Genome Based Meta-QTL Analysis of Grain Weight in Tetraploid Wheat Identifies Rare Alleles of GRF4 Associated with Larger Grains

 ,
, 
 and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Tetraploid Wheat Mapping Populations
2.2. Growing Conditions and Experimental Design
2.3. QTL Analysis
2.4. Genome-Based Meta- Quantitative Trait Loci Analysis
2.5. Identification of Wheat Orthologs to Yield-Related Genes in Rice
2.6. GRF4-A Single Nucleotide Polymorphism Marker Development
2.7. Development and Evaluation of Introgression Line IL-21.1
2.8. Allelic Variation Studies
3. Results
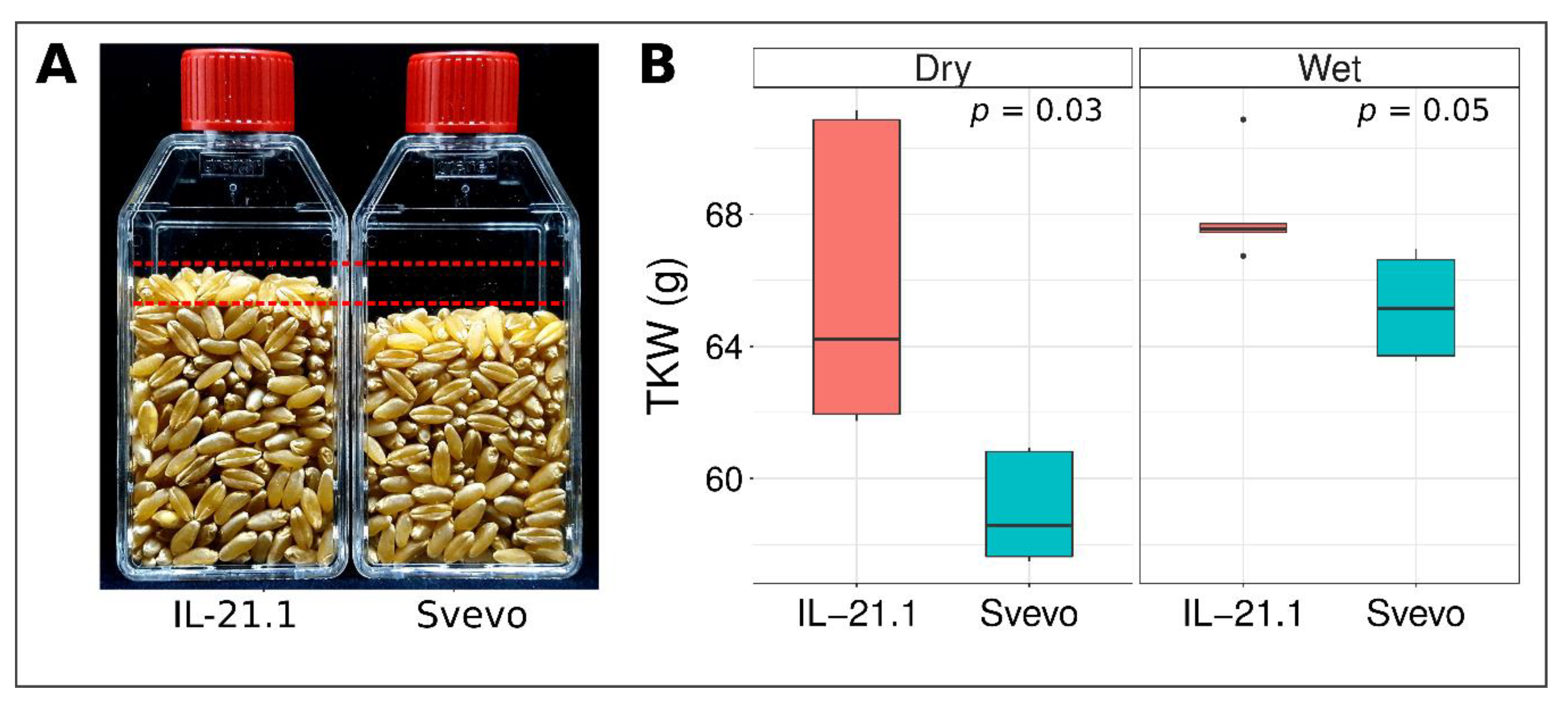
3.1. Phenotypic Characterization of Grain Parameters in Parental Lines
3.2. QTL Analysis
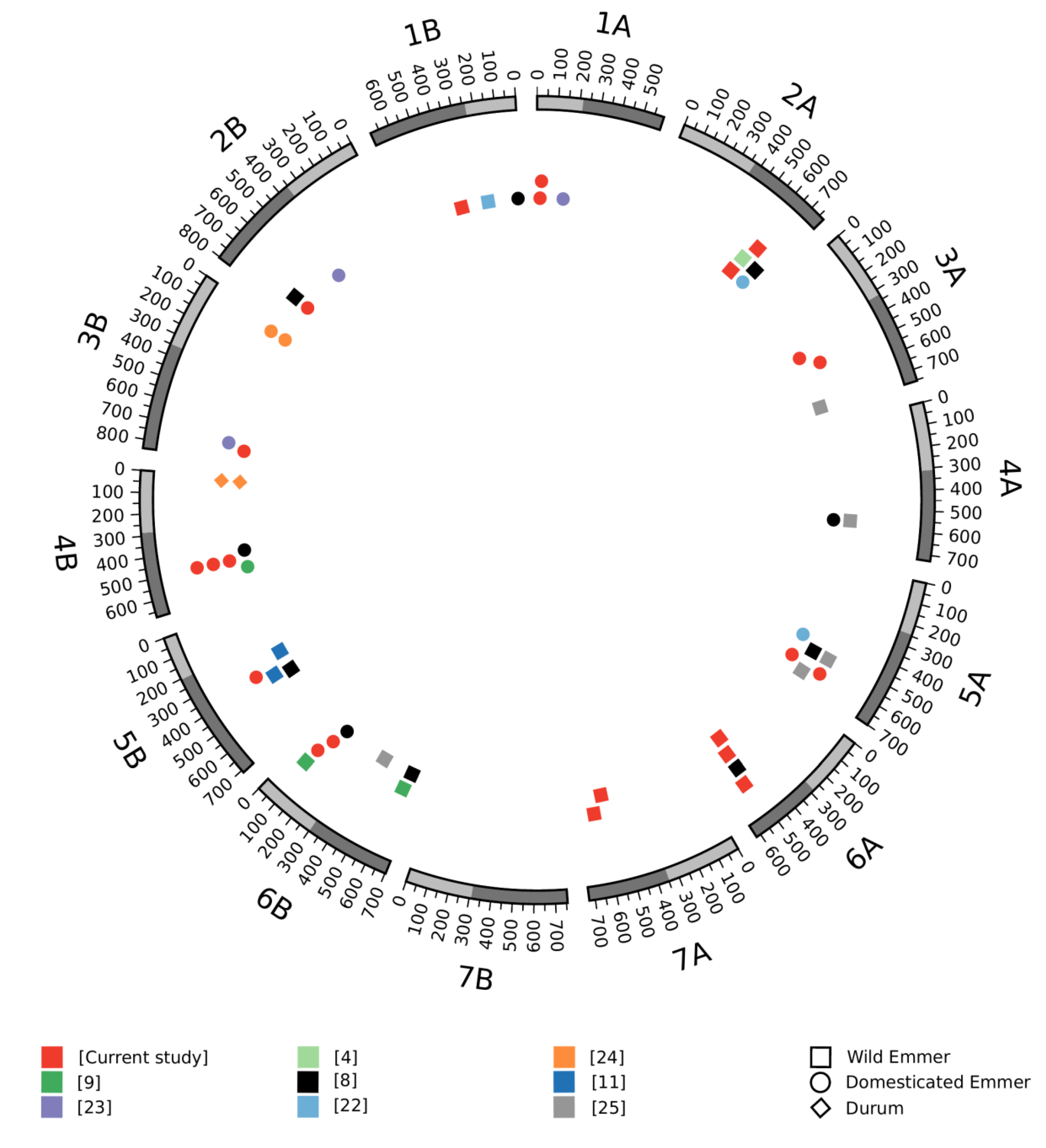
3.3. Meta-QTL Analysis
3.4. Validation of mQTL-GW-6A Using Sv × Zv Introgression Lines
3.5. Wheat Orthologs to Yield-Related Genes in Rice
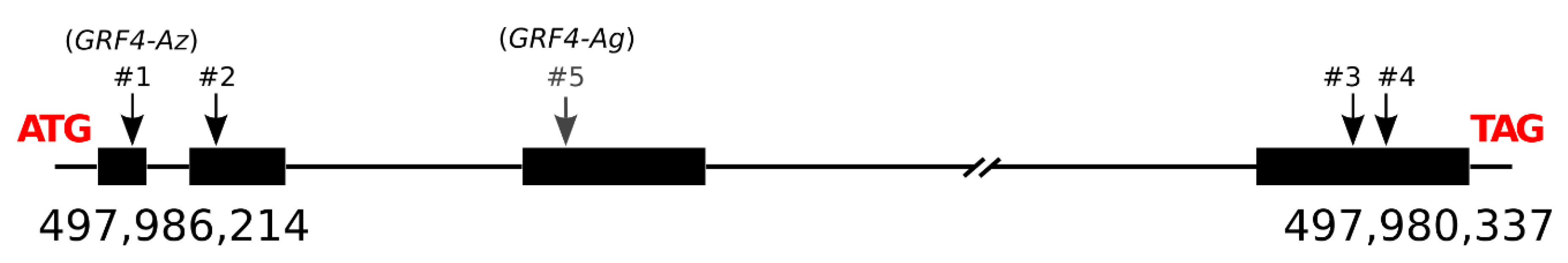
3.6. GRF4-A Polymorphisms
3.7. Allelic Diversity Study of GRF4-A
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Campbell, K.G.; Bergman, C.J.; Gualberto, D.G.; Anderson, J.A.; Giroux, M.J.; Hareland, G.; Fulcher, R.G.; Sorrells, M.E.; Finney, P.L. Quantitative trait loci associated with kernel traits in a soft x hard wheat cross. Crop Sci. 1999, 39, 1184–1195. [Google Scholar] [CrossRef]
- Gegas, V.C.; Nazari, A.; Griffiths, S.; Simmonds, J.; Fish, L.; Orford, S.; Sayers, L.; Doonan, J.H.; Snape, J.W. A genetic framework for grain size and shape variation in wheat. Plant Cell 2010, 22, 1046–1056. [Google Scholar] [CrossRef] [PubMed]
- Abbo, S.; Pinhasi van-Oss, R.; Gopher, A.; Saranga, Y.; Ofner, I.; Peleg, Z. Plant domestication versus crop evolution: A conceptual framework for cereals and grain legumes. Trends Plant Sci. 2014, 19, 351–360. [Google Scholar] [CrossRef]
- Golan, G.; Oksenberg, A.; Peleg, Z. Genetic evidence for differential selection of grain and embryo weight during wheat evolution under domestication. J. Exp. Bot. 2015, 66, 5703–5711. [Google Scholar] [CrossRef] [PubMed]
- Ozkan, H.; Brandolini, A.; Pozzi, C.; Effgen, S.; Wunder, J.; Salamini, F. A reconsideration of the domestication geography of tetraploid wheats. Theor. Appl. Genet. 2005, 110, 1052–1060. [Google Scholar] [CrossRef]
- Poyarkova, H.; Gerechter-Amitai, Z.K. Two variants of wild emmer (Triticum dicoccoides) native to Israel: Morphology and distribution. Can. J. Bot. 1991, 69, 2772–2789. [Google Scholar] [CrossRef]
- Aaronsohn, A. Agricultural and botanical explorations in Palestine. Bur. Plant Ind. Bull. USDA 1910, 180, 1–63. [Google Scholar]
- Peleg, Z.; Fahima, T.; Korol, A.B.; Abbo, S.; Saranga, Y. Genetic analysis of wheat domestication and evolution under domestication. J. Exp. Bot. 2011, 62, 5051–5061. [Google Scholar] [CrossRef]
- Elouafi, I.; Nachit, M.M. A genetic linkage map of the Durum x Triticum dicoccoides backcross population based on SSRs and AFLP markers, and QTL analysis for milling traits. Theor. Appl. Genet. 2004, 108, 401–413. [Google Scholar] [CrossRef]
- Nave, M.; Avni, R.; Ben-Zvi, B.; Hale, I.; Distelfeld, A. QTLs for uniform grain dimensions and germination selected during wheat domestication are co-located on chromosome 4B. Theor. Appl. Genet. 2016, 129, 1303–1315. [Google Scholar] [CrossRef]
- Thanh, P.T.; Vladutu, C.I.; Kianian, S.F.; Thanh, P.T.; Ishii, T.; Nitta, M.; Nasuda, S.; Mori, N. Molecular genetic analysis of domestication traits in emmer wheat. I: Map construction and QTL analysis using an F2 population. Agric. Environ. Biotechnol. 2013, 27, 3627–3637. [Google Scholar] [CrossRef]
- Hu, M.J.; Zhang, H.P.; Cao, J.J.; Zhu, X.F.; Wang, S.X.; Jiang, H.; Wu, Z.Y.; Lu, J.; Chang, C.; Sun, G.; et al. Characterization of an IAA-glucose hydrolase gene TaTGW6 associated with grain weight in common wheat (Triticum aestivum L.). Mol. Breed. 2016. [Google Scholar] [CrossRef]
- Nadolska-Orczyk, A.; Rajchel, I.K.; Orczyk, W.; Gasparis, S. Major genes determining yield-related traits in wheat and barley. Theor. Appl. Genet. 2017, 130, 1081–1098. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Jiang, L.; Zheng, J.; Wang, T.; Wang, H.; Huang, Y.; Hong, Z. Genetic bases of rice grain shape: So many genes, so little known. Trends Plant Sci. 2013, 18, 218–226. [Google Scholar] [CrossRef]
- Zheng, J.; Zhang, Y.; Wang, C. Molecular functions of genes related to grain shape in rice. Breed. Sci. 2015, 65, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Zhao, M.; Zhang, Q.; Xu, Z.; Xu, Q. The DENSE AND ERECT PANICLE 1 (DEP1) gene offering the potential in the breeding of high-yielding rice. Breed. Sci. 2016, 66, 659–667. [Google Scholar] [CrossRef]
- Wang, E.; Wang, J.; Zhu, X.; Hao, W.; Wang, L.; Li, Q.; Zhang, L.; He, W.; Lu, B.; Lin, H.; et al. Control of rice grain-filling and yield by a gene with a potential signature of domestication. Nat. Genet. 2008, 40, 1370–1374. [Google Scholar] [CrossRef]
- Duan, P.; Ni, S.; Wang, J.; Zhang, B.; Xu, R.; Wang, Y.; Chen, H.; Zhu, X.; Li, Y. Regulation of OsGRF4 by OsmiR396 controls grain size and yield in rice. Nat. Plants 2016, 2, 15203. [Google Scholar] [CrossRef]
- Che, R.; Tong, H.; Shi, B.; Liu, Y.; Fang, S.; Liu, D.; Xiao, Y.; Hu, B.; Liu, L.; Wang, H.; et al. Control of grain size and rice yield by GL2-mediated brassinosteroid responses. Nat. Plants 2015, 2, 15195. [Google Scholar] [CrossRef]
- Tsukaya, H. Yield increase: GRFs provide the key. Nat. Plants 2015, 2, 15210. [Google Scholar] [CrossRef]
- Avni, R.; Nave, M.; Eilam, T.; Sela, H.; Alekperov, C.; Peleg, Z.; Dvorak, J.; Korol, A.; Distelfeld, A. Ultra-dense genetic map of durum wheat × wild emmer wheat developed using the 90K iSelect SNP genotyping assay. Mol. Breed. 2014, 34, 1549–1562. [Google Scholar] [CrossRef]
- Peng, J.; Ronin, Y.; Fahima, T.; Röder, M.S.; Li, Y.; Nevo, E.; Korol, A. Domestication quantitative trait loci in Triticum dicoccoides, the progenitor of wheat. Proc. Natl. Acad. Sci. USA 2003, 100, 2489–2494. [Google Scholar] [CrossRef]
- Faris, J.D.; Zhang, Q.; Chao, S.; Zhang, Z.; Xu, S.S. Analysis of agronomic and domestication traits in a durum × cultivated emmer wheat population using a high-density single nucleotide polymorphism-based linkage map. Theor. Appl. Genet. 2014, 127, 2333–2348. [Google Scholar] [CrossRef]
- Russo, M.A.; Ficco, D.B.M.; Laido, G.; Marone, D.; Papa, R.; Blanco, A.; Gadaleta, A.; De Vita, P.; Mastrangelo, M.A. Dense durum wheat × T. dicoccum linkage map based on SNP markers for the study of seed morphology. Mol. Breed. 2014, 34, 1579–1597. [Google Scholar] [CrossRef]
- Tzarfati, R.; Barak, V.; Krugman, T.; Fahima, T.; Abbo, S.; Saranga, Y.; Korol, A.B. Novel quantitative trait loci underlying major domestication traits in tetraploid wheat. Mol. Breed. 2014, 34, 1613–1628. [Google Scholar] [CrossRef]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An information aesthetic for comparative genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef]
- Avni, R.; Moran Nave, M.; Barad, O.; Baruch, K.; Twardziok, S.O.; Gundlach, H.; Hale, I.; Mascher, M.; Spannagl, M.; Wiebe, K.; et al. Wild emmer genome architecture and diversity elucidate wheat evolution and domestication. Science 2017, 97, 93–97. [Google Scholar] [CrossRef]
- Wang, S.; Wong, D.; Forrest, K.; Allen, A.; Chao, S.; Huang, B.E.; Maccaferri, M.; Salvi, S.; Milner, S.G.; Cattivelli, L.; et al. Characterization of polyploid wheat genomic diversity using a high-density 90 000 single nucleotide polymorphism array. Plant Biotechnol. J. 2014, 12, 787–796. [Google Scholar] [CrossRef]
- Sun, P.; Zhang, W.; Wang, Y.; He, Q.; Shu, F.; Liu, H.; Wang, J.; Wang, J.; Yuan, L.; Deng, H. OsGRF4 controls grain shape, panicle length and seed shattering in rice. J. Integr. Plant Biol. 2016, 58, 836–847. [Google Scholar] [CrossRef]
- Arcade, A.; Labourdette, A.; Falque, M.; Mangin, B.; Chardon, F.; Charcosset, A.; Joets, J. BioMercator: Integrating genetic maps and QTL towards discovery of candidate genes. Bioinformatics 2004, 20, 2324–2326. [Google Scholar] [CrossRef]
- Goffinet, B.; Gerber, S. Quantitative trait loci: A meta-analysis. Genetics 2000, 155, 463–473. [Google Scholar]
- Maccaferri, M.; Ricci, A.; Salvi, S.; Milner, S.G.; Noli, E.; Martelli, P.L.; Casadio, R.; Akhunov, E.; Scalabrin, S.; Vendramin, V.; et al. A high-density, SNP-based consensus map of tetraploid wheat as a bridge to integrate durum and bread wheat genomics and breeding. Plant Biotechnol. J. 2015, 13, 648–663. [Google Scholar] [CrossRef]
- Mester, D.; Ronin, Y.; Schnable, P.; Aluru, S.; Korol, A. Fast and accurate construction of ultra-dense consensus genetic maps using evolution strategy optimization. PLoS ONE 2015, 10, e0122485. [Google Scholar] [CrossRef]
- Bezant, J.H.; Laurie, D.A.; Pratchett, N.; Chojecki, J.; Kearsey, M.J. Mapping of QTL controlling NIR predicted hot water extract and grain nitrogen content in a spring barley cross using marker-regression. Plant Breed. 1997, 116, 141–145. [Google Scholar] [CrossRef]
- Cooper, M.; Woodruff, D.R.; Eisemann, R.L.; Brennan, P.S.; DeLacy, I.H. A selection strategy to accommodate genotype-by-environment interaction for grain yield of wheat: Managed-environments for selection among genotypes. Theor. Appl. Genet. 1995, 90, 492–502. [Google Scholar] [CrossRef]
- Simmonds, J.; Scott, P.; Brinton, J.; Mestre, T.C.; Bush, M.; del Blanco, A.; Dubcovsky, J.; Uauy, C. A splice acceptor site mutation in TaGW2-A1 increases thousand grain weight in tetraploid and hexaploid wheat through wider and longer grains. Theor. Appl. Genet. 2016, 129, 1099–1112. [Google Scholar] [CrossRef]
- Distelfeld, A.; Uauy, C.; Olmos, S.; Schlatter, A.R.; Dubcovsky, J.; Fahima, T. Microcolinearity between a 2-cM region encompassing the grain protein content locus Gpc-6B1 on wheat chromosome 6B and a 350-kb region on rice chromosome 2. Funct. Integr. Genom. 2004, 4, 59–66. [Google Scholar] [CrossRef]
- Lu, L.; Shao, D.; Qiu, X.; Sun, L.; Yan, W.; Zhou, X.; Yang, L.; He, Y.; Yu, S.; Xing, Y. Natural variation and artificial selection in four genes determine grain shape in rice. New Phytol. 2013, 200, 1269–1280. [Google Scholar] [CrossRef]
- Ozkan, H.; Willcox, G.; Graner, A.; Salamini, F.; Kilian, B. Geographic distribution and domestication of wild emmer wheat (Triticum dicoccoides). Genet. Resour. Crop Evol. 2011, 58, 11–53. [Google Scholar] [CrossRef]
- Sela, H.; Ezrati, S.; Ben-Yehuda, P.; Manisterski, J.; Akhunov, E.; Dvorak, J.; Breiman, A.; Korol, A. Linkage disequilibrium and association analysis of stripe rust resistance in wild emmer wheat (Triticum turgidum ssp. dicoccoides) population in Israel. Theor. Appl. Genet. 2014, 127, 2453–2463. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Trait | Environment | Sv | Zv |
|---|---|---|---|
| Mean spike weight (g) | 2014R | 3.9 ± 0.1 | 2.5 ± 0.1 |
| 2015A | 4.0 ± 0.1 | 1.6 ± 0.8 | |
| TKW (g) | 2014R | 61.9 ± 0.2 | 45.8 ± 0.1 |
| 2015A | 55.7 ± 0.1 | 29.1 ± 0.1 | |
| Grain area (mm2) | 2014R | 21.6 ± 0.2 | 23.3 ± 0.3 |
| 2015A | 23.0 ± 0.3 | 18.6 ± 0.4 | |
| Grain width (mm) | 2014R | 3.2 ± 0.03 | 2.8 ± 0.0 |
| 2015A | 3.7 ± 0.03 | 2.5 ± 0.0 | |
| Grain length (mm) | 2014R | 8.1 ± 0.08 | 10.2 ± 0.1 |
| 2015A | 8.5 ± 0.05 | 10.1 ± 0.1 |
| Rice Gene | Rice Gene Function | Source | Wheat Chr. | Wheat Alignment Start | Wheat Alignment End | Wheat Gene Function | WEW Gene ID |
|---|---|---|---|---|---|---|---|
| D11/DWARF11 | Cytochrome P450 (CYP724B1) enzyme | Reviewed by [14] | 2A | 561795447 | 561798557 | Cytochrome P450 superfamily protein | TRIDC2AG048380 |
| 2B | 496938005 | 496941148 | Cytochrome P450 superfamily protein | TRIDC2BG050840 | |||
| D2 | Cytochrome P450 (CYP90D) enzyme | Reviewed by [14] | 2A | 4843464 | 5209969 | Cytochrome P450 superfamily protein | TRIDC2AG001470 |
| 2B | 5686865 | 5987817 | Cytochrome P450 superfamily protein | TRIDC2BG001370 | |||
| D61 | BR insensitive (BRI)-like leucine-rich repeat (LRR) receptor kinase | Reviewed by [14] | 3A | 465976238 | 465979780 | Leucine-rich receptor-like protein kinase family protein | TRIDC3AG036670 |
| 3B | 453931439 | 453935096 | receptor-like protein kinase 2 | TRIDC3BG041310 | |||
| GIF1 | Cell wall invertase | Reviewed by [14] | 2A | 503854205 | 503855081 | Beta-fructofuranosidase, insoluble isoenzyme 2 (Cell wall invertase 2) | TRIDC2AG042730 |
| 2B | 447195335 | 447196211 | Beta-fructofuranosidase, insoluble isoenzyme 2 (Cell wall invertase 2) | TRIDC2BG045820 | |||
| GRF4/GS2 | Growth-Regulating Factor 4 (OsGRF4) | [18,29] | 2A | 680343644 | 680346735 | Growth-regulating factor 3 | TRIDC2AG062550 |
| 2B | 649416512 | 649417723 | Growth-regulating factor 3 | TRIDC2BG066890 | |||
| 6A | 497985063 | 497985958 | Growth-regulating factor 4 | TRIDC6AG041360 | |||
| 6B | 517412993 | 517416246 | Growth-regulating factor 4 | TRIDC6BG048340 | |||
| GS3 | Membrane protein with multiple domains | Reviewed by [14] | 4A | 714924235 | 714925670 | Grain length protein | TRIDC4AG069340 |
| 7A | 5283743 | 5283958 | Grain length protein | TRIDC7AG001510 | |||
| GS5 | Serine carboxypeptidase | Reviewed by [14] | 3A | 182355936 | 182359086 | serine carboxypeptidase-like 33 | TRIDC3AG023140 |
| 3B | 212372375 | 212373474 | Carboxypeptidase Y homolog A | TRIDC3BG026960 | |||
| GW2 | RING-type E3 ubiquitin ligase | Reviewed by [14] | 6A | 230789449 | 230809149 | Protein SIP5 (*TaGW2) | TRIDC6AG027660 |
| 6B | 294434000 | 294448424 | Protein SIP5 | TRIDC6BG033820 | |||
| GW5 | Arginine-rich protein of 144 amino acids | Reviewed by [14] | 1A | 142379896 | 142381359 | IQ-domain 26 | TRIDC1AG017640 |
| 1B | 185320338 | 185321816 | IQ-domain 26 | TRIDC1BG021520 | |||
| 3A | 69160021 | 69161092 | IQ-domain 26 | TRIDC3AG013280 | |||
| 3B | 111226601 | 111227636 | IQ-domain 26 | TRIDC3BG017740 | |||
| GW8/SPL16 | SQUAMOSA promoter-binding protein-like 16 | Reviewed by [14] | 7A | 251030195 | 251034936 | undescribed protein | TRIDC7AG033770 |
| 7B | 230000953 | 230005263 | Squamosa promoter-binding-like protein 16 | TRIDC7BG025060 | |||
| qGL3 | Ser/Thr phosphatase of the protein phosphatase kelch-like (PPKL) family | Reviewed by [15] | 5A | 683802818 | 683803388 | Bifunctional inhibitor/lipid-transfer protein/seed storage 2S albumin superfamily protein | TRIDC5AG075900 |
| SRS3 | Kinesin 13 protein | Reviewed by [14] | 1A | 131830406 | 131835083 | Kinesin-related protein 6 | TRIDC1AG016970 |
| 1B | 142745967 | 142751957 | Kinesin-related protein 6 | TRIDC1BG017960 | |||
| 3A | 274848912 | 274857469 | Kinesin-related protein 6 | TRIDC3AG027550 | |||
| 3B | 295258442 | 295261011 | Kinesin-related protein 6 | TRIDC3BG032430 | |||
| DEP1 | G protein γ subunit | [16] | 5A | 422466437 | 422469555 | Guanine nucleotide-binding protein subunit gamma 3 | TRIDC5AG033880 |
| 5B | 391766206 | 391769237 | undescribed protein | TRIDC5BG035790 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Avni, R.; Oren, L.; Shabtay, G.; Assili, S.; Pozniak, C.; Hale, I.; Ben-David, R.; Peleg, Z.; Distelfeld, A. Genome Based Meta-QTL Analysis of Grain Weight in Tetraploid Wheat Identifies Rare Alleles of GRF4 Associated with Larger Grains. Genes 2018, 9, 636. https://doi.org/10.3390/genes9120636
Avni R, Oren L, Shabtay G, Assili S, Pozniak C, Hale I, Ben-David R, Peleg Z, Distelfeld A. Genome Based Meta-QTL Analysis of Grain Weight in Tetraploid Wheat Identifies Rare Alleles of GRF4 Associated with Larger Grains. Genes. 2018; 9(12):636. https://doi.org/10.3390/genes9120636
Chicago/Turabian StyleAvni, Raz, Leah Oren, Gai Shabtay, Siwar Assili, Curtis Pozniak, Iago Hale, Roi Ben-David, Zvi Peleg, and Assaf Distelfeld. 2018. "Genome Based Meta-QTL Analysis of Grain Weight in Tetraploid Wheat Identifies Rare Alleles of GRF4 Associated with Larger Grains" Genes 9, no. 12: 636. https://doi.org/10.3390/genes9120636
APA StyleAvni, R., Oren, L., Shabtay, G., Assili, S., Pozniak, C., Hale, I., Ben-David, R., Peleg, Z., & Distelfeld, A. (2018). Genome Based Meta-QTL Analysis of Grain Weight in Tetraploid Wheat Identifies Rare Alleles of GRF4 Associated with Larger Grains. Genes, 9(12), 636. https://doi.org/10.3390/genes9120636