Membrane-Associated Proteins in Giardia lamblia

Abstract
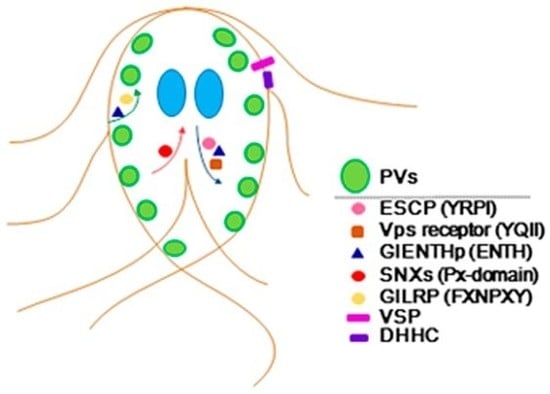
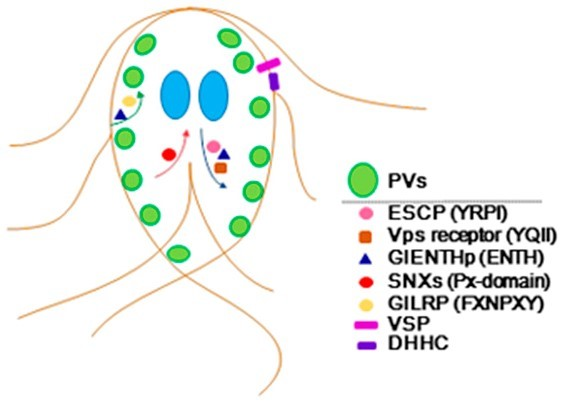
:
1. Introduction
2. Fat Attachment: Phosphoinositide-Binding Proteins
3. Take Me Home: Integral Protein Targeting
4. The Ultimate Fate: Default Targeting to the Plasma Membrane
5. Conclusions and Future Directions
Funding
Conflicts of Interest
References
- Adl, S.M.; Simpson, A.G.; Farmer, M.A.; Andersen, R.A.; Anderson, O.R.; Barta, J.R.; Bowser, S.S.; Brugerolle, G.; Fensome, R.A.; Fredericq, S.; et al. The new higher level classification of eukaryotes with emphasis on the taxonomy of protists. J. Eukaryot. Microbiol. 2005, 52, 399–451. [Google Scholar] [CrossRef] [PubMed]
- Ankarklev, J.; Jerlstrom-Hultqvist, J.; Ringqvist, E.; Troell, K.; Svard, S.G. Behind the smile: Cell biology and disease mechanisms of Giardia species. Nat. Rev. Microbiol. 2010, 8, 413–422. [Google Scholar] [CrossRef] [PubMed]
- Muller, J.; Ley, S.; Felger, I.; Hemphill, A.; Muller, N. Identification of differentially expressed genes in a Giardia lamblia WB C6 clone resistant to nitazoxanide and metronidazole. J. Antimicrob. Chemother. 2008, 62, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Soltys, B.J.; Falah, M.; Gupta, R.S. Identification of endoplasmic reticulum in the primitive eukaryote Giardia lamblia using cryoelectron microscopy and antibody to bip. J. Cell Sci. 1996, 109 Pt 7, 1909–1917. [Google Scholar]
- Tovar, J.; Leon-Avila, G.; Sanchez, L.B.; Sutak, R.; Tachezy, J.; van der Giezen, M.; Hernandez, M.; Muller, M.; Lucocq, J.M. Mitochondrial remnant organelles of Giardia function in iron-sulphur protein maturation. Nature 2003, 426, 172–176. [Google Scholar] [CrossRef] [PubMed]
- Lanfredi-Rangel, A.; Attias, M.; de Carvalho, T.M.; Kattenbach, W.M.; De Souza, W. The peripheral vesicles of trophozoites of the primitive protozoan Giardia lamblia may correspond to early and late endosomes and to lysosomes. J. Struct. Biol. 1998, 123, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Touz, M.C.; Zamponi, N. Sorting without a Golgi complex. Traffic 2017, 18, 637–645. [Google Scholar] [CrossRef] [PubMed]
- Yichoy, M.; Duarte, T.T.; De Chatterjee, A.; Mendez, T.L.; Aguilera, K.Y.; Roy, D.; Roychowdhury, S.; Aley, S.B.; Das, S. Lipid metabolism in giardia: A post-genomic perspective. Parasitology 2011, 138, 267–278. [Google Scholar] [CrossRef] [PubMed]
- Yichoy, M.; Nakayasu, E.S.; Shpak, M.; Aguilar, C.; Aley, S.B.; Almeida, I.C.; Das, S. Lipidomic analysis reveals that phosphatidylglycerol and phosphatidylethanolamine are newly generated phospholipids in an early-divergent protozoan, Giardia lamblia. Mol. Biochem. Parasitol. 2009, 165, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Behnia, R.; Munro, S. Organelle identity and the signposts for membrane traffic. Nature 2005, 438, 597–604. [Google Scholar] [CrossRef] [PubMed]
- Di Paolo, G.; De Camilli, P. Phosphoinositides in cell regulation and membrane dynamics. Nature 2006, 443, 651–657. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, Y.; Zamora, G.; Ray, S.; Chapoy, J.; Chavez, E.; Valvarde, R.; Williams, E.; Aley, S.B.; Das, S. Transcriptional analysis of three major putative phosphatidylinositol kinase genes in a parasitic protozoan, Giardia lamblia. J. Eukaryot. Microbiol. 2007, 54, 29–32. [Google Scholar] [CrossRef] [PubMed]
- Cox, S.S.; van der Giezen, M.; Tarr, S.J.; Crompton, M.R.; Tovar, J. Evidence from bioinformatics, expression and inhibition studies of phosphoinositide-3 kinase signalling in Giardia intestinalis. BMC Microbiol. 2006, 6, 45. [Google Scholar] [CrossRef] [PubMed]
- Sinha, A.; Mandal, S.; Banerjee, S.; Ghosh, A.; Ganguly, S.; Sil, A.K.; Sarkar, S. Identification and characterization of a fyve domain from the early diverging eukaryote Giardia lamblia. Curr. Microbiol. 2011, 62, 1179–1184. [Google Scholar] [CrossRef] [PubMed]
- Gillooly, D.J.; Morrow, I.C.; Lindsay, M.; Gould, R.; Bryant, N.J.; Gaullier, J.M.; Parton, R.G.; Stenmark, H. Localization of phosphatidylinositol 3-phosphate in yeast and mammalian cells. EMBO J. 2000, 19, 4577–4588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Itoh, T.; Koshiba, S.; Kigawa, T.; Kikuchi, A.; Yokoyama, S.; Takenawa, T. Role of the ENTH domain in phosphatidylinositol-4,5-bisphosphate binding and endocytosis. Science 2001, 291, 1047–1051. [Google Scholar] [CrossRef] [PubMed]
- Hirst, J.; Motley, A.; Harasaki, K.; Peak Chew, S.Y.; Robinson, M.S. EpsinR: An ENTH domain-containing protein that interacts with AP-1. Mol. Biol. Cell 2003, 14, 625–641. [Google Scholar] [CrossRef] [PubMed]
- Feliziani, C.; Zamponi, N.; Gottig, N.; Ropolo, A.S.; Lanfredi-Rangel, A.; Touz, M.C. The giardial ENTH protein participates in lysosomal protein trafficking and endocytosis. Biochim. Biophys. Acta 2015, 1853, 646–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebneter, J.A.; Hehl, A.B. The single epsin homolog in Giardia lamblia localizes to the ventral disk of trophozoites and is not associated with clathrin membrane coats. Mol. Biochem. Parasitol. 2014, 197, 24–27. [Google Scholar] [CrossRef] [PubMed]
- Odorizzi, G.; Babst, M.; Emr, S.D. Phosphoinositide signaling and the regulation of membrane trafficking in yeast. Trends Biochem. Sci. 2000, 25, 229–235. [Google Scholar] [CrossRef]
- Worby, C.A.; Dixon, J.E. Sorting out the cellular functions of sorting nexins. Nat. Rev. Mol. Cell Biol. 2002, 3, 919–931. [Google Scholar] [CrossRef] [PubMed]
- Karathanassis, D.; Stahelin, R.V.; Bravo, J.; Perisic, O.; Pacold, C.M.; Cho, W.; Williams, R.L. Binding of the PX domain of p47(phox) to phosphatidylinositol 3,4-bisphosphate and phosphatidic acid is masked by an intramolecular interaction. EMBO J. 2002, 21, 5057–5068. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Liu, D.; Gill, G.; Songyang, Z. Regulation of cytokine-independent survival kinase (CISK) by the Phox homology domain and phosphoinositides. J. Cell Biol. 2001, 154, 699–705. [Google Scholar] [CrossRef] [PubMed]
- Seet, L.F.; Hong, W. The phox (PX) domain proteins and membrane traffic. Biochim. Biophys. Acta 2006, 1761, 878–896. [Google Scholar] [CrossRef] [PubMed]
- Jana, A.; Sinha, A.; Sarkar, S. Phosphoinositide binding profiles of the PX domains of Giardia lamblia. Parasitol. Int. 2017, 66, 606–614. [Google Scholar] [CrossRef] [PubMed]
- Teasdale, R.D.; Loci, D.; Houghton, F.; Karlsson, L.; Gleeson, P.A. A large family of endosome-localized proteins related to sorting nexin 1. Biochem. J. 2001, 358, 7–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallon, M.; Cullen, P.J. Retromer and sorting nexins in endosomal sorting. Biochem. Soc. Trans. 2015, 43, 33–47. [Google Scholar] [CrossRef] [PubMed]
- Bonifacino, J.S.; Hurley, J.H. Retromer. Curr. Opin. Cell Biol. 2008, 20, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Harterink, M.; Port, F.; Lorenowicz, M.J.; McGough, I.J.; Silhankova, M.; Betist, M.C.; van Weering, J.R.; van Heesbeen, R.G.; Middelkoop, T.C.; Basler, K.; et al. A SNX3-dependent retromer pathway mediates retrograde transport of the Wnt sorting receptor Wntless and is required for Wnt secretion. Nat. Cell Biol. 2011, 13, 914–923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, S.A.; Barr, V.A.; Haft, D.H.; Taylor, S.I.; Haft, C.R. Identification and characterization of SNX15, a novel sorting nexin involved in protein trafficking. J. Biol. Chem. 2001, 276, 5074–5084. [Google Scholar] [CrossRef] [PubMed]
- Van Kerkhof, P.; Lee, J.; McCormick, L.; Tetrault, E.; Lu, W.; Schoenfish, M.; Oorschot, V.; Strous, G.J.; Klumperman, J.; Bu, G. Sorting nexin 17 facilitates LRP recycling in the early endosome. EMBO J. 2005, 24, 2851–2861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miras, S.L.; Merino, M.C.; Gottig, N.; Ropolo, A.S.; Touz, M.C. The giardial VPS35 retromer subunit is necessary for multimeric complex assembly and interaction with the vacuolar protein sorting receptor. Biochim. Biophys. Acta 2013, 1833, 2628–2638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheever, M.L.; Sato, T.K.; de Beer, T.; Kutateladze, T.G.; Emr, S.D.; Overduin, M. Phox domain interaction with Ptdins(3)P targets the Vam7 t-SNARE to vacuole membranes. Nat. Cell Biol. 2001, 3, 613–618. [Google Scholar] [CrossRef] [PubMed]
- Elias, E.V.; Quiroga, R.; Gottig, N.; Nakanishi, H.; Nash, T.E.; Neiman, A.; Lujan, H.D. Characterization of SNARES determines the absence of a typical Golgi apparatus in the ancient eukaryote Giardia lamblia. J. Biol. Chem. 2008, 283, 35996–36010. [Google Scholar] [CrossRef] [PubMed]
- Settembre, C.; Fraldi, A.; Medina, D.L.; Ballabio, A. Signals from the lysosome: A control centre for cellular clearance and energy metabolism. Nat. Rev. Mol. Cell Biol. 2013, 14, 283–296. [Google Scholar] [CrossRef] [PubMed]
- Blott, E.J.; Griffiths, G.M. Secretory lysosomes. Nat. Rev. Mol. Cell Biol. 2002, 3, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Rivero, M.R.; Jausoro, I.; Bisbal, M.; Feliziani, C.; Lanfredi-Rangel, A.; Touz, M.C. Receptor-mediated endocytosis and trafficking between endosomal-lysosomal vacuoles in Giardia lamblia. Parasitol. Res. 2013, 112, 1813–1818. [Google Scholar] [CrossRef] [PubMed]
- Rivero, M.R.; Vranych, C.V.; Bisbal, M.; Maletto, B.A.; Ropolo, A.S.; Touz, M.C. Adaptor protein 2 regulates receptor-mediated endocytosis and cyst formation in Giardia lamblia. Biochem. J. 2010, 428, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Bonifacino, J.S.; Lippincott-Schwartz, J. Coat proteins: Shaping membrane transport. Nat. Rev. Mol. Cell Biol. 2003, 4, 409–414. [Google Scholar] [CrossRef] [PubMed]
- Janvier, K.; Kato, Y.; Boehm, M.; Rose, J.R.; Martina, J.A.; Kim, B.Y.; Venkatesan, S.; Bonifacino, J.S. Recognition of dileucine-based sorting signals from HIV-1 NEF and LIMP-II by the AP-1 γ-σ1 and AP-3 δ- σ3 hemicomplexes. J. Cell Biol. 2003, 163, 1281–1290. [Google Scholar] [CrossRef] [PubMed]
- Doray, B.; Lee, I.; Knisely, J.; Bu, G.; Kornfeld, S. The γ/σ1 and α/σ2 hemicomplexes of clathrin adaptors AP-1 and AP-2 harbor the dileucine recognition site. Mol. Biol. Cell 2007, 18, 1887–1896. [Google Scholar] [CrossRef] [PubMed]
- Touz, M.C.; Lujan, H.D.; Hayes, S.F.; Nash, T.E. Sorting of encystation-specific cysteine protease to lysosome-like peripheral vacuoles in Giardia lamblia requires a conserved tyrosine-based motif. J. Biol. Chem. 2003, 278, 6420–6426. [Google Scholar] [CrossRef] [PubMed]
- Touz, M.C.; Kulakova, L.; Nash, T.E. Adaptor protein complex 1 mediates the transport of lysosomal proteins from a Golgi-like organelle to peripheral vacuoles in the primitive eukaryote Giardia lamblia. Mol. Biol. Cell 2004, 15, 3053–3060. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, P.; Dahms, N.M.; Kornfeld, S. Mannose 6-phosphate receptors: New twists in the tale. Nat. Rev. Mol. Cell Biol. 2003, 4, 202–212. [Google Scholar] [CrossRef] [PubMed]
- Rivero, M.R.; Miras, S.L.; Feliziani, C.; Zamponi, N.; Quiroga, R.; Hayes, S.F.; Ropolo, A.S.; Touz, M.C. Vacuolar protein sorting receptor in Giardia lamblia. PLoS ONE 2012, 7, e43712. [Google Scholar] [CrossRef] [PubMed]
- Rivero, M.R.; Miras, S.L.; Quiroga, R.; Ropolo, A.S.; Touz, M.C. Giardia lamblia low-density lipoprotein receptor-related protein is involved in selective lipoprotein endocytosis and parasite replication. Mol. Microbiol. 2011, 79, 1204–1219. [Google Scholar] [CrossRef] [PubMed]
- Martincova, E.; Voleman, L.; Pyrih, J.; Zarsky, V.; Vondrackova, P.; Kolisko, M.; Tachezy, J.; Dolezal, P. Probing the biology of Giardia intestinalis mitosomes using in vivo enzymatic tagging. Mol. Cell. Biol. 2015, 35, 2864–2874. [Google Scholar] [CrossRef] [PubMed]
- Touz, M.C.; Conrad, J.T.; Nash, T.E. A novel palmitoyl acyl transferase controls surface protein palmitoylation and cytotoxicity in Giardia lamblia. Mol. Microbiol. 2005, 58, 999–1011. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Saraiya, A.A.; Wang, C.C. Experimental verification of the identity of variant-specific surface proteins in Giardia lamblia trophozoites. mBio 2013, 4, e00321-13. [Google Scholar] [CrossRef] [PubMed]
- Mowatt, M.R.; Aggarwal, A.; Nash, T.E. Carboxy-terminal sequence conservation among variant-specific surface proteins of Giardia lamblia. Mol. Biochem. Parasitol. 1991, 49, 215–227. [Google Scholar] [CrossRef]
- Adam, R.D. Biology of Giardia lamblia. Clin. Microbiol. Rev. 2001, 14, 447–475. [Google Scholar] [CrossRef] [PubMed]
- Nash, T.E.; Mowatt, M.R. Characterization of a Giardia lamblia variant-specific surface protein (VSP) gene from isolate GS/M and estimation of the VSP gene repertoire size. Mol. Biochem. Parasitol. 1992, 51, 219–227. [Google Scholar] [CrossRef]
- Aley, S.B.; Gillin, F.D. Specialized surface adaptations of Giardia lamblia. Infect. Agents Dis. 1995, 4, 161–166. [Google Scholar] [PubMed]
- Svard, S.G.; Meng, T.C.; Hetsko, M.L.; McCaffery, J.M.; Gillin, F.D. Differentiation-associated surface antigen variation in the ancient eukaryote Giardia lamblia. Mol. Microbiol. 1998, 30, 979–989. [Google Scholar] [CrossRef] [PubMed]
- Nash, T.E.; Banks, S.M.; Alling, D.W.; Merritt, J.W., Jr.; Conrad, J.T. Frequency of variant antigens in Giardia lamblia. Exp. Parasitol. 1990, 71, 415–421. [Google Scholar] [CrossRef]
- Nash, T.E.; Herrington, D.A.; Levine, M.M.; Conrad, J.T.; Merritt, J.W., Jr. Antigenic variation of Giardia lamblia in experimental human infections. J. Immunol. 1990, 144, 4362–4369. [Google Scholar] [PubMed]
- Nash, T. Surface antigen variability and variation in Giardia lamblia. Parasitol. Today 1992, 8, 229–234. [Google Scholar] [CrossRef]
- Gottstein, B.; Harriman, G.R.; Conrad, J.T.; Nash, T.E. Antigenic variation in Giardia lamblia: Cellular and humoral immune response in a mouse model. Parasite Immunol. 1990, 12, 659–673. [Google Scholar] [CrossRef] [PubMed]
- Papanastasiou, P.; Hiltpold, A.; Bommeli, C.; Kohler, P. The release of the variant surface protein of Giardia to its soluble isoform is mediated by the selective cleavage of the conserved carboxy-terminal domain. Biochemistry 1996, 35, 10143–10148. [Google Scholar] [CrossRef] [PubMed]
- Marti, M.; Li, Y.; Schraner, E.M.; Wild, P.; Kohler, P.; Hehl, A.B. The secretory apparatus of an ancient eukaryote: Protein sorting to separate export pathways occurs before formation of transient Golgi-like compartments. Mol. Biol. Cell 2003, 14, 1433–1447. [Google Scholar] [CrossRef] [PubMed]
- Touz, M.C.; Ropolo, A.S.; Rivero, M.R.; Vranych, C.V.; Conrad, J.T.; Svard, S.G.; Nash, T.E. Arginine deiminase has multiple regulatory roles in the biology of Giardia lamblia. J. Cell Sci. 2008, 121, 2930–2938. [Google Scholar] [CrossRef] [PubMed]
- Papanastasiou, P.; McConville, M.J.; Ralton, J.; Kohler, P. The variant-specific surface protein of Giardia, VSP4A1, is a glycosylated and palmitoylated protein. Biochem. J. 1997, 322 Pt 1, 49–56. [Google Scholar] [CrossRef]
- Hiltpold, A.; Frey, M.; Hulsmeier, A.; Kohler, P. Glycosylation and palmitoylation are common modifications of Giardia variant surface proteins. Mol. Biochem. Parasitol. 2000, 109, 61–65. [Google Scholar] [CrossRef]
- Carranza, P.G.; Feltes, G.; Ropolo, A.; Quintana, S.M.; Touz, M.C.; Lujan, H.D. Simultaneous expression of different variant-specific surface proteins in single Giardia lamblia trophozoites during encystation. Infect. Immun. 2002, 70, 5265–5268. [Google Scholar] [CrossRef] [PubMed]
- Davids, B.J.; Reiner, D.S.; Birkeland, S.R.; Preheim, S.P.; Cipriano, M.J.; McArthur, A.G.; Gillin, F.D. A new family of giardial cysteine-rich non-VSP protein genes and a novel cyst protein. PLoS ONE 2006, 1, e44. [Google Scholar] [CrossRef] [PubMed]
- Merino, M.C.; Zamponi, N.; Vranych, C.V.; Touz, M.C.; Ropolo, A.S. Identification of Giardia lamblia DHHC proteins and the role of protein S-palmitoylation in the encystation process. PLoS Negl. Trop. Dis. 2014, 8, e2997. [Google Scholar] [CrossRef] [PubMed]
- Bartels, D.J.; Mitchell, D.A.; Dong, X.; Deschenes, R.J. Erf2, a novel gene product that affects the localization and palmitoylation of Ras2 in Saccharomyces cerevisiae. Mol. Cell. Biol. 1999, 19, 6775–6787. [Google Scholar] [CrossRef] [PubMed]
- Lobo, S.; Greentree, W.K.; Linder, M.E.; Deschenes, R.J. Identification of a Ras palmitoyltransferase in Saccharomyces cerevisiae. J. Biol. Chem. 2002, 277, 41268–41273. [Google Scholar] [CrossRef] [PubMed]
- Linder, M.E.; Jennings, B.C. Mechanism and function of DHHC S-acyltransferases. Biochem. Soc. Trans. 2013, 41, 29–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coelho, C.H.; Costa, A.O.; Silva, A.C.; Pucci, M.M.; Serufo, A.V.; Busatti, H.G.; Durigan, M.; Perales, J.; Chapeaurouge, A.; de Silva a Silva, D.A.; et al. Genotyping and descriptive proteomics of a potential zoonotic canine strain of Giardia duodenalis, infective to mice. PLoS ONE 2016, 11, e0164946. [Google Scholar] [CrossRef] [PubMed]
- Emery, S.J.; Lacey, E.; Haynes, P.A. Quantitative proteomics in Giardia duodenalis-achievements and challenges. Mol. Biochem. Parasitol. 2016, 208, 96–112. [Google Scholar] [CrossRef] [PubMed]
- Wampfler, P.B.; Tosevski, V.; Nanni, P.; Spycher, C.; Hehl, A.B. Proteomics of secretory and endocytic organelles in Giardia lamblia. PLoS ONE 2014, 9, e94089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jerlstrom-Hultqvist, J.; Ankarklev, J.; Svard, S.G. Is human giardiasis caused by two different Giardia species? Gut Microbes 2011, 1, 379–382. [Google Scholar] [CrossRef] [PubMed]
- Franzen, O.; Jerlstrom-Hultqvist, J.; Castro, E.; Sherwood, E.; Ankarklev, J.; Reiner, D.S.; Palm, D.; Andersson, J.O.; Andersson, B.; Svard, S.G. Draft genome sequencing of Giardia intestinalis assemblage B isolate GS: Is human giardiasis caused by two different species? PLoS Pathog. 2009, 5, e1000560. [Google Scholar] [CrossRef] [PubMed]
- Singer, S.M.; Yee, J.; Nash, T.E. Episomal and integrated maintenance of foreign DNA in Giardia lamblia. Mol. Biochem. Parasitol. 1998, 92, 59–69. [Google Scholar] [CrossRef]
- Kulakova, L.; Singer, S.M.; Conrad, J.; Nash, T.E. Epigenetic mechanisms are involved in the control of Giardia lamblia antigenic variation. Mol. Microbiol. 2006, 61, 1533–1542. [Google Scholar] [CrossRef] [PubMed]
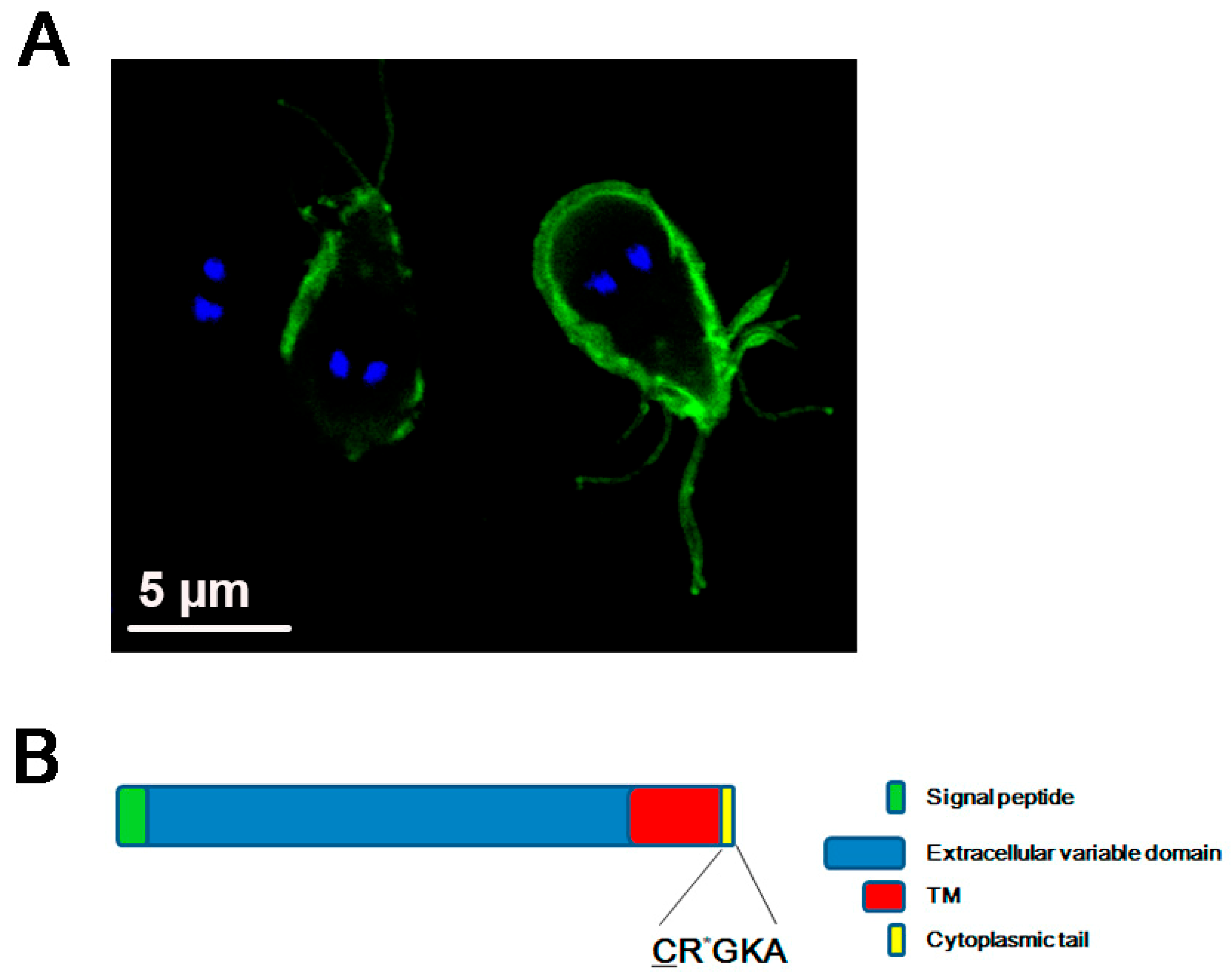
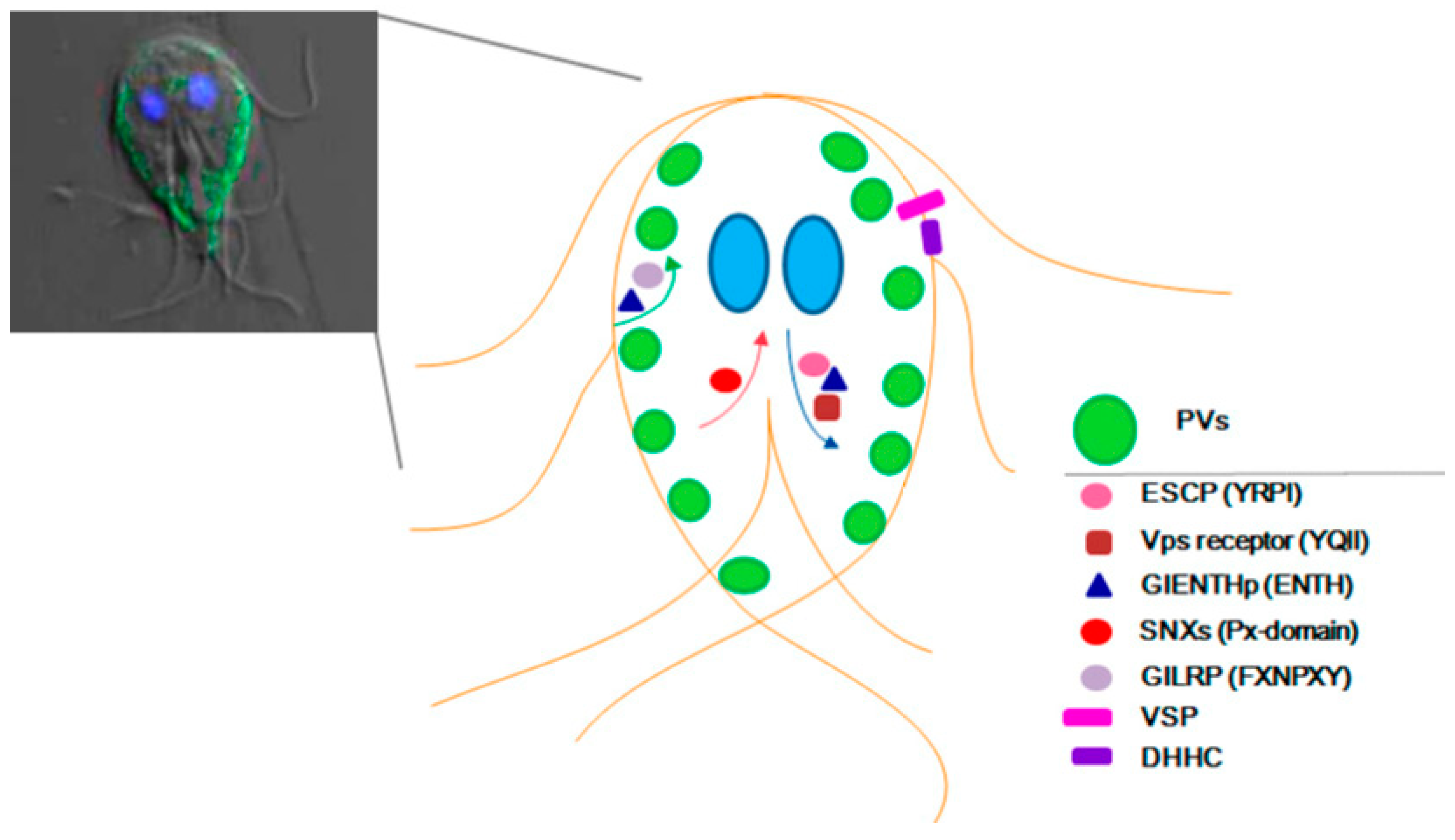

{kind=link}
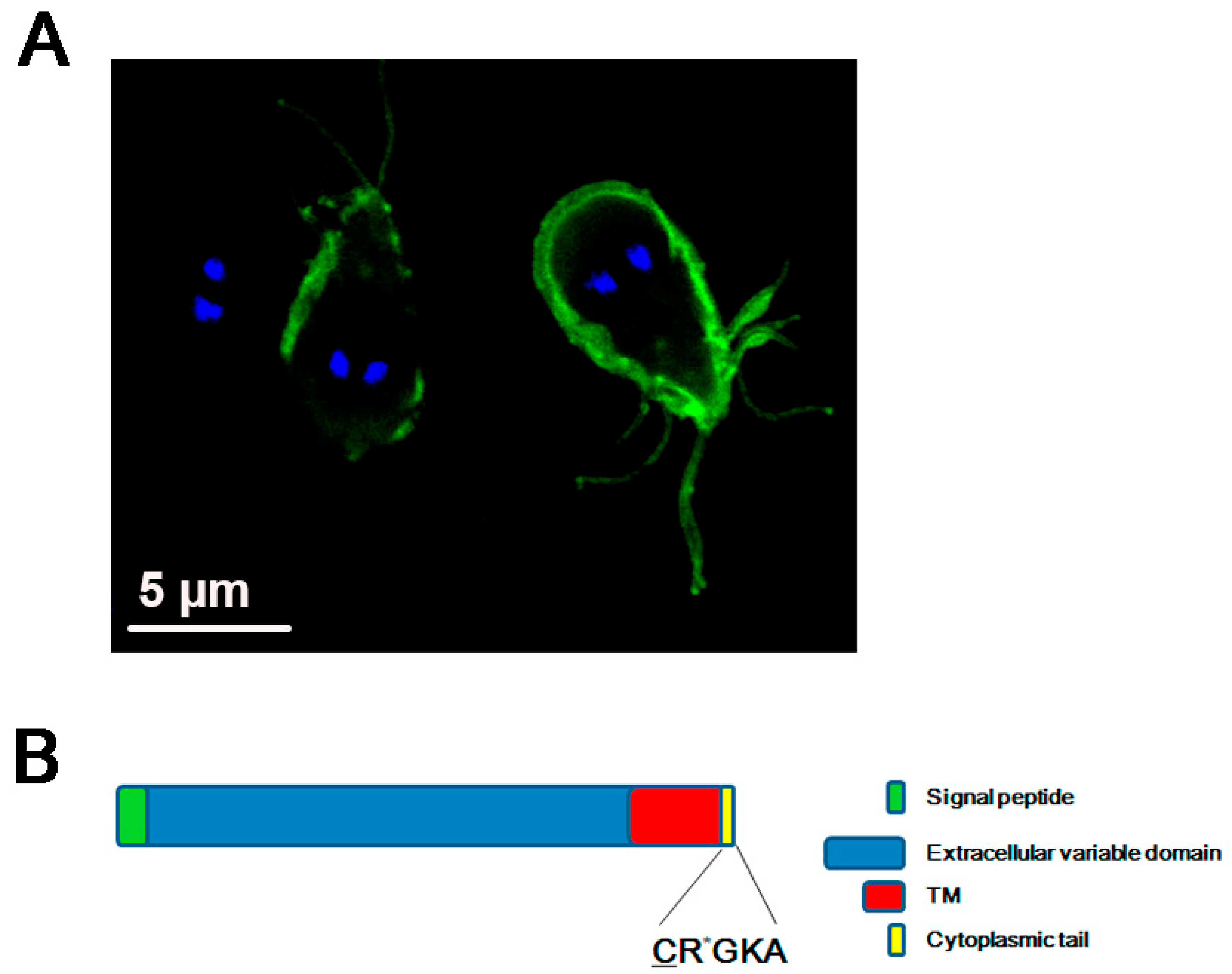{kind=link}
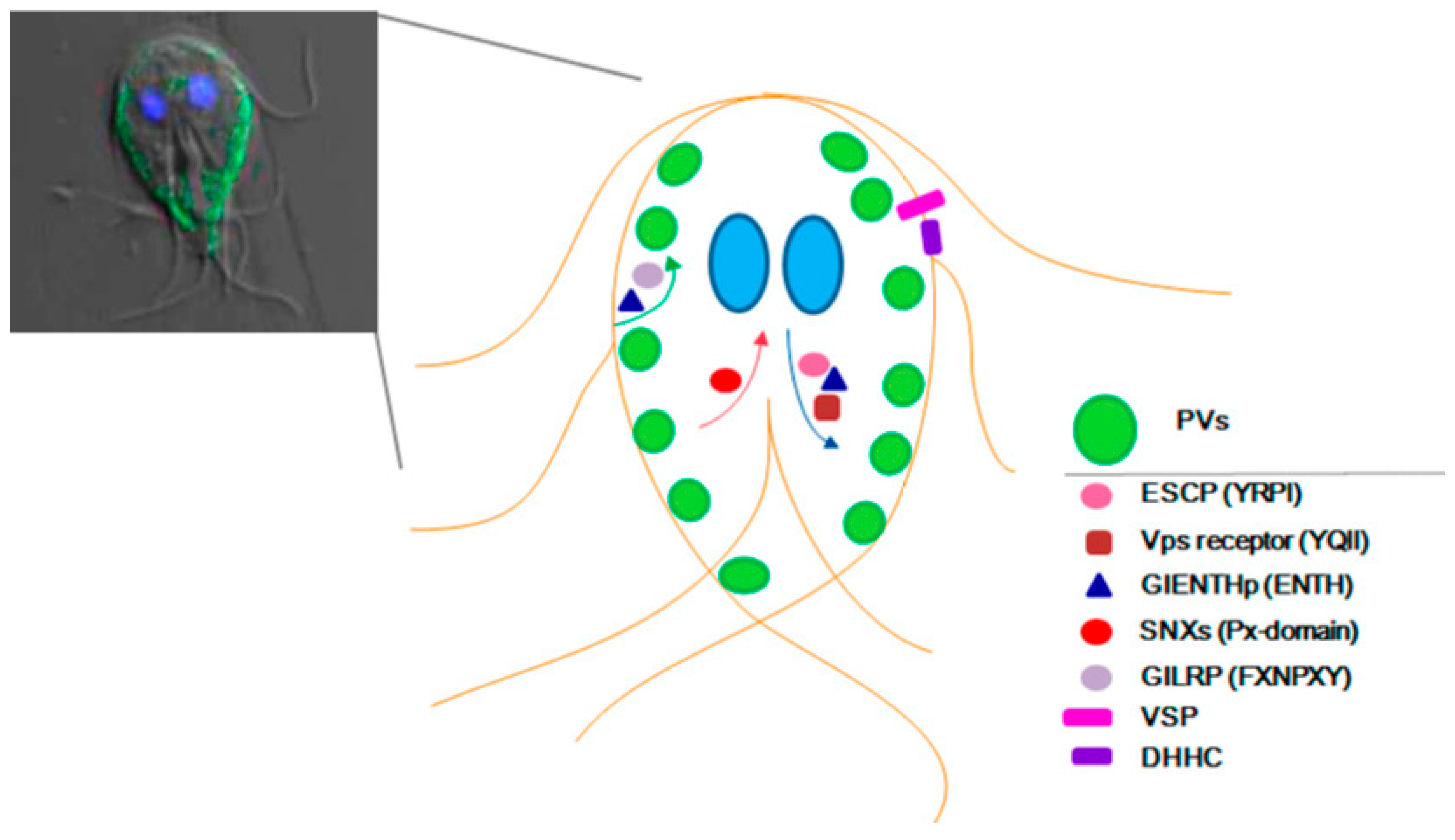{kind=link}
| GL50803_16596 | GL50803_42357 | GL50803_16548 | GL50803_24488 | GL50803_16595 | GL50803_7723 | |
|---|---|---|---|---|---|---|
| Basic amino acids | - | - | + | + | + | + |
| PX-domain PIP binding (ordered by affinity) | PI3P | PI3P PI5P PI3,4,5P3 PI3,5P2 PI4,5P2 PI4P | PI3,4P2 PI3,5P2 PI4,5P2 PI3P PI4P PI3P | PI3,4,5P3 PI3,4P2 PI3,5P2 PI4,5P2 PI3P PI4P PI3P | PI3,5P2 PI3P PI4P PI3P PI4,5P2 | ND |
| Localization in yeast | Endosomes | Endosomes | ND | ND | Cytosolic vesicles Golgi? | Vacuole membrane |
| Localization in G. lamblia | ND | ND | PVs | ND | ND | ND |
| mRNA expression | + | + (mayor expression in cysts) | + | + (mayor expression in cysts) | + | + |
| Giardia Ortholog Groups | |||||
|---|---|---|---|---|---|
| assemblage A | assemblage A2 | assemblage B | assemblage E | ||
| Motiv | Isolate WB | Isolate DH | Isolate GS_B | Isolate GS | Isolate P15 |
| FYVE | GL50803_16653 | DHA2_16653 | GSB_16653 | GL50581_4212 | GLP15_3543 |
| ENTH | GL50803_3256 | DHA2_151051 | GSB_151711 | GL50581_49 | GLP15_2242 |
| PX | GL50803_16596 | DHA2_150243 | GSB_150230 | NO | GLP15_442 |
| GL50803_42357 | DHA2_150236 | GSB_150235 | GL50581_3587 | GLP15_433 | |
| GL50803_16548 | DHA2_16548 | GSB_16548 | GL50581_1134 | GLP15_3433 | |
| GL50803_24488 | DHA2_152720 | GSB_154119 | GL50581_3300 | GLP15_1942 | |
| GL50803_7723 | DHA2_150244 | GSB_150229 | GL50581_444 | GLP15_443 | |
| GL50803_16595 | DHA2_150245 | GSB_150228 | GL50581_445 | GLP15_444 | |
| YXXØ | GL50803_14566 | DHA2_14566 | GSB_14566 | GL50581_1260 | GLP15_3781 |
| GL50803_28954 | DHA2_28954 | GSB_28954 | GL50581_1526 | GLP15_2327 | |
| FXNPXY | GL50803_113565 | DHA2_152495 | GSB_153075 | NO | GLP15_1378 |
| GL50803_94510 | DHA2_153693 | GLP15_1686 | |||
| CRGKA (VSP) | - | - | GSB_150963 * | - | - |
| GL50803_101074 * | - | - | - | - | |
| GL50803_112208 * | - | - | - | - | |
| CRRSKAV (HCNCp) | GL50803_40376 | DHA2_40376 | GSB_40376 | GL50581_1041 | GLP15_486 |
| DHHC | GL50803_42184 | DHA2_153342 | GSB_154139 | GL50581_1824 | GLP15_189 |
| GL50803_8711 | DHA2_150040 | GSB_150746 | GL50581_3650 | GLP15_196 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Touz, M.C.; Feliziani, C.; Rópolo, A.S. Membrane-Associated Proteins in Giardia lamblia. Genes 2018, 9, 404. https://doi.org/10.3390/genes9080404
Touz MC, Feliziani C, Rópolo AS. Membrane-Associated Proteins in Giardia lamblia. Genes. 2018; 9(8):404. https://doi.org/10.3390/genes9080404
Chicago/Turabian StyleTouz, María C., Constanza Feliziani, and Andrea S. Rópolo. 2018. "Membrane-Associated Proteins in Giardia lamblia" Genes 9, no. 8: 404. https://doi.org/10.3390/genes9080404
APA StyleTouz, M. C., Feliziani, C., & Rópolo, A. S. (2018). Membrane-Associated Proteins in Giardia lamblia. Genes, 9(8), 404. https://doi.org/10.3390/genes9080404