 9, 870, 1–14. [Google Scholar] [CrossRef]
- Kumari, P.; Choi, H.L. Manure removal system influences the abundance and composition of airborne biotic contaminants in swine confinement buildings. Environ. Monit. Assess. 2015, 187, 1–10. [Google Scholar] [CrossRef]
- Hong, P.Y.; Li, X.; Yang, X.; Shinkai, T.; Zhang, Y.; Wang, X.; Mackie, R.I. Monitoring airborne biotic contaminants in the indoor environment of pig and poultry confinement buildings. Environ. Microbiol. 2012, 14, 1420–1431. [Google Scholar] [CrossRef]
- Pilote, J.; Létourneau, V.; Girard, M.; Duchaine, C. Quantification of airborne dust, endotoxins, human pathogens and antibiotic and metal resistance genes in Eastern Canadian swine confinement buildings. Aerobiologia 2019, 35, 283–296. [Google Scholar] [CrossRef]
- Mbareche, H.; Veillette, M.; Bilodeau, G.; Wösten, H.; Duchaine, C. Fungal cells recovery from air samples, a tale of loss and gain. Appl. Environ. Microbiol. 2019. under review. [Google Scholar] [CrossRef] [PubMed]
- Just, N.; Blais Lecours, P.; Marcoux-Voiselle, M.; Kirychuk, S.; Veillette, M.; Singh, B.; Duchaine, C. Archaeal characterization of bioaerosols from cage-housed and floor-housed poultry operations. Can. J. Microbiol. 2012, 59, 46–50. [Google Scholar] [CrossRef] [PubMed]
- Wise, M.G.; Siragusa, G.R. Quantitative detection of Clostridium perfringens in the broiler fowl gastrointestinal tract by real-time PCR. Appl. Environ. Microbiol. 2005, 71, 3911–3916. [Google Scholar] [CrossRef] [PubMed]
- Malinen, E.; Kassinen, A.; Rinttilä, T.; Palva, A. Comparison of real-time PCR with SYBR Green I or 5′-nuclease assays and dot-blot hybridization with rDNA-targeted oligonucleotide probes in quantification of selected faecal bacteria. Microbiology 2003, 149, 269–277. [Google Scholar] [CrossRef]
- Rinttilä, T.; Kassinen, A.; Malinen, E.; Krogius, L.; Palva, A. Development of an extensive set of 16S rDNA-targeted primers for quantification of pathogenic and indigenous bacteria in faecal samples by real-time PCR. J. Appl. Microbiol. 2004, 97, 1166–1177. [Google Scholar] [CrossRef]
- Bach, H.-J.; Tomanova, J.; Schloter, M.; Munch, J. Enumeration of total bacteria and bacteria with genes for proteolytic activity in pure cultures and in environmental samples by quantitative PCR mediated amplification. J. Microbiol. Methods 2002, 49, 235–245. [Google Scholar] [CrossRef]
- Reysenbach, A.; Pace, N.; Robb, F.; Place, A. Archaea: A laboratory manual—Thermophiles. Cold Spring Harb. Protoc 1995, 16, 101–107. [Google Scholar]
- Baker, G.; Smith, J.J.; Cowan, D.A. Review and re-analysis of domain-specific 16S primers. J. Microbiol. Methods 2003, 55, 541–555. [Google Scholar] [CrossRef]
- Gagnon, C.A.; Del Castillo, J.R.; Music, N.; Fontaine, G.; Harel, J.; Tremblay, D. Development and use of a multiplex real-time quantitative polymerase chain reaction assay for detection and differentiation of Porcine circovirus-2 genotypes 2a and 2b in an epidemiological survey. J. Vet. Diagn. Investig. 2008, 20, 545–558. [Google Scholar] [CrossRef]
- Joly, P.; Falconnet, P.-A.; André, J.; Weill, N.; Reyrolle, M.; Vandenesch, F.; Maurin, M.; Etienne, J.; Jarraud, S. Quantitative real-time Legionella PCR for environmental water samples: Data interpretation. Appl. Environ. Microbiol. 2006, 72, 2801–2808. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, K.; Ahmed, W.; Palmer, A.; Sidhu, J.; Hodgers, L.; Toze, S.; Haas, C. Public health implications of Acanthamoeba and multiple potential opportunistic pathogens in roof-harvested rainwater tanks. Environ. Res. 2016, 150, 320–327. [Google Scholar] [CrossRef] [PubMed]
- Friedman, S.D.; Cooper, E.M.; Calci, K.R.; Genthner, F.J. Design and assessment of a real time reverse transcription-PCR method to genotype single-stranded RNA male-specific coliphages (Family Leviviridae). J. Virol. Methods 2011, 173, 196–202. [Google Scholar] [CrossRef] [PubMed]
- Verreault, D.; Létourneau, V.; Gendron, L.; Massé, D.; Gagnon, C.A.; Duchaine, C. Airborne porcine circovirus in Canadian swine confinement buildings. Vet. Microbiol. 2010, 141, 224–230. [Google Scholar] [CrossRef] [PubMed]
- Comeau, A.M.; Li, W.K.; Tremblay, J.-É.; Carmack, E.C.; Lovejoy, C. Arctic Ocean microbial community structure before and after the 2007 record sea ice minimum. PLoS ONE 2011, 6, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef]
- Lévesque, A.; Girard, M.; Létourneau, V.; Pilote, J.; Duchaine, C.; Godbout, S.; Lemay, S.P. Achieving a Greater Reduction of Airborne Emissions from Swine Buildings by the Combination of Different Technologies. In Proceedings of the 2017 ASABE Annual International Meeting, Spokane, WA, USA, 16–19 July 2017; pp. 1–7. [Google Scholar]
- Amezcua, R.; Friendship, R.M.; Dewey, C.E.; Gyles, C.; Fairbrother, J.M. Presentation of postweaning Escherichia coli diarrhea in southern Ontario, prevalence of hemolytic E. coli serogroups involved, and their antimicrobial resistance patterns. Can. J. Vet. Res. 2002, 66, 73–78. [Google Scholar]
- DAF. Piglet Scours—General Information. Available online: https://www.daf.qld.gov.au/business-priorities/agriculture/animals/pigs/health-diseases/diseases/piglet-scours (accessed on 2 April 2019).
- Cornick, N.A.; VuKhac, H. Indirect transmission of Escherichia coli O157: H7 occurs readily among swine but not among sheep. Appl. Environ. Microbiol. 2008, 74, 2488–2491. [Google Scholar] [CrossRef]
- Gibbs, S.G.; Green, C.F.; Tarwater, P.M.; Mota, L.C.; Mena, K.D.; Scarpino, P.V. Isolation of antibiotic-resistant bacteria from the air plume downwind of a swine confined or concentrated animal feeding operation. Environ. Health Perspect. 2006, 114, 1032–1037. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Microorganisms | Primers and Probes (Final Concentration) | Sequence (5′–3′) | Thermoprotocol | References |
|---|---|---|---|---|
| Archaea | A751F (500 nM) | CCG ACG GTG AGR GRY GAA | 1) 95 °C for 5 min 2) 95 °C for 10 s 3) 55.5 °C for 20 s 4) 72 °C for 30 s Repeat 2)–4) 35 times 5) 72 °C for 10 min 6) 50–95 °C (0.5 °C increase per 2 s) | [44,45] |
| A976R (500 nM) | YCC GGC GTT GAM TCC AAT T | |||
| Bacteria | EUBF (625 nM) | GGT AGT CYA YGC MST AAA CG | 1) 94 °C for 3 min 2) 95 °C for 20 s 3) 62 °C for 60 s Repeat 2)–3) 40 times | [43] |
| EUBR (625 nM) | GAC ARCC ATG CAS CAC CTG | |||
| EUB probe (62.5 nM) | FAM-TKCGCGTTGCDTCGAATTAAWCCAC-TAMRA | |||
| Clostridium perfringens | CPerf165F (250 nM) | CGC ATA ACG TTG AAA GAT GG | 1) 95 °C for 3 min 2) 95 °C for 15 s 3) 57 °C for 30 s 4) 72 °C for 30 s Repeat 2)–4) 40 times 5) 72 °C for 10 min 6) 50–95 °C (0.5 °C increase per 2 s) | [40] |
| CPerf269R (250 nM) | CCT TGG TAG GCC GTT ACC C | |||
| Enterococcus | Ent1 (250 nM) | CCC TTA TTG TTA GTT GCC ATC ATT | [42] | |
| Ent2 (250 nM) | ACT CGT TGT ACT TCC CAT TGT | |||
| Escherichia coli | EC1 (300 nM) | GTT AAT ACC TTT GCT CAT TGA | 1) 95 °C for 3 min 2) 95 °C for 15 s 3) 55 °C for 30 s 4) 72 °C for 30 s Repeat 2)–4) 40 times 5) 72 °C for 10 min 6) 50–95 °C (0.5 °C increase per 2s) | [41] |
| EC2 (300 nM) | ACC AGG GTA TCT AAT CCT GTT | |||
| Legionella longbeachae | LLongF (500 nM) | AGA TGG GAT GTC TGG TGC TC | 1) 95 °C for 3 min 2) 95 °C for 15 s 3) 60 °C for 1 min Repeat 2)–3) 40 times | [48] |
| LlongR (500 nM) | ACC TGG TTT TGC ACC AGT TC | |||
| LlongProbe (50 nM) | 56-FAM/ACA AAG CAA/ZEN AAG GTG ACG CT/3IABkFQ | |||
| Legionella pneumophila | LpneuF (500 nM) | GCA TTG GTG CCG ATT TGG | 1) 95 °C for 5 min 2) 95 °C for 15 s 3) 57 °C for 10 s Repeat 2)–3) 45 times | [47] |
| LpneuR (500 nM) | GYT TTG CCA TCA AAT CTT TCT GAA | |||
| LpneuProbe (50 nM) | 56-FAM/CCA CTC ATA/ZEN/ GCG TCT TGC ATG CCT TTA/3IABkFQ | |||
| Coliphages | Colip-F (250 nM) | ATCCATTTTGGTAACGCCG | 1) 95 °C for 3 min 2) 94 °C for 15 s 3) 60 °C for 30 s Repeat 2)–3) 40 times | [49] |
| Colip-R (250 nM) | TGCAATCTCACTGGGACATAT | |||
| Colip-Probe (50 nM) | TAGGCATCTACGGGGACGA | |||
| Porcine Circovirus | Circo-Gen-F (400 nM) | GGC CAC CTG GGT GTG GTA AA | 1) 95 °C for 3 min 2) 95 °C for 30 s 3) 60 °C for 60 s 4) 72 °C for 60 s Repeat 2)–4) 45 times | [46] |
| Circo-Gen-R (400 nM) | CCC ACC ACT TGT TTC TAG GTG GTT | |||
| Circo-Gen-Probe (120 nM) | 56-FAM/TTT GCA GAC CCG GAA ACC ACA TAC TGG A/3BHQ |
| Visit. | Unit | Air Flow (CFM) | EBRT (s) | Sampling Rate (L/min) | Relative Humidity (%) | Temperature Before (°C) | Temperature After (°C) |
|---|---|---|---|---|---|---|---|
| V1 (26/5/2017) | A | 1540 | 8.4 | 150 | 64.0 | 20.0 | 19.0 |
| B | 1090 | 11.9 | 150 | 63.0 | 20.0 | 20.0 | |
| V2 (21/6/2017) | A | 1350 | 9.6 | 100 | 63.5 | 22.2 | 20.5 |
| B | 1160 | 11.2 | 100 | 64.5 | 23.1 | 22.6 | |
| V3 (5/7/2017) | A | 1400 | 9.3 | 150 | 45.0 | 25.5 | 21.0 |
| B | 1170 | 11.1 | 100 | 51.0 | 26.7 | 23.5 | |
| V4 (17/7/2017) | A | 1350 | 9.6 | 150 | 63.1 | 26.2 | 23.4 |
| B | 1240 | 10.5 | 100 | 68.9 | 27.4 | 25.5 | |
| V5 (3/8/2017) | A | 1390 | 9.3 | 150 | 64.5 | 24.7 | 22.2 |
| B | 1370 | 9.5 | 150 | 67.1 | 25.2 | 23.7 | |
| V6 (14/8/2017) | A | 1350 | 9.6 | 150 | 61.7 | 24.6 | 21.1 |
| B | 1230 | 10.5 | 100 | 66.2 | 23.4 | 22.2 | |
| V7 (30/8/2017) | A | 3375 | 3.8 | 300 | 48.9 | 26.2 | 21.0 |
| B | 3090 | 4.2 | 250 | 51.5 | 25.5 | 21.4 | |
| V8 (13/9/2017) | A | 3325 | 3.9 | 300 | 50.5 | 27.2 | 21.7 |
| B | 3080 | 4.2 | 250 | 54.7 | 24.6 | 21.9 | |
| V9 (28/9/2017) | A | 3300 | 3.9 | 300 | 54.7 | 22.5 | 19.5 |
| B | 3050 | 4.2 | 250 | 57.8 | 21.5 | 19.0 | |
| V10 (17/10/2017) | A | 1130 | 11.5 | 100 | 56.0 | 20.4 | 17.0 |
| B | 490 | 26.4 | 150 | 59.0 | 19.2 | 15.0 | |
| V11 (3/11/2017) | A | 1100 | 11.8 | 100 | 58.0 | 21.7 | 20.0 |
| B | 540 | 24.0 | 200 | 59.3 | 21.4 | 22.0 | |
| V12 (18/12/2017) | A | 255 | 50.8 | 100 | 80.3 | 11.6 | 16.7 |
| B | 770 | 16.8 | 250 | 79.9 | 15.6 | 17.5 | |
| V13 (25/1/2018) | A | 385 | 33.7 | 150 | 88.8 | 13.8 | 17.6 |
| B | 460 | 28.2 | 200 | 94.4 | 13.3 | 16.7 | |
| V14 (14/2/2018) | A | 290 | 44.7 | 100 | 87.5 | 14.2 | 16.8 |
| B | 870 | 14.9 | 300 | 92.5 | 15.1 | 17.0 | |
| V15 (7/3/2018) | A | 575 | 22.5 | 150 | 82.0 | 15.2 | 17.3 |
| B | 1263 | 10.3 | 150 | 83.5 | 16.9 | 17.5 | |
| V16 (23/3/2018) | A | 455 | 28.5 | 150 | 81.5 | 15.8 | 17.2 |
| B | 1420 | 9.1 | 150 | 71.0 | 19.2 | 18.1 | |
| V17 (13/4/2018) | A | 285 | 45.5 | 100 | 83.0 | 16.8 | 19.0 |
| B | 1690 | 7.7 | 150 | 55.8 | 19.6 | 17.5 |
| Percent Reduction Efficiency (%) | |||||||
|---|---|---|---|---|---|---|---|
| Group | AirFlow Rate (CFM) | Total Bacteria | Culturable | Enterococcus | E. coli | Archaea | Coliphages |
| A | 1350 | 66 (66) | −218 (559) | 32 (139) | 20 (99) | 42 (0) | 40.8 (168.2) |
| B | 3195 | 77 (34) | 26 (50) | 84 (8) | −87 (274) | 42 (26) | 28.8 (54.5) |
| C | 557.5 | 75 (17) | 29 (96) | 76 (31) | 56 (27) | 47 (23) | 17.6 (100.4) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vyskocil, J.M.; Létourneau, V.; Girard, M.; Lévesque, A.; Duchaine, C. Reduction of Bioaerosols Emitted from a Swine Confinement Building by a Percolating Biofilter During a 10-Month Period. Atmosphere 2019, 10, 525. https://doi.org/10.3390/atmos10090525
Vyskocil JM, Létourneau V, Girard M, Lévesque A, Duchaine C. Reduction of Bioaerosols Emitted from a Swine Confinement Building by a Percolating Biofilter During a 10-Month Period. Atmosphere. 2019; 10(9):525. https://doi.org/10.3390/atmos10090525
Chicago/Turabian StyleVyskocil, Jonathan M, Valérie Létourneau, Matthieu Girard, Ariane Lévesque, and Caroline Duchaine. 2019. "Reduction of Bioaerosols Emitted from a Swine Confinement Building by a Percolating Biofilter During a 10-Month Period" Atmosphere 10, no. 9: 525. https://doi.org/10.3390/atmos10090525
APA StyleVyskocil, J. M., Létourneau, V., Girard, M., Lévesque, A., & Duchaine, C. (2019). Reduction of Bioaerosols Emitted from a Swine Confinement Building by a Percolating Biofilter During a 10-Month Period. Atmosphere, 10(9), 525. https://doi.org/10.3390/atmos10090525